
Abstract
Kinorhynchs are marine, microscopic ecdysozoan animals that are found throughout the world’s ocean. Cephalorhyncha flosculosa sp. nov. is described from the Aegean Coast of Turkey. Samples were collected from intertidal zones at two localities. The new species is distinguished from its congeners by having flosculi in midventral positions on segment 3–8, and by differences in its general spine and sensory spot positions. Until now, species of Cephalorhyncha were only known from the Pacific Ocean, hence, this record of the genus at the Aegean Sea not only expands its geographic distribution to Turkey, but is likely to expand it throughout the Mediterranean Sea and much of southern Europe. The new species of Cephalorhyncha represents the fifth kinorhynch species recorded from Turkey, and increases also the number of known Cephalorhyncha species to four.
Similar content being viewed by others


Background
The phylum Kinorhyncha is classified within the invertebrate animals. They are microscopic marine worms generally not longer than 1 mm. Kinorhynchs live throughout the world’s ocean, from intertidal areas to 8000 m in depth. They are most common in subtidal muddy sediments, but some have been found in algal holdfasts, sponges, or other invertebrates. [1–3]. The phylum currently comprises about 239 species distributed across 29 genera [4–11]. Cephalorhyncha is a small genus accommodating only three species. The genus (type species Cephalorhyncha asiatica [12] was proposed by Adrianov [13]. Later, Neuhaus and Blasche [14] reassigned the Californian species Echinoderes nybakkeni Higgins, 1986 [15] to Cephalorhyncha. The third species, C. liticola Sørensen, 2008 [16] was described from Hawaii by Sørensen [16]. Species of Cephalorhyncha are characterized by possessing a sternal plate of segment 2 that is only incompletely differentiated into one tergal and two sternal plates, i.e., fissures of the tergosternal junctions are fully developed whereas the midsternal junction is incomplete. Segments 3–10 consist of one tergal and two sternal plates [3, 13, 14].
Effective management and conservation of biodiversity and ecosystems depends largely on the taxonomic determination of species composition. Unfortunately, insufficient taxonomic information intercepts our ability to make informed decisions about conservation and sustainable management of biodiversity and ecosystems. Turkey has numerous sandy beaches distributed along its coastline, but the marine meiofauna has been poorly explored despite the fact that the country has a vast coastline of about 8300 km. From a taxonomic perspective, mostly the Copepoda have been studied to a large extent [17, 18]. Oppositely, the marine and meiobenthic Kinorhyncha have been very poorly explored. Băcescu [19] reported the first kinorhynch species, Pycnophyes communis Zelinka, 1908, from the Black Sea. Later, Sönmez et al. [20] reported Echinoderes gerardi Higgins, 1978 and a species resembling Echinoderes bispinosus Higgins, 1982 from the Aegean coast of Turkey, and Ürkmez et al. [21] found Echinoderes dujardinii Claparède, 1863 from subtidal sediments of Sinop Bay, Black Sea. In the present paper, a new species of Cephalorhyncha collected at two intertidal localities from the Aegean Coast of Turkey is described.
Methods
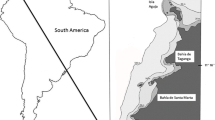
Samples were collected from two intertidal localities (Fig. 1): the north coast of Kara Island, ca. 5 km southeast of Bodrum on the Turkish mainland (36°59′08″N, 27°27′13″E), and at Çalış on the beach of Fethiye (36°39′33″N, 29°06′35″E) using the Karaman–Chappius method [22]. This method consists of a funnel-shaped excavation into the sediment down to the groundwater level, allowing sea water to fill the bottom of the pit, and then collecting that water with a suitable cup and filtering it through plankton net. The filtered samples were fixed in 70% alcohol and kinorhynch specimens picked up using an Olympus SZ-16 stereomicroscope. Specimens for light microscopic examination were dehydrated through a graded glycerin series, and mounted in Fluoromount-G between two cover slips, and attached to a plastic H–S slide mount which enables from viewing from both sides. The specimens were examined with an Olympus BX51 compound microscope equipped with DIC optics. Photos were taken with an Olympus DP27 camera, and line art illustrations were made in Adobe Illustrator CS6. Specimens for scanning electron microscopy were dehydrated through a graded alcohol-acetone series, and were subsequently critical point dried, mounted on aluminium stubs, sputter coated with a platinum–palladium mix, and examined with a JEOL JSM-6335F Field emission scanning electron microscope.

