
Abstract
Protein N-glycosylation (PNG) is crucial for protein folding and enzymatic activities, and has remarkable diversity among eukaryotic species. Little is known of how unique PNG mechanisms arose and evolved in eukaryotes. Here we demonstrate a picture of onset and evolution of PNG components in Golgi apparatus that shaped diversity of eukaryotic protein N-glycan structures, with an emphasis on roles that domain emergence and combination played on PNG evolution. 23 domains were identified from 24 known PNG genes, most of which could be classified into a single clan, indicating a single evolutionary source for the majority of the genes. From 153 species, 4491 sequences containing the domains were retrieved, based on which we analyzed distribution of domains among eukaryotic species. Two domains in GnTV are restricted to specific eukaryotic domains, while 10 domains distribute not only in species where certain unique PNG reactions occur and thus genes harboring these domains are supoosed to be present, but in other ehkaryotic lineages. Notably, two domains harbored by β-1,3 galactosyltransferase, an essential enzyme in forming plant-specific Lea structure, were present in separated genes in fungi and animals, suggesting its emergence as a result of domain shuffling.
Similar content being viewed by others

Introduction
Genes with new functions emerge continuously throughout the tree of life. A new gene arises within a specific phylogenetic lineage, which is not similar in sequence with any other genes in organisms that have split evolutionarily before this time1,2. In terms of how origin of gene novelties occurs, there are two major models: duplication-divergence model, which proposes an initial duplication of an existing gene followed by rapid divergence, and de novo evolution model, which assumes that a new gene emerges out of non-coding DNA1,3,4. Phylogenetic analyses suggested that de novo evolution of new genes occurred throughout evolutionary time, although non-coding DNA sources are not always identified for some claimed de novo genes5. Although less common, gene fusion, by which multiple transcription units merge into one compact unit, is an important source of new gene emergence6. Considering functional and structural significance of evolution, modularity of protein evolution plays a remarkable role in shaping the genomic make-up, which is generally associated with domains. Domains are functional constituents of proteins more conserved than other parts of genes in sequence, and are thus evolutionarily conserved across taxa. Genes arising by duplication-divergence is attributable enormously to domain duplication and divergence, and de novo gene births constantly give rise to new domains7. Likewise, gene fusion essentially occurs through recombination of conserved domains, namely domain shuffling, that are found to be present in different instances8,9.
In this article, we investigate how genes responsible for protein N-glycosylation (PNG) arose in eukaryotes. Particularly, we study the roles that domain emergence and combination as well as sequence divergence played in gene evolution in this process. Glycosylation is one of the most complex post-translation modifications of proteins, which is common for secretory proteins in eukaryotes10. N-linked glycans are widely observed to be crucial for proper folding of proteins, which provide blueprints for precise instruction of protein folding and discrimination signals for quality control systems11. Thus, it can be important for the function of individual glycoproteins, which would further have physiological effects on eukaryotic cells. Biological activities of many therapeutic proteins rely heavily on their glycosylation status. As a result, protein glycosylation is one of the main focuses in the biopharmaceutical research community10,12. Carbohydrates attached to the proteins can be classified into two categories, N-glycans, which are linked to the amide group of asparagine residues, and O-glycans, present on the hydroxyl group of serine, threonine, hydroxylysine and hydroxyproline residues10.
PNG is catalyzed with the form of a rational orchestration of multiple enzymatic formation and breakdown of glycan linkages, which is achieved by glycosyltransferases and glycosidases, occurring in the endoplasmic reticulum (ER) firstly and then in Golgi apparatus. Reaction mechanisms in ER are largely conserved in yeasts, mammals and plants. The mechanisms are initialized at the cytosolic side of the ER membrane by transferring an oligosaccharide precursor, Man5GlcNAc2, from a dolichol lipid carrier onto specific Asn residues constitutive of the consensus sequence Asn-X-Ser/Thr/Cys in nascent proteins10,13,14,15. Afterward, reactions proceed in ER lumen with complete assembly of the Glc3Man9GlcNAc2 precursor catalyzed by sequential orchestration of multiple enzymatic steps10,12,16.
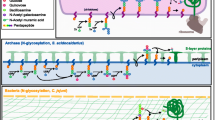
While the assembly mechanism of core N-glycan precursor is conserved in ER, further modifications in Golgi apparatus vary enormously in different eukaryotic lineages, depending on a rich genetic toolbox of enzymes that are used to generate different types of N-glycans; how genes encoding these enzymes emerged and evolved is the focus of this article. In yeasts, a single α1,6 mannose unit is first attached to the glycan by Och1; then, it is elongated by multi-enzyme complexes, M-PolI and M-PolII, to form the α1,6 outer chain backbone containing up to 50 additional mannose residues. It is further decorated with side chains mainly consisting of homopolymeric α1,2 mannosides and heteropolymeric α1,2/α1,3 or α1,2/β1,2 mannosides, catalyzed by Mnn1, Mnn2, Mnn5 and Mnn6, respectively17,18. This machinery confers N-glycans distinguished properties of immense proportions of mannose redsidues in yeasts (Fig. 1a). In higher plants and mammals, however, N-glycans are made up by an enormously greater variety of monosaccharide residues (Fig. 1b and c)10,19,20. Firstly, the high-mannose N-glycan core is trimmed and acetylglucosamine is attached, which are catalyzed by β1,2 N-acetylglucosaminyltransferase I, Golgi α-mannosidase II and β1,2 N-acetylglucosaminyltransferase II sequentially, before it is further modified where they have remarkable differences in plants and animals. In plants, the glycan core is usually substituted by a β1,2 xylose, which is catalyzed by β1,2 xylosyltransferase (β1,2-XylT), and the proximal N-acetylglucosamine is replaced by an α1,3-fucose through catalysis of α1,3-fucosyltransferase (α1,3-FucT); also, in higher plants a typical N-glycan usually contains a Lewis a (Lea) structure, which is formed by attachment of β1,3 galactose and α1,4 fucose to the terminal GlcNAc, facilitated by β1,3-GalT and α1,4-FucT, respectively (Fig. 1b). In mammals, a β1,4-galactose, combined with a sialic acid, is often attached to GlcNAc residue, which is catalyzed by β1,4 galactosyltransferase and α2,6 sialyltransferase sequentially. Also, tri- and tetra-antennary branched complex N-glycans are common extensions in mammals, which are facilitated by GnTIII, GnTIV and GnTV, respectively (Fig. 1c).

Typical structure of N-glycans of fungal (a), plant (b) and animal (c) proteins. The depiction is adapted from Gomord et al. and Castilho et al.19,65. The N-glycans are attached to contiguous asparagine residues with the consensus sequence Asn-X-Ser/Thr/Cys. Labels in red are enzymes for the steps involved in lineage-specific N-glycan modifications. Glycan residue representation for icons is shown at bottom.
Evolutionary origins of N-glycosylation occurring in ER have been demonstrated to be conserved among eukaryotic lineages21. We believe that diversity and novelties of PNG, and hence structures of N-glycans of proteins in different domains of eukaryotic lives, should be reflected by gene novelties coding for enzymes in the PNG pathways in Golgi. However, our knowledge of origin and evolution of PNG reactions in Golgi apparatus is dispatched. In this article, we aim to systematically investigate how molecular mechanisms of PNG in Golgi emerged and evolved, based on which we propose how they shaped diversity and novelty of protein N-glycans in different eukaryotic lineages. On the basis of identification of conserved domains in the PNG genes already characterized, we systematically identified genes containing the domains through combination of BLAST and HMMER, facilitated by whole genome sequencing and assembly data available. Based on the gene sequences retrieved, we sought to answer when novel mechanisms of PNG possibly occurred, and how they evolved in fungi, animals and plants.
Results
Reference sequence collection and domain identification
Reference PNG gene sequences with known enzymatic functions were retrieved based on recent articles10,12,16,19,22. In total, 11 sequences from Saccharomyces cerevisiae, 8 from Arabidopsis thaliana, and 5 from Homo sapiens were collected (Table 1, Fig. 1, Supplementary Table S1). Of the 24 genes, all encode glycosyltranseferases, except for two mannosidases (MNS1 and GMII). 23 Pfam domains were identified in the peptide sequences of the 24 genes. Among them, combination of domains were identified in Och1, Van1, α1,4-FucT, and GnTV, respectively, while several domains were shared by multiple (2–3) enzymes (Table 1).
