
Abstract
EV-D68 is associated with respiratory tract infections (RTIs). Since its first isolation, EV-D68 has been detected sporadically. However, the US and Canada have experienced outbreaks of EV-D68 infections between August and December 2014. This study aimed to investigate the molecular epidemiology and clinical characteristics of EV-D68 in Chongqing, Southwestern China. From January 2012 to November 2014, 1876 nasopharyngeal aspirate specimens (NPAs) were collected from hospitalized children with RTIs. Among the 1876 NPAs, EV-D68 was detected in 19 samples (1.0%, 19/1876). Of these, 13 samples were detected in September and October 2014 (9.8%, 13/132). Phylogenetic analysis showed that all 13 strains detected in the 2014 Chongqing had high homology with the main strains of the 2014 US outbreak. Among the children with EV-D68 infection, 13 (68%) had a history of recurrent wheezing. A total of 13 children had a discharge diagnosis of asthma. Of these, 11 children were diagnosed with acute asthma exacerbation. EV-D68 was the predominant pathogen that evoked asthma exacerbation in September and October 2014. In conclusion, our results found that a history of recurrent wheezing may be a risk factor for the detection of EV-D68 and viral-induced asthma exacerbation may be a clinical feature of EV-D68 infection.
Similar content being viewed by others


Introduction
Enterovirus D68 (EV-D68) belongs to human enterovirus species D of the genus Enterovirus within the family Picornaviridae. EV-D68 was originally isolated from four children with pneumonia and bronchiolitis in California, United States (US), in 19621. In contrast to other enteroviruses, EV-D68 is associated with respiratory tract infections (RTIs) because EV-D68 is an acid-sensitive virus that is biologically more similar to human rhinoviruses2. Since its first isolation, EV-D68 has been detected sporadically and only 26 isolates were identified in the US enterovirus surveillance between 1970 and 20053. However, between 2008 and 2010, increased reports of worldwide circulation of EV-D68 have been published in Asia, Europe, Africa and the US4,5,6. Besides RTIs, EV-D68 infection has been associated with rare cases of central nervous system (CNS) disease3,7.
The US and Canada have experienced widespread outbreaks of EV-D68 infections associated with severe respiratory disease since mid-August 2014. Underlying conditions such as asthma or wheezing have been reported in approximately 70% to 80% of EV-D68 cases8. More critically, Messacar K et al. found that the outbreak of EV-D68 was geographically and temporally associated with acute flaccid paralysis (AFP) and cranial nerve dysfunction in children9. There was a case report of AFP following EV-D68 infection in Europe in which EV-D68 was isolated from nasopharyngeal aspirate, stool and bronchoalveolar fluid specimens of the child10.
In a previous hospital-based viral surveillance study of RTI cases in Chongqing, China, from 2009 to 2012, the detection rate of EV-D68 was 0.4% (7/1565)11. However, the specific clinical characteristics and gene structural features of EV-D68 in China remain unclear. Therefore, the current study aimed to investigate the molecular epidemiology and clinical characteristics of EV-D68 in Chongqing, China.
Results
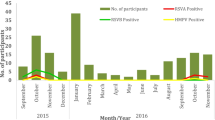
A total of 1876 NPAs were collected from children with RTIs between January 2012 and November 2014 (559 from 2012, 631 from 2013, 686 from 2014). Children enrolled in this study were of ages ranging from 1 month to 16 years and 11 months (median, 9 months). The overall male-to-female ratio was 1238: 638, with a 1.9 bias towards males. Among all samples, 1476 (78.7%) were positive for at least one viral agent. RSV was the most common viral agent (detected in 572 children [30.5%]), followed by HRV (473 children [25.2%]), PIV (384 children [20.5%]), HBoV (258 children [13.8%]), IV (190 children [10.1%]), ADV (110 children [5.9%]), HMPV (44 children [2.3%]), HCoV (27 children [1.4%]) and HEV (26 children [1.4%]). Among the 26 HEV positive samples, 19 were positive for EV-D68 (1.0%, 19/1876, Fig. 1). The frequency of EV-D68 detection was 0.4% (6/1681) from January 2012 through August 2014 and 9.8% (13/132) from September through October 2014. Moreover, the detection rate of EV-D68 in September-October 2014 was higher than in September-October 2012 (9.8%, 13/132 vs. 0%, 0/42, P = 0.04) and in September-October 2013 (9.8%, 13/132 vs. 2.4%, 3/127, P = 0.012).