The sampling localities of C. flosculosa sp. nov
Terminology follows Sørensen and Pardos [3] and Neuhaus [2]. Specimens used for morphological comparison include type material of Cephalorhyncha liticola (Natural History Museum of Denmark, Cat. Nos. KIN-205 to KIN-207) and topotype specimens of C. asiatica, kept in the personal collection of MVS.
Cephalorhyncha flosculosa sp. nov
Material examined
Holotype, adult female, collected on May 16, 2011 from intertidal sand at Çalış (Fig. 1), Aegean Coast of Turkey (36°39′33″N, 29°06′35″E) by Dr. Serdar Sönmez and Dr. Serdar Sak, mounted in Fluoromount-G and deposited at the Natural History Museum of Denmark under catalogue numbers ZMUC KIN-1005. Paratypes include one female and two males collected on the same date and locality as the holotype, mounted in Fluoromount-G and deposited at the Natural History Museum of Denmark under catalogue numbers ZMUC KIN-1006 to KIN-1008. Additional material from same date and locality as holotype includes two specimens mounted in Fluoromount-G and stored in the personal reference collection of the first author, and three specimens, mounted for SEM, and stored in the personal reference collection of the second author. In addition, three specimens were collected on June 29, 2011 from intertidal sand at Kara Island (Fig. 1), 36°59′08″N, 27°27′13″E by Nuran Özlem Yıldız, mounted for SEM, and stored in the personal reference collection of the second author.
Diagnosis
Segment 1 forming complete ring; segment 2 with one tergal and one sternal plate, with complete tergosternal junctions, but only partially developed midsternal junction; segments 3–11 with one tergal and two sternal plates. Middorsal spines present on segments 4–8; ventrolateral tubes present on segments 2, lateroventral tubes on segment 5, lateroventral spines on segments 6–9, lateral accessory tubes on segment 8, and midlateral tubes on segment 10. Flosculum-shaped sensory spots present at posterior segment margins in midventral positions of segments 3–8.
Etymology
The species is named flosculosa (L.), meaning “having or bearing little flowers”. The name was inspired by the flower-shaped sensory spots in the midventral positions of segments 3–8.
Description
Body consists of head, neck and trunk with 11 segments (Figs. 2a, b, 3a, 4a, b). Segment and spine lengths are summarized in Table 1. For spines, sensory spots and glandular cell outlets positions see Table 2.

Line art illustrations of C. flosculosa sp. nov. a Female, dorsal view; b female, ventral view; c male, segments 10–11, dorsal view; d male, segments 10–11, ventral view. fl Flosculum, gcoI glandular cell outlet type I, lat lateral accessory tube, ld laterodorsal sensory spot, ltas lateral terminal accessory spine, lvs lateroventral spine, lvt lateroventral tube, mds middorsal spine, mls midlateral spine, pd paradorsal sensory spot, pmf partially developed midventral fissure, ps penile spines, sd subdorsal sensory spot, vlt ventrolateral tube

Light micrographs of C. flosculosa sp. nov. a Holotypic female KIN-1005, b paratypic female KIN-1006, c–g paratypic male KIN-1007, h paratypic male KIN-1008. a Lateral overview; b head and segments 1 and 2 in an un-squeezed specimen, ventral view; c neck and segments 1–5, dorsal view; d neck and segments 1–4, ventral view; e segments 4–8, dorsal view; f segments 3–6, ventral view; g segments 8–10, dorsal view; h segments 8–11, lateral view. gcoI Glandular cell outlet type I, lvs8 lateroventral spine on segment 8, mds middorsal spine, mls midlateral spine on segment 10, mvp midventral placid, pf pectinate fringe, pmf partially developed midventral fissure, ps penile spine, vm ventromedial sensory spot

Scanning electron micrographs of female C. flosculosa sp. nov. a Ventral view, b laterodorsal view. mc Mouth cone, s01 segment 1, s05–06 segment 5 and 6
Mouth cone with nine outer oral styles; outer oral styles alternate in length between longer and shorter ones (five long and four short), but all of them consist of two joined units (Figs. 4a, 7e). Exact arrangement of scalids could not be examined in any of the mounted specimens, since the introverts were either partly retracted or too dirty. Neck with 16 placids with conspicuously rounded anterolateral corners (Figs. 3c, d); midventral placids broader than remaining ones (Figs. 2b, 3b, d). The trunk is laterally compressed, which makes the specimens appear broad in lateral perspective, but rather narrow in dorsal or ventral view. This lateral compression also makes mounting for LM in a dorso-ventral orientation quite challenging.
