
Abstract
Transport of macromolecules through the nuclear envelope (NE) is mediated by nuclear pore complexes (NPCs) consisting of nucleoporins (Nups). Elys/Mel-28 is the Nup that binds and connects the decondensing chromatin with the reassembled NPCs at the end of mitosis. Whether Elys links chromatin with the NE during interphase is unknown. Here, using DamID-seq, we identified Elys binding sites in Drosophila late embryos and divided them into those associated with nucleoplasmic or with NPC-linked Elys. These Elys binding sites are located within active or inactive chromatin, respectively. Strikingly, Elys knockdown in S2 cells results in peripheral chromatin displacement from the NE, in decondensation of NE-attached chromatin, and in derepression of genes within. It also leads to slightly more compact active chromatin regions. Our findings indicate that NPC-linked Elys, together with the nuclear lamina, anchors peripheral chromatin to the NE, whereas nucleoplasmic Elys decompacts active chromatin.
Similar content being viewed by others

Introduction
Although the idea that chromosomes in interphase nucleus are attached to the nuclear envelope (NE) was proposed a long ago1, mechanisms mediating this attachment are still poorly understood. Accumulating evidence suggests that peripheral chromatin is in intimate association with both the nuclear lamina (NL) and the nuclear pore complexes (NPCs). NL consists of Lamins and Lamin-associated proteins2. Genomic regions interacting with the NL (lamina-associated domains, LADs) were successfully identified in different organisms, including Drosophila, mammals, and nematode3,4,5,6,7,8. Depletion of specific components of the NL resulted in the displacement of chromatin from the NE to the nuclear interior9,10,11,12,13,14,15,16,17,18,19, thus indicating that chromatin is attached to the NL and that interphase chromosomes are slightly stretched by this attachment19,20. In addition, chromosomes from yeast, Drosophila, and mammals interact with various nucleoporins (Nups) (i.e., proteins composing NPCs)21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37. However, contrarily to yeast, Nups-chromatin interactions in metazoans may take place not only at the NE but also in the nuclear interior27,28,29,31. For example, in Drosophila, a nucleoplasmic fraction of Nup98 interacts with active chromatin, whereas Nup98, as a component of NPCs, mostly associates with inactive chromatin28.
Several findings point to the involvement of NPCs in chromatin tethering to the NE in metazoans. The dosage-compensated single X chromosome in Drosophila SL-2 cells is stronger bound with Nup153 and is localized closer to the NE than autosomes29, whereas Nup153 depletion leads to repositioning from the NE of several Nup153-target loci in both Drosophila SL-2 and mouse embryonic stem cells29,33. Similarly, Nup Elys/Mel-28 (further Elys) interacts stronger with C. elegans male single X chromosome than with two X chromosomes of hermaphrodites which correlates with a less distant position of the male X chromosome from the NE38. In addition, Nup155 is involved in chromatin tethering to the NE in Drosophila S2 and ovarian germline cells39. However, despite these indications, the contribution of NPC-chromatin interactions to the maintenance of peripheral chromatin positioning as well as the impact of these interactions on the overall genome architecture and gene expression remain largely unexplored.
Elys may be the unique Nup that directly tethers chromatin to the NE40. Initially, it was identified as the AT-hook-containing transcription factor expressed in the embryonic hematopoietic tissues of mouse41. More recent experiments have shown that, during mitosis, Elys, together with the Nup107-160 complex, are concentrated on kinetochores in mammals, Xenopus, and the nematode42,43,44,45. Importantly, upon mitotic exit, Elys participates in the NPC reassembly by connecting the Nup107-160 complex to decondensing chromatin42,45,46,47. Accordingly, in the nematode, Xenopus, and mammalian cells, Elys depletion results in the partial loss of nuclear pores from the NE42,44,45,48. Partial loss is explained by the existence of an Elys-independent pathway of NPC incorporation into the NE during interphase48. Knockdown of Elys in Drosophila salivary glands caused the enhanced apoptosis and the disappearance of Lamins and Nups from the NE49.
Eight Elys molecules, as the components of NPC nuclear ring, face peripheral chromatin50. Elys contains β-propeller and α-helical domains at the N-terminal part, which are responsible for its association with the Nup107-160 complex51,52. The C-terminal part of Elys bears an AT-hook DNA-binding motif recognizing A/T-rich DNA sequences47 and an additional domain, able to bind with the acidic patch of a nucleosome52,53,54,55,56. Drosophila Elys also possesses β-propeller and α-helical domains but contains three non-canonical AT-hook-like motifs which were shown to be required for Elys binding to A/T-rich sequences in vitro49.