Monthly distribution of EV-D68 in Chongqing, China, January 2012 to November 2014.
Bars indicate the number of specimens that were positive for EV-D68 and the line indicates the number of specimens tested.
Sequence analysis of VP1 revealed that the 19 strains in Chongqing could be grouped into two clades (clade A and clade B) that were the main epidemic genotypes in Chongqing from 2009 to 2012 (Fig. 2)11. Thirteen strains of 2014 Chongqing detected, seven strains from the 2014 US outbreak and most strains from the 2014 Netherlands detected were grouped into clade B. However, only two Chongqing strains from 2012 and 2013 could be grouped into clade B. The remaining four Chongqing strains were grouped into clade A (two in 2012 and two in 2013). Unfortunately, only thirteen Chongqing strains were nearly full-length nucleotide sequences, including ten strains from 2014, two strains from 2013 and one strain from 2012. Sequence analysis of nearly full-length nucleotide sequences found that the 13 clinical isolates detected in the current study had nucleotide sequence similarities of 89.9–99.8% with each other and similarities of 87.8–88.2% with the Fermon strain. Strains of clade B demonstrated similarities ranging between 94.3–99.8% (nucleotides). A phylogenetic tree constructed with nearly full-length nucleotide sequences revealed that the division of EV-D68 was consistent with Fig. 2 (Fig. 3).

Phylogenetic tree of selected EV-D68 strains based on the nucleotide sequence of VP1 genome regions.
The 369-bp fragments, which correspond to nt 2518-2886 of the EV-D68 prototype strain (GenBank accession no. AY426531), were used to construct the phylogenetic tree. ● Strains detected in this study. ■ EV-D68 previously detected in Chongqing from 2009–2012. ▲ Sequences from the US in 2014. ♦ Sequences from the Netherlands in 2014. ▼ Sequences from France in 2014.

Phylogenetic tree of selected EV-D68 strains based on the nucleotide sequence of 5′UTR to 3D genome regions.
The 7059-bp fragments, which correspond to nt 145-7203 of the EV-D68 prototype strain (GenBank accession no. AY426531), were used to construct the phylogenetic tree. ● Strains detected in this study ▲ Sequences from the US in 2014.
Kaida et al. first reported that EV-D68 Osaka strains have two deletions at nt 681–704 and 717–727 in contrast to the Fermon strain in the 5′UTR12. These EV-D68 Osaka strains were grouped into clade C according to phylogenetic analysis (Fig. 2). In the current analysis, fifteen Chongqing strains and seven strains from the 2014 US outbreak that were grouped into clade B also had two blocks of deletions at nt 681–703 and 717–728 in contrast with the Fermon strain (Fig. 4), which differed from clade C at nt 704 and 728. Tokarz et al. found that clade A only had a deletion at nt 681–7044. In the current analysis, four Chongqing strains (CQ2874, CQ2929, CQ5508 and CQ5753) and one strain from the 2014 US outbreak (US/KY/14-18953) that were grouped in clade A had a deletion at nt 682–704, which contrasted with the Fermon strain by a nucleic acid substitution at position 681 (Fig. 4).

Nucleotide regions of 5′untranslated regions of EV-D68 showed the deletion blocks preceding VP4.
The difference in deletions (position 704 and 728) between clade B and clade C are indicated by boxes.
The complete structural viral protein (VP4-VP1) amino acid sequences were also analyzed. Clade B had clearly different amino acid signatures relative to the prototype Fermon strain and had different signatures relative to clades A and C (Figure S1). However, all clade B amino acid residue substitutions have been reported in previous studies4,5,6,13,14,15.