Segment 1 consists of a closed ring (Figs. 2a, b, 3c, d, 4a). Glandular cell outlet type 1 present in middorsal (Figs. 2a, 6a) and lateroventral positions near anterior margin of segment (Figs. 2b, 5a). Sensory spots present in subdorsal and laterodorsal positions (Figs. 2a, 6a, 7a); sensory spots large, with a central pore, numerous short micropapillae, and a few considerably longer ones along the posterior margin. Cuticular hairs lightly scattered on the dorsal and lateral sides (Fig. 6a); ventral side completely devoid of hairs. Posterior segment margin terminates into a well-defined pectinate fringe with short fringe tips.

Scanning electron micrographs of C. flosculosa sp. nov. a Segments 1–2, lateroventral view; b segments 3–4, ventral view; c segment 9, lateroventral view; d segment 5–6, ventral view; e segment 10, lateroventral view; f segment 7–8, ventral view. aft Acuminate fringe tip, fl flosculum, gcoI glandular cell outlet type I, lvt5 lateroventral tubes on segment 5, lvs 6–9 lateroventral spine on segment 6–9, pf pectinate fringe, vl ventrolateral sensory spot, vlt2 ventrolateral tubes on segment 2, vm ventromedial sensory spot

Scanning electron micrographs of C. flosculosa sp. nov. a Segments 1–2, dorsal view; b segments 3–4, dorsal view; c segment 6, dorsal view; d segments, 7–8, dorsal view; e segments 9–10, dorsal view; f segment 11, dorsal view. ch Cuticular hair, gcoI glandular cell outlet type I, mds 4–8 middorsal spine on segment 4–8, mls10 midlateral spine on segment 10, pd paradorsal sensory spot, sd subdorsal sensory spot, lts lateral terminal spine

Scanning electron micrographs of C. flosculosa sp. nov. a Segment 1–2, lateral view; b laterodorsal–lateroventral parts of segments 5–6; c laterodorsal–lateroventral parts of segments 7–8 lateral view; d segments 9–10, lateral view; e mouth cone with outer oral styles; f midventral flosculum on segment 6. fl 3–8 Flosculum on segment 3–8, ld laterodorsal sensory spot, lvs 6–9 lateroventral acicular spines on segment 6–9, lvat 8 lateroventral accessory tubes on segment 8, lvt 5 lateroventral tubes on segment 5, mls10 midlateral spine on segment 10, pd paradorsal sensory spot, sd subdorsal sensory spot
Segment 2 consists of one tergal and one sternal plate. Sternal plate partially differentiated into two plates by an epicuticular midsternal junction that, opposed to the midsternal junctions on the following segments, does not divide the ventral side into two articulating sternal plates (Figs. 2b, 3b, d). Tergosternal lines well-developed. Long tubes (23 µm from SEM) present in ventrolateral positions (Figs. 2b, 5a). Sensory spots present in middorsal, laterodorsal (Figs. 2a, 6a, 7a) and ventromedial (Figs. 2b, 5a) positions; sensory spots more droplet-shaped than those on the preceding segment, and without the extended marginal hairs. Cuticular hairs lightly scattered on the dorsal and lateral sides; ventral side with very few hairs only, near the tergosternal junctions; ventromedial and paraventral areas devoid of hairs but with longitudinal wrinkles in the cuticle. Posterior segment margin extend into midventral tip; pectinate fringe well-developed, with conspicuously long and flexible fringe tips, especially in sublateral–ventromedial positions; a special lanceolate or acuminate fringe tip is located at each tergosternal junction (Fig. 5a, b, d).