Interestingly, 9 domains, which were embodied in sequences encoding glycosyltransferases, belonged to the same clan (Pfam ID: CL0110) (Table 1). Based on Pfam definition, a clan contains multiple Pfam families that have descendent from a single evolutionary origin23. In the Pfam database, the clan CL0110 contains 46 families of glycosyltransferases possessing a Rossmann-like fold structure24. In total, domains in 15 out of 22 glycosyltransferase genes in PNG could be classified into the clan CL0110, suggesting that these genes have risen from a single evolutionary origin23. Notably, all domains in genes responsible for fungus-specific modifications belong to this clan, suggesting that the metabolic pathway leading to the branched structures with dense mannosylation took shape by duplication and divergence of a single sequence evolutionarily in fungi (Fig. 1, Table 1).
Identification of PNG genes by domain recognition
Based on the reference peptide sequences and the identified domains, we sought a comprehensive identification from selected sequenced genomes. We chose species of representative taxa in the tree of life with high-quality genome assembly data publicly available. Peptide gene model files of 15 Archaea and 52 Bacteria were downloaded, whose genomes are completely assembled, and those of 24 fungal species and 34 animal species were obtained, whose chromosome-level genome assembly data are available. Gene model files of 28 plant species were downloaded, among which genomes of 21 species are assembled to chromosome-level, and 7 are assembled to scaffold-level whose taxa represent algae, lower vascular plants, and Amborella which is close to the base of the flowering plant lineage. Hence, in total, we used 153 genomes for identification of PNG domain-containing genes (Supplementary Table S2).
Based on the identified domains in reference genes of PNG enzymes, we used HMMER, a domain-centric method to compare profile hidden Markov models (HMMs) of PfamA to peptide datasets, to identify homologous sequences, by which 4491 sequences were obtained in total25,26. BLAST searches were performed too, which did not generate any sequences beyond HMMER search results. 6 domains or domain combinations as contained in PNG sequences are confined in a specific eukaryotic lineage; this distribution is consistent with that of genes containing these domains related to PNG (Fig. 2, Supplementary Figure S1). For example, Mnn9, Van1 and Anp1, which are involved in protein N-mannosylation in fungi, are supposed to be present in fungi; all these genes contain the same domain PF03452, and genes with this domain were only identified in fungi18,27,28. Likewise, activity of β1,4-GalT was only identified in animals, and the peptide sequence contains domains PF13733 and PF02709; genes containing both the domains were only identified in animals29,30. Although some genes are only present in specific lineages, sequences containing the domains in these genes were identified in other lineages too. For example, β1,2-XylT is only present in plants, which has the domain PF04577, but genes containing this domain were identified in animals as well as in plants (Fig. 2)31. Generally, if the genes containing the domains are present in kingdoms where a specific PNG reaction is not supposed to occur, genes would be remarkably more abundant in kingdoms where genes responsible for specific PNG mechanisms are present, except domains like PF05637 and PF00852 (Fig. 2).

The heat map depicts values of 10 of the logarithm of absolute gene number counts for each genome. Black represents no sequences identified for the gene in the genome. Brown dot below the Pfam domain IDs indicates that the domains are classified under the clan CL0110. A, PF01074; B, PF09261; C, PF07748; D, PF00337; E, PF01762; F, PF13733; G, PF02709; H, PF15027; I, PF15024. (a) Genes containing the domains related to PNG are only present in fungi; (b) genes related to PNG are only in plants and animals; (c) only in plants; (d) only in animals.
Gene evolution in fungal N-mannosylation
Fungal glycosylation is characterized by dense mannosylation which mainly takes place in Golgi apparatus. 11 enzymes were characterized to be involved in fungal Golgi N-mannosylation. Peptide sequences of these enzymes were classified into 5 pfam families, and could be further assigned to the same clan, CL0110, which indicates that the glycosylation enzymes in sophisticated fungal PNG mechanism could be traced back to a single source evolutionarily, which probably occur by gene duplication and divergence.
While genes containing PF03452 and PF01793 were only identified in fungi, PF05637 and PF11051 were identified in plants as well, and PF04488 were identified in plants and animals as well as in fungi.
Mnn9 and Van1 make up mannan polymerase I (Man Pol-I), and Anp1 is a component of mannan polymerase II (Man Pol-II)18,27,28. All these 3 enzymes belong to the family PF03452. Interestingly, PF03452 genes were identified only in Saccharomyceta; over 3 copies of genes were present in each genome of these species. In phylogeny, these genes were classified into 3 major clades (Supplementary Figure S1). Genes in Clade II were split into 2 minor groups in Saccharomucetales (Clades IIa and IIb, Supplementary Figure S1). Only leotiomyceta genes were identified in Clade III. Mnn6 contain domain PF01793, which was clustered in Clade I of the 3 major clades of the phylogeny tree of PF01793, as shown in Supplementary Figure S2.
Both Mnn10 and Mnn11 contain domain PF05637. These genes were shown to be present not only in fungi but in plants. In E. cuniculi and Basidiomycota representing basal fungi clades, no homologs were identified. The phylogeny of this family resolved multiple distinctive clades, which indicated that the members have evolved into different biological/enzymatic roles. Mnn10 and Mnn11 represented the only 2 members of PF05637 in S. cerevisiae, which are in Clade I and IV, respectively. Only plant members are represented in Clade II. In Clade III, only leotiomyceta homologs were included, while in Clade V, there are leotiomyceta and plant homologs (Supplementary Figure S3). Indeed, the plant homologs have shown to not play roles in glycoprotein biosynthesis, but are involved in plant cell wall biosynthesis32,33.
6 S. cerevisiae genes contain domain PF11051. Among them, MNN2 (YBR015C) and MNN5 (YJL186W) are specifically involved in N-glycan formation, which are responsible for the addition of the first and second α1,2-linked mannose, respectively, to form the branches on the mannan backbone of oligosaccharides34. MNN1 are involved in both N- and O-glycosylation, while the other 3 are specifically involved in O-glycosylation35,36. The phylogeny partitioned the PF11051 members into 2 groups largely, in which MNN2 and MNN5 are placed in the same group (Clade I), and the other 4 S. cerevisiae genes are in the other group (Clade II) (Supplementary Figure S4). An Arabidopsis gene was identified, which was grouped in the clade II, whose biological function has not been reported yet, to our knowledge (Supplementary Figure S4).
Och1 (YGL038C) initiates N-mannosylation in Golgi by attaching an α1,6 mannose unit to the oligosaccharide core37. Hoc1 (YJR075W) is a component of M-PolII which adds α1,6 mannose residues to the core38,39. Both peptides were identified to contain PF04488 domain. Besides these genes, 2 more were identified from S. cerevisiae, which have been characterized to play as Mannosylinositol phosphorylceramide (MIPC) synthase catalytic subunits, and be involved in sphingolipid biosynthesis40; these genes form a single clade in phylogeny, in which plant and animal genes are included too (Clade II) (Supplementary Figure S5).
Gene Evolution of PNG enzymes shared by plants and animals
In Golgi apparatus, several α1,2-linked mannose residues need to be removed to provide the Man5GlcNAc2 substrate for the formation of complex N-glycans in animals and plants. In human, this reaction is catalyzed by 3 isoforms of Golgi-α-mannosidase41. In Arabidopsis, 2 isoforms of this enzyme are present in the genome42,43. These enzymes belong to class I α-mannosidases, which harbor a conserved domain PF01532. A thorough retrieval of sequences containing PF01532 was conducted, and the results indicated that genes in this family are present not only in plants and animals, but also in fungi. Phylogeny inference, however, indicated that plant and animal genes encoding Golgi-α-mannosidases were in 2 clusters close to each other (Supplementary Figure S6, Clades I and II). In these clusters, no fungi genes were contained. The clade close to Clades I and II comprises of fungal genes as well as animal and plant genes (Supplementary Figure S6, Clade III). In this clade, α-mannosidases from yeast (YJR131W), Arabidopsis (AT1G30000) and human (ENSG00000177239.14) are included, which all have been demonstrated to reside in ER and involved in ER-associated degradation (ERAD) of misfolded glycoproteins44,45,46,47. This indicates genes in the clade close to Golgi-α-mannosidase clade encode ER-associated α-mannosidases involved in protein quality control. Clade IV only contains fungal genes; in this clade, a yeast gene encoding mannosidase (YLR057W) is comprised, which was proved to be a novel component of ERAD pathway. Clade V is distant to any other clades, comprises genes in all three kingdoms (Supplementary Figure S6). In this clade, three human genes encode enzymes playing roles as ER degradation enhancers (EDEMs, ENSG00000134109, ENSG00000088298 and ENSG00000116406)48,49,50. Also, the yeast and Arabidopsis genes are involved in ERAD46,51. These results indicate that genes in all the clades are involved in ERAD, except genes in Clades I and II encoding Golgi-α-mannosidases, which only contain genes from animals and plants.