The clinical and epidemiological characteristics of 19 children positive for EV-D68 are shown in Table 1 and 2. The children with EV-D68 ranged in age from 1 month to 12 years and 8 months (median, 32 months). The male-to-female ratio was 10:9. The most common signs and symptoms exhibited by the children were cough (100%) and wheezing (90%), with only five (26%) children having fever. Thirteen (68%) children had a history of recurrent wheezing (Table 2). Thirteen children had a discharge diagnosis of asthma, in which twelve children dually diagnosed with pneumonia. Among these thirteen children, eleven were diagnosed with acute asthma exacerbation. Moreover, three children were diagnosed with pneumonia, one child was diagnosed with bronchiolitis, one child was diagnosed with Mycoplasma pneumonia and one child was diagnosed with bronchitis (Table 1). EV-D68 was detected as a sole pathogen in 8/19 cases (with the exception of other common respiratory viruses, bacteria and Mycoplasma). Children diagnosed with asthma were more commonly associated with cases of EV-D68 single detection than co-detection (100% vs. 45%, P = 0.018, Table 2). The clinical isolates of EV-D68 in 2014 were all in clade B according to phylogenetic analysis. Comparisons were made between clades B and A (Table 2). Children infected with clade B viruses were significantly older than those infected with clade A viruses (median of 43 months old vs. 4.5 months old, P = 0.001). The duration of hospitalization was shorter for clade B-infected children than for clade A-infected children (median of 5 days vs. 8.5 days, P = 0.027). Diagnoses of asthma and acute asthma exacerbation were more common among children infected with clade B viruses than those with clade A (87% vs. 0%, P = 0.004; 73% vs. 0%, P = 0.018). Moreover, among the thirteen clinical isolates of EV-D68 in 2014, eleven cases were among children diagnosed with acute asthma exacerbation. Of these, five and six cases were among children diagnosed with severe and moderate acute asthma exacerbations, respectively (Table 1).
Among the 1876 NPAs samples, 75 were obtained from children admitted to the hospital with acute asthma exacerbation. The pathogen spectrum among the children with acute asthma exacerbation is shown in Table 3 with HRV and RSV predominating. HRV and RSV were the predominant pathogens associated with acute asthma exacerbation in September and October of 2012 and 2013. EV-D68 was the predominant pathogen in September and October of 2014 following the decline of HRV.
Discussion
Combined with the results of the previous study in Chongqing, thirteen cases of EV-D68 were detected during the period 2009–2013: two in 2010, four in 2011, four in 2012 and three in 201311. EV-D68 infection was substantially higher in September and October 2014 than in previous years.
Recent studies have shown that EV-D68 infection presents from mild respiratory illness to severe acute lower respiratory tract infection and even death3,8,11,12,13,14,15,16,17,18. Additionally, increasing evidence supports the relationship between EV-D68 infection and AFP3,7,9,10. Hasegawa et al. found a strong association between EV-D68 infection and exacerbation of asthma19. In the 2014 EV-D68 outbreak in the US, the percentage of children who had a history of asthma or wheezing in Kansas City and Chicago was 68% and 73%, respectively and most of these children were admitted to the pediatric intensive care unit8. At the St. Louis Children’s Hospital in St. Louis, Missouri, EV-D68 was detected in seven of ten samples from patients with severe disease20. In our study, most children infected with EV-D68 had a history of recurrent wheezing or asthma and a diagnosis of acute asthma exacerbation. In EV-D68 single detection, the phenomenon was more obvious. Moreover, EV-D68 became the predominant pathogen associated with asthma exacerbation in September and October of 2014. Our study found that a history of recurrent wheezing or asthma may be a risk factor for the detection of EV-D68 and viral-induced acute asthma exacerbation may be a clinical feature of EV-D68 infection in Chongqing area.
The current study found that clade B viruses had a stronger association with asthma exacerbation than clade A, but children infected with clade A viruses were too young to be diagnosed with asthma21. Therefore, the difference between clade B- and clade A-induced asthma exacerbations could not be inferred. In addition, the duration of hospitalization was longer in clade A-infected children. However, the number of cases was too small to estimate differences in disease severity between clade B and clade A. Further studies are thus needed to explore additional relationships between different clades.
