
Abstract
Enteroviruses (EVs) are important human pathogens associated with various clinical syndromes. This study represents an overview of non-polio enteroviruses (NPEVs) isolated from acute flaccid paralysis (AFP) surveillance in Shandong Province, China from 1988 to 2013. Altogether 792 and 170 NPEV isolates were isolated from stool specimens of 9263 AFP cases and 1059 contacts, respectively. Complete VP1 sequencing and typing on all 962 isolates revealed 53 NPEV types in which echovirus (E) 6 (7.6%), E14 (7.6%), E11 (7.4%), coxsackievirus (CV) B3 (7.4%), E25 (5.6%), CVB5 (4.9%), E7 (4.5%) and EV-A71 (4.4%) were the eight most commonly reported serotypes. Distinct summer–fall seasonality was observed, with June–October accounting for 79.3% of isolation from AFP cases with known month of specimen collection. Increase of isolation of EV-A71 and CVA—the predominant pathogens for the hand, foot and mouth disease—was observed in recent years. Sequence analysis on VP1 coding region of EV-A71 and E6 suggested Shandong strains had great genetic divergence with isolates from other countries. The results described in this study provide valuable information on the circulation and emergence of different EV types in the context of limited EV surveillance in China.
Similar content being viewed by others
Introduction
Enteroviruses (EVs) (genus Enterovirus, family Picornaviridae) are common human pathogens. EV infection is usually asymptomatic, but sometimes it is associated with diverse clinical syndromes ranging from minor febrile illness to severe, potentially fatal diseases such as aseptic meningitis, encephalitis, paralysis, myocarditis and neonatal enteroviral sepsis1,2. EV infections maintain to be an important public health problem. It is estimated that 10–15 million symptomatic enterovirus infections occurred in the United States each year3.
EVs are small, nonenveloped, single-stranded RNA viruses. EV RNA is approximately 7.5 kb long and encodes a polyprotein that is processed to yield the mature structural (VP1 to VP4) and nonstructural proteins1. EV serotypes have traditionally been classified into echoviruses (E), coxsackieviruses (CV) group A and B and polioviruses (PV) by neutralization test and molecular typing method based on VP1 sequences has led to the discovery of a large number of new EV types4,5,6. So far, human EVs comprises more than 100 types which are classified into 4 species, EV-A to EV-D7.
PV infection is known to be associated with acute paralytic poliomyelitis. The standard approach recommended by World Health Organization (WHO) for polio surveillance is the detection and investigation of acute flaccid paralysis (AFP) cases, which includes standardized virological analysis of faecal samples of the patient, and/or sometimes those from contacts. Systematic AFP surveillance was initiated in China in 1994 and earlier experience on a small scale in Shandong Province could be dated back to 1988. Since 1996, annual reported AFP cases remained stable around 400. Besides PVs, a considerable proportion of non-polio enteroviruses (NPEVs) can be obtained from AFP surveillance as well. Since there is no specific EV surveillance system in mainland China yet, the AFP surveillance is the only data source for understanding EV circulations.
In China, large-scale outbreaks of hand, foot and mouth disease (HFMD) occurred repeatedly since 2007 and subgenogroup C4 was revealed to be the sole viral genetic lineage circulating in mainland China since then8. However, existence of C2 subgenogroup of EV-A71 in Shandong in 1996 was demonstrated in AFP surveillance9, suggesting retrospective investigation on EVs from AFP surveillance can provide valuable information on EV circulation. A study in Yunnan Province has identified 46 NPEV serotypes from AFP surveillance in 1997–2000 and 200410. However, to the best of our knowledge, no similar study has been reported in other provinces of China yet.
Shandong is a coastal province with a large population (~96 million) and major ports that could potentially serve as portals for importation of exogenous viruses. Here we describe the molecular typing and transmission patterns of all NPEV isolates detected during 26 years surveillance along with the phylogenetic analysis of two pre-dominant types EV-A71 and E6.