In animals and plants, β1-2-GlcNAc by GlcNAc transferase I (GnTI) starts the diversification of Man5GlcNAc252. In both Arabidopsis and human, a single gene is responsible for this role (AT4G38240 and ENSG00000131446, respectively)53,54,55. All these genes contain the domain PF03071. Genes were only identified in animals and plants. Phylogenetic analyses resolved four major clades. Plant genes are all in a single clade (Supplementary Figure S7, Clade I), and Chordata genes containing human GnTI gene (ENSG00000131446) was in another single clade (Supplementary Figure S7, Clade II). 3 C. elegans genes encoding GnTI were in an independent clade (Supplementary Figure S7, Clade III)56,57. The Clade IV, as shown in Supplementary Figure S7, comprising Chordata genes, contains a human gene encoding protein O-linked mannose N-acetylglucosaminyltransferase (ENSG00000085998); alterations of this gene have been shown to cause muscle-eye-brain disease and several congenital muscular dystrophies58,59.
Following the addition of a β1,2-GlcNAc by GnTI, α1,3- and α1,6-Man were removed from the core N-glycan substrate by α-mannosidase II (GMII)10. In human genome, 2 genes code for this enzyme60,61. Peptide sequences of these genes contain 3 domains: PF01074, PF09261 and PF07748. Genes harboring all the domains are identified in fungi as well as in animals and plants. However, as shown in phylogenetic tree, only plant and animal genes were present in a major clade, in which no genes were identified in fungi, indicating that genes encoding GMIIs are not present in fungi (Supplementary Figure S8, Clade I). The clade close to the group GMII contains genes encoding vacuolar α-mannosidases (YGL156w in yeast, and ENSG00000140400 in human, as shown in Supplementary Figure S8, Clade II)62,63. In another major clade, genes were only present in plants and animals, which likely encode α-mannosidases hydrolyzing terminal non-reducing α-D-mannose residues64,65.
Golgi β1,2-N-acetylglucosaminyltransferase II (GnTII) catalyzes the conversion from hybrid to complex N-glycans10. Peptide sequences of this gene contain a domain PF05060, and sequences of this enzyme were only identified in plants and animals; in each species, only 1–2 copies of genes were present in the genome.
Evolution of genes encoding PNG machinery specific for animals
β-1,4-galactosyltransferases (β1,4-GalT) form a family with seven members, which all have exclusive specificity for the donor substrate UDP-Gal, and all transfer Gal in β-1,4 linkage to GlcNac, Glc and Xyl29. One of them, β1,4-GalT I, catalyzes attachment of β1,4-galactose to GlcNAc residue, which is absent in plants and fungi30,66. In humans, this enzyme is encoded by ENSG0000008606267,68. The peptide sequence of this enzyme contains 2 Pfam domains (Domain IDs: PF13733 and PF02709). Genes containing these domains were identified in animals, while no genes were identified in fungi and plants. In phylogeny, the genes closest to the clade containing ENSG00000086062 likely encoded β1,4-GalT II which synthesizes N-acetyllactosamine in glycolipids and glycoproteins (Supplementary Figure S9)69,70.
Another animal-specific glycosyltransferase is α-2,6 sialyltransferase (α2,6-SialT), which catalyzes the transfer of sialic acid residue to terminal nonreducing positions of oligosaccharide chains of glycoproteins71. In humans, this peptide is encoded by gene ENSG00000117069, which is a Type II membrane protein and belongs to a family with multiple members71,72,73. Every known peptide sequence of these proteins was identified to harbor domain PF00777. Phylogenetic analyses of the sequences containing this domain showed that animal α2,6-SialT genes formed a monophyletic group (Supplementary Figure S10). Interestingly, plant homologs were present, which were placed close to the animal α2,6-SialT genes in the phylogenetic tree (Supplementary Figure S10). In plants, no sialic acid has been detected74. 2 Arabidopsis SiaT-like genes were suggested to be involved in transfer of 2-keto-3-deoxylyxo-heptulosaric acid and 2-keto-3-deoxymanno-octulosonic acid to Rhamnogalacturonan-II in pectic polymer biosynthesis and to be required for proper pollen tube elongation75.
Formation of tri- and tetra-antenary complex N-glycans are common in mammalian glycoprotein modification, while plant and fungal glycoproteins lack these multiantenary glycans10,76. These branched structures are associated with various physiological processes such as cancer metastasis and T-cell activation, and the glycans influence protein propertities including immunogenicity, stability and pharmacokinetics20,77,78. Branching of these N-glycans are catalyzed by several acetylglucosaminyltransferases (GnTIII, GnTIV and GnTV, respectively)79. GnTIII catalyzes the addition of N-acetylglucosamine in β1-4 linkage to the β-linked mannose of the trimannosyl core of N-linked sugar chains to produce a bisecting GlcNAc residue80. Domain PF04724 was identified in human GnTIII peptide sequences. Genes containing this domain were identified in animals as well as in plants and fungi; in most of the animal genomes, only 1–2 copies of the genes were present, while multiple copies of the genes were identified in every genome of the plants we investigated. Phylogenetic topology of animal genes in this family largely conformed with animal taxonomy (Supplementary Figure S11). In the phylogenetic tree, plant homologs were clustered together, none of which have been functionally characterized, to our knowledge. However, reports suggested that some genes were possibly involved in pollen germination and pollen tube development (Supplementary Figure S11)81. GnTIV catalyzes the transfer of GlcNAc from UDP-GlcNAc in β1-4 linkage to α1,3-D-mannoside on GlcNAcβ1-2Manα1-6(GlcNAcβ1-2Manα1-3)Manβ1-4 GlcNAcβ1-4GlcNAcβ1-Asn82,83. The peptide sequences of this gene were identified to contain domain PF04666. 4 homologs of GnTIV were present in human genome84,85. No genes were identified in land plants, while the genes were widely present in animal genomes. Phylogenetic inference showed that the four GnTIV genes were clustered into two major clades, with GnTIVA and GnTIVB, the two function-characterized genes clustered in Clade I as shown in Supplementary Figure S12. GnTV transfers N-acetylglucosamine (GlcNAc) to the C-6 position of the α1,6-linked mannosyl residue in the trimannosyl core structure of complex N-glycans to generate GlcNAc(α1,6) mannose86. 2 isoforms of GnTV gene are present in the human genome87,88. Two domains, namely PF05027 and PF15024, were identified in peptide sequences of GnTV. Genes harboring PF15027 were only identified in animals, while genes harboring PF15024 were identified in some dicotyledonous plants, ferns, algae as well as in animals. These domains could not be classified into any clans, and no reports indicate genes containing these domains in other eukaryotic kingdoms have any other activities, thus allowing us to conclude that this gene emerged de novo. Phylogeny of genes with the domain PF15024 showed that the animal genes were mostly split into 2 clades, with most species we studied in this article represented in each clade. This shows that animal GnTV experienced duplications in early stage of animal evolution, with genes in both clades retained in animal genomes (Supplementary Figure S13). In the phylogenetic tree, plant genes harboring domain PF15024 were present in a monophyletic clade; the plant genes have lost the domain PF15027, suggesting loss of GnTV activities with these genes (Supplementary Figure S13).
Evolution of PNG Genes specific for plants
In plants, complex-type N-glycans are structurally unique. β-mannose of the glycan core is attached by a bisecting β1,2-xylose, and proximal N-acetylglucosamine of the glycan core is substituted by an α1,3-fucose. Also, β1,3 galactose and α1,4-fucose link to the terminal GlcNAc of N-glycans, which form the Lewis a (Lea) oligosaccharide structure. These unique characteristics of the N-glycan structure are believed to be conferred by the plant-specific components of the enzymatic machinery in plants. Progresses have been made to identify the genes responsible for the formation of these structures in model plant Arabidopsis31. However, few reports were available on the genes in these steps in other plant species, whose origins and evolutions are still unknown, to our knowledge.