The 5′UTR contains the internal ribosome entry site (IRES) responsible for cap-independent initiation of translation as well as other secondary structural elements responsible for genome replication22. Studies have found that the enterovirus IRES is associated with translational efficiency and virulence of the enterovirus4,23,24. The 5′UTR spacer region of EV-D68 ranged from the end of the IRES and the polyprotein open reading frame (ORF)4. The role of the spacer region in viral fitness remains unclear4. Deletions in the spacer region might be correlated with the worldwide increase in EV-D684,12. Our study showed different deletions among clades A, B and C. Clade A only had one deletion (nt 682–704)4, clade C had two deletions (nt 681–704 and 717–727)12 and clade B had two deletions (nt 681–703 and 717–728)5. The 5′UTR was considered a conserved region but the spacer region likely had more mutations. Therefore, more sequences would need to be collected to explore the nucleic acid differences of 5′UTR sequences among different clades. More studies are needed to expound the relationship between the deletions within the 5′UTR of EV-D68 and efficiency of translation, or virulence.
The VP1 contains a serotype specific neutralization site, the BC and DE loops, associated with viral antigenicity6. Previous studies found some distinctively different amino acid substitutions among the three clades, particularly in the BC and DE loops of the VP1 protein4,5,6,13,14,15. Such changes might modify the immune response and help to explain the biological basis for the increase in EV-D68 incidence4,13. However, an association between the substitutions and the severity of infection has not been established25. Phylogenetic analysis of VP1 sequences suggested that the major EV-D68 epidemic clade varied across time. The epidemic clades of EV-D68 in the Philippines (2008), New York (2009), Japan (2010) and the Netherlands (2010) were clade B, clade A, clade C and clade A, respectively12,16,17,26. The predominant clade might shift from clades A and C to clade B and clade B might expand worldwide. Although the main 2014 EV-D68 epidemic clade in Chongqing, the US and the Netherlands was clade B, it was different from the epidemic cluster observed in the Philippines in 2008 and no prior increase was reported. Moreover, the EV-D68 strains in certain areas had regional features as evidenced by the fact that the EV-D68 epidemic strains in Chongqing and US belonged to a different cluster than the Netherlands strains. This phenomenon may be linked to the rapid evolution of enteroviruses27.
Our study has several limitations. First, as this study was limited to inpatients, we could not analyze the clinical characteristics in outpatients and asymptomatic children. Viral surveillance was performed in a tertiary referral hospital where diseases seen were often more severe, thus more EV-D68 might have been detected. Second, because we only collected the NPAs of children with RTIs, no children positive for EV-D68 in our study had muscle weakness or paralysis. We could not find any relationship between AFP and EV-D68.
In summary, we found that clade B of EV-D68 increased in September and October 2014 in Chongqing, China. A history of recurrent wheezing or asthma appeared to be a risk factor for the detection of EV-D68 and viral-induced acute asthma exacerbation might be a clinical feature of EV-D68 infection. Our study provides additional epidemiological and clinical data on EV-D68 infection in China and a resource of genome sequences for genomic comparison of EV-D68. At present, EV-D68 is not included in the viral surveillance of ARTIs in China, but it has emerged as a considerable global public health threat. Additional surveillance of EV-D68 is needed.
Methods
Ethics statement
This study was approved by the Ethics Committee of the Children’s Hospital of Chongqing Medical University. The study was conducted in compliance with the principles of the Declaration of Helsinki. Informed consent was obtained from each parent or guardian prior to enrollment.
Patients and sample collection
The study was conducted at the Department of Respiratory Medicine at the Children’s Hospital of Chongqing Medical University (CHCMU) in Chongqing, China. Chongqing is located in a subtropical region and CHCMU is the largest children’s hospital in southeastern China. Nasopharyngeal aspirates (NPAs) were collected from children with RTIs11. A standardized questionnaire was designed to obtain the patients’ information from medical records, including demographic characteristics, underlying medical conditions, symptoms and signs, laboratory tests, chest radiograph results, hospital course and discharge diagnoses.
