
Abstract
The objective was to investigate the levels of TWIST1 in normal and OA cartilage and examine its role in regulating gene expression in chondrocytes. Human cartilage tissues and chondrocytes were obtained at autopsy from normal knee joints and from OA-affected joints at the time of total knee arthroplasty. TWIST1 expression was increased in human OA knee cartilage compared to normal knee cartilage. TWIST1 induced matrix metalloproteinase 3 (MMP3) expression without direct binding to MMP3 promoter and increased the 5-hydroxymethylcytosine (5hmC) level at the MMP3 promoter. The effect of TWIST1 on expression of TET family (TET1, 2 and 3) was measured in stable TWIST1 transfected TC28 cells, and TET1 expression was up-regulated. TWIST1 dependent upregulation of Mmp3 expression was suppressed in Tet triple KO fibroblast derived from mouse ES cells. Increased TWIST1 expression is a feature of OA-affected cartilage. We identified a novel mechanism of catabolic reaction where TWIST1 up-regulates MMP3 expression by enriching 5hmC levels at the MMP3 promoter via TET1 induction. These findings implicate TWIST1 as an important factor regulating OA related gene expression. Clarifying epigenetic mechanisms of 5hmC induced by TWIST1 is a critical molecule to understanding OA pathogenesis.
Similar content being viewed by others


Introduction
Dysregulated chondrocyte activation and de-differentiation in articular cartilage are important factors in Osteoarthritis (OA) pathogenesis1,2. Twist-related protein1 (TWIST1) is known as Class A basic helix-loop-helix protein 38 (bHLHa38), and necessary for the embryonic development through epithelial-mesenchymal transitions (EMT), and regulates the formation of mesoderm and morphogenesis3,4,5,6. TWIST1 regulates cellular determination and differentiation of several lineages including myogenesis and osteogenesis7. A critical role in mesoderm development is indicated by the association of TWIST1 gene mutations with Saethre-Chotzen syndrome, characterized by a broad range of congenital anomalies, including fusion of the skin between the second and third fingers, short stature, craniosynostosis, head and facial asymmetry and ptosis8,9. Twist1 is expressed in immature chondrocytes, but decreases in mature chondrocytes10. Twist1 is known not only as a suppresser of Runx2 which is necessary for osteoblast differentiation and bone formation, but also as a regulator of mesenchymal stem cell differentiation into chondrocytes11. Twist1 interferes with chondrocyte gene expression through Wnt and BMP-dependent signaling10. Twist1 also binds to the 3′UTR and the HMG DNA-binding domain of Sox9, a master regulator of chondrogenesis, and suppresses Sox9-dependent gene activation12,13. However, expression of TWIST1 in mature normal and OA cartilage and its effects in mature chondrocytes are unknown.
Recently, Twist1 was also identified as a key factor of cancer metastasis and stemness through EMT14,15, correlating tumor invasion with MMP1, MMP2 and MMP9 regulation16,17. MMPs are classified based on substrate specificity (Collagenase (MMP1, 8 and 13), Gelatinase (MMP2 and 9), Stromelysin (MMP3, 10 and 11)), and mediate cartilage degradation in OA18,19,20. In this regard, we were interested in the potential link between TWIST1 and MMPs in human chondrocytes and OA pathogenesis.
Cytosine methylation changes the interactions between proteins and DNA, and methylation changes at CpG sites in promoter regions have been suggested to mediate OA related gene expression21. In OA chondrocytes, some MMP promoters show decreased methylation at CpG loci compared to normal chondrocytes22. Recently, 5-hydroxymethylcytosine (5hmC) has been identified as a new constituent of mammalian DNA, and recognized as the sixth base. Conversion of 5mC to 5hmC by TET (ten eleven translocation) family of Fe(II) and 2-oxoglutarate-dependent enzymes facilitates DNA demethylation, and regulates gene expression23,24,25. The 5hmC levels were enriched in the central nervous system, and play important roles in embryonic stem cells26,27. In OA chondrocytes, a global increase of 5hmC and increased locus specific 5hmC at CpG sites in the MMP1 and MMP3 were reported compared with normal chondrocytes28. However, the mechanisms leading to 5hmC changes in chondrocytes are largely unknown.
Here, we found that TWIST1 was upregulated in human OA knee cartilage tissue compared with normal knee cartilage. In this study, we focused on the relationship between TWIST1 and MMP expression through 5hmC status in MMP3 promoter CpG sites.
Results
TWIST1 is highly expressed in human OA cartilage
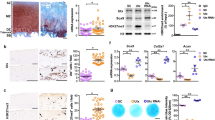
We analyzed TWIST1 expression in cartilage by real-time qRT-PCR and IHC. TWIST1 mRNA levels were significantly increased in human OA knee cartilages compared to normal knee cartilages (approximately 10-fold up-regulation) (Fig. 1A). By IHC, TWIST1 positive cells were observed in human OA cartilage tissues, mainly in chondrocyte clusters, but few in normal cartilage regions (Fig. 1B). The TWIST1 expression was up-regulated after IL-1β (interleukin-1β) and TNFα (tumor necrosis factor α) stimulation (Fig. 1C,D).