Results
Cases and NPEV isolation
The AFP surveillance was conducted in all 138 counties of Shandong Province and more than 600 sentinel hospitals were included. Hospitalized patients < 15 years of age were reported and male to female ratio was 1.9:1 for all cases from 1990 to 2013. The data for the cases in AFP surveillance in the years 1988 and 1989 were unavailable. Nevertheless, 4 NPEV strains were isolated during that period. During 1990–2013, a total of 9263 AFP cases were reported. EV isolation was performed on all stool specimens and 788 NPEV strains were isolated from stool specimens of these cases. Also, specimens from 1059 contacts of AFP cases were collected during this period and 170 NPEVs were isolated. The annual numbers of reported AFP cases and isolates were illustrated in Fig. 1. AFP surveillance in China increased substantially throughout the 1990s, but the surveillance and report before 1995 was not so active, resulting in the low level of NPEV isolation. It should also be notified that the absence of information of contacts and related isolates before 2007 (Fig. 1) are due to the unavailable information, not indicating that no specimens from contacts were collected at that time.

Annual numbers of AFP cases and contacts and isolation in Shandong Province, China, 1988–2013.
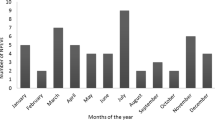
The monthly distributions of AFP cases and NPEV isolation were illustrated in Fig. 2. AFP reports increased in summer and autumn months with a peak in July (1100 [11.9%] of 9255 reports with known month during 1990–2013). However, a more prominent seasonality of NPEV detection from AFP cases was observed with June–October accounting for 79.3% (629 of 792) of isolation with known month of specimen collection.

Numbers of AFP cases and NPEV isolates from AFP cases by month of specimen collection — Shandong, 1990–2013.
Serotype
Molecular typing was performed successfully on all 962 isolates from AFP cases and contacts during 1988–2013 and a total of 53 NPEV types were identified (Table 1, 2). Of these, the 16 most commonly reported serotypes (E6, E14, E11, CVB3, E25, CVB5, E7, EV-A71, E3, E30, CVB1, CVA9, CVB2, CVA24, E13 and CVA4) accounted for 74.6% of all isolation. The eight most common serotypes accounted for approximately half of all reports (49.3%). A total of 19 isolates belonging to 9 new EV types was identified including EV-A76, EV-A90, EV-B73, EV-B74, EV-B75, EV-B80, EV-B87, EV-B97 and EV-C96 (Table 1, 2). Co-infection of different serotypes was observed in 15 patients and 1 contact.
During 1988–2013, NPEVs belonging to species A, B and C accounted for 10.5% (101 of 962), 84.4% (812 of 962) and 5.1% (49 of 962) of total isolation, respectively. No EV-D virus was identified. EV-B is the major species in most years of the study period. However, dramatic increase in EV-A constitution in AFP cases was observed in recent years (Table 1). Especially in 2011 and 2013, EV-A accounted for 39.0% (16 of 41) and 56.3% (18 of 32) of annual isolation, respectively. Common HFMD pathogens, such as EV-A71, CVA2, CVA4, CVA6, CVA10 and CVA16 were all present in the 26-y surveillance. Interestingly, several EV-A viruses had been observed in AFP surveillance before the onset of national HFMD epidemic in China in 2007. The first isolation of EV-A71, CVA2, CVA4, CVA6 and CVA10 can be dated back to the year 1996, 1996, 1996, 1992 and 2004, respectively.
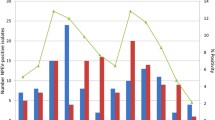
The most commonly isolated serotypes changed over time. Long-term trends of circulation of eight predominant NPEV serotypes were illustrated in Fig. 3 and epidemic pattern of circulation was observed for all serotypes including EV-A71, which remained rare before 2010 whereas increased dramatically since 2011. In 2011 and 2013, EV-A71 accounted for 14.6% (6 of 41) and 31.2% (10 of 32) of annual isolation, respectively. Epidemic pattern of circulation of E6, E11 and CVB5 was most distinct with regular sharp increases every 3–6 years, which usually last for 1–3 years (Fig. 3). No geographical aggregation was observed for these NPEV types.

Annual number of isolation of 8 predominant serotypes from AFP cases, Shandong, 1990–2013.
Annual isolation of E6, E11, E14 and EV-A71 is illustrated in Fig. 3A and that of CVB3, CVB5, E7 and E25 is illustrated in Fig. 3B.