Elys interacts with SWI/SNF chromatin remodeling complex PBAP36 known to remove nucleosomes from the regulatory regions of the genome and to switch chromatin to a more open state57,58,59. Consistent with this ability, artificial tethering of Sec13, the partner of Elys in the Nup107-160 complex, to several sites on Drosophila polytene chromosomes resulted in the recruitment of Elys to these sites followed by local chromatin decondensation36. Yet, the ATAC-seq assay did not reveal notable changes in chromatin accessibility upon Elys depletion in S2 cells60. Elys binding sites across the entire Drosophila genome were recently identified by ChIP-seq in brains from 3rd instar larvae and in embryonic S2 cell line35,37. Elys ChIP-seq profile appeared to be highly correlated with the profile of H3K27 acetylation35. At the same time, a fraction of ChIP-seq sites common between Elys and Nup93 or between Elys and Nup107 was found to reside within LADs, thus indicating peripheral localization of the chromatin carrying these sites37. However, Elys ChIP-seq sites were not explicitly classified as bound by nucleoplasmic or NPC-linked Elys molecules. Therefore, the influence of Elys on chromatin state and on gene expression specifically in these two different locations was unclear. Moreover, it was not previously explored whether Elys tethers chromatin to the NE and maintains its peripheral positioning during interphase.
Here, we report that the knockdown of Elys (Elys-KD) in Drosophila S2 cells does not notably affect the abundance of NPCs at the NE and does not result in increased apoptosis, thereby keeping cells alive. These findings allow us to consider S2 cells as an appropriate model to analyze changes in genome architecture upon the lack of Elys. Using S2 cells and late embryos, we found that Elys, as a component of NPCs, binds with numerous genomic sites within LADs and this binding is required for the retention of peripheral chromatin at the NE during interphase. Consistent with these results, we found that, upon Elys-KD in S2 cells, the topologically associating domains (TADs)61,62,63,64 that are attached to the NL become less condensed, and genes located within them are slightly derepressed. These effects are similar to what was observed upon disruption of the NL in S2 cells19. Taken together, these findings support the model according to which the components of the NL, in cooperation with the NPC-linked Elys, bind peripheral chromatin and maintain its proper localization and functions in interphase nuclei.
Results
Elys-KD in Drosophila S2 cells does not lead to considerable loss of NPCs at the NE
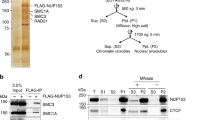
Knockdown of Elys in salivary glands from Drosophila third instar larvae impairs localization of major NE components, such as Nups, Lamins, and Lamin-B-receptor (LBR)49. Since salivary glands contain non-dividing cells undergoing endoreplication, we examined whether Elys is similarly required for the correct localization of NE components in the mitotically dividing embryonic S2 cells. We stained S2 cells with anti-Elys antibodies generated earlier65 and revealed Elys localization mostly at the nuclear rim (Fig. 1a, upper left panel). Elys colocalizes with the NPCs (stained with Mab414 antibodies which detect the phenylalanine-glycine-rich Nups) at the NE giving a characteristic punctate pattern (Fig. 1a, upper right panel). RNAi knockdown of Lamin Dm0 (Lam-KD; Supplementary Fig. 1a) in S2 cells results in NPC clustering66 at the NE manifested in the gaps in Mab414 and Elys staining (Fig. 1a, middle panel). However, contrary to the observations in salivary glands, Elys-KD in S2 cells resulting in a 6–13 fold decrease of Elys protein level (Supplementary Fig. 1) does not impair Lamin localization (Fig. 1a, low left panel; Supplementary Fig. 2a, b; Supplementary Data 1) and only slightly reduces LBR presence at the NE (Supplementary Fig. 2c, d; Supplementary Data 1). Strikingly, unlike in nematode, Xenopus, and mammals42,43,44,45,46, and unlike in Drosophila salivary glands49, Elys-KD in S2 cells does not lead to a notable loss of NPCs at the NE and to the emergence of cells lacking nuclear pores (Fig. 1a, low right panel; Fig. 1b, c; Supplementary Fig. 2c, d (for Nup153 localization); Supplementary Data 1). Only knockdown of both Lamin Dm0 and Elys (Fig. 1d, upper panel) results in the pronounced reduction of NPC staining at the nuclear rim (Fig. 1d, lower panel).

a, b, d Immunostaining of control, Lam-KD, Elys-KD, or (Lam-KD plus Elys-KD) S2 cells with anti-Elys, anti-Lamin and Mab414 antibodies (a), with anti-Elys and Mab414 antibodies (b), or with anti-LBR, anti-Elys and anti-Lamin antibodies (d). Scale bars 1 µm (a, d), 10 µm (b). c ImageJ quantification of Mab414 average fluorescence intensity (normalized on average Dapi fluorescence) across the NE in Elys-KD (two biological replicates, n = 75) and control (two biological replicates, n = 75) S2 cells. The ratio of median values is indicated. P-value was estimated in an M-W U-test. e Western-blot analysis of proteins, co-immunoprecipitated with anti-Elys or anti-Nup107 antibodies from S2 extracts, probed by anti-Elys, or anti-Nup107 antibodies (n.i. – non-immune serum, IP/input ratio 1:4.5). Note that the spliced bands ran in parallel on the same gel. The sizes of molecular weight markers are indicated to the right. f, g Immunostaining of S2 cells with anti-CenpA (kinetochores, violet), anti-Elys (red), anti-α-Tubulin (green) in metaphase (f), with anti-Elys (red), anti-Mab414 (green), anti-LBR (violet), Dapi (blue) in anaphase (g). Scale bars 1 µm (f, g). Arrows point to the Elys concentrated around decondensing chromatin (g). h Immunostaining of control and Elys-KD S2 cells with Mab414 antibodies (green) counterstained with Dapi (blue) in telophase. Scale bar 1 µm.