(A) TWIST1 gene expression in cartilage tissues from 7 normal donors and 8 OA donors were analyzed by real-time qRT-PCR. TWIST1 gene expression levels in OA cartilage tissues are relative to normal cartilage tissues and normalized to GAPDH. Values are the mean ± SEM ratio. (B) Immunohistochemical staining for TWIST1 in human normal and OA cartilage tissue. **P < 0.01. Left scale bars, 500 μm. Right scale bar, 200 μm. (C) The TWIST1 expression changes after IL-1β and TNFα stimulation in human primary chondrocyte were confirmed by real-time PCR. Values are the mean ± SEM. *P < 0.05. (D) The TWIST1 expression changes after 12 hours IL-1β and TNFα stimulation in human primary chondrocyte were confirmed by Western blotting.
TWIST1 induces MMP3 expression
Matrix metalloproteinases are major factors contributing joint tissue destruction29. Immunohistochemistry showed, in OA cartilage, MMP3 is highly expressed, similar to TWIST1 (Fig. 2A). This prompted us to test whether TWIST1 would be a regulator of MMPs in chondrocytes. We examined MMP gene expression by real-time qRT-PCR in human primary chondrocytes following Ad-TWIST1 infection compared with Ad-GFP infection. MMP3 was significantly up-regulated by Ad-TWIST1 (5–10 fold change) (Fig. 2B). In TC28 cells, MMP3 mRNA level was also increased by Ad-TWIST1 infection by approximately 3 times (Fig. 2C). Ad-TWIST1 up-regulated almost all MMPs, but MMP1 and MMP3 increased much more compared with MMP2, 8, 9, 10, 11, 12 and 13 in human primary chondrocytes (Fig. 3).

(A) Immunohistochemistry analysis of human cartilage tissues. Cartilage sections were obtained from normal and OA-affected cartilages. Sections were stained with MMP3 antibody. Left scale bars, 500 μm. Right scale bar, 200 μm. (B) MMP3 expression was detected by real-time qRT-PCR in human chondrocytes from 3 donors after Ad-GFP or Ad-TWIST1 infection at 72 hours. Values are the mean ± SEM of three independent experiments for each donor. (C) Relative MMP3 expression level in TC28 after Ad-GFP or Ad-TWIST1 infection. Values are the mean ± SEM. *P < 0.05, ***P < 0.001.

Normal human chondrocytes from 3 donors were infected Ad-GFP or Ad-TWIST1 for 72 hours. MMP expression was analyzed by real-time qRT-PCR. Values are the mean ± SEM of three independent experiments for each donor. *P < 0.05, **P < 0.01, ***P < 0.001.
Methylation status of CpG loci in MMP3 promoter
We confirmed 5hmC enrichment in human OA cartilage by immunohistochemistry (Fig. 4A). In TC28 cells, a bisulfite DNA sequencing analysis showed the MMP3 promoter CpG loci were methylated, and Ad-TWIST1 didn’t affect the methylation status compared with non-treated cells (NTC) and Ad-GFP-treated group (Fig. 4B). However, 5hmC status level around a CCGG site in MMP3 promoter was specifically increased, but 5mC status level didn’t change by Ad-TWIST1 compared with Ad-GFP (Fig. 4C). In human chondrocytes, TET1 and TET3 mRNA level were increased by Ad-TWIST1 infection by approximately 2 fold (Fig. 4D). These results suggest TWIST1 promotes 5hmC of specific MMP3 promoter sites through TET1 and TET3 up-regulation.

(A) Immunohistochemistry analysis of human cartilage tissues. Cartilage sections were obtained from normal and OA-affected cartilages. Sections were stained with 5hmC anti body. Left scale bars, 500 μm. Right scale bar, 200 μm. (B) Bisulfite genomic sequencing results of the MMP3 promoter regions in TC28 cells after Ad-GFP or Ad-TWIST1 infection at 72 hours. Black, methylated; white, unmethylated. (C) To detect locus-specific 5hmC and 5mC status at CCGG site in MMP3 promoter, restriction enzyme digestion and quantitative polymerase chain reaction were performed. *P < 0.05. (D) TET1, TET2, and TET3 expressions were detected by real-time qRT-PCR in human chondrocytes after Ad-GFP or Ad-TWIST1 infection at 72 hours. Values are the mean ± SEM of three independent experiments. *P < 0.05.
TWIST1 regulates TET1 expression in TC28 cells
To further investigate the mechanism of 5hmC enrichment by TWIST1, we generated Myc-tagged TWIST1 stably transfected TC28 cells (Myc-TWIST1-TC28) using piggyBac transposon (Fig. 5A,B). In Myc-TWIST1-TC28 cells TET1 expression was dramatically increased, but there were no differences in TET2 and TET3 in western blot (Fig. 5C,D). This result suggests that TWIST1 increases 5hmC levels at the MMP3 promoter through TET1 up-regulation. Immunohistochemistry showed, in human OA cartilage tissue, TET1 is highly expressed, similar to TWIST1 (Fig. 5E). The TET1 expression increased in both chondrocytes and TC28 cell after TWIST1 overexpression. Therefore we focused on the relationship TET1 and MMP3.

(A) Schematic representation of the human TWIST1 protein. (B) Myc-tagged TWIST1 or TWIST1-ΔbHLH stably transfected TC28 cells were generated. Myc expression was analyzed by Western blotting. (C) TET1, 2, and 3 gene expression levels in TWIST1 stably transfected TC28 cells (TC28-TWIST1) were determined by quantitative PCR. Stable TWIST1-ΔHLH transfected TC28 cells (TC28-TWIST1-ΔHLH) were used as control. (D) Western blot analysis of TET 1, 2 and 3 expression in TC28-TWIST1 and TC28-TWIST1-ΔHLH cells. All gels for western blot analysis were run under the same experimental conditions, and western blot images were cropped with a grey cropping line. (E) Immunohistochemistry analysis of human cartilage tissues. Cartilage sections were obtained from normal and OA-affected cartilages. Sections were stained with TET1 anti body. Scale bar, 200 μm.