VP1 sequence analysis of EV-A71
Altogether 42 EV-A71 isolates, 32 from AFP patients and the rest 10 from contacts, were identified during the AFP surveillance in 1988–2013. During the period from 1988 to 2002, no EV-A71 strains were isolated except for one C2 subgenogroup strain in 19969. Since 2003, continuous detection of EV-A71 was observed and they all belonged to C4a subgenogroup. Homologous comparison revealed 91.8% to 99.8% VP1 nucleotide sequence similarities among C4 subgenogroup strains from AFP surveillance and 81.4% to 83.6% with prototype strain BrCr. Phylogenetic analysis was performed on the complete VP1 sequences of the isolates from AFP surveillance with global reference sequences and those previously obtained from patients with HFMD and aseptic meningitis in Shandong Province11 (Fig. 4). Except for the C2 subgenogroup strain in 1996, Shandong AFP strains were closely related to local HFMD and aseptic meningitis isolates from 2007 to 2010 and belonged to C4a subgenogroup.

Phylogenetic tree based on EV71 VP1 sequences of Shandong isolates and reference strains of genogroup C.
Circle indicates the prototype strain, triangles indicate strains from AFP surveillance, squares indicates Shandong HFMD reference strains from 2007 to 2010 and diamonds indicate Shandong strains from aseptic meningitis patients. The years of isolation are shown as the first two digits in the names of the strains for AFP isolates and as the last two digits in the names of the strains for strains from HFMD and aseptic meningitis.
VP1 sequence analysis of E6
A total of 73 E6 strains were isolated during the AFP surveillance in 1988–2013. Homologous comparison revealed 77.3% to 100% VP1 nucleotide sequence similarities among themselves and the overall mean p-distance value is 0.162. Phylogenetic analysis was conducted on the complete VP1 sequences of the isolates with global reference E6 sequences. Shandong E6 strains segregated into 4 clusters, A–D (Fig. 5). Cluster A consisted of isolates from 2007 to 2012, indicating their circulation in Shandong is recently established. Nine strains from other provinces of China also belonged to this cluster. Cluster B consisted of isolates from 1988 to 2002. Members of this cluster disappeared for 11 years, suggesting it might become extinct in Shandong. Cluster C was composed of isolates from 1993 to 2011, indicating this cluster had circulated in Shandong province for a long time. Great genetic divergence was observed with other E6 strains (mean p-distance between groups, 0.204). Cluster D consisted of only 1 strain from 2007. It has close relationship with a strain from aseptic meningitis patient in Anhui Province, China in 2005. They were located in a branch consisted of strains from Australia, France, Russia and Korea. However, no other Chinese strain of this branch was observed, suggesting this cluster might be imported into China recently and circulated at a low level.

Phylogenetic tree based on E6 VP1 sequences of Shandong isolates and global strains.
Red triangles indicate Shandong strains from AFP surveillance and green squares indicate strains from other provinces of China. Circle indicates the prototype strain and VP1 sequence of E29 strain JV-10 serves as an outgroup.
Discussion
Since the Global Polio Eradication Initiative (GPEI) launched in 1988, the global incidence of poliomyelitis has dropped a lot. No cases due to wild-type poliovirus 2 (WPV2) have been identified since 199912. However, AFP surveillance is still of great importance in monitoring re-emergence of WPV poliomyelitis in many previously polio-free countries and emergence of vaccine-derived polioviruses (VDPVs)13. During AFP surveillance in Shandong, no WPV has been isolated since 1992 and VPDV had been detected in 2007 (P1, 9 nt; P1, 13 nt), 2009 (P2, 11 nt) and 2011 (P1, 10 nt), respectively. They were all ambiguous VPDV (aVDPV) which were clinical isolates from persons with no known immunodeficiency. In USA, the National Enterovirus Surveillance System (NESS) has provided valuable information for investigating temporal patterns of circulation of different serotypes, guiding outbreak investigations and identifying targets for development of diagnostic assays and antivirals2. Whereas in China, EV surveillance based on human specimens is very limited and AFP surveillance provided the only information on EV circulation.
Since the correlation of serotype with VP1 sequence was demonstrated5, molecular techniques of EV typing are becoming increasingly available and new EV types continue to be identified14,15,16,17,18. In the 26-y surveillance, a total of 9 new EV types were identified. The numbers of isolates of each new type ranged from 1 to 4, accounting for a small proportion of total isolation (2.0%, 19/962). According to a previous 5-y study in Yunnan Province10, some of these new EV types such as EV-A76, EV-B75, EV-B80 and EV-C96 were also found. There is a long geographic distance (>1800 km) between the two provinces and the co-detection of these serotypes suggest they had a wide spread in mainland China, although with low levels of activity. However, the eight most common serotypes in the Yunnan study were E13, E14, E12, CVB3, E2, E6, EV-C96 and E1, which are quite different with the results of this study (E6, E14, E11, CVB3, E25, CVB5, E7 and EV-A71). Considering the results of E11, E13, E7, E6, CVA24, E30, E19 and E29 as the eight most common serotypes in Philippines in a 17-y study19, the variation in constitution of most common serotypes suggests EV serotypes have incongruent epidemic magnitude under different spatio-temporal conditions.