β1,2-xylosyltransferase (XylT) catalyzes the transfer of xylose to the N-glycans in glycoproteins in plants. Only the gene in Arabidopsis was enzymatically identified to be XylT in vitro and in vivo, despite some reports of enzyme purification of XylTs from other plants31,89,90. Domain PF04577 was identified in Arabidopsis XylT peptide sequence (AT5G55500). Genes containing the domain PF04577 were not identified in fungi. Only 1–3 genes were present in each animal genome, while in plants, genes are abundant (as many as over 30 copies in each genome). Phylogenetic analyses resolved the gene family into 3 major groups, and Group I could be further split to 3 clades, namely Clades Ia, Ib and Ic, respectively (Supplementary Figure S14). Arabidopsis XylT gene was placed under the Clade Ib in Group I; in this clade, only plant genes were present, indicating that genes in this clade represent XylT genes in plants. Clade Ia contains animal and plant genes; in this clade, no plant genes have been enzymatically or physiologically characterized, but the human gene ENSG00000144647 was identified to encode a protein O-linked mannose N-acetylglucosaminyltransferase, suggesting roles of the genes in this clade involved in protein O-glycans91,92. In Clade Ic, animal genes are grouped together, among which human gene ensg00000163378 was identified as a EGF domain specific O-linked N-acetylglucosamine transferase93. Genes in Groups II and III are all derived from plant genomes: Group II genes consist of Klebsormidium flaccidum genes, while genes in Group III consist of genes from species spanning from lower to higher land plants (Supplementary Figure S14). In this group, although no genes were definitely identified enzymatically, AT3G10320 was demonstrated as a putative xylosyltransferase which was recently characterized as MUCI21, while the genes AT3G18170 and AT3G18180 are expressed highly in a heteroxylan containing mucilaginous tissues, which indicated that the genes in this group are related to mucilage production in terrestrial plants94,95.
α1,3-fucose transferase (α1,3-FucT) and α1,4-fucose transferase (α1,4-FucT) add fucose residue to the basal and terminal part of the glycan core, respectively. In Arabidopsis, the genes encoding these enzymes were identified (AT3G19280 and AT1G49710 for α1,3-FucT, and AT1G71990 for α1,4-FucT)96,97. The Arabidopsis genes encoding both the enzymes conferred domain PF00852. Genes containing this domain were identified in both plants and animals, but not in other eukaryotic species. Copy numbers in animals are slightly more than those in plants. Phylogenetic analysis results showed that the plant sequences were clustered into two groups, and each contained one of Arabidopsis fucose transferases, respectively, indicating that the genes in these two groups represent genes encoding α1,3-FucT and α1,4-FucT, respectively, which play roles in plant-specific N-glycan modifications (Supplementary Figure S15, Clades I and II). In animal-specific Clades III, IV and V, genes likely code for fucoses too, whose substrates include polysaccharides and sphingolipids, and mutations in these genes have been demonstrated to be associated with a variety of human diseases98,99,100,101,102.
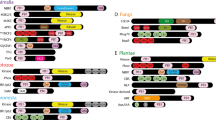
The other component of Lea structure, β1,3-galactose, is attached by β-1,3 galactosyltransferase (β1,3-GalT). In Arabidopsis, this enzyme is encoded by AT1G26810103. The peptide sequence of this gene was identified to contain 2 Pfam domains, PF01762 and PF00337 (Fig. 3a). The sequences containing both these domains were only identified in land plants. There were 6 β1,3-GalT genes in Arabidopsis, consistent with Strasser et al.103. Phylogenetic analysis indicated that the genes split into 2 groups shortly after the origination of the genes containing the 2 domains (Fig. 3b). Strasser et al. posited that only 1 out of the 6 genes has β1,3-galactose activity103. In the phylogeny tree, this gene was clustered in Clade I, forming a monophyletic clade (Fig. 3b). Interestingly, both the domains PF01762 and PF00337 are present in animal genomes, but they were contained in separated genes (Fig. 3d; Supplementary Figure S16). The animal genes harboring the domain PF01762 include type II membrane-bound glycoproteins with diverse enzymatic functions which use different donor substrates including UDP-galactose and UDP-N-acetylglucosamine, and different acceptor sugars such as N-acetylglucosamine, galactose and N-acetylgalactosamine (Supplementary Figure S16)104. Genes containing the PF00337 domain are a gene family coding for β-galactoside-binding proteins, which are implicated in modulating cell-cell and cell-matrix interactions105,106. In algae, genes were identified containing only domain PF01762. In land plants, a group of genes were identified containing two domains, PF01762 and PF13334; PF13334 emerged in land plants, and were not identified in any other eukaryotic lineages. This group of genes contained O-galactosyltransferases involved in cell wall formation and embryo development (Supplementary Figure S16)107,108. Our results indicated that the β-1,3 galactosyltransferase genes, which are essential in construction of Lea structure in land plants, originated by combination of two domains, PF01762 and PF00337, followed by sequence duplication and divergence of new genes.

(a) Linear structure of Arabidopsis β1,3-GalT (AT1G26810), highlighting the two domains it contains. (b) Unrooted tipless phylogenetic tree of plant genes containing both domains PF01762 and PF00337, which shows that genes could be classified into two major groups. Group I is shaded in red, which includes β1,3-GalT, and the Group II is shaded in grey. (c) Phylogenetic tree with tip labels of plant genes containing both domains PF01762 and PF00337, in which the known Arabidopsis β1,3-GalT gene (AT1G26810.1, labeled in red) is placed in group I. (d) Abundance of genes containing domains PF01762, PF00337 and PF13334, respectively. The heat map depicts values of 10 of the logarithm of absolute gene number counts for each genome. Black shade represents no gene identified for the gene in a specific genome.
Discussion
Structural diversity of protein N-glycans is believed to be attributable to differences of protein N-glycosylation (PNG) mechanisms, which largely is confined to Golgi apparatus, among eukaryotic lineages. A recent work has been performed to characterize origin and early evolution of PNG machinery in ER, which is largely consistent among fungi, plants and animals21. Although extensive studies have been made to characterize genes constituting unique PNG machinery components, little is known of how the genes expressed in Golgi apparatus emerged and evolved in eukaryotes. With the availability of high-quality genome assembly data, we carried out a comprehensive identification of PNG genes in Golgi, and studied evolution of the genes with the emphasis on evolution of domains25. A domain is a conserved region which is more conserved than other regions in a gene. We believe that domain-centric approaches could result in much more comprehensive identification of interested genes, as domains are conserved regions which are much more slowly diverged in sequence of evolution in the tree of life. Homology-based approaches like BLAST rely heavily on parameters such as e-value, which may introduce subjectivity. Hence, besides identification of homologs of PNG genes using BLAST, we carried out comprehensive survey of genes containing the domains that the PNG genes harbor in genomes of high quality which represent major species from archaea, bacteria, fungi, animals and plants through HMMER. In total, we obtained 4491 genes from 153 genomes, of which most were identified in eukaryotes. Further, we conducted phylogenetic analyses, together with extensive literature investigation, to help us to infer fucntions of the genes. This way, we believe it offers basis to provide fuller picture of eukaryotic PNG machinery evolution than by only retrieve of homologs of known genes.
24 domains were identified in the 23 known PNG genes. In yeast, 11 known PNG genes contained 5 conserved domains, which could be further classified into the same clan, CL0110. Also, GnTI and GnTII, the two glycosyltransferases shared by plants and animals, together with β1,3-GalT in plants and β1,4-GalT in animals could be classified into the clan CL0110, too. In total, 17 out of the 22 glycosyltransferase genes in PNG could be classified into the same clan. This clearly suggests single evolutionary source of the majority of genes constituting the complex and divergent PNG machinery. Although some genes, such as β1,2-XylT and α1,3-FucT in plants, and α2,6-SialT in animals, have kingdom-specific presence, genes containing domains that these genes contain could have genes identified in other kingdoms, indicating that emergence of these genes followed gene duplication and divergence model. Of all the 23 genes, only GnTV in animals, which harbors two domains, cannot be traced to any source, which indicates that this gene emerged de novo in animal lineage. Although less acknowledged, domain shuffling is an important way of new genes arising. In this study, β1,3-GalT is an example of domain shuffling, which takes an essential role in plant-specific Lea formation. Peptide sequence of β1,3-GalT contains two domains, PF00337 and PF01762. Genes containing PF01762 were identified in fungi and animals too, and PF00337 was also present in animals. In animals, the two domains were in separated genes, and only in plants the two domains were identified to be fused, probably through domain shuffling.
Overall, this is an example that shows component novelty, which shapes uniqueness of a pathway in a lineage, could be achieved by varied evolutionary mechanisms. In this article, we showed that PNG genes in Golgi evolved mainly by duplication and further fast divergence, but there are cases of de novo evolution and domain shuffling in shaping novelty of PNG pathways in animals and plants. We believe that the protein N-glycosylation pathway is still fast evolving, and there should be unknown PNG elements awaiting identification. Our work provides foundation for further characterization of PNG mechanisms in Eukaryotes, and the results may have important implications for our understanding of evolution of genetic novelties shaping uniqueness of PNG pathways in Eukaryotes.