These surprising results prompted us to examine Elys behavior during mitosis in S2 cells. Post-mitotic NPC reassembly in other organisms is initiated in anaphase by Elys binding to the decondensing chromatin, followed by Elys-mediated recruitment of the Nup107-160 subcomplex of NPC42,45,46,47,48,67,68. We found that Elys and Nup107 reciprocally co-immunoprecipitate each other from protein extracts of S2 cells (Fig. 1e and Supplementary Fig. 3), therefore indicating a physical association between these Nups, at least during the interphase. In mammals, Xenopus and nematode, a fraction of Elys, being in the complex with Nup107-160, is localized on kinetochores during mitosis42,43,44,45,52. However, Mehta et al.49 revealed Elys does not stay on kinetochores in Drosophila. We confirmed their findings by staining S2 cells with anti-α-tubulin and anti-Elys antibodies, which demonstrates diffuse Elys distribution around condensed chromosomes, but not at the kinetochores during metaphase (Fig. 1f). In anaphase, Elys appears to concentrate around decondensing chromatin earlier than other Nups or LBR (Fig. 1g), thus pointing to the Elys leading role in determining seeding sites on chromatin upon the post-mitotic NPCs reassembly in Drosophila S2 cells, as shown in other organisms42,45,46,47,48,67,68. Nevertheless, the lack of Elys in S2 cells does not prevent the appearance of NPCs at the NE at the end of mitosis (Fig. 1b). Moreover, upon Elys-KD, the incorporation of NPCs in the NE occurs when cytokinesis is not yet complete (Fig. 1h). Consistent with these results, we did not reveal any disturbance of mRNA export from the nucleus upon Elys-KD (Supplementary Fig. 4), thus indicating that nuclear-cytoplasmic traffic of RNA via nuclear pores functions normally. Furthermore, using flow cytometry, we found only minor changes in cell cycle progression upon Elys-KD (Supplementary Fig. 5). We conclude that Elys-dependent incorporation of NPCs in the NE at the end of mitosis may be bypassed in S2 cells by rapidly acting Elys-independent mechanism.
RNAi knockdown of Elys using ubiquitous act5C-GAL4 driver was shown to induce apoptosis in Drosophila salivary glands49. We explored whether Elys-KD in S2 cells also results in enhanced cell death. Unexpectedly, the TUNEL assay did not detect any increase in the proportion of apoptotic cells upon Elys-KD in comparison with the control cells (3.6% versus 3.3%, respectively, P = 0.99, M-W U-test, Supplementary Fig. 6; Supplementary Data 2).
In summary, the absence of notable NPC and NL defects, as well as normal cell viability upon Elys-KD makes S2 cells an appropriate model to explore the influence of Elys depletion on the genome architecture.
Elys binds to numerous sites on polytene chromosomes
Several Nups, including Sec13, occupy dozens of sites on polytene chromosomes mostly located away from the NE27. Artificial tethering of Sec13 to several ectopic sites on polytene chromosomes results in recruiting of Elys to these sites, correlated with local chromatin decondensation36. Using immunostaining of squashed polytene chromosomes with anti-Elys antibodies, we revealed that Elys by itself binds to the plethora of sites, some of them are colocalized with polytene chromosome bands (pink strips on Fig. 2a), while others are colocalized with inter-bands (red strips on Fig. 2a). Polytene chromosome bands and inter-bands are known to contain silent and expressed genes, respectively69. Since numerous fluorescence in situ hybridization (FISH) experiments indicate that active chromatin is located far from the NE70, we propose that similarly to some other Nups, Elys interacts with chromatin positioned both at the NE and in the nuclear interior.

a Immunostaining of squashed polytene chromosomes with anti-Elys (red) antibodies. Elys binding sites located within polytene chromosome bands, counterstained with Dapi (blue), are revealed as pink strips (marked by arrows in a chromosomal region), while Elys binding sites within inter-bands are revealed as red strips (marked by asterisks in a chromosomal region). b Screenshot from UCSC genome browser showing Nup98_nucl (pink), Elys_embryo (red), and Nup98_NPC (violet) profiles, as well as the corresponding domains or sites of enrichment (rectangles over profiles) for the representative region of chromosome 3R. The 9-state chromatin model and RefSeq genes are indicated below. The overlapped regions between Elys_embryo and Nup98_nucl sites or between Elys_embryo and Nup98_NPC sites (Elys_nucl and Elys_NPC sites, respectively) are outlined by translucent rectangles. c Venn diagram showing the degree of overlap (as a percentage of Elys_NPC or Elys_nucl site length) between Elys_NPC or Elys_nucl sites and LADs, Pc, and H3K27Ac domains.