In mainland China, nationwide epidemic of HFMD has occurred repeatedly since 2007. EV-A71 and CVA16 are the two major causative agents in different areas and other coxsackieviruses of species A, such as CVA2, CVA4, CVA6, CVA10, CVA12, etc, have also been demonstrated to be involved20,21,22. Consistently, an increase of isolation of EV-A71 and other EV-A serotypes from AFP cases was also observed (Table 1, Fig. 3). EV-A71 has been demonstrated to be associated with neurological diseases in young children including aseptic meningitis23, poliomyelitis-like paralysis24,25 and fatal brain-stem encephalitis26. The increase in isolation from AFP cases suggests since the large-scale epidemic of HFMD in 2007, EV-A71 associated serious neurological diseases have become a major public health concern in mainland China, which is consistent with the observation of emerging EV-A71 detection in aseptic meningitis patients in a previous study in Shandong Province27.
Shandong Province is located in temperate zone. EV infections are peaked in summer-fall seasons in temperate climates and the results from our study are consistent with the seasonality (Fig. 2). According to results from EV surveillance in USA in 1970–2005, long-term trends of circulation of different EV serotypes have two major patterns: epidemic and endemic. Epidemic pattern is characterized by substantial fluctuations in circulation levels over time, including large peaks when the serotype was among the most prevalent enteroviruses reported for a given year. Endemic pattern has stable and usually low-levels of circulation with few distinct peaks2. In this study, all eight most common serotypes displayed epidemic pattern of long-term trends, although these data are from AFP surveillance, not specific EV surveillance system. EV-A71 had large-scale nationwide epidemic in current China and an increase of isolation was observed in AFP cases in this study, whereas no similar trend was observed in the surveillance from USA2,28. Further surveillance will help understand the long-term trend of EV-A71 circulation in China under the circumstance of continuous epidemics of HFMD. In mainland China, C4 was demonstrated to be the predominant subgenogroup responsible for most documented HFMD outbreaks and epidemic since 19988 and Shandong strains from meningitis patients in 2010 also belonged to subgenogroup C4a11. VP1 sequence analysis in this study revealed close genetic relationship among EV-A71 C4 subgenogroup strains from AFP, HFMD and aseptic meningitis patients, suggesting C4 is an important pathogen of multiple clinical manifestations.
E6 is one of the most frequently isolated serotype in many regions of the world. Its infection includes aseptic meningitis, meningoencephalitis, rashes, gastrointestinal illnesses, neonatal hepatitis and pneumonitis. In China, outbreaks and sporadic aseptic meningitis cases caused by E6 have been reported recently27,29. In the VP1 phylogenetic tree, Shandong strains were grouped into four clusters. Except that the only member of cluster D had close relationship with strains from France, Australia, Russia and South Korea, the rest 3 clusters contained Chinese strains exclusively, suggesting the existence of geographic barrier between mainland China and other regions of the world. Frequent travel might increase importation or exportation of different E6 lineages.
Co-infection was detected in 15 patients and 1 contact. It was identified in two kinds of situations. First, in this study, entire VP1 sequence of an EV strain was obtained via combination of 5′ end and 3′ end sequences. So, if molecular typing on 5′ end and 3′ end PCR sequences revealed different types, this indicates a mixture of different EV types in one isolate. Second, RD and HEp-2 cell lines have been demonstrated to possess different sensitivity to different serotypes. RD is sensitive to echovirus and EV-A CVAs, while HEp-2 is sensitive to CVBs and some EV-C types. So, isolation of different EV types from a single specimen was observed from different cell lines. In comprehending the surveillance results of this study, the limitation of virus isolation should be considered. Some serotypes, such as some coxsackie A viruses of EV-C species, cannot produce visible CPE in these cell lines. So, these viruses cannot be recovered in the cell culture method. Hence, the actual EV distribution in the population might not be completely identical with that reflected by this study. Also, this study describes EV typing results from AFP surveillance. Since some NPEVs, such as EV-A71, had been demonstrated to be the pathogens of AFP, the results of this study might be different from usual EV surveillance (infections often milder) and do not necessarily directly reflect to general EV circulation.