TET1 suppression by siRNA inhibited MMP3 enhancement by TWIST1 in human chondrocytes
To test if TET1 plays a critical role in TWIST1 dependent MMP3 upregulation, first we treated human chondrocytes TET1 siRNA for 36 hours, the cells were infected with Ad-TWIST1 and cultured for 72 hours. TWIST1 dependent MMP3 upregulation was suppressed by siTET1 treatment (Fig. 6A).

(A) The siRNA for TET1 was transfected to human primary chondrocytes, and after 36 hours, cells were infected with Ad-GFP or Ad-TWIST1 and cultured for 72 hours. The MMP3 gene expression was determined by real-time qRT-PCR. (B) Fibroblasts were derived from Tet1, 2 and 3 triple knock out mice embryonic stem cells, and were transfected Ad-GFP or Ad-TWIST1 for 72 hours. Mmp3 gene expression was determined by quantitative PCR. *P < 0.05.
TWIST1 does not induce Mmp3 expression in Tet-deficient fibroblasts
To reveal the role of TETs more clearly, we obtained fibroblasts from wild-type or Tet1, 2 and 3 triple knockout mutant embryonic stem cells (ESCs)30,31. Wild-type and Tet-KO fibroblasts were infected with Ad-GFP or Ad-TWIST1 and cultured for 72 hours. TWIST1 dependent upregulation of Mmp3 expression was markedly inhibited in Tet-KO fibroblast cells (Fig. 6B), suggesting that Tet proteins are essential for driving TWIST1 dependent MMP3 induction.
Discussion
Some reports showed that Twist1 inhibited transactivator function of master chondrogenic regulator Sox912,13, and knockdown of Twist1 transcript levels caused increased expression of cartilage-specific genes, such as aggrecan and type II collagen10,32. Our data shows that TWIST1 was expressed at much higher levels in OA cartilage compared with normal cartilage, providing the first evidence to reveal a correlation between TWIST1 and OA.
Methylation of DNA in the promoter regions of a target gene is well known to regulate gene expression33. In cartilage and chondrocytes, some reports have shown that the DNA methylation status was regulated in individual gene promoters34,35,36. This implicates DNA methylation as a mechanism of OA-related gene expression. For instance, at the COL10A1 promoter, two CpG sites were demethylated during chondrocyte differentiation from mesenchymal stem cells, and correlated with COL10A1 expression37. Hypermethylated COL9A1 enhancer was associated with transcriptional repression in OA cartilage38, and the SOX9 promoter was hypomethylated during chondrogenesis of human synovium-derived mesenchymal stem cells39. Several studies reported DNA methylation changes in OA cartilage34,40,41,42,43. Jeffries et al. performed genome-wide DNA methylation study in cartilage from hip OA patients44. They identified 550 differentially methylated sites in OA, and approximately one-third of known OA susceptibility genes showed differential methylation. The major enzymes that mediate the destructive processes in joint tissues in OA MMPs. Roach et al. examined the methylation status of the promoter regions of MMP3, MMP9, and MMP13 in OA chondrocytes, and revealed the overall percentage of non-methylated site was increased22. We checked MMP3 promoter methylation status by bisulfite sequencing analysis, but the percentage of non-methylated CpG sites did not change after Ad-TWIST1 infection (Fig. 3B). This suggested that TWIST1 regulates MMP3 through other epigenetic mechanisms.