Methods
Sequence and domain identification
Reference peptide sequences were retrieved from Saccharomyces Genome Database (S. cerevisiae), TAIR (A. thaliana) and Ensembl Genomes (H. sapiens), respectively109,110. Pfam domain IDs were retrieved using pfam_scan.pl and PfamA database111,112. Carbohydrate-active enzyme categorization for reference genes were fulfilled with sequence-based annotation tool using CAZy Database through both BLAST and HMMER approaches using default values113,114.
Gene model files of 15 Archaea, 52 Bacteria, 24 fungal, 34 animal and 28 plant genomes were obtained, sources of which are recorded in Supplementary Table S2. The files were cleaned to only contain locus IDs in comment line, and removed symbols other than Roman letters in sequence lines which would otherwise interfere with gene identification and sequence analyses. Standalone BLAST searches were performed against the gene model files, using BLASTp in BLAST + suite, using reference peptide sequences obtained involved in protein N-glycosylation as queries, with E-values 1e-3115. Also, HMM searches of the 153 gene model files against the 23 domains in genes related to protein N-glycosylation, plus PF13334, which is fused with PF01762, were conducted respectively, with the “trusted cutoff” of the domains established by Pfam-A (ftp://sanger.ac.uk/) as the threshold for detecting the domain. Combination of BLAST and HMMER search results resulted in raw data files for each domain. Searches against Pfam-A family database using perl script pfam_scan.pl using “trusted cutoff” as the threshold, deletion of repeated sequences with 100% homology and visual inspection resulted in final versions of sequence data, with 4491 sequences obtained in total.
Bioinformatic analyses
Sequences were analyzed by Probcons v1.12, and the alignments were visualized by BioEdit 7.2.5 ((http://www.mbio.ncsu.edu/)116. If needed, the alignment results were converted between from FASTA to NEXUS and/or PHYLIP formats. Bayesian phylogenetic analyses were conducted using MrBayes 3.1.2, with four Markov chains and two runs, with parameters set as default unless otherwise mentioned117. Standard deviation of split frequencies was checked after each 1,000,000 generations of each run to make sure they are below 0.05. The trees generated were visualized using Figtree v1.4.2 (http://tree.bio.ed.ac.uk/).
Gene structure was drawn using IBS 1.0.1, based on domain information retrieved from Pfam domain identification118. Gene number counts were converted to 10 of logarithmic values before heatmap illustration using pheatmap, an R package (https://cran.r-project.org/).
Nomenclature
“Seven-kingdom system” developed by Michael et al. was used for classification of living organisms, in which the division of empire Prokaryota was introduced into two kingdoms, Bacteria and Archaea, and the empire Eukaryota was divided into five kingdoms, Protozoa, Chromista, Plantae, Fungi and Animalia119. We focus on three eukaryotic kingdoms, fungi, animals (Animalia) and plants (Plantae).
Gene family whose member containing N-glycosylation domain was named after the domain ID. For example, the gene family containing a domain PF01532, which encode mannosidases, is named PF01532 family. Unless otherwise indicated, genes were named after locus IDs, which were prefixed by abbreviations of species names and double underlines. For example, Arabidopsis thaliana gene AT1G26810 was labeled as Ath__AT1G26810. Human gene nomenclature was based on HUGO Gene Nomenclature Committee (HGNC) Database120,121.
Additional Information
How to cite this article: Wang, P. et al. Evolution of protein N-glycosylation process in Golgi apparatus which shapes diversity of protein N-glycan structures in plants, animals and fungi. Sci. Rep. 7, 40301; doi: 10.1038/srep40301 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Tautz, D. & Domazet-Loso, T. The evolutionary origin of orphan genes. Nature reviews. Genetics 12, 692–702, doi: 10.1038/nrg3053 (2011).
Khalturin, K., Hemmrich, G., Fraune, S., Augustin, R. & Bosch, T. C. More than just orphans: are taxonomically-restricted genes important in evolution? Trends in genetics: TIG 25, 404–413, doi: 10.1016/j.tig.2009.07.006 (2009).
Siepel, A. Darwinian alchemy: Human genes from noncoding DNA. Genome research 19, 1693–1695, doi: 10.1101/gr.098376.109 (2009).
Domazet-Loso, T. & Tautz, D. An evolutionary analysis of orphan genes in Drosophila. Genome research 13, 2213–2219, doi: 10.1101/gr.1311003 (2003).
Neme, R. & Tautz, D. Phylogenetic patterns of emergence of new genes support a model of frequent de novo evolution. BMC genomics 14, 117, doi: 10.1186/1471-2164-14-117 (2013).
Nakamura, Y., Itoh, T. & Martin, W. Rate and polarity of gene fusion and fission in Oryza sativa and Arabidopsis thaliana. Molecular biology and evolution 24, 110–121, doi: 10.1093/molbev/msl138 (2007).
Moore, A. D. & Bornberg-Bauer, E. The dynamics and evolutionary potential of domain loss and emergence. Molecular biology and evolution 29, 787–796, doi: 10.1093/molbev/msr250 (2012).
de Souza, S. J. Domain shuffling and the increasing complexity of biological networks. BioEssays: news and reviews in molecular, cellular and developmental biology 34, 655–657, doi: 10.1002/bies.201200006 (2012).
Kawashima, T. et al. Domain shuffling and the evolution of vertebrates. Genome research 19, 1393–1403, doi: 10.1101/gr.087072.108 (2009).
Costa, A. R., Rodrigues, M. E., Henriques, M., Oliveira, R. & Azeredo, J. Glycosylation: impact, control and improvement during therapeutic protein production. Critical reviews in biotechnology 34, 281–299, doi: 10.3109/07388551.2013.793649 (2014).
Xu, C. & Ng, D. T. Glycosylation-directed quality control of protein folding. Nature reviews. Molecular cell biology 16, 742–752, doi: 10.1038/nrm4073 (2015).
Lannoo, N. & Van Damme, E. J. N-glycans: The making of a varied toolbox. Plant science: an international journal of experimental plant biology 239, 67–83, doi: 10.1016/j.plantsci.2015.06.023 (2015).
Matsui, T. et al. N-glycosylation at noncanonical Asn-X-Cys sequences in plant cells. Glycobiology 21, 994–999, doi: 10.1093/glycob/cwq198 (2011).
Sato, C. et al. Characterization of the N-oligosaccharides attached to the atypical Asn-X-Cys sequence of recombinant human epidermal growth factor receptor. Journal of biochemistry 127, 65–72 (2000).
Chi, Y. H. et al. N-glycosylation at non-canonical Asn-X-Cys sequence of an insect recombinant cathepsin B-like counter-defense protein. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology 156, 40–47, doi: 10.1016/j.cbpb.2010.01.017 (2010).
Kellokumpu, S., Hassinen, A. & Glumoff, T. Glycosyltransferase complexes in eukaryotes: long-known, prevalent but still unrecognized. Cellular and molecular life sciences: CMLS, doi: 10.1007/s00018-015-2066-0 (2015).
Nakayama, K., Nagasu, T., Shimma, Y., Kuromitsu, J. & Jigami, Y. OCH1 encodes a novel membrane bound mannosyltransferase: outer chain elongation of asparagine-linked oligosaccharides. The EMBO journal 11, 2511–2519 (1992).
Stolz, J. & Munro, S. The components of the Saccharomyces cerevisiae mannosyltransferase complex M-Pol I have distinct functions in mannan synthesis. The Journal of biological chemistry 277, 44801–44808, doi: 10.1074/jbc.M208023200 (2002).
Gomord, V. et al. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant biotechnology journal 8, 564–587, doi: 10.1111/j.1467-7652.2009.00497.x (2010).
Zhao, Y. et al. Branched N-glycans regulate the biological functions of integrins and cadherins. The FEBS journal 275, 1939–1948, doi: 10.1111/j.1742-4658.2008.06346.x (2008).
Lombard, J. The multiple evolutionary origins of the eukaryotic N-glycosylation pathway. Biology direct 11, 36, doi: 10.1186/s13062-016-0137-2 (2016).
Fabre, E., Hurtaux, T. & Fradin, C. Mannosylation of fungal glycoconjugates in the Golgi apparatus. Current opinion in microbiology 20, 103–110, doi: 10.1016/j.mib.2014.05.008 (2014).