China has a vast territory and a large population. The EV circulation in Shandong Province is not necessarily consistent with that of other regions in China. However, EV has the ability of transmission over long distance and frequent transmissions over cities or provinces in China have been observed previously8,30. So, the results of serotype spectrum, long-term pattern of circulation and molecular epidemiology can serve as a reflection of the situation in other parts of Eastern China. Further similar studies from other regions will help understand the EV circulation in the country.
Methods
Ethics statement
Ethical approval was given by the Ethics Review Committee of Shandong Center for Disease Control and Prevention and the methods were carried out in accordance with the principles of the Declaration of Helsinki. Written informed consents for the use of their clinical samples were obtained from all subjects (the legal guardians of the patients and contacts).
Virus isolation
Stool samples from the AFP patients and contacts were collected and processed according to standard procedures recommended by the WHO31. Contacts were collected for AFP cases with (1) <5 years of age, (2) <3 doses of OPV immunization or unknown OPV history, (3) unqualified stool specimen, or (4) clinical diagnosis of poliomyelitis. RD (human rhabdomyosarcoma cell line), HEp-2 (human epidermoid carcinoma cell line) and L20B (mouse cell line expressing the gene for the human cellular receptor for poliovirus) cell lines were used for virus isolation31. All cell lines were gifts from the WHO Global Poliovirus Specialized Laboratory in the USA and were all originally purchased from the American Type Culture Collection. A total of 200 μl of chloroform-treated stool solution was added to each of the cell culture tubes and the inoculated cells were examined daily. After 7 d, the tubes were frozen, thawed and re-passaged and another 7-d examination was performed. Infected cell cultures were harvested and used for further examination until complete cytopathic effect was obtained. To avoid cross contamination, cell tubes of normal L20B, RD and HEp-2 cells served as negative controls. Isolates from RD or HEp-2 cell lines were re-passaged to L20B cell line and were designated as NPEV if no CPE was observed.
VP1 amplification, sequencing and molecular typing
Total RNA was extracted from 140 μl of the infected cell culture using QIAamp viral RNA mini kit (Qiagen, Valencia, CA, USA) and reverse transcription-PCR (RT-PCR) was performed using Access RT-PCR System (Promega, USA) according to the manufacturers' procedures.
Primer pairs 486/488, 487/489 and 040/011 for EV-A strains and primer pairs 008/013, 490/492, 187/011, 491/493 and 012/011 for EV-B strains were used for amplification of partial VP1 sequences4,5,32. As for EV-C VP1 amplification, previously designed primer pairs 494/496, 495/497 and 040/0114,32 are not always efficient. So, we designed primer pairs UF1/UR1, UF2/UR2 and 040/DR1 (Table 3) in this study based on alignment of all available EV-C complete genome sequences from GenBank. These primers turned out to be efficient not only for all EV-C strains described in this study (CVA17, CVA20, CVA21, CVA24 and EV-C96), but also for Sabin PV1–3 and other available CVA13 and EV-C99 strains (from healthy population) of EV-C in our lab. Nevertheless, due to limited EVs in our lab, their performance on other EV-C viruses beyond the listed 10 serotypes had not been testified. Combination of 5′ end and 3′ end sequences yielded entire VP1 coding sequences. As the serotype and species were unknown before typing, EV-B primers were first used for RT-PCR amplification. And EV-A and EV-C primers were used in case of negative results by using EV-B primers. In order to avoid cross contamination, a RT-PCR reaction using the RNA extracted from normal RD cell served as a blank control and a negative control containing all the components of the reaction except for the template was also included.
PCR positive products were purified and sequenced bi-directionally with the BigDye Terminator v3.0 Cycle Sequencing kit (Applied Biosystems, Foster City, CA) and sequences were analyzed by ABI 3130 genetic analyzer (Applied Biosystems, Hitachi, Japan). Molecular typing based on VP1 sequences was performed using online Enterovirus Genotyping Tool version 0.133.