Recently, 5hmC was discovered as a sixth base of the genome, and converted from 5mC by TET family proteins. Many reports showed 5hmC and TET proteins have important roles of epigenetic reprogramming and regulating tissue-specific gene expression. The highest levels of 5hmC were found in the brain and in embryonic stem cells45,46,47,48. Taylor et al. reported genome-wide mapping of DNA hydroxymethylation in OA chondrocytes49. DNA methylation at promoter regions is associated with gene silencing50. In OA chondrocytes, higher global 5hmC level was observed, and also 5hmC level was increased particularly in the MMP1 and MMP3 promoter28. In our study, bisulfite sequencing results suggest that TWIST1 does not lead to demethylation, but 5hmC levels were increased by TWIST1 in the MMP3 promoter (Fig. 3B,D). This suggests that 5hmC gain may be important but demethylation is not essential for transcriptional regulation of MMP3 by TWIST1. Taylor et al. identified 5hmC distribution to be enriched in the regulatory regions of genes preceding the transcription start site (TSS), as well as in the gene bodies during chondrogenic differentiation in ATDC5 cells51. To clarify 5mC and 5hmC states in the MMP3 promoter, additional TAB-seq (Tet-assisted bisulfite sequencing) analysis should be performed in future studies. Genome-wide mapping results showed a gain in 5hmC in the gene bodies of some OA susceptibility genes49. In our results, 5hmC gain in MMP3 gene body is unknown, but MMP3 induction by TWIST1 was suppressed in human chondrocytes with TET1 knock-down (Fig. 6A) and in Tet triple KO fibroblasts (Fig. 6B), and this results strongly suggests that TET catalyzed enrichment of 5hmC is critical for MMP3 regulation by TWIST1. Previous studies described that globally increased 5hmC level were observed, but TET1 was down-regulated, and there were no significant changes in the expression of TET2 and TET3 in OA chondrocytes28. We also confirmed 5hmC enrichment in OA cartilage by immunohistochemistry (Fig. 3A), but in our case, the mRNA levels of TET1 were only mildly increased in human OA knee cartilages compared to normal knee cartilages (Figure S1). These differences of TET1 expression might be caused by the difference between chondrocytes and cartilage tissues, but we also confirmed by immunohistochemistry that TET1 is highly expressed in OA cluster cells (Fig. 5E). TWIST1 dramatically induced TET1 expression in human chondrocytes and TC28 cells, so our results imply that the 5hmC gain is mediated by TWIST1. Two recent reports described that the inflammatory cytokines IL-1β, TNFα or oncostatin M suppressed TET1 gene expression in chondrocytes28,52, but our results showed TET1 expression didn’t change after IL-1β and TNFα stimulation (Figure S1). These cytokines up-regulated TWIST1 expression (Fig. 1C,D), and TWIST1 overexpression induced TET1 gene expression in human chondrocytes (Figs 4D, 5C and D). Thus, only cytokine stimulation might not be sufficient to regulate TET1 expression through TWIST1 up-regulation. We confirmed TET1 had a critical role in TWIST1 dependent MMP3 upregulation in human primary chondrocytes with siRNA-mediated TET1 knock down (Fig. 6A), and in fibroblasts with TET triple knock-out (Fig. 6B). Therefore, TET expression change through TWIST1 could be an important mechanism for the development of OA.
We observed 5hmC enrichment in the MMP3 promoter (Fig. 3D), but not in the MMP1 promoter (data not shown). Wu et al. performed denovo motif discovery analysis, and identified a CpG-rich sequence bound TET153. The MMP3 promoter has two potential TET1 binding sequences (5′-CCGCAGC-3′ and 5′-GCGCAGC-3′) in reverse complement, but the MMP1 promoter has no TET1 binding sequences. These differences between MMP1 and MMP3 promoters are one potential reason that TET1 induces only MMP3 expression.
Conclusions
This study provides the first evidence that increased expression of TWIST1 is a novel feature of OA-affected cartilage, and TWIST1 promotes catabolic reactions by inducing MMP3 expression through 5hmC gain in MMP3 promoter via regulation of TET1. TWIST1 may be one of core regulators of DNA hydroxymethylation in chondrocytes. Thus, TWIST1 is a promising transcription factor to reveal the detailed epigenetic mechanism in chondrocytes. Dysregulated TWIST1 may be an important factor in OA pathogenesis and a novel treatment target for OA.
Methods
Cartilage tissue procurement, cell isolation and culture
The study was conducted in accordance with the Declaration of Helsinki, and the Scripps Institutional Review Board approved the protocol 09–5162 and informed consent form prior to study initiation. Informed consent was obtained from all patients prior the study. Normal human knee joints were procured at autopsy from 7 donors (mean ± SD age 35.1 ± 11.8 years). OA-affected cartilage was collected at the time of total knee arthroplasty from 8 donors with knee OA (mean ± SD age 69.7 ± 5.0 years). We scored articular cartilage by dividing it into 39 regions based on the International Cartilage Repair Society knee map, and evaluated the cartilage macroscopically using a modified Outerbridge scoring system. Each area was scored on a scale of 1–4, where 1 = intact surface, 2 = fibrillation, 3 = fissuring, and 4 = erosion. The total knee cartilage score could thus ranges from 39 (normal) to 156 (maximum severity). Total knee cartilage scores were then translated into grades 0–IV, where grade 0 = normal (total score 39), grade I = minimal change (total score 40–58), grade II = mild change (total score 59–78), grade III = moderate change (total score 79–97), and grade IV = severe change (total score ≥ 98). We considered grade I and II as normal, and grade III and IV as OA.
Chondrocytes were isolated by collagenase digestion of cartilage and cultured in monolayer as described54. First passage cells were used in the experiments.
TC28 cells (immortalized human primary juvenile costal chondrocytes) were obtained from the American Type Culture Collection (ATCC). These cells were propagated as monolayer cultures in Dulbecco’s modified Eagle’s medium (DMEM). All media were supplemented with 10% heat-inactivated FBS, 100 units/mL penicillin, and 100 mg/mL streptomycin. The cells were maintained at 37 °C in a humidified atmosphere with 5% CO2.
RNA isolation and quantitative real time-PCR analysis
Total RNA was isolated from cartilage tissue or cells by extracting the homogenate in TRIzol (Life Technologies). Complementary DNA (cDNA) was synthesized using a PrimeScript RT Reagent Kit (Takara Bio). Quantitative polymerase chain reaction (qPCR) was performed using a LightCycler 96 (Roche). TaqMan Gene Expression Assay probes for TWIST1 (Hs00361186_m1), MMP1 (Hs00899658_m1), MMP2 (Hs01548727_m1), MMP3 (Hs00968305_m1), MMP8 (Hs01029057_m1), MMP9 (Hs00234579_m1), MMP10 (Hs00233987_m1), MMP11 (Hs00968295_m1), MMP12 (Hs00159178_m1), MMP13 (Hs00233992_m1), TET1 (Hs00286756_m1), TET2 (Hs00325999_m1) and TET3 (Hs00379125_m1) according to the instructions of the manufacturer (Applied Biosystems). Gene expression levels of GAPDH (Hs02758991_g1) were assessed as an internal control.