Identification of Elys binding sites in late embryos
We aimed to separate genomic sites bound by Elys in the nuclear interior from those bound by NPC-linked Elys molecules. To this end, we employed publicly available data obtained by DamID technique71 for Nup98-interacting regions in embryonic Kc167 cells which were successfully divided into two distinct pools28, either interacting with Nup98 in the nucleoplasm (Nup98_nucl sites) or interacting with Nup98 at the NPCs (Nup98_NPC sites). Since almost all Nup98 ChIP-seq sites coincide with Elys ChIP-seq sites35, we compared Elys ChIP-seq sites in S2 cells37 with Nup98_nucl or Nup98_NPC sites in Kc167 cells (note that S2 and Kc167 are cognate cell cultures of embryonic origin). Surprisingly, while the overlap of Elys ChIP-seq sites with Nup98_nucl sites was highly non-random (P < 10−4, permutation test), the overlap of Elys ChIP-seq sites with Nup98_NPC sites was small and statistically non-significant (the overlapped regions constitute only 5.4% of total Elys peak coverage in the euchromatic chromosome arms, P ~ 1 for their occasional colocalization, permutation test). Moreover, the overlap of Nup98 or Nup93 ChIP-seq sites identified in S2 cells35,37 with Nup98_NPC sites determined in Kc167 cells was also non-significant (P ~ 1, or P = 0.2, respectively, permutation test). The absence of colocalization between binding sites of different Nups identified by ChIP-seq in S2 cells with Nup98_NPC sites determined by DamID in Kc167 may be caused either by high variability of NPC sites in various cell types or by poor detection of NPC-bound chromatin by ChIP procedure.
To examine which assumption is correct, we tried to identify Elys binding sites in S2 cells using transient transfection of these cells with DamID constructs72, but, by unknown reasons, this approach gave unreliable results. Then, by applying the DamID-seq technique adapted for transgenic flies73, we successfully generated the Elys profile in late (16–18-h) embryos (Fig. 2b). Using a three-state Hidden Markov model (HMM) algorithm, we identified ~10,600 Elys binding sites with median length 1.8 kb (Supplementary Data 3). Since late embryos represent a mixed population of diverse cell types, we reasoned that a fraction of Elys binding sites identified in late embryos might coincide with Nup98_NPC and Nup98_nucl sites in Kc167 cells. Indeed, we found that the majority of Elys peaks in embryos were colocalized either with Nup98_NPC or with Nup98_nucl peaks in Kc167 (Fig. 2b). Whole-genome analysis revealed that ~30% and ~40% out of all Elys binding sites in embryos were highly non-randomly overlapped with Nup98_nucl and Nup98_NPC sites, respectively (P < 10−4 in both cases, permutation test). Hereinafter, these Elys binding sites conservative between embryos and Kc167 cells will be referred to as Elys_nucl and Elys_NPC sites (Supplementary Data 3). We also identified a fraction of ambivalent Elys_NPC/nucl sites (constituting ~13% out of all Elys sites, Supplementary Data 3), apparently corresponding to Elys interactions with the genome in both locations. Yet, ~17% of Elys sites did not span Nup98 sites and were excluded from further analysis.
NPC-linked Elys binds numerous genomic sites located within LADs
Recently, Gozalo et al.37 have reported that 17% of Nup93 ChIP-seq sites and 22% of the shared Elys/Nup107 ChIP-seq sites in S2 cells were localized within LADs, thereby indicating peripheral positioning of a fraction of Nup-bound regions. Moreover, in some cases, positions of Nup93 ChIP-seq peaks coincided with the dips in the Lamin Dm0 (Lam) profile37. We determined the distribution of Elys_NPC and Elys_nucl sites across different types of chromatin by comparing their genome positions with that of various types of chromatin domains. More than half of Elys_NPC sites appear to locate within LADs (Fig. 2c). In agreement with the reported overlap of some Nup93 binding sites with the repressive Polycomb (Pc) domains37, 9% of Elys_NPC sites were localized within Pc domains (Fig. 2c). However, only a few Elys_NPC sites contain active, highly acetylated chromatin (Fig. 2c). Strikingly, the degree of intersection (from the total length) of Elys_NPC sites with the inactive chromatin revealed in Kc167 cells72 reaches 94%, while it was ~70% with that revealed in S2 cells74 (Supplementary Fig. 7). Contrary to Elys_NPC sites, about a half of Elys_nucl sites overlap with H3K27 acetylated regions in S2 cells (Fig. 2c), and 86% or 95% of Elys_nucl sites correspond to active chromatin types in Kc167 or S2 cells, respectively (Supplementary Fig. 7). Therefore, Elys_NPC and Elys_nucl sites are almost completely located either within inactive or within active chromatin, respectively.