Finn, R. D. et al. Pfam: clans, web tools and services. Nucleic acids research 34, D247–251, doi: 10.1093/nar/gkj149 (2006).
Liu, J. & Mushegian, A. Three monophyletic superfamilies account for the majority of the known glycosyltransferases. Protein science: a publication of the Protein Society 12, 1418–1431, doi: 10.1110/ps.0302103 (2003).
Mistry, J. & Finn, R. Pfam: a domain-centric method for analyzing proteins and proteomes. Methods Mol Biol 396, 43–58, doi: 10.1007/978-1-59745-515-2_4 (2007).
Krogh, A., Brown, M., Mian, I. S., Sjolander, K. & Haussler, D. Hidden Markov models in computational biology. Applications to protein modeling. Journal of molecular biology 235, 1501–1531, doi: 10.1006/jmbi.1994.1104 (1994).
Jungmann, J. & Munro, S. Multi-protein complexes in the cis Golgi of Saccharomyces cerevisiae with alpha-1,6-mannosyltransferase activity. The EMBO journal 17, 423–434, doi: 10.1093/emboj/17.2.423 (1998).
Chapman, R. E. & Munro, S. The functioning of the yeast Golgi apparatus requires an ER protein encoded by ANP1, a member of a new family of genes affecting the secretory pathway. The EMBO journal 13, 4896–4907 (1994).
Amado, M., Almeida, R., Schwientek, T. & Clausen, H. Identification and characterization of large galactosyltransferase gene families: galactosyltransferases for all functions. Biochimica et biophysica acta 1473, 35–53 (1999).
Bakker, H. et al. Galactose-extended glycans of antibodies produced by transgenic plants. Proc Natl Acad Sci USA 98, 2899–2904, doi: 10.1073/pnas.031419998 (2001).
Strasser, R. et al. Molecular cloning and functional expression of beta1, 2-xylosyltransferase cDNA from Arabidopsis thaliana. Febs Lett 472, 105–108 (2000).
Vuttipongchaikij, S. et al. Arabidopsis GT34 family contains five xyloglucan alpha-1,6-xylosyltransferases. New Phytol 195, 585–595, doi: 10.1111/j.1469-8137.2012.04196.x (2012).
Voiniciuc, C. et al. MUCILAGE-RELATED10 Produces Galactoglucomannan That Maintains Pectin and Cellulose Architecture in Arabidopsis Seed Mucilage. Plant Physiol 169, 403, doi: 10.1104/pp.15.00851 (2015).
Rayner, J. C. & Munro, S. Identification of the MNN2 and MNN5 mannosyltransferases required for forming and extending the mannose branches of the outer chain mannans of Saccharomyces cerevisiae. Journal of Biological Chemistry 273, 26836–26843, doi: 10.1074/jbc.273.41.26836 (1998).
Lussier, M., Sdicu, A. M. & Bussey, H. The KTR and MNN1 mannosyltransferase families of Saccharomyces cerevisiae. Bba-Gen Subjects 1426, 323–334, doi: 10.1016/S0304-4165(98)00133-0 (1999).
Romero, P. A. et al. Mnt2p and Mnt3p of Saccharomyces cerevisiae are members of the Mnn1p family of alpha-1,3-mannosyltransferases responsible for adding the terminal mannose residues of O-linked oligosaccharides. Glycobiology 9, 1045–1051, doi: 10.1093/glycob/9.10.1045 (1999).
Lehle, L., Eiden, A., Lehnert, K., Haselbeck, A. & Kopetzki, E. Glycoprotein-Biosynthesis in Saccharomyces-Cerevisiae - Ngd29, an N-Glycosylation Mutant Allelic to Och1 Having a Defect in the Initiation of Outer Chain Formation. Febs Lett 370, 41–45, doi: 10.1016/0014-5793(95)00789-C (1995).
Jungmann, J. & Munro, S. Multi-protein complexes in the cis Golgi of Saccharomyces cerevisiae with alpha-1,6-mannosyltransferase activity. Embo Journal 17, 423–434, doi: 10.1093/emboj/17.2.423 (1998).
Stolz, J. & Munro, S. The components of the Saccharomyces cerevisiae mannosyltransferase complex M-Pol I have distinct functions in mannan synthesis. Journal of Biological Chemistry 277, 44801–44808, doi: 10.1074/jbc.M208023200 (2002).
Uemura, S., Kihara, A., Inokuchi, J. & Igarashi, Y. Csg1p and newly identified Csh1p function in mannosylinositol phosphorylceramide synthesis by interacting with Csg2p. Journal of Biological Chemistry 278, 45049–45055, doi: 10.1074/jbc.M305498200 (2003).
Mast, S. W. & Moremen, K. W. In Glycobiology Vol. 415 Methods in Enzymology (ed. Fukuda, M. ) 31–46 (2006).
Liebminger, E. et al. Class I alpha-mannosidases are required for N-glycan processing and root development in Arabidopsis thaliana. The Plant cell 21, 3850–3867, doi: 10.1105/tpc.109.072363 (2009).
Kajiura, H. et al. Two Arabidopsis thaliana Golgi alpha-mannosidase I enzymes are responsible for plant N-glycan maturation. Glycobiology 20, 235–247, doi: 10.1093/glycob/cwp170 (2010).
Knop, M., Hauser, N. & Wolf, D. H. N-Glycosylation affects endoplasmic reticulum degradation of a mutated derivative of carboxypeptidase yscY in yeast. Yeast 12, 1229–1238, doi: 10.1002/(SICI)1097-0061(19960930)12:12<1229::AID-YEA15>3.0.CO;2-H (1996).
Camirand, A., Heysen, A., Grondin, B. & Herscovics, A. Glycoprotein biosynthesis in Saccharomyces cerevisiae. Isolation and characterization of the gene encoding a specific processing alpha-mannosidase. The Journal of biological chemistry 266, 15120–15127 (1991).
Huttner, S. et al. Arabidopsis Class I alpha-Mannosidases MNS4 and MNS5 Are Involved in Endoplasmic Reticulum-Associated Degradation of Misfolded Glycoproteins. The Plant cell 26, 1712–1728, doi: 10.1105/tpc.114.123216 (2014).
Benyair, R. et al. Mammalian ER mannosidase I resides in quality control vesicles, where it encounters its glycoprotein substrates. Molecular biology of the cell 26, 172–184, doi: 10.1091/mbc.E14-06-1152 (2015).
Guiliano, D. B. et al. Endoplasmic reticulum degradation-enhancing alpha-mannosidase-like protein 1 targets misfolded HLA-B27 dimers for endoplasmic reticulum-associated degradation. Arthritis Rheumatol 66, 2976–2988, doi: 10.1002/art.38809 (2014).
Ninagawa, S. et al. EDEM2 initiates mammalian glycoprotein ERAD by catalyzing the first mannose trimming step. The Journal of cell biology 206, 347–356, doi: 10.1083/jcb.201404075 (2014).
Hirao, K. et al. EDEM3, a soluble EDEM homolog, enhances glycoprotein endoplasmic reticulum-associated degradation and mannose trimming. The Journal of biological chemistry 281, 9650–9658, doi: 10.1074/jbc.M512191200 (2006).
Gauss, R., Kanehara, K., Carvalho, P., Ng, D. T. & Aebi, M. A complex of Pdi1p and the mannosidase Htm1p initiates clearance of unfolded glycoproteins from the endoplasmic reticulum. Molecular cell 42, 782–793, doi: 10.1016/j.molcel.2011.04.027 (2011).
Burke, J., Pettitt, J. M., Humphris, D. & Gleeson, P. A. Medial-Golgi retention of N-acetylglucosaminyltransferase I. Contribution from all domains of the enzyme. The Journal of biological chemistry 269, 12049–12059 (1994).
von Schaewen, A., Sturm, A., O’Neill, J. & Chrispeels, M. J. Isolation of a mutant Arabidopsis plant that lacks N-acetyl glucosaminyl transferase I and is unable to synthesize Golgi-modified complex N-linked glycans. Plant Physiol 102, 1109–1118, doi: 10.1104/pp.102.4.1109 (1993).
Strasser, R. et al. Molecular basis of N-acetylglucosaminyltransferase I deficiency in Arabidopsis thaliana plants lacking complex N-glycans. The Biochemical journal 387, 385–391, doi: 10.1042/BJ20041686 (2005).