Western blot analysis
TC28 cells and chondrocytes were seeded in 24-well dishes at a density of 1 × 104 cells/dish 24 hours before transfection. Whole cell lysates were prepared in a lysis buffer (50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% Triton X-100) containing a protease/phosphatase inhibitor cocktail (Cell Signaling Technology) at the indicated time points. Proteins were electrophoresed on 4–20% SDS polyacrylamide gels and were transferred to nitro cellulose membranes (Bio-Rad). Blots were blocked with Odyssey Blocking Buffer (LI-COR) at room temperature for 30 minutes. The primary antibodies used were: mouse anti-GAPDH monoclonal antibody (mAb) (Ambion), mouse anti-TWIST1 mAb (Sigma-Aldrich), rabbit anti-MMP3 mAb (Cell Signaling Technology), rabbit anti-Myc-Tag polyclonal antibody (pAb) (Cell Signaling Technology), mouse anti-TET1 mAb (GeneTex), rabbit anti-TET2 pAb (GeneTex), rabbit anti-TET3 pAb (GeneTex). The secondary antibodies (goat anti-rabbit – IRDye 800 (1:5000 dilution) and goat anti mouse – IRDye 680 (1:10,000 dilution)) were applied. Bands on the blots were visualized using LiCor Odyssey.
Plasmid
TWIST1 (Myc-DDK-tagged)-Human twist 1 was obtained from Origene. For making piggyBac Myc-TWIST1 transposon vector, the human TWIST1 coding region was obtained from the cDNA with KOD Hot Start DNA Polymerase (Novagen), and subcloned into a piggyBac vector. The primer sequences to clone the coding region of TWIST1 were as follows: Forward primer: 5′-ATGATGCAGGACGTGTCCAGCTCGCC-3′; Reverse primer: 5′-CTAGTGGGACGCGGACATGGACCAGGC-3′. To induce DNA binding defective mutant of TWIST1, basic helix-loop-helix region deleted TWIST1 open reading frame (ORF) over expression vector (piggyBac TWIST1-ΔbHLH vector) was constructed55.
Adenovirus
TWIST1-adenovirus (Ad-TWIST1) (SignaGen Laboratories) and eGFP-adenovirus (Ad-GFP) (VECTOR Biolabs) were used in this study. TC28 cells were infected with adenovirus at a multiplicity of infections (MOI) of 100 plaque forming units (PFU)/cell, and human chondrocytes were infected at 1000 MOI.
Bisulfite genomic sequencing analysis
To study DNA methylation status by methylation-specific PCR in TC28 cells after TWIST1 over expression, the DNA was treated with bisulfite using the EZ DNA methylation-Gold KIT (ZYMO Research Corp). After PCR amplification, the target DNA fragment was subcloned into pCR4-TOPO (Life Technologies), and sequenced.
Analyzing and quantitating 5hmC in MMP3 promoter
5hmC levels were detected using EpiMark 5hmC and 5mC Analysis Kit (New England Biolabs). TC28 cells were seeded in 6-well plates at a density of 2 × 105 cells/well 24 hours before Ad-GFP or Ad-TWIST1 infection. Genomic DNA was extracted from the cells 2 days after virus infection by using PureLink Genomic DNA Mini Kit. Genomic DNA (10 μg) was treated with 30 units of T4 β-glucosyltransferase (T4-BGT) at 37 °C for 18 hours. 100 units of MspI or 50 units of HpaII was added to the glucosylated DNA, and incubated the sample at 37 °C for 12 hours. Proteinase K treatment was done at 40° for 30 minutes and inactivated at 95° for 10 minutes. The percentages of 5hmC were calculated EpiMark comparative Ct method. The sequences of primers for MMP3 promoter were 5′-GGAGGGGAAAAGGTTGAAAG-3′ (forward) and 5′-CCACGTAGCTGCTCCATAAATAG-3′ (reverse).
Immunohistochemistry
Immunohistochemistry studies were performed to investigate the expression pattern of TWIST1 in human cartilage tissue. The tissue sections were incubated with 10% trypsin at 37 °C for 60 minutes for antigen retrieval. After washing with phosphate buffered saline, the sections were blocked with 10% normal goat serum for 60 minutes at room temperature. Rabbit anti-human TWIST1 polyclonal antibody (Cell Signaling Technology), rabbit polyclonal anti-MMP3 antibody (Cell Signaling Technology), rabbit polyclonal anti-5hmC antibody (Active Motif) and mouse anti-TET1 antibody (GeneTex) were applied and incubated overnight at 4 °C. The sections were incubated with secondary antibody, substrate and hematoxylin were performed.