Homologous comparison and phylogenetic analysis
Sequence analysis was performed on two predominant serotypes, EV-A71 and E6. The entire list of VP1 sequences of global reference E6 and EV-A71 strains used in sequence analysis can be found as Supplementary Table S1 online. Nucleotide sequence alignments were carried out by BioEdit 7.0.5.3 software34. Phylogenetic trees were constructed by Mega 4.0 using neighbor-joining method after estimation of genetic distance using the Kimura two-parameter method35. A bootstrapping test was performed with 1,000 duplicates.
Additional Information
Nucleotide accession numbers. VP1 sequences used in this study were deposited in GenBank under accession numbers GQ253420–GQ253423, GQ329778–GQ329785, JQ326298–JQ326306, HQ829954–HQ829961, HQ399492–HQ399495 and KJ772420–KJ772493.
References
Enteroviruses: polioviruses, coxsackieviruses, echoviruses and newer enteroviruses. Fields Virology, 5th ed. (Lippincott Williams & Wilkins, Philadelphia, 2007).
Khetsuriani, N., Lamonte-Fowlkes, A., Oberst, S. & Pallansch, M. A. Centers for Disease Control and Prevention. Enterovirus surveillance–United States, 1970–2005. MMWR Surveill. Summ. 55, 1–20 (2006).
Strikas, R. A., Anderson, L. & Parker, R. A. Temporal and geographic patterns of isolates of nonpolio enteroviruses in the United States, 1970–1983. J. Infect. Dis. 153, 346–351 (1986).
Oberste, M. S. et al. Comparison of classic and molecular approaches for the identification of untypeable enteroviruses. J. Clin. Microbiol. 38, 1170–1174 (2000).
Oberste, M. S., Maher, K., Kilptrick, D. R. & Pallansch, M. A. Molecular evolution of the human enteroviruses: correlation of serotype with VP1 sequence and application to picornavirus classification. J. Virol. 73, 1941–1948 (1999).
Brown, B. A. et al. Resolving ambiguities in genetic typing of human enterovirus species C clinical isolates and identification of enterovirus 96, 99 and 102. J. Gen. Virol. 90, 1713–1723 (2009).
Picornaviridae. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses. (Elsevier, San Diego, 2011).
Zhang, Y. et al. Emergence and transmission pathways of rapidly evolving evolutionary branch C4a strains of human enterovirus 71 in the central plain of China. PLoS ONE 6, e27895 (2011).
Tao, Z., Wang, H. & Xu, A. Identification of a C2 subgenogroup strain of enterovirus 71 in a retrospective study in Shandong province, China, from 1990 to 2010. J. Clin. Microbiol. 50, 1823–1824 (2012).
Bingjun, T. et al. Molecular typing and epidemiology of non-polio enteroviruses isolated from Yunnan province, the People's Republic of China. J. Med.Virol. 80, 670–679 (2008).
Tao, Z. et al. Molecular epidemiology of human enterovirus associated with aseptic meningitis in Shandong Province, China, 2006–2012. PloS ONE 9, e89766 (2014).
Roberts, L. Polio eradication. Looking for a little luck. Science 323, 702–705 (2009).
WHO. Field Guide for Supplementary Activities Aimed at Achieving Polio Eradication. (World Health Organization, Geneva, 1995).
Oberste, M. S. et al. Molecular identification of 13 new enterovirus types, EV79-88, EV97 and EV100-101, members of the species Human Eneterovirus B. Virus Res. 128, 34–42 (2007).
Oberste, M. S. et al. Molecular identification and characterization of two proposed new enterovirus serotypes, EV74 and EV75. J. Gen. Virol. 85, 3205–3212 (2004).
Oberste, M. S. et al. Enteroviruses 76, 89, 90 and 91 represent a novel group within the species human enterovirus A. J. Gen. Virol. 86, 445–451 (2005).
Norder, H. et al. Sequencing of ‘untypable’ enteroviruses reveals two new types, EV-77 and EV-78, within human enterovirus type B and substitutions in the BC loop of the VP1 protein for known types. J. Gen. Virol. 84, 827–836 (2003).
Tang, J. et al. Complete genome characterization of a novel enterovirus type EV-B106 isolated in China, 2012. Sci. Rep. 4, 4255 (2014).
Apostol, L. N. et al. Detection of non-polio enteroviruses from 17 years of virological surveillance of acute flaccid paralysis in the Philippines. J. Med. Virol. 84, 624–631 (2012).
Lu, Q. B. et al. Circulation of coxsackievirus A10 and A6 in hand-foot-mouth disease in China, 2009–2011. PLoS ONE 7, e52073 (2012).