To validate that many Elys_NPC sites are indeed present within LADs, we compared the Elys DamID profile in late embryos with the Lam DamID profile in Kc167 cells5. In addition, we performed DamID-seq mapping of genomic regions interacting with Lam in 16–18-h embryos and identified embryonic LADs (Supplementary Data 4) covering 55.8% of the non-repetitive genome. Visual examination of the profiles in UCSC genome browser indicates that Elys_NPC peaks, positioned within LADs or at LAD boundaries, frequently correspond to the dips in the Lam profile from both Kc167 cells and late embryos (Fig. 3a). Strikingly, the corresponding peaks were mostly absent in Elys ChIP-seq profile from S2 cells (Fig. 3a). At the whole-genome level, the peak in the averaged Elys profile, centered at Elys_NPC sites positioned within LADs (±2 kb from LAD boundaries), corresponds to the dip in the averaged Lam profile from both Kc167 cells and late embryos (although in the latter case the dip is not so perfect and pronounced; Fig. 3b). The dip is not revealed when the Lam profile is averaged around randomly chosen positions within Kc167 LADs (Supplementary Fig. 8a, left panel). Importantly, the dip in the Lam profile around Elys_NPC sites within LADs corresponds to the dip in the H3K27 acetylation profile (Fig. 3b, most right panel). Therefore, Elys_NPC sites within LADs fail to interact with the NL not because of the increased histone acetylation correlating with chromatin positioning in the nuclear interior70. Rather, these genomic regions are bound by the NPC-linked Elys, and this binding partially prevents their interactions with the NL. In this case, the presence of Elys_NPC sites within LADs reflects the inaccuracy of the two-state HMM algorithm, applied for LADs calling, which merges the neighboring LADs separated by short gaps corresponding to Elys_NPC sites.

a Screenshot from UCSC genome browser showing H3K27Ac (orange), Nup98_nucl (pink), Nup98_NPC (violet), Elys_embryo (red), Elys_S2 (yellow), Lam_embryo and Lam_Kc167 (brown) profiles, as well as the corresponding domains or sites of enrichment (rectangles over profiles) for the representative region of chromosome 2R. The 9-state chromatin model, RNA-seq in control S2 cells (from this work) and RefSeq genes are indicated below. The overlapped regions between Elys_embryo and Nup98_NPC sites (containing Elys_NPC peaks) are outlined by translucent rectangles (Elys_NPC peaks mostly coincide with the dips in Lam profiles). b, c Averaged Lam_Kc167, Lam_embryo, Elys_embryo, Elys_S2 and H3K27Ac profiles around Elys_NPC sites (b) or Elys_nucl sites (c) located within LADs (±2 kb from LAD boundaries) determined in Kc167 or in late embryos. d Screenshot from UCSC genome browser showing H3K27Ac (orange), Nup98_nucl (pink), Elys_embryo (red), and Nup98_NPC (violet) profiles, as well as the corresponding domains or sites of enrichment (rectangles over profiles) for the representative region of chromosome 2L. The 9-state chromatin model, RNA-seq in control S2 cells (from this work), and RefSeq genes are indicated below. The regions of high concordance between H3K27Ac, Nup98_nucl, and Elys_embryo profiles are outlined by translucent rectangles.
To explore whether the dips in the Lam profile are located at the same positions during development, we analyzed previously obtained Lam DamID profiles in the central brain, neurons, glia, and the fat body of Drosophila third instar larvae6. The dips in the averaged Lam profile around Elys_NPC sites located within LADs (±2 kb from LAD boundaries) were detected in all these organs/cell types, including terminally differentiated neurons (Supplementary Fig. 9). Therefore, during cell differentiation, NPC-linked Elys is bound to a similar set of genomic sites within LADs, although some variability associated with gene activation may be present.
It is noteworthy that the averaged Elys ChIP-seq profile from S2 cells37 does not display the peak at Elys_NPC sites positioned within LADs (±2 kb from their boundaries) (Fig. 3b). Moreover, the dip in the averaged Lam profile, centered at Elys ChIP-seq sites located within LADs (±2 kb from their boundaries) (Supplementary Fig. 8b), or within LADs overlapping with inactive chromatin from S2 cells (Supplementary Fig. 8a, right panel), is far less pronounced than that centered at Elys_NPC sites (Supplementary Fig. 8a, middle panel). Furthermore, in contrast to Elys_NPC sites, Elys ChIP-seq sites located within LADs (±2 kb from LAD boundaries) possess high levels of histone acetylation (Supplementary Fig. 8a, most right panel). An increased histone acetylation is also revealed around Nup93 ChIP-seq sites from S2 cells (Supplementary Fig. 8c). The latter findings imply that the majority of these sites are likely located distantly from the NE in the active chromatin of inter-LADs adjoining to LAD boundaries, or in the transition zones between LADs and inter-LADs, but not within the genuine LADs.
Supporting this notion, Elys_nucl sites positioned in the nuclear interior demonstrate negative values of the averaged Lam profile and high level of histone acetylation (Fig. 3c). High consistency between Elys_nucl profile and H3K27 acetylation is exemplified in Fig. 3d.