Fibroblasts from Tet1, 2 and 3 triple knock out embryonic stem cells
Mesenchymal cells were produced through induction of embryoid bodies (EBs)56. Undifferentiated Tet triple knock out (TKO) embryonic stem (ES) cells were dissociated into single cell suspension by treatment with 0.25% trypsin (Invitrogen) and 0.02% EDTA (SIGMA-Aldrich) in PBS (trypsin-EDTA), resuspended in 10% FBS-DMEM (Invitrogen), and then transferred to suspension culture on Petri dishes. After 3 days of EB formation, cell aggregates were cultured on the gelatin-coated dishes with 10% FBS-DMEM supplemented with 10–6 M all-trans retinoic acid (RA, SIGMA-Aldrich). After 8 days, the outgrowths were dissociated, filtered by 40-μm meshes (BD Biosciences Discovery Labware, MA, USA) and cultured briefly (~1 hour) on the gelatin-coated culture dishes to select only the fibroblast cells. Then, the attached cells were cultured in 10% FBS-αMEM supplemented with 10 ng/ml bFGF (Upstate Biotechnology, Lake Placid, NY, USA) until confluence.
Statistical analysis
Data are reported as mean ± SEM of the indicated number of independent experiments. Some experiments were conducted with primary chondrocytes samples from three different donors, each performed in triplicate. Statistically differences were determined by t-tests or one-way ANOVA with post hoc comparisons using Tukey’s test. P values less than 0.05 were considered significant.
Additional Information
How to cite this article: Hasei, J. et al. TWIST1 induces MMP3 expression through up-regulating DNA hydroxymethylation and promotes catabolic responses in human chondrocytes. Sci. Rep. 7, 42990; doi: 10.1038/srep42990 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Serra, R. et al. Expression of a truncated, kinase-defective TGF-beta type II receptor in mouse skeletal tissue promotes terminal chondrocyte differentiation and osteoarthritis. The Journal of cell biology 139, 541–552 (1997).
Yang, X. et al. TGF-beta/Smad3 signals repress chondrocyte hypertrophic differentiation and are required for maintaining articular cartilage. The Journal of cell biology 153, 35–46 (2001).
Leptin, M. twist and snail as positive and negative regulators during Drosophila mesoderm development. Genes & development 5, 1568–1576 (1991).
Bialek, P. et al. A twist code determines the onset of osteoblast differentiation. Developmental cell 6, 423–435 (2004).
Chen, Z. F. & Behringer, R. R. twist is required in head mesenchyme for cranial neural tube morphogenesis. Genes & development 9, 686–699 (1995).
Martin, A. & Cano, A. Tumorigenesis: Twist1 links EMT to self-renewal. Nature cell biology 12, 924–925, doi: 10.1038/ncb1010-924 (2010).
Lee, M. S., Lowe, G. N., Strong, D. D., Wergedal, J. E. & Glackin, C. A. TWIST, a basic helix-loop-helix transcription factor, can regulate the human osteogenic lineage. Journal of cellular biochemistry 75, 566–577 (1999).
Gallagher, E. R., Ratisoontorn, C. & Cunningham, M. L. In GeneReviews(R) (eds R. A. Pagon et al.) (1993).
Howard, T. D. et al. Mutations in TWIST, a basic helix-loop-helix transcription factor, in Saethre-Chotzen syndrome. Nature genetics 15, 36–41, doi: 10.1038/ng0197-36 (1997).
Reinhold, M. I., Kapadia, R. M., Liao, Z. & Naski, M. C. The Wnt-inducible transcription factor Twist1 inhibits chondrogenesis. The Journal of biological chemistry 281, 1381–1388, doi: 10.1074/jbc.M504875200 (2006).
Hinoi, E. et al. Runx2 inhibits chondrocyte proliferation and hypertrophy through its expression in the perichondrium. Genes & development 20, 2937–2942, doi: 10.1101/gad.1482906 (2006).
Gu, S., Boyer, T. G. & Naski, M. C. Basic helix-loop-helix transcription factor Twist1 inhibits transactivator function of master chondrogenic regulator Sox9. The Journal of biological chemistry 287, 21082–21092, doi: 10.1074/jbc.M111.328567 (2012).
Goodnough, L. H. et al. Twist1 mediates repression of chondrogenesis by beta-catenin to promote cranial bone progenitor specification. Development 139, 4428–4438, doi: 10.1242/dev.081679 (2012).
Yang, J. et al. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell 117, 927–939, doi: 10.1016/j.cell.2004.06.006 (2004).
Jung, H. Y. & Yang, J. Unraveling the TWIST between EMT and cancer stemness. Cell Stem Cell 16, 1–2, doi: 10.1016/j.stem.2014.12.005 (2015).
Weiss, M. B. et al. TWIST1 is an ERK1/2 effector that promotes invasion and regulates MMP-1 expression in human melanoma cells. Cancer research 72, 6382–6392, doi: 10.1158/0008-5472.CAN-12-1033 (2012).
Zhao, X. L. et al. Promotion of hepatocellular carcinoma metastasis through matrix metalloproteinase activation by epithelial-mesenchymal transition regulator Twist1. Journal of cellular and molecular medicine 15, 691–700, doi: 10.1111/j.1582-4934.2010.01052.x (2011).
Blanco Garcia, F. J. Catabolic events in osteoarthritic cartilage. Osteoarthritis and cartilage/OARS, Osteoarthritis Research Society 7, 308–309, doi: 10.1053/joca.1998.0174 (1999).
Konttinen, Y. T. et al. Analysis of 16 different matrix metalloproteinases (MMP-1 to MMP-20) in the synovial membrane: different profiles in trauma and rheumatoid arthritis. Annals of the rheumatic diseases 58, 691–697 (1999).
Kevorkian, L. et al. Expression profiling of metalloproteinases and their inhibitors in cartilage. Arthritis and rheumatism 50, 131–141, doi: 10.1002/art.11433 (2004).