Clinical assessments
We defined four clinical variables related to wheezing: (1) a history of asthma, which was determined by asking the parents whether their child had ever been given a definitive diagnosis of asthma by clinicians or by documentation of a previous diagnosis of asthma in the medical record; (2) a history of recurrent wheezing, which was determined by asking the parents whether their child had ever had the symptom of wheezing at least three times; (3) a discharge diagnosis of asthma, which was defined as any discharge diagnosis of asthma following the guidelines of the Global Initiative for Asthma (GINA);21 and (4) hospitalization for acute asthma exacerbation using criteria in accordance with previous and current published international guidelines28. The degrees of acute asthma exacerbations were divided into mild, moderate and severe28.
Molecular analysis
Viral DNA and RNA were extracted from 200-μL aliquots of the NPA samples by a QIAampMinElute Virus Spin kit (Qiagen, Hilden, Germany). RNA was applied as the template for cDNA synthesis using the SuperScript III First-Strand Synthesis System (Invitrogen, California, USA). All experimental methods were carried out according to the manufacturers’ instructions.
All samples were screened for influenza viruses (IVA-C), human parainfluenza viruses (PIV1-4), respiratory syncytial virus (RSV-A and RSV-B), human coronaviruses (HCoV-229E and HCoV-OC43), human metapneumovirus (HMPV), adenovirus (ADV) and human bocavirus (HBoV) using molecular methods, as described previously29,30. A primer pair targeting 400 bp of the 5′untranslated regions (5′UTR) of the human enterovirus (HEV) and human rhinovirus (HRV) genome was used to distinguish between HEV and HRV16. To determine EV-D68 sequences, we synthesized cDNA using specific primer pairs (Table 4)13,17. The sequenced regions were all structural viral proteins (VP4-VP1), partial 5′UTR regions and nonstructural viral proteins (2A-3D). PCR was performed using a TaKaRa Ex Taq PCR kit (Takara Biotechnology, Dalian, China) and all PCR products were sequenced by Shanghai Majorbio Bio-Pharm Technology.
Detection of bacteria and yeasts
Qualitative and semiquantitative cultures for bacteria and yeasts were performed using standard microbiological methods. For all samples, macroscopically distinct colonies were isolated from pure culture and standard methods for identification, typing and establishing antibiotic sensitivity patterns were used31.
Sequence analysis
Sequence data for each clinical strain were formatted and assembled by the Seqman program of DNASTAR Software (v5.0). The sequences of EV-D68 strains that were newly obtained in Chongqing have been deposited in GenBank under the accession numbers KT803582–KT803606. To study the distribution and diversity of EV-D68 identified in this study compared to other EV-D68 strains, multiple sequence alignments were made using the ClustalW function of the BioEdit program (v7.0.9). Phylogenetic trees were constructed using the MEGA neighbor-joining algorithm (5.05). The statistical significances of the tree topologies were tested via bootstrapping (1000 replicates). Only bootstrap values >70% are shown in each tree. A phylogenetic tree that was constructed with 369 bp of VP1 gene nucleotide sequences showed that modern EV-D68 strains were divided into three primary clades (A, B and C) according to the classification previously described by Tokarz (Fig. 2)4. Nearly full-length nucleotide sequences that correspond to nt 145–7203 of the EV-D68 prototype strain (GenBank accession no. AY426531) were obtained from 13 strains of EV-D68 in the current study. A phylogenetic tree was constructed based on this region of nucleotide sequences with the 13 strains from Chongqing (Ten strains from 2014, two strains from 2013 and one strain from 2012), seven strains from the 2014 US outbreak and available EV-D68 sequences that were long enough (Fig. 3).
Statistical analysis
Data were analyzed using the SPSS 17.0 software package. Categorical variables were compared using chi-square or Fisher’s exact tests and continuous variables were compared using the nonparametric Mann-Whitney U-test. Two-sided P-values of <0.05 were considered statistically significant.
Additional Information
How to cite this article: Xiao, Q. et al. Prevalence and molecular characterizations of enterovirus D68 among children with acute respiratory infection in China between 2012 and 2014. Sci. Rep. 5, 16639; doi: 10.1038/srep16639 (2015).