Thus, we suppose that weak correspondence between Elys_NPC sites determined by DamID in embryos and Elys ChIP-seq sites in S2 cells is not a consequence of a high level of polymorphism between different cell types. Rather, the ChIP procedure poorly reveals Elys_NPC sites, whereas the DamID technique readily detects them. The robust identification of Elys_NPC sites by DamID allows us to make a confident conclusion that peripheral chromatin interacts with the NE through multiple alternating sites. Some of them are attached to the NL, while others are attached to NPCs.
Both NPC-linked and nucleoplasmic Elys likely recognize A/T-rich DNA motifs
Since Elys is the Nup known to bind chromatin directly47,49,53,54,55,56, we analyzed DNA sequence motifs potentially responsible for its binding. Previously, Rasala et al.47 have shown that antibiotic distamycin A, capable of binding with A/T-rich DNA sequences, inhibits Xenopus Elys interactions with chromatin. Furthermore, using EMSA, Mehta et al.49 have revealed that two out of three non-canonical motifs in Drosophila Elys can bind A/T-rich sequences. Consistent with these results, we found that the most high-scoring motif identified by MEME75 in Elys_NPC or Elys_nucl sites is represented by poly(A) tracks (Fig. 4a). ~30% of Elys_NPC and Elys_nucl sites contain at least one 20-bp A/T-rich sequence within ±150 bp region from their centers. We also found that A/T-base content is increased in both Elys_NPC and, more notably, Elys_nucl sites compared to random genomic sites (Fig. 4b). Therefore, a significant fraction of genomic targets of both NPC-linked and nucleoplasmic Elys contains A/T-rich DNA motifs, which likely mediate Elys binding to these sites.

a Top five MEME motifs identified in Elys_NPC or Elys_nucl sites (within ±150 bp from their centers). b Average A/T profile (percentage of A/T within 20-bp bins) around Elys_NPC, Elys_nucl or random genomic sites. Dashed lines delimit the median lengths of corresponding sites.
Binding of inactive chromatin with NPC-linked Elys maintains peripheral chromatin positioning at the NE
DamID technique detects genomic sites, which are either bound by or in contact with the protein of interest76. To confirm that Elys, as a component of NPCs, binds peripheral chromatin, but not just contacts it, we assayed by FISH the radial position of three loci, carrying Elys_NPC sites within LADs (Supplementary Fig. 10), after Elys-KD in S2 cells (Supplementary Fig. 1a, b). These loci were previously shown to localize mostly at the NE and to lose peripheral localization upon Lam-KD in S2 cells9,19. We found that the lack of Elys leads to a notable displacement of these loci from the NE to the nuclear interior (Fig. 5a–d; Supplementary Fig. 11; Supplementary Data 5), and this shift resembles that observed upon Lam-KD (Fig. 5b). Moreover, simultaneous depletion of Elys and Lamin causes a more noticeable removal of the locus from the NE (Fig. 5b).

a Confocal images of FISH signals (red) detected by the probe for 60D region in nuclei stained with anti-Lamin (green) and anti-LBR (violet) antibodies in Elys-KD or control S2 cells. Scale bar 1 µm. b–d Violin-plots showing distribution of radial-normalized distances between 60D (b), 22A (c), or 36C (d) FISH signals and the NE in control (blue; n = 141, or n = 151, or n = 125 for 60D, 22A and 36C probes, respectively), Elys-KD (red; n = 137, or n = 150, or n = 125 for 60D, 22A and 36C probes, respectively), Lam-KD (brown; n = 100), and simultaneous Elys-KD and Lam-KD (dark brown; n = 100) S2 cells. e, f Averaged fluorescence intensity profiles along the nuclear diameter in Elys-KD (n = 50) and control (n = 50) S2 cells immunostained with anti-histone H4 and anti-LBR antibodies (e), or in Elys-KD (n = 170) and control (n = 170) S2 cells immunostained with anti-H3K27Ac and anti-Lamin antibodies (f). To estimate P-values, two fluorescence intensity distributions (control vs Elys-KD) on the interval of radial positions from 0 (position of the NE identified by the peak of LBR or Lamin Dm0 staining) to 0.15R, were compared. This interval is delimited by the dashed lines. g Violin-plots showing the distribution of volumes of control (n = 250) and Elys-KD (n = 250) S2 nuclei manually outlined by the NL and further automatically reconstructed in IMARIS. White dots represent median values, upper and lower ends of black bars show the upper and lower quartiles, the ends of the thin line indicate the maximum and minimum values. h A schematic illustrating mechanisms of peripheral chromatin attachment to the NE. All measurements in (b–g) were performed in two biological replicates. P-values in (b–g) were estimated in an M-W U-test.
Next, we examined whether the removal of peripheral chromatin from the NE upon Elys-KD is a general phenomenon, or it affects only particular loci. To this end, methanol-fixed Elys-KD and control S2 cells were stained with anti-histone H4 antibodies to visualize total chromatin and with anti-LBR antibodies to visualize the NE. The averaged fluorescence intensity of histone H4, measured along the diameter of nuclei and normalized on the diameter of those nuclei, shows that total chromatin is localized slightly more distantly from the NE upon Elys-KD, as compared to control cells (Fig. 5e; Supplementary Fig. 12a; Supplementary Data 6). However, staining of nuclei with anti-H3K27Ac antibodies indicates that active chromatin, located further away from the NE (Supplementary Fig. 12b), occupies nearly the same radial position in control and Elys-KD cells (Fig. 5f; Supplementary Data 6). Therefore, the lack of Elys results in the inactive peripheral chromatin redistribution towards the nuclear interior.