Segment 3 and following eight segments consisting of one tergal and two sternal plates. Segment with regular sensory spots in subdorsal positions (Figs. 2a, 6b), and glandular cell outlets type 1 in paraventral positions (Figs. 2b, 3f). An unpaired flosculum-shaped sensory spot furthermore present midventrally (Figs. 2b, 5b, d, f); the flosculum is small and cone-shaped, located between the posteroventral corners of the two sternal plates, at the very posterior margin of the soft cuticle that forms the articulation between the plates (Fig. 7f). Cuticular hairs lightly scattered on the dorsal and lateral sides; sternal plates with 3–4 lines of cuticular hairs, anteriorly on segment, in the ventrolateral and ventromedial areas; paraventral areas without hairs, but with hair-like cuticular extensions, not emerging through perforation sites. Dorsal, lateral and ventral parts of posterior segment margins with conspicuously long and flexible fringe tips, being even longer than on segment 2 (Fig. 3f); acuminate fringe tips present at each tergosternal junction (Fig. 5b).
Segment 4 with flexible acicular spine in middorsal position (Figs. 2a, 3e, 6b). Regular sensory spots present in paradorsal positions, flanking the middorsal spine (Fig. 2a). Glandular cell outlets type 1 present in paraventral positions (Figs. 2b, 3f). Flosculum-shaped sensory spot present in midventral position as on preceding segment (Figs. 2b, 5b). Cuticular hairs as on preceding segment. Fringe tips of pectinate fringe on posterior segment margin even longer than those on preceding segment (Figs. 3e, 5b); acuminate fringe tips present at each tergosternal junction (Fig. 5b).
Segment 5 with flexible acicular spine in middorsal position (Figs. 2a, 3e, 4b), flanked by paradorsal sensory spots (Fig. 2a), and long tubes (22 µm from SEM) in lateroventral positions (Figs. 2b, 5d, 7b). Glandular cell outlets type 1 present in paraventral positions (Figs. 2b, 3f, 5d). Cuticular hairs, posterior segment margin and flosculum-shaped sensory spot as on preceding segment (Figs. 2b, 5d). Acuminate fringe tips present at each tergosternal junction (Fig. 5d).
Segment 6 with middorsal and lateroventral acicular spines (Figs. 2a, b, 3e, 4b, 5d, 6c, 7b). Regular sensory spots present in paradorsal (Figs. 2a, 6c), laterodorsal and ventromedial positions (Figs. 2a, b), and glandular cell outlets type 1 in paraventral positions (Figs. 2b, 3f, 5d, 7b). Cuticular hairs, posterior segment margin and flosculum-shaped sensory spot as on preceding segment.
Segment 7 with middorsal (Figs. 2a, 3e, 4d) and lateroventral spines (Figs. 2b, 5f). Regular sensory spots present in paradorsal (Figs. 2a, 6d) and ventromedial positions (Figs. 2a, 5f), and glandular cell outlets type 1 in paraventral positions (Figs. 2b, 5f). Cuticular hairs, posterior segment margin and flosculum-shaped sensory spot (Figs. 2b, 5f) as on preceding segment.
Segment 8 with middorsal (Figs. 2a, 3e, g, 6d) and lateroventral acicular spines, and lateral accessory tubes (Figs. 2b, 5f, 7c). Regular sensory spots present in paradorsal positions (Figs. 2a, 6d), and glandular cell outlets type 1 in paraventral positions (Figs. 2b, 5f). Small sieve plate present in lateral accessory position, partly covered by fringes from preceding segment. Cuticular hairs, posterior segment margin and flosculum-shaped sensory spot (Figs. 2b, 5f) as on preceding segment.
Segment 9 with lateroventral acicular spines (Figs. 2b, 5c, 7d). Regular sensory spots present in paradorsal, subdorsal (Figs. 2a, 6e, 7d), laterodorsal (Figs. 2a, 7d) and ventrolateral positions (Figs. 2b, 5c), and glandular cell outlets type 1 in ventromedial positions (Figs. 2b, 5c). Flosculum-shaped sensory spot not present. Cuticular hairs and posterior segment margin as on preceding segment (Fig. 3g).
Segment 10 with strong and rigid midlateral acicular spine (Figs. 2b, c, d, 3h, 6e, 7d). Regular sensory spots present in subdorsal positions (Figs. 2a, c, 6e, 7d), and glandular cell outlets type 1 in ventromedial positions (Figs. 2b, d, 5e). Cuticular hairs on dorsal and lateral sides as on preceding segments; ventral side only with few, marginal hair-like cuticular extensions. Pectinate fringe of posterior segment margin with rather short fringe tips, differing considerably from those on preceding segments (Fig. 3h), and acuminate fringe tips at tergosternal junctions absent.