Kumar, R., Yang, J., Larsen, R. D. & Stanley, P. Cloning and expression of N-acetylglucosaminyltransferase I, the medial Golgi transferase that initiates complex N-linked carbohydrate formation. Proceedings of the National Academy of Sciences of the United States of America 87, 9948–9952, doi: 10.1073/pnas.87.24.9948 (1990).
Zhu, S., Hanneman, A., Reinhold, V. N., Spence, A. M. & Schachter, H. Caenorhabditis elegans triple null mutant lacking UDP-N-acetyl-D-glucosamine:alpha-3-D-mannoside beta1,2-N-acetylglucosaminyltransferase I. The Biochemical journal 382, 995–1001, doi: 10.1042/BJ20040793 (2004).
Chen, S., Zhou, S., Sarkar, M., Spence, A. M. & Schachter, H. Expression of three Caenorhabditis elegans N-acetylglucosaminyltransferase I genes during development. The Journal of biological chemistry 274, 288–297 (1999).
Raducu, M., Baets, J., Fano, O., Van Coster, R. & Cruces, J. Promoter alteration causes transcriptional repression of the POMGNT1 gene in limb-girdle muscular dystrophy type 2O. European journal of human genetics: EJHG 20, 945–952, doi: 10.1038/ejhg.2012.40 (2012).
Vervoort, V. S. et al. POMGnT1 gene alterations in a family with neurological abnormalities. Annals of neurology 56, 143–148, doi: 10.1002/ana.20172 (2004).
Moremen, K. W., Touster, O. & Robbins, P. W. Novel purification of the catalytic domain of Golgi alpha-mannosidase II. Characterization and comparison with the intact enzyme. The Journal of biological chemistry 266, 16876–16885 (1991).
Misago, M. et al. Molecular cloning and expression of cDNAs encoding human alpha-mannosidase II and a previously unrecognized alpha-mannosidase IIx isozyme. Proc Natl Acad Sci USA 92, 11766–11770 (1995).
Yoshihisa, T. & Anraku, Y. A novel pathway of import of alpha-mannosidase, a marker enzyme of vacuolar membrane, in Saccharomyces cerevisiae. The Journal of biological chemistry 265, 22418–22425 (1990).
Wang, L. & Suzuki, T. Dual functions for cytosolic alpha-mannosidase (Man2C1): its down-regulation causes mitochondria-dependent apoptosis independently of its alpha-mannosidase activity. The Journal of biological chemistry 288, 11887–11896, doi: 10.1074/jbc.M112.425702 (2013).
Borgwardt, L. et al. Alpha-mannosidosis: correlation between phenotype, genotype and mutant MAN2B1 subcellular localisation. Orphanet Journal of Rare Diseases 10, doi: 10.1186/s13023-015-0286-x (2015).
Rojo, E., Zouhar, J., Carter, C., Kovaleva, V. & Raikhel, N. V. A unique mechanism for protein processing and degradation in Arabidopsis thaliana. Proc Natl Acad Sci USA 100, 7389–7394, doi: 10.1073/pnas.1230987100 (2003).
Jacobs, P. P. & Callewaert, N. N-glycosylation engineering of biopharmaceutical expression systems. Current molecular medicine 9, 774–800 (2009).
Masri, K. A., Appert, H. E. & Fukuda, M. N. Identification of the full-length coding sequence for human galactosyltransferase (beta-N-acetylglucosaminide: beta 1,4-galactosyltransferase). Biochemical and biophysical research communications 157, 657–663 (1988).
Liu, W. et al. Elevated expression of beta1,4-galactosyltransferase-I in cartilage and synovial tissue of patients with osteoarthritis. Inflammation 35, 647–655, doi: 10.1007/s10753-011-9357-x (2012).
Sasaki, N. et al. beta4GalT-II is a key regulator of glycosylation of the proteins involved in neuronal development. Biochemical and biophysical research communications 333, 131–137, doi: 10.1016/j.bbrc.2005.05.082 (2005).
Jiang, J. et al. beta4GalT-II increases cisplatin-induced apoptosis in HeLa cells depending on its Golgi localization. Biochemical and biophysical research communications 358, 41–46, doi: 10.1016/j.bbrc.2007.04.044 (2007).
Harduin-Lepers, A., Mollicone, R., Delannoy, P. & Oriol, R. The animal sialyltransferases and sialyltransferase-related genes: a phylogenetic approach. Glycobiology 15, 805–817, doi: 10.1093/glycob/cwi063 (2005).
Wong, S. H., Low, S. H. & Hong, W. The 17-residue transmembrane domain of beta-galactoside alpha 2,6-sialyltransferase is sufficient for Golgi retention. The Journal of cell biology 117, 245–258 (1992).
Munro, S. Sequences within and adjacent to the transmembrane segment of alpha-2,6-sialyltransferase specify Golgi retention. The EMBO journal 10, 3577–3588 (1991).
Seveno, M. et al. Glycoprotein sialylation in plants? Nature biotechnology 22, 1351–1352 author reply 1352–1353, doi: 10.1038/nbt1104-1351 (2004).
Dumont, M. et al. The cell wall pectic polymer rhamnogalacturonan-II is required for proper pollen tube elongation: implications of a putative sialyltransferase-like protein. Annals of botany 114, 1177–1188, doi: 10.1093/aob/mcu093 (2014).
Pedrazzini, E. et al. The Arabidopsis tonoplast is almost devoid of glycoproteins with complex N-glycans, unlike the rat lysosomal membrane. Journal of Experimental Botany 67, 1769–1781, doi: 10.1093/jxb/erv567 (2016).
Demetriou, M., Granovsky, M., Quaggin, S. & Dennis, J. W. Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature 409, 733–739, doi: 10.1038/35055582 (2001).
Lau, K. S. & Dennis, J. W. N-Glycans in cancer progression. Glycobiology 18, 750–760, doi: 10.1093/glycob/cwn071 (2008).
Brockhausen, I., Narasimhan, S. & Schachter, H. The biosynthesis of highly branched N-glycans: studies on the sequential pathway and functional role of N-acetylglucosaminyltransferases I, II, III, IV, V and VI. Biochimie 70, 1521–1533 (1988).
Ihara, Y. et al. cDNA cloning, expression, and chromosomal localization of human N-acetylglucosaminyltransferase III (GnT-III). Journal of biochemistry 113, 692–698 (1993).
Wang, Y. et al. Transcriptome Analyses Show Changes in Gene Expression to Accompany Pollen Germination and Tube Growth in Arabidopsis. Plant Physiol 148, 1201–1211, doi: 10.1104/pp.108.126375 (2008).
Yoshida, A. et al. Tissue specific expression and chromosomal mapping of a human UDP-N-acetylglucosamine: alpha1,3-d-mannoside beta1, 4-N-acetylglucosaminyltransferase. Glycobiology 9, 303–310 (1999).
Zheng, Z. et al. Genetic variation in a4GnT in relation to Helicobacter pylori serology and gastric cancer risk. Helicobacter 14, 120–125, doi: 10.1111/j.1523-5378.2009.00708.x (2009).
Oguri, S., Yoshida, A., Minowa, M. T. & Takeuchi, M. Kinetic properties and substrate specificities of two recombinant human N-acetylglucosaminyltransferase-IV isozymes. Glycoconjugate journal 23, 473–480, doi: 10.1007/s10719-006-6216-3 (2006).
Yoshida, A. et al. A novel second isoenzyme of the human UDP-N-acetylglucosamine:alpha1,3-D-mannoside beta1,4-N-acetylglucosaminyltransferase family: cDNA cloning, expression, and chromosomal assignment. Glycoconjugate journal 15, 1115–1123 (1998).
Cummings, R. D., Trowbridge, I. S. & Kornfeld, S. A mouse lymphoma cell line resistant to the leukoagglutinating lectin from Phaseolus vulgaris is deficient in UDP-GlcNAc: alpha-D-mannoside beta 1,6 N-acetylglucosaminyltransferase. The Journal of biological chemistry 257, 13421–13427 (1982).
Kaneko, M. et al. A novel beta(1,6)-N-acetylglucosaminyltransferase V (GnT-VB)(1). Febs Lett 554, 515–519 (2003).
Saito, H. et al. cDNA cloning and chromosomal mapping of human N-acetylglucosaminyltransferase V+. Biochemical and biophysical research communications 198, 318–327 (1994).
Tezuka, K., Hayashi, M., Ishihara, H., Akazawa, T. & Takahashi, N. Studies on synthetic pathway of xylose-containing N-linked oligosaccharides deduced from substrate specificities of the processing enzymes in sycamore cells (Acer pseudoplatanus L.). European journal of biochemistry/FEBS 203, 401–413 (1992).