References
Schieble, J. H., Fox, V. L. & Lennette, E. H. A probable new human pi-cornavirus associated with respiratory diseases. Am J Epidemiol. 85, 297–310 (1967).
Oberste, M. S. et al. Enterovirus 68 is associated with respiratory illness and shares biological features with both the enteroviruses and the rhinoviruses. J Gen Virol. 85, 2577–84 (2004).
Khetsuriani, N., Lamonte-Fowlkes, A., Oberst, S. & Pallansch, M. A. Enterovirus surveillance-United States, 1970-2005. MMWR Surveill Summ. 55, 1–20 (2006).
Tokarz, R. et al. Worldwide emergence of multiple clades of enterovirus 68. J Gen Virol. 93, 1952–8 (2012).
Lauinger, I. L., Bible, J. M., Halligan, E. P., Aarons, E. J., MacMahon, E. & Tong, C. Y. Lineages, sub-lineages and variants of enterovirus 68 in recent outbreaks. PLoS One. 7, e36005 (2012).
Opanda, S. M., Wamunyokoli, F., Khamadi, S., Coldren, R. & Bulimo, W. D. Genetic diversity of human enterovirus 68 strains isolated in Kenya using the hypervariable 3′-end of VP1 gene. PloS One. 9, e102866 (2014).
Kreuter, J. D. et al. A fatal central nervous system enterovirus 68 infection. Arch Pathol Lab Med. 135, 793–6 (2011).
Midgley, C. M. et al. Severe respiratory illness associated with enterrovirus D68-Missouri and Illinois. MMRW Morb Mortal Wkly Rep. 63, 798–9 (2014).
Messacar, K. et al. A cluster of acute flaccid paralysis and cranial nerve dysfunction temporally associated with an outbreak of enterovirus D68 in children in Colorado, USA. Lancet. 10.1016/S0140-6736(14)62457-0 (2015).
Lang, M. et al. Acute flaccid paralysis following enterovirus D68 associated pneumonia, France, 2014. Euro Surveill. 19, 20952 (2014).
Lu, Q. B. et al. Detection of enterovirus 68 as one of the commonest types of enterovirus found in patients with acute respiratory tract infection in China. J Med Microbiol. 63, 408–14 (2014)
Kaida, A., Enterovirus 68 in children with acute respiratory tract infections, Osaka, Japan. Emerg Infect Dis. 17, 1494–7 (2011).
Imamura, T. et al. Antigenic and receptor binding properties of enterovirus 68. J Virol. 88, 2374–84 (2014).
Meijer, A. et al. Emergence and epidemic occurrence of enterovirus 68 respiratory infections in The Netherlands in 2010. Virology. 423, 49–57 (2012).
Linsuwanon, P. et al. Molecular epidemiology and evolution of human enterovirus serotype 68 in Thailand, 2006-2011. PLoS One. 7, e35190 (2012).
Imamura, T. et al. Enterovirus 68 among children with severe acute respiratory infection, the Philippines. Emerg Infect Dis. 17, 1430–5 (2011).
Rahamat-Langendoen, J. et al. Upsurge of human enterovirus 68 infections in patients with severe respiratory tract infections. J Clin Virol. 52, 103–6 (2011).
Centers for Disease Control and Prevention (CDC). Clusters of acute respiratory illness associated with human enterovirus 68--Asia, Europe and United States, 2008-2010. MMWR Morb Mortal Wkly Rep. 60, 1301–4 (2011).
Hasegawa, S., Hirano, R., Okamoto-Nakagawa, R., Ichiyama, T. & Shirabe, K. Enterovirus 68 infection in children with asthma attacks: virus-induced asthma in Japanese children. Allergy. 66, 1618–20 (2011).
Wylie, K. M. et al. Genome sequence of enterovirus D68 from st. Louis, Missouri, USA. Emerg Infect Dis. 21, 184–6 (2015)
Bateman, E. D. et al. Global strategy for asthma management and prevention: GINA executive summary. Eur Respir J. 31, 143–78 (2008).