Zhang, Y. et al. An outbreak of hand, foot and mouth disease associated with subgenotype C4 of human enterovirus 71 in Shandong, China. J. Clin. Virol. 44, 262–267 (2009).
Zhang, Y. et al. Molecular evidence of persistent epidemic and evolution of subgenotype B1 coxsackievirus A16-associated hand, foot and mouth disease in China. J. Clin. Microbiol. 48, 619–622 (2010).
Gilbert, G. L. et al. Outbreak of enterovirus 71 in Victoria, Australia, with a high incidence of neurologic involvement. Pediatr. Infect. Dis. J. 7, 484–488 (1988).
Samuda, G. M., Chang, W. M., Yeung, C. M. & Tang, P. Monoplegia caused by enterovirus 71: an outbreak in Hong Kong. Pediatr. Infect. Dis. J. 6, 206–208 (1987).
Chumakov, M. et al. Enterovirus 71 isolated from cases of epidemic poliomyelitis-like disease in Bulgaria. Arch. Virol. 60, 329–360 (1979).
Lum, L. C. et al. Fatal enterovirus 71 encephalomyelitis. J. Pediatr. 133, 795–798 (1998).
Tao, Z. et al. Molecular epidemiology of human enterovirus associated with aseptic meningitis in Shandong Province, China, 2006–2012. PloS ONE 9, e89766 (2014).
Centers for Disease Control and Prevention. Nonpolio enterovirus and human parechovirus surveillance — United States, 2006—2008. MMWR Morb. Mortal. Wkly. Rep. 59, 1577–1580 (2010).
Mao, N. et al. An aseptic meningitis outbreak caused by echovirus 6 in Anhui province, China. J. Med. Virol. 82, 441–445 (2010).
Tao, Z. et al. Intercity spread of echovirus 6 in Shandong Province, China: application of environmental surveillance in tracing circulating enteroviruses. Appl Environ Microbiol 78, 6946–6953 (2012).
Isolation and identification of polioviruses. WHO Polio Laboratory Manual, 4th edn. (World Health Organization, Geneva, 2004).
Oberste, M. S. et al. Species-specific RT-PCR amplification of human enteroviruses: a tool for rapid species identification of uncharacterized enteroviruses. J. Gen. Virol. 87, 119–128 (2006).
Kroneman, A. et al. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 51, 121–125 (2011).
Hall, T. A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98 (1999).
Tamura, K., Dudley, J., Nei, M. & Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599 (2007).
Acknowledgements
This study was supported by a grant from the Science and Technology Department of Shandong Province (2012YD18044), a grant from the National Natural Science Foundation of China (81302481), two grants from the Health Department of Shandong Province (2011QZ013, 2011HZ058) and the Key Technologies Reaseach & Development Program of National Ministry of Science and Technology (Grant no. 2013ZX10004-202).
Author information
Authors and Affiliations
Contributions
Z.T., H.W., Y.L. (Yao Liu), Y.L. (Yan Li), A.X. and L.S. conceived the study and drafted the paper, P.J., X.L., M.L., S.W., Y.Z., Y.F., Q.F., H.Y., J.Y., P.C. and W.L. gathered and analyzed the data and G.L., F.J, L.F. and P.X. helped to interpret results and contributed to the writing. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Table S1
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Tao, Z., Wang, H., Liu, Y. et al. Non-Polio Enteroviruses from Acute Flaccid Paralysis Surveillance in Shandong Province, China, 1988–2013. Sci Rep 4, 6167 (2014). https://doi.org/10.1038/srep06167
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06167
- Springer Nature Limited
This article is cited by
-
Genotyping of non-polio enteroviruses associated with acute flaccid paralysis in Thailand in 2013 and 2014
Virology Journal (2021)
-
Prevalence and Bayesian Phylogenetics of Enteroviruses Derived From Environmental Surveillance Around Polio Vaccine Switch Period in Shandong Province, China
Food and Environmental Virology (2020)
-
Molecular characterization of non-polio enteroviruses isolated from children with acute flaccid paralysis in IRAN, 2015–2018
Virus Genes (2020)
-
Multiple genotypes of Echovirus 11 circulated in mainland China between 1994 and 2017
Scientific Reports (2019)
-
Complete genome characterization of three enterovirus C96 isolates in China
Archives of Virology (2019)