Segment 11 with long lateral terminal spines (Fig. 6f); females in addition with lateral terminal accessory spines (Fig. 2a), males with three pairs of penile spines (Figs. 2c, d, 3h). Sensory spots present in subdorsal (Figs. 2a, c, 6e) and ventromedial positions near the posterior margin of the segment (Figs. 2b, d, 5e), and glandular cell outlets type 1 present in subdorsal (Figs. 2a, 6f), ventrolateral and ventromedial (Fig. 2d). Tergal plate covered by short cuticular hair-like structures, and extending into short pointed tergal extensions; sternal plates without cuticular hairs, but with short, densely set hair-like cuticular extensions near margins of rounded sternal extensions.
Discussion
Differential diagnosis
The composition of segment 2, consisting of one tergal and one sternal plate, with the sternal plate being partially divided by an incomplete midsternal junction, clearly places the new species in the echinoderid genus Cephalorhyncha [13, 14, 25]. Currently, the genus accommodates three species: C. asiatica (Adrianov, 1989), C. nybakkeni (Higgins, 1986), and C. liticola Sørensen, 2008. Cephalorhyncha flosculosa sp. nov. can be differentiated from its three congeners by its midventral flosculi on segments 3–8.
Cephalorhyncha flosculosa sp. nov. is most easily distinguished from C. liticola by its midventral flosculi, and other minor differences in distributions of sensory spots. For example, C. flosculosa sp. nov. has paradorsal sensory spots on segment 9 and no paradorsal sensory spots on segment 10, whereas C. liticola shows the opposite, with paradorsal sensory spots on segment 10 only. C. flosculosa sp. nov. also has subdorsal sensory spots on segments 9 and 10; these are missing in C. liticola.
Cephalorhyncha flosculosa sp. nov. clearly shows the closest resemblance with C. liticola. Especially the strongly laterally compressed trunk and the exceptionally long fringe tips from the posterior segment margins are highly characteristic for both species (compare Figs. 1, 3 in the present contribution with Figures 10 and 11 in Sørensen [16]). They also have their minute sizes in common. With trunk lengths of 139–209 µm in C. flosculosa sp. and 127–210 µm in C. liticola, they are indeed the two smallest kinorhynch species.
The new species is distinguished from C. nybakkeni by the presence of lateral accessory tubes on segment 8. Such tubes are absent in C. nybakkeni (Higgins, 1986). C. asiatica (Adrianov, 1989) is distinguished from C. flosculosa sp. nov. by the presence of strong midlateral spines on segment 10; C. asiatica has short, laterodorsal tubes on this segment [12, 13].
Unpublished SEM images of C. asiatica show that this species also tends to be laterally compressed, but to a lesser degree than C. flosculosa sp. nov. and C. liticola, and that C. asiatica also shows conspicuously long fringe tips from the posterior segment margins, but only from the sternal plates. SEM images of C. nybakkeni are not available, but LM images shown by Neuhaus and Blasche [14; Figure 11f–m] suggest that long fringe tips are present on the sternal plates in this species as well. Although it may seem as a rather vague character, the conspicuously long fringe tips, at least from the sternal plates, could potentially represent an additional diagnostic trait for the genus.
Flosculi
Van der Land [26] described flosculi from the priapulid Tubiluchus corallicola as “small organs in the shape of a flower”. Since then, various types and shapes of flosculi have been reported from species of Priapulida and Loricifera [27–30]. In kinorhynchs, Adrianov [12] considers the mushroom-shaped sensory structures (type 3 sensory spots sensu Nebelsick [31] to be a kind of flosculi. These flosculi are typically found on the most posterior segments, and appear to be more numerous in juvenile specimens [2, 32, 33].