Martínez-Salas, E., Pacheco, A., Serrano, P. & Fernandez, N. New insights into internal ribosome entry site elements relevant for viral gene expression. J Gen Virol. 89, 611–626 (2008).
Gromeier, M., Alexander, L. & Wimmer, E. Internal ribosomal entry site substitution eliminates neurovirulence in intergeneric poliovirus recombinants. Proc Natl Acad Sci USA 93, 2370–2375 (1996).
Li, R., Zou, Q., Chen, L., Zhang, H. & Wang, Y. Molecular analysis of virulent determinants of enterovirus 71. PLoS One. 6, e26237 (2011).
Gimferrer, L. et al. First Enterrovirus D68 (EV-D68) cases detected in hospitalized patients in a tertiary care university hospital in Spain, October 2014. Enferm Infecc Microbiol Clin. 10.1016/j.eimc.2015.01.008 (2015).
Tokarz, R. et al. Longitudinal molecular microbial analysis of influenza-like illness in New York City, May 2009 through May 2010. Virol J. 9, 8:288 (2011).
Kyriakopoulou, Z., Pliaka, V., Amoutzias, G. D. & Markoulatos, P. Recombination among human non-polio enteroviruses: implications for epidemiology and evolution. Virus Genes. 50, 177–88 (2015).
Reddel, H. K. et al. An official American Thoracic Society/European Respiratory Society statement: asthma control andexacerbations: standardizing endpoints for clinical asthma trials and clinical practice. Am J Respir Crit Care Med. 180, 59–99 (2009).
Tiveljung-Lindell, A. et al. Development and implementation of a molecular diagnostic platform for daily rapid detection of 15 respiratory viruses. J Med Virol. 81, 167–75 (2009).
Hao, R. et al. Correlation between nucleotide mutation and viral loads of human bocavirus 1 in hospitalized children with respiratory tract infection. J Gen Virol. 94, 1079–85 (2013).
Yu, D. et al. Impact of bacterial colonization on the severity and accompanying airway inflammation, of virus-induced wheezing in children. Clin Microbiol Infect. 16, 1399–1404 (6) (2010).
Acknowledgements
The authors acknowledge the assistance of the patients and their caregivers involved in the study, the staff of the Department of Respiratory Medicine of Children’s Hospital of Chongqing and the Key Laboratory of Developmental Diseases in Childhood at Chongqing Medical University and the Ministry of Education. This project was supported by the China Special Grant for the Prevention and Control of Infection Diseases (2012zx10004212) and National Key Specialty [2011] 873.
Author information
Authors and Affiliations
Contributions
Q.X. carried out the initial analyses and drafted the initial manuscript. Q.X., S.Z. and L.W. collected the nasopharyngeal aspirates and clinical informations, finished the detection of virus. L.R., X.X., Y.D. and Y.Z. advised on data analyses. X.Z., Z.L., Z.F. and A.H. reviewed and revised the manuscript. E.L. conceptualized and designed the study, reviewed and revised the manuscript. All authors approved the final manuscript as submitted and agree to be accountable for all aspects of the work.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Xiao, Q., Ren, L., Zheng, S. et al. Prevalence and molecular characterizations of enterovirus D68 among children with acute respiratory infection in China between 2012 and 2014. Sci Rep 5, 16639 (2015). https://doi.org/10.1038/srep16639
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep16639
- Springer Nature Limited
This article is cited by
-
Clinical and molecular epidemiology of enterovirus D68 from 2013 to 2020 in Shanghai
Scientific Reports (2024)
-
IL-17A plays a critical role in RSV infection in children and mice
Virology Journal (2023)
-
Clinical and molecular epidemiologic features of enterovirus D68 infection in children with acute lower respiratory tract infection in China
Archives of Virology (2023)
-
Upsurge of Enterovirus D68 and Circulation of the New Subclade D3 and Subclade B3 in Beijing, China, 2016
Scientific Reports (2019)
-
A cross-sectional seroepidemiology study of EV-D68 in China
Emerging Microbes & Infections (2018)