Barter, M. J., Bui, C. & Young, D. A. Epigenetic mechanisms in cartilage and osteoarthritis: DNA methylation, histone modifications and microRNAs. Osteoarthritis and cartilage/OARS, Osteoarthritis Research Society 20, 339–349, doi: 10.1016/j.joca.2011.12.012 (2012).
Roach, H. I. et al. Association between the abnormal expression of matrix-degrading enzymes by human osteoarthritic chondrocytes and demethylation of specific CpG sites in the promoter regions. Arthritis and rheumatism 52, 3110–3124, doi: 10.1002/art.21300 (2005).
Szwagierczak, A., Bultmann, S., Schmidt, C. S., Spada, F. & Leonhardt, H. Sensitive enzymatic quantification of 5-hydroxymethylcytosine in genomic DNA. Nucleic acids research 38, e181, doi: 10.1093/nar/gkq684 (2010).
Bhutani, N., Burns, D. M. & Blau, H. M. DNA demethylation dynamics. Cell 146, 866–872, doi: 10.1016/j.cell.2011.08.042 (2011).
Tahiliani, M. et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324, 930–935, doi: 10.1126/science.1170116 (2009).
Pastor, W. A. et al. Genome-wide mapping of 5-hydroxymethylcytosine in embryonic stem cells. Nature 473, 394–397, doi: 10.1038/nature10102 (2011).
Szulwach, K. E. et al. 5-hmC-mediated epigenetic dynamics during postnatal neurodevelopment and aging. Nature neuroscience 14, 1607–1616, doi: 10.1038/nn.2959 (2011).
Taylor, S. E., Smeriglio, P., Dhulipala, L., Rath, M. & Bhutani, N. A global increase in 5-hydroxymethylcytosine levels marks osteoarthritic chondrocytes. Arthritis & rheumatology 66, 90–100, doi: 10.1002/art.38200 (2014).
Yoshihara, Y. et al. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in synovial fluids from patients with rheumatoid arthritis or osteoarthritis. Annals of the rheumatic diseases 59, 455–461 (2000).
Horii, T. et al. Genome engineering of mammalian haploid embryonic stem cells using the Cas9/RNA system. PeerJ 1, e230, doi: 10.7717/peerj.230 (2013).
Kafer, G. R. et al. 5-Hydroxymethylcytosine Marks Sites of DNA Damage and Promotes Genome Stability. Cell reports 14, 1283–1292, doi: 10.1016/j.celrep.2016.01.035 (2016).
Chakraborty, S. et al. Twist1 promotes heart valve cell proliferation and extracellular matrix gene expression during development in vivo and is expressed in human diseased aortic valves. Developmental biology 347, 167–179, doi: 10.1016/j.ydbio.2010.08.021 (2010).
Jones, P. A. & Takai, D. The role of DNA methylation in mammalian epigenetics. Science 293, 1068–1070, doi: 10.1126/science.1063852 (2001).
Alvarez-Garcia, O. et al. Increased DNA methylation and reduced expression of transcription factors in human osteoarthritis cartilage. Arthritis & rheumatology, doi: 10.1002/art.39643 (2016).
Bonin, C. A. et al. Identification of differentially methylated regions in new genes associated with knee osteoarthritis. Gene 576, 312–318, doi: 10.1016/j.gene.2015.10.037 (2016).
Kim, H., Kang, D., Cho, Y. & Kim, J. H. Epigenetic Regulation of Chondrocyte Catabolism and Anabolism in Osteoarthritis. Molecules and cells 38, 677–684, doi: 10.14348/molcells.2015.0200 (2015).
Zimmermann, P. et al. Correlation of COL10A1 induction during chondrogenesis of mesenchymal stem cells with demethylation of two CpG sites in the COL10A1 promoter. Arthritis and rheumatism 58, 2743–2753, doi: 10.1002/art.23736 (2008).
Imagawa, K. et al. Association of reduced type IX collagen gene expression in human osteoarthritic chondrocytes with epigenetic silencing by DNA hypermethylation. Arthritis & rheumatology 66, 3040–3051, doi: 10.1002/art.38774 (2014).
Ezura, Y., Sekiya, I., Koga, H., Muneta, T. & Noda, M. Methylation status of CpG islands in the promoter regions of signature genes during chondrogenesis of human synovium-derived mesenchymal stem cells. Arthritis and rheumatism 60, 1416–1426, doi: 10.1002/art.24472 (2009).
Rushton, M. D. et al. Characterization of the cartilage DNA methylome in knee and hip osteoarthritis. Arthritis & rheumatology 66, 2450–2460, doi: 10.1002/art.38713 (2014).
den Hollander, W., Ramos, Y. F. & Bos, S. D. Knee and hip articular cartilage have distinct epigenomic landscapes: implications for future cartilage regeneration approaches. 73, 2208–2212, doi: 10.1136/annrheumdis-2014-205980 (2014).
Fernandez-Tajes, J. et al. Genome-wide DNA methylation analysis of articular chondrocytes reveals a cluster of osteoarthritic patients. Annals of the rheumatic diseases 73, 668–677, doi: 10.1136/annrheumdis-2012-202783 (2014).
Kim, K. I., Park, Y. S. & Im, G. I. Changes in the epigenetic status of the SOX-9 promoter in human osteoarthritic cartilage. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research 28, 1050–1060, doi: 10.1002/jbmr.1843 (2013).