The flosculi which is reported from C. flosculosa sp. nov. differ from the typical type 3 sensory spots. Whereas type 3 sensory spots are mushroom-shaped, with a relatively long stalk equipped with short, terminal micropapillae, the flosculi in C. flosculosa sp. nov. have a shorter stalk, and the terminal micropapillae are longer and pointier. Nebelsick [31] already stressed that the homology of flosculi across the scalidophoran groups still remained to be examined, and likewise, we find it premature to suggest that the flosculi of C. flosculosa sp. nov. are homologous with type 3 sensory spots found on posterior segments of allomalorhagid and kentrorhagid kinorhynchs. What is interesting though, is that the flosculi of C. flosculosa sp. nov. are rather unusual structures, in regard to their shape as well as their position. This kind of flosculi has to our knowledge not been shown from any kinorhynch before, which in itself makes them unusual, and also their position is completely unique. Whereas all kinorhynchs typically have cuticular structures distributed around their trunk segments—from middorsal to paraventral position—it is not very common to possess unpaired, midventral structures, and we are not aware of any corresponding examples of midventral structures on the trunk segments.
Habitat preferences and zoogeography
Cephalorhyncha flosculosa sp. nov. was collected from the intertidal zone and from a sandy beach with gravel at 40–70 cm sediment depth. This special habitat preference is shared with two of its congeners: C. nybakkeni was described from the intertidal zone of a Californian beach [15] and C. liticola is intertidal as well, from a Hawaiian beach [16]. In all three cases, specimens could be found in the sand down to 70 cm depth. This suggests that at least some species of Cephalorhyncha share a certain preference for this kind of habitats. However, C. asiatica is described from subtidal mud [13], hence the habitat choice does not characterize the genus as such.
Zoogeographically, the finding of C. flosculosa sp. nov. in the Aegean Sea also attracts special attention, as it represents the first recording of the genus outside the Pacific Ocean. The finding of a representative of Cephalorhyncha in the Mediterranean Sea, basically as far away from the Pacific Ocean as possible, suggests that species of the genus could be found in any other World ocean as well. This confirms a trend that appears to be general for many kinorhynch genera, namely that their species may have a rather patchy distribution, but still on the generic level show an almost global distribution. Similar examples are the genus Dracoderes, which since its discovery appeared to be an East Asian genus [13, 34], but has more recently been found in samples from Spain [35]; Paracentrophyes was originally described from the Atlantic Ocean [36, 37], but later was recorded from the West Pacific also [38]; and Fissuroderes that was known from the Pacific only [14], until it showed up in the West Atlantic [39]. Other kinorhynch genera are still known from only single localities or restricted areas, but we find it very likely that they over time will be recorded from other regions of the world as well, and that most, if not all, kinorhynch genera will turn out to have a nearly cosmopolitan distribution. The number of taxonomists working on obscure taxa such as kinorhynchs has always been rather low which explains our limited knowledge about them.
References
Higgins RP. Kinorhyncha. In: Higgins RP, Thiel H, editors. Introduction to the study of Meiofauna. Washington, DC: Smithsonian Institution Press; 1988. p. 328–31.
Neuhaus B. Kinorhyncha (=Echinodera). In: Schmidt-Rhaesa A, editor. Handbook of zoology. Gastrotricha, Cycloneuralia and Gnathifera, vol 1: Nematomorpha, Priapulida, Kinorhyncha, Loricifera. Berlin: De Gruyter; 2013. p. 181–348.
Sørensen MV, Pardos F. Kinorhynchs systematic and biology—an introduction to the study of kinorhynchs, inclusive identification keys to genera. Meiofauna Mar. 2008;6:21–73.
Landers SC, Sørensen MV. Two new species of Echinoderes (Kinorhyncha, Cyclorhagida), E. romanoi sp. n. and E. joyceae sp. n., from the Gulf of Mexico. ZooKeys. 2016;594:51–71. doi:10.3897/zookeys.594.8623.
Neves R, Sørensen MV, Herranz M. First account on kinorhynchs from Portugal, with the description of two new species: Echinoderes lusitanicus sp. nov. and E. reicherti sp. nov. Mar. Biol Res. 2016. doi:10.1080/17451000.2016.1154973.
Sánchez N, Yamasaki H. Two new Pycnophyidae species (Kinorhyncha: Allomalorhagida) from Japan lacking ventral tubes in males. Zool Anz A J Comp Zool. 2016. doi:10.1016/j.jcz.2016.04.001.