Zeng, Y. et al. Purification and specificity of beta1,2-xylosyltransferase, an enzyme that contributes to the allergenicity of some plant proteins. The Journal of biological chemistry 272, 31340–31347 (1997).
Ogawa, M. et al. GTDC2 modifies O-mannosylated alpha-dystroglycan in the endoplasmic reticulum to generate N-acetyl glucosamine epitopes reactive with CTD110.6 antibody. Biochemical and biophysical research communications 440, 88–93, doi: 10.1016/j.bbrc.2013.09.022 (2013).
Yoshida-Moriguchi, T. et al. SGK196 is a glycosylation-specific O-mannose kinase required for dystroglycan function. Science 341, 896–899, doi: 10.1126/science.1239951 (2013).
Ogawa, M., Furukawa, K. & Okajima, T. Extracellular O-linked beta-N-acetylglucosamine: Its biology and relationship to human disease. World journal of biological chemistry 5, 224–230, doi: 10.4331/wjbc.v5.i2.224 (2014).
Jensen, J. K., Johnson, N. & Wilkerson, C. G. Discovery of diversity in xylan biosynthetic genes by transcriptional profiling of a heteroxylan containing mucilaginous tissue. Frontiers in plant science 4, 183, doi: 10.3389/fpls.2013.00183 (2013).
Ralet, M. C. et al. The affinity of xylan branches on rhamnogalacturonan I for cellulose provides the structural driving force for mucilage adhesion to the Arabidopsis seed coat. Plant Physiol, doi: 10.1104/pp.16.00211 (2016).
Wilson, I. B. et al. Cloning and expression of cDNAs encoding alpha1,3-fucosyltransferase homologues from Arabidopsis thaliana. Biochimica et biophysica acta 1527, 88–96 (2001).
Both, P. et al. Distantly related plant and nematode core alpha1,3-fucosyltransferases display similar trends in structure-function relationships. Glycobiology 21, 1401–1415, doi: 10.1093/glycob/cwr056 (2011).
Mollicone, R. et al. Activity, splice variants, conserved peptide motifs, and phylogeny of two new alpha1,3-fucosyltransferase families (FUT10 and FUT11). The Journal of biological chemistry 284, 4723–4738, doi: 10.1074/jbc.M809312200 (2009).
Baboval, T. & Smith, F. I. Comparison of human and mouse Fuc-TX and Fuc-TXI genes, and expression studies in the mouse. Mammalian genome: official journal of the International Mammalian Genome Society 13, 538–541, doi: 10.1007/s00335-001-2152-5 (2002).
Padro, M., Cobler, L., Garrido, M. & de Bolos, C. Down-regulation of FUT3 and FUT5 by shRNA alters Lewis antigens expression and reduces the adhesion capacities of gastric cancer cells. Biochimica et biophysica acta 1810, 1141–1149, doi: 10.1016/j.bbagen.2011.09.011 (2011).
Toivonen, S., Nishihara, S., Narimatsu, H., Renkonen, O. & Renkonen, R. Fuc-TIX: a versatile alpha1,3-fucosyltransferase with a distinct acceptor- and site-specificity profile. Glycobiology 12, 361–368 (2002).
Li, W. et al. Alpha1,3 fucosyltransferase VII plays a role in colorectal carcinoma metastases by promoting the carbohydration of glycoprotein CD24. Oncology reports 23, 1609–1617 (2010).
Strasser, R. et al. A unique beta1,3-galactosyltransferase is indispensable for the biosynthesis of N-glycans containing Lewis a structures in Arabidopsis thaliana. The Plant cell 19, 2278–2292, doi: 10.1105/tpc.107.052985 (2007).
Amado, M. et al. A family of human beta3-galactosyltransferases. Characterization of four members of a UDP-galactose:beta-N-acetyl-glucosamine/beta-nacetyl-galactosamine beta-1,3-galactosyltransferase family. The Journal of biological chemistry 273, 12770–12778 (1998).
Elola, M. T., Blidner, A. G., Ferragut, F., Bracalente, C. & Rabinovich, G. A. Assembly, organization and regulation of cell-surface receptors by lectin-glycan complexes. The Biochemical journal 469, 1–16, doi: 10.1042/BJ20150461 (2015).
Barondes, S. H., Cooper, D. N., Gitt, M. A. & Leffler, H. Galectins. Structure and function of a large family of animal lectins. The Journal of biological chemistry 269, 20807–20810 (1994).
Ogawa-Ohnishi, M. & Matsubayashi, Y. Identification of three potent hydroxyproline O-galactosyltransferases in Arabidopsis. The Plant journal: for cell and molecular biology 81, 736–746, doi: 10.1111/tpj.12764 (2015).
Geshi, N. et al. A galactosyltransferase acting on arabinogalactan protein glycans is essential for embryo development in Arabidopsis. Plant Journal 76, 128–137, doi: 10.1111/tpj.12281 (2013).
Kersey, P. J. et al. Ensembl Genomes 2016: more genomes, more complexity. Nucleic acids research 44, D574–580, doi: 10.1093/nar/gkv1209 (2016).
Berardini, T. Z. et al. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 53, 474–485, doi: 10.1002/dvg.22877 (2015).
Finn, R. D. et al. The Pfam protein families database. Nucleic acids research 36, D281–288, doi: 10.1093/nar/gkm960 (2008).
Mistry, J., Bateman, A. & Finn, R. D. Predicting active site residue annotations in the Pfam database. BMC bioinformatics 8, 298, doi: 10.1186/1471-2105-8-298 (2007).
Park, B. H., Karpinets, T. V., Syed, M. H., Leuze, M. R. & Uberbacher, E. C. CAZymes Analysis Toolkit (CAT): web service for searching and analyzing carbohydrate-active enzymes in a newly sequenced organism using CAZy database. Glycobiology 20, 1574–1584, doi: 10.1093/glycob/cwq106 (2010).
Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P. M. & Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic acids research 42, D490–495, doi: 10.1093/nar/gkt1178 (2014).
Camacho, C. et al. BLAST+: architecture and applications. BMC bioinformatics 10, 421 (2008).
Do, C. B., Mahabhashyam, M. S., Brudno, M. & Batzoglou, S. ProbCons: Probabilistic consistency-based multiple sequence alignment. Genome research 15, 330–340, doi: 10.1101/gr.2821705 (2005).
Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574 (2003).
Liu, W. et al. IBS: an illustrator for the presentation and visualization of biological sequences. Bioinformatics 31, 3359–3361, doi: 10.1093/bioinformatics/btv362 (2015).
Ruggiero, M. A. et al. A Higher Level Classification of All Living Organisms. Plos One 10, doi: 10.1371/journal.pone.0119248 (2015).
Gray, K. A., Seal, R. L., Tweedie, S., Wright, M. W. & Bruford, E. A. A review of the new HGNC gene family resource. Human genomics 10, 6, doi: 10.1186/s40246-016-0062-6 (2016).
Gray, K. A., Yates, B., Seal, R. L., Wright, M. W. & Bruford, E. A. Genenames.org: the HGNC resources in 2015. Nucleic acids research 43, D1079–1085, doi: 10.1093/nar/gku1071 (2015).
Acknowledgements
We appreciate valuable advice from Professor Richard Strasser at University of Natural Resources and Life Sciences (BOKU) in Vienna, Austria. We are also grateful to S. Varney and N. Rachel for their English edting of the manuscript. We thank Computational Center of Xinjiang Institute of Ecology and Geography, Supercomputing Environment of Chinese Academy of Sciences for the uses of supercomputing resources. The work was supported by the Fundamental Scientific Research Funds for CATAS-TCGRI (1630032013009) and National Science Foundation of China (31300260).
Author information
Authors and Affiliations
Contributions
P.W. designed the study, analysed and interpreted the data; P.W. and H.W. wrote the manuscript; J.G., X.T., X.Z., Y.L. and Y.J. participated in the study designs and contributed in drafting and revising the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, P., Wang, H., Gai, J. et al. Evolution of protein N-glycosylation process in Golgi apparatus which shapes diversity of protein N-glycan structures in plants, animals and fungi. Sci Rep 7, 40301 (2017). https://doi.org/10.1038/srep40301
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40301
- Springer Nature Limited
This article is cited by
-
Overexpression of wheat α-mannosidase gene TaMP impairs salt tolerance in transgenic Brachypodium distachyon
Plant Cell Reports (2020)
-
Reconstruction of the sialylation pathway in the ancestor of eukaryotes
Scientific Reports (2018)