Interestingly, volumes of nuclei were not significantly diminished upon Elys-KD compared to control S2 cells (Fig. 5g; Supplementary Data 7), similar to what was observed upon Lam-KD in S2 cells19.
Collectively, our findings indicate that, in interphase nuclei, inactive peripheral chromatin is attached not only to the NL but also, through Elys, to the NPCs. Without each type of multiple anchorages, chromatin is slightly shifted from the nuclear periphery to the nuclear interior (Fig. 5h).
Elys-KD in S2 cells results in the decompactization of the NE-attached TADs
Previously, conflicting data were obtained regarding Elys influence on chromatin compaction. Artificial targeting of Elys via Sec13 to a particular site on polytene chromosomes was found to cause local chromatin decondensation36. However, ATAC-seq analysis upon Elys depletion in S2 cells did not reveal notable changes in chromatin accessibility at Elys ChIP-seq peaks or at all ATAC peaks60. To resolve this issue, we generated high-throughput chromosome conformation capture (Hi-C)77 heatmaps at 4-kb resolution containing information about spatial chromatin organization in control and Elys-KD S2 cells (Supplementary Fig. 1c). TADs in Drosophila cells are known to represent units of chromatin folding with length from tens to hundreds of kb (Fig. 6a). Using Armatus software78, ~3600 TADs in control and ~3500 TADs in Elys-KD cells with 24-kb median length were identified (Fig. 6b; Supplementary Data 8). Next, we calculated the average contact frequency (ACF, see “Methods” section) for each TAD that has exactly the same positions of boundaries in control and Elys-KD cells (~57% of all control TADs). We reasoned that changes in ACF values for each TAD upon Elys-KD would reflect changes in chromatin packaging density within that TAD. For this analysis, TADs with the same boundaries were divided into three groups depending on the proportion of coverage with both active chromatin (states 1 and 2 according to the 9-state model in S2 cells74) and LADs5. For each TAD, the Jaccard coefficient based on these two metrics was calculated. After Elys-KD, TADs from group A, mostly composed of active chromatin, became slightly more compact (i.e., their ACF values increased), whereas TADs mostly corresponding to LADs (group C), on the contrary, became less densely organized (Fig. 6c). The same trends in chromatin compaction were detected when TADs were divided into four groups based on quartiles of total gene expression within them (Supplementary Fig. 13a). Since the X chromosome in S2 cells is enriched in H4K16 acetylated histones due to the dosage compensation79, we analyzed alterations in TAD density for the X chromosome separately and revealed the same ACF changes as for all TADs (Supplementary Fig. 13b). We conclude that the loss of Elys leads to more compact active TADs and less compact inactive TADs.

a A schematic showing TADs and compartments. b Hi-C heatmap for the representative region of 3L chromosome in Elys-KD (above the diagonal) and control (below the diagonal) cells. TADs (demarcated by black dotted lines), RNA-seq profile in control cells (in RPM, blue peaks), RefSeq genes, and chromatin annotation in S2 cells according to the 9-state model are indicated below the Hi-C map. c Box-plots showing log2FC of ACF (Elys-KD/control) for TADs divided into three groups (A, B, C). Black lines represent median values, upper and lower ends of box-plot show the upper and lower quartiles, the whiskers indicate the maximum and minimum values, and individual points show the outliers. P-values were estimated in a Wilcoxon signed-rank test. d Saddle-plot showing subtraction of aggregated contact frequency (Elys-KD minus control) log2FC(observed/expected) for autosomes. Dashed lines indicate areas that were analyzed on (e). e Box-plots showing subtraction of aggregated contact frequency (Elys-KD minus control) log2FC(observed/expected) for autosomes within active (AA), inactive (BB), and between active and inactive (AB) chromatin compartments. Black lines represent median values, upper and lower ends of box-plot show the upper and lower quartiles, the whiskers indicate the maximum and minimum values, and individual points show the outliers. P-values were estimated in a Wilcoxon signed-rank test. f, g Heatmaps showing subtraction of ACF (Elys-KD minus control) log2FC(observed/expected) around Elys_NPC sites located in the inactive chromatin (states 6-9) (f) or around Elys_nucl sites containing > 50% of active chromatin (states 1 and 2) (g). h, i Averaged ISm profiles around Elys_NPC sites located within inactive (states 6–9) chromatin (h) or around Elys_nucl sites (i) in control (blue curve) and Elys-KD (red curve) cells. ISm profiles were calculated for 2-kb resolution Hi-C heatmaps. Only contacts between regions located 2–8 kb away from both sides of the central bin were considered. j A schematic showing how the attachment to NPCs may facilitate contacts between regions located on both sides from the Elys_NPC site.