Sánchez N, Yamasaki H, Pardos F, Sørensen MV, Martínez A. Morphology distangles the systematics of a ubiquitous but elusive meiofaunal group (Kinorhyncha: Pycnophyidae). Cladistics. 2016. doi:10.1111/cla.12143.
Sørensen MV, Dal Zotto M, Rho HS, Herranz M, Sánchez N, Pardos F, Yamasaki H. Phylogeny of Kinorhyncha based on morphology and two molecular loci. PLoS One. 2015;10(7):e0133440. doi:10.1371/journal.pone.0133440.
Sørensen MV, Gąsiorowski L, Randsø PV, Sánchez N, Neves RC. First report of kinorhynchs from Singapore, with the description of three new species. Raffles Bull Zool. 2016;64:27–30. doi:10.1016/j.jcz.2016.02.004.
Sørensen MV, Herranz M, Landers SC. A new species of Echinoderes (Kinorhyncha: Cyclorhagida) from the Gulf of Mexico, with a redescription of Echinoderes bookhouti Higgins, 1964. Zool Anz A J Comp Zool. 2016. doi:10.1016/j.jcz.2016.04.004.
Yamasaki H. Ryuguderes iejimaensis, a new genus and species of Campyloderidae (Xenosomata: Cyclorhagida: Kinorhyncha) from a submarine cave in the Ryukyu Islands, Japan. Zool Anz A J Comp Zool. 2016. doi:10.1016/j.jcz.2016.02.003.
Adrianov AV. First record of kinorhynchs from the Sea of Japan. Zool Zhur. 1989;68:17–27.
Adrianov AV, Malakhov VV. Cephalorhyncha of the world ocean. Moscow: KMK Scientific Press; 1999.
Neuhaus B, Blasche T. Fissuroderes, a new genus of Kinorhyncha (Cyclorhagida) from the deep sea and continental shelf of New Zealand and from the continental shelf of Costa Rica. Zool Anz A J Comp Zool. 2006;245:19–52. doi:10.1016/j.jcz.2006.03.003.
Higgins RP. A new species of Echinoderes (Kinorhyncha: Cyclorhagida) from a coarse-sand California beach. Trans Am Microsc Soc. 1986;105:266–73.
Sørensen MV. A new kinorhynch genus from the Antarctic deep sea and a new species of Cephalorhyncha from Hawaii (Kinorhyncha: Cyclorhagida: Echinoderidae). Org Divers Evol. 2008;8(230):c1–18. doi:10.1016/j.ode.2007.11.003.
Alper A, Sönmez S, Sak S, Karaytuğ S. Marine harpacticoid (Copepoda, Harpacticoida) fauna of Dilek peninsula (Aydın, Turkey). Turk J Zool. 2015;39(4):580–6. doi:10.3906/zoo-1405-67.
Sönmez S, Sak S, Karaytuğ S. A new species of Arenosetella Wilson, 1932 from Turkey with notes on the genus (Copepoda, Harpacticoida, Ectinosomatidae). Zoosyst Evol. 2016;2(1):119–29. doi:10.3897/zse.92.7003.
Băcescu, M. Le role des iles dans la dispersion récente des espèces indo-pacifiques en Méditerranée occidentale et quelques observations sur la faune marine de L’ile des serpents, en comparaison avec celle peuplants les parages prébosphoriques de la Mer Noire. In: V. Le peuplement des iles Méditerranéennes et la problème de l’insularité. Banyuls-sur-Mer. Paris: Centre National de la Recherche Scientifique; 1961. p. 243–53.
Sönmez S, Köroğlu NO, Karaytuğ S. First record of Family Echinoderidae Zelinka, (1894) (Kinorhyncha: Cyclorhagida) from Turkish marine coasts. Biharean Biol. 2016;10:151205.
Ürkmez D, Pardos F, Sezgin M, Karaçuha M, Öksüz İ. Echinoderes dujardinii Claparède, 1863 (Kinorhyncha, Cyclorhagida): a new record for the kinorhynch fauna of Turkey. Ecol Montenegrina. 2016;6:1–8.
Delamare-Deboutteville C. Recherches sur l’écologie et la répartition du mystacocaride Derocheilocaris remanei Delamare et Chappius en Méditerranée. Vie Milieu. 1954;4:459–69.
Dujardin F. Sur un petit animal marin, l’Echinodère formant un type intermédiaire entre les Crustacés et les Vers. Ann Sci Nat (3) Zool. 1851;15:158–73.