Jeffries, M. A. et al. Genome-wide DNA methylation study identifies significant epigenomic changes in osteoarthritic subchondral bone and similarity to overlying cartilage. Arthritis & rheumatology, doi: 10.1002/art.39555 (2015).
Tu, J. et al. MicroRNA-29b/Tet1 regulatory axis epigenetically modulates mesendoderm differentiation in mouse embryonic stem cells. Nucleic acids research 43, 7805–7822, doi: 10.1093/nar/gkv653 (2015).
Dawlaty, M. M. et al. Loss of Tet enzymes compromises proper differentiation of embryonic stem cells. Developmental cell 29, 102–111, doi: 10.1016/j.devcel.2014.03.003 (2014).
Sherwani, S. I. & Khan, H. A. Role of 5-hydroxymethylcytosine in neurodegeneration. Gene 570, 17–24, doi: 10.1016/j.gene.2015.06.052 (2015).
Hahn, M. A. et al. Dynamics of 5-hydroxymethylcytosine and chromatin marks in Mammalian neurogenesis. Cell reports 3, 291–300, doi: 10.1016/j.celrep.2013.01.011 (2013).
Taylor, S. E., Li, Y. H., Wong, W. H. & Bhutani, N. Genome-wide mapping of DNA hydroxymethylation in osteoarthritic chondrocytes. Arthritis & rheumatology 67, 2129–2140, doi: 10.1002/art.39179 (2015).
Branco, M. R., Ficz, G. & Reik, W. Uncovering the role of 5-hydroxymethylcytosine in the epigenome. Nature reviews. Genetics 13, 7–13, doi: 10.1038/nrg3080 (2012).
Taylor, S. E. et al. Stable 5-Hydroxymethylcytosine (5hmC) Acquisition Marks Gene Activation During Chondrogenic Differentiation. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research, doi: 10.1002/jbmr.2711 (2015).
Haseeb, A., Makki, M. S. & Haqqi, T. M. Modulation of ten-eleven translocation 1 (TET1), Isocitrate Dehydrogenase (IDH) expression, alpha-Ketoglutarate (alpha-KG), and DNA hydroxymethylation levels by interleukin-1beta in primary human chondrocytes. The Journal of biological chemistry 289, 6877–6885, doi: 10.1074/jbc.M113.512269 (2014).
Wu, H. & Zhang, Y. Mechanisms and functions of Tet protein-mediated 5-methylcytosine oxidation. Genes & development 25, 2436–2452, doi: 10.1101/gad.179184.111 (2011).
Akasaki, Y. et al. Dysregulated FOXO transcription factors in articular cartilage in aging and osteoarthritis. Osteoarthritis and cartilage/OARS, Osteoarthritis Research Society 22, 162–170, doi: 10.1016/j.joca.2013.11.004 (2014).
Maia, A. M., da Silva, J. H., Mencalha, A. L., Caffarena, E. R. & Abdelhay, E. Computational modeling of the bHLH domain of the transcription factor TWIST1 and R118C, S144R and K145E mutants. BMC bioinformatics 13, 184, doi: 10.1186/1471-2105-13-184 (2012).
Teramura, T. et al. Induction of mesenchymal progenitor cells with chondrogenic property from mouse-induced pluripotent stem cells. Cellular reprogramming 12, 249–261, doi: 10.1089/cell.2009.0086 (2010).
Acknowledgements
We gratefully acknowledge Shantanu Patil MD, Stuart Duffy, Merissa Olmer, and Lilo Creighton for their technical assistance. This work was supported by Uehara Memorial Foundation Postdoctoral Fellowship and grants from the Japan Rheumatism Foundation, NIH (grant numbers AG007996, AG049617, AR050631 and AR065379), the Core Research for the Evolutionary Science and Technology (CREST), JST (grant number 26113008), JSPS KAKENHI (grant numbers 15H02560 and 15K15544), the Takeda Science Foundation, and Bristol-Myers K.K. RA Clinical Investigation Grant to Hiroshi Asahara.
Author information
Authors and Affiliations
Contributions
All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published. Drs. Asahara and Lotz had full access to all of the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis. Study conception and design. Hasei, Teramura, Asahara Lotz. Acquisition of data. Hasei, Teramura, Takehara, Onodera, Olmer. Analysis and interpretation of data. Hasei, Teramura.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Hasei, J., Teramura, T., Takehara, T. et al. TWIST1 induces MMP3 expression through up-regulating DNA hydroxymethylation and promotes catabolic responses in human chondrocytes. Sci Rep 7, 42990 (2017). https://doi.org/10.1038/srep42990
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep42990
- Springer Nature Limited
This article is cited by
-
Alterations in DNA methylation machinery in a rat model of osteoarthritis of the hip
Journal of Orthopaedic Surgery and Research (2024)
-
The essential anti-angiogenic strategies in cartilage engineering and osteoarthritic cartilage repair
Cellular and Molecular Life Sciences (2022)
-
Oral administration of N-acetyl cysteine prevents osteoarthritis development and progression in a rat model
Scientific Reports (2019)
-
hnRNPK S379 phosphorylation participates in migration regulation of triple negative MDA-MB-231 cells
Scientific Reports (2019)
-
Targeting of chondrocyte plasticity via connexin43 modulation attenuates cellular senescence and fosters a pro-regenerative environment in osteoarthritis
Cell Death & Disease (2018)