Next, we analyzed distant interactions within and between active (A) and inactive (B) chromatin compartments77 (Fig. 6a). As was reported earlier19, contact frequency in S2 cells is higher than expected only for the A compartment (Supplementary Fig. 13c). Similar to the impact of Lam-KD19, Elys-KD results in enhanced or relaxed distant interactions within inactive or active autosomal compartments, respectively (Fig. 6d, e). The same trend is observed for the inactive compartment of the X chromosome (Supplementary Fig. 13d). Yet, the enhanced intermingling of A and B compartments reported upon Lam-KD, is not observed upon Elys-KD.
To track the influence of Elys on local chromatin compaction, we built averaged Hi-C heatmaps around Elys_NPC and Elys_nucl sites. Upon Elys-KD, ACF in Elys_NPC sites located within inactive chromatin (states 6–9)74 decreases stronger than in adjacent inactive regions (P = 0.002, M-W U-test) (Fig. 6f). Moreover, chromatin attachment to NPCs facilitates contacts between regions located 2–8 kb away from both ends of Elys_NPC sites, as determined by the peak on the curve of modified insulation score (ISm, see “Methods” section for details) reflecting contact frequency between these regions (Fig. 6h, j). Such contacts become weaker upon Elys-KD (P = 9.1 × 10−7, Wilcoxon signed-rank test). These results support the model according to which Elys_NPC sites are attached to NPCs.
At the same time, ACF within Elys_nucl sites containing ≥50% of active chromatin (states 1 and 2)74 as well as within adjacent active regions is increased upon Elys-KD (Fig. 6g). Elys occupancy of Elys_nucl sites, which coincides with a fraction of TAD boundaries80, enhances spatial isolation of neighboring regions (P = 1.5 × 10−61, Wilcoxon signed-rank test) (Fig. 6i). Therefore, Elys binding with active chromatin leads to its slight decompactization, which may be mediated by the PBAP complex, shown to be associated with Elys36.
Elys is enriched at the 5′- and 3′-ends of genes
Nups in mammals and Drosophila were found to interact with enhancers or super-enhancers (the latter represent enhancer clusters)81, marked by increased H3K27 acetylation34,35. Using the modified DamID technique, super-enhancers in mammals were recently shown to associate with the NPC-linked Nups82. It remained unclear whether enhancers in Drosophila are also associated with NPCs. We examined whether Elys_nucl or Elys_NPC sites are colocalized with strong active enhancers which were identified in S2 cells by self-transcribing active regulatory region sequencing (STARR-seq)83. We found highly statistically significant overlap only between STARR-seq enhancers and Elys_nucl sites (Fig. 7a; 42% of enhancers overlap with Elys_nucl sites, P < 10−4 for their occasional colocalization, permutation test), but not between STARR-seq enhancers and Elys_NPC sites (5% of enhancers overlap with Elys_NPC sites, P ~ 1 for their occasional colocalization, permutation test). However, there was still a possibility that enhancers induced by external stimuli may be attached to the NPCs. To test this idea, we compared Elys_nucl and Elys_NPC sites with STARR-seq enhancers activated by ecdysone treatment in S2 cells84. Similar to active enhancers, the ecdysone-responsive enhancers strongly colocalize with Elys_nucl sites, but not with Elys_NPC sites (Fig. 7a; P < 10−4 and P ~ 1, respectively, for their occasional colocalization, permutation test). Therefore, both active and inducible enhancers in Drosophila S2 cells are mostly bound by the nucleoplasmic Elys fraction.

a Screenshot from the UCSC genome browser demonstrating that active or ecdysone-responsive STARR-seq enhancers mainly coincide with Elys_nucl sites. Nup98_nucl (pink), Elys_embryo (red), and Nup98_NPC (violet) profiles are shown, as well as the corresponding sites of enrichment (rectangles over profiles) and active or ecdysone-responsive STARR-seq enhancers in S2 cells (black rectangles) for the representative region of chromosome 2R. RNA-seq in control S2 cells (this work) and RefSeq genes are indicated below. b Pie chart showing the percentage of genes overlapping with indicated Elys sites. c Screenshot from UCSC genome browser with an example of the gene containing Elys_nucl, Elys_NPC, and Elys_NPC/nucl sites. H3K27Ac (orange), Nup98_nucl (pink), Nup98_NPC (violet), Elys_embryo (red), and Lam_Kc167 profiles are shown, as well as the corresponding domains or sites of enrichment (rectangles over profiles) and ecdysone-responsive STARR-seq enhancers in S2 cells (black rectangles). RNA-seq in control S2 cells (this work) and RefSeq genes are indicated below. Regions corresponding to Elys_NPC, Elys_nucl, and Elys_NPC/nucl sites are outlined by violet, pink, or merged color translucent rectangles, respectively. d Averaged Elys profiles over metagene carrying sites of Elys_NPC (violet), Elys_nucl (pink), their combinations (wine-colored), or for randomly