Zelinka K. Über die Organisation von Echinoderes. Verh Dtsch Zool Ges. 1894;4:46–9.
Sørensen MV. Phylogenetic analysis of the Echinoderidae (Kinorhyncha: Cyclorhagida). Org Divers Evol. 2008;8:233–46. doi:10.1016/j.ode.2007.11.002.
Van der Land J. Systematics, zoogeography, and ecology of the Priapulida. Zool Verh. 1970;112(1):1–118.
Bang-Berthelsen IH, Schmidt-Rhaesa A, Kristensen RM. Loricifera. In: Schmidt-Rhaesa A, editor. Handbook of zoology. Gastrotricha, Cycloneuralia and Gnathifera, vol 1: Nematomorpha, Priapulida, Kinorhyncha. Berlin: De Gruyter; 2013. p. 349–71.
Kristensen RM. Loricifera, a new phylum with Aschelminthes characters from the meiobenthos. Zool Syst Evol-Forsch. 1983;21:163–80.
Schmidt-Rhaesa A. Priapulida. In: Schmidt-Rhaesa A, editor. Handbook of zoology. Gastrotricha, Cycloneuralia and Gnathifera, vol 1: Nematomorpha, Priapulida, Kinorhyncha, Loricifera. Berlin: De Gruyter; 2013. p. 147–80.
Storch V, Alberti G, Rosito RM, Sotto FB. Some ultrastructural observations on Tubiluchus philippinensis (Priapulida), a new faunal element of Philippine coastal waters. Philipp Sci. 1985;22:144–56.
Nebelsick M. Sensory spots of Echinoderes capitatus (Zelinka, 1928) (Kinorhyncha, Cyclorhagida). Acta Zool. 1992;73(3):185–95.
Neuhaus B. Postembriyonic development of Paracentrophyes praedictus (Homalorhagida): neoteny questionable among the Kinorhyncha. Zool Scr. 1995;24:179–92.
Sørensen MV, Accogli G, Hansen JG. Postembryonic development in Antygomonas incomitata (Kinorhyncha: Cyclorhagida). J Morphol. 2010;271:863–82. doi:10.1002/jmor.10844.
Higgins RP, Shirayama Y. Dracoderidae, a new family of the cyclorhagid Kinorhyncha from the Inland Sea of Japan. Zool Sci. 1990;7:939–46.
Sørensen MV, Herranz M, Rho HS, Min W, Yamasaki H, Sánchez N, Pardos F. On the genus Dracoderes Higgins & Shirayama, 1990 (Kinorhyncha: Cyclorhagida) with a redescription of its type species, D. abei, and a description of a new species from Spain. Mar Biol Res. 2012;8:210–32. doi:10.1080/17451000.2011.615328.
Higgins RP. The Atlantic barrier reef ecosystem at Carrie Bow Cay, Belize, II: Kinorhyncha. Smithson Contrib Mar Sci. 1983;18:1–131.
Zelinka K. Monographie der Echinodera. Leipzig: Verlag Wilhelm Engelmann; 1928.
Sørensen MV, Pardos F, Herranz M, Rho HS. New data on the genus Paracentrophyes (Homalorhagida, Kinorhyncha), with the description of a new species from the West Pacific. Open Zool J. 2010;3:42–59. doi:10.2174/1874336601003010042.
Herranz M, Pardos F. Fissuroderes sorenseni sp. nov. and Meristoderes boylei sp. nov.: first Atlantic recording of two rare kinorhynch genera, with new identification keys. Zool Anz A J Comp Zool. 2013;253:93–111. doi:10.1016/j.jcz.2013.
Authors’ contributions
NOY did scanning electron micrographs, MVS did measurements and light micrographs, draw line art, NOY, MVS and SK read and wrote the final manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank to Dr. Serdar Sak and Dr. Serdar Sönmez for their help in collecting material from Çalış.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Yıldız, N.Ö., Sørensen, M.V. & Karaytuğ, S. A new species of Cephalorhyncha Adrianov, 1999 (Kinorhyncha: Cyclorhagida) from the Aegean Coast of Turkey. Helgol Mar Res 70, 24 (2017). https://doi.org/10.1186/s10152-016-0476-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10152-016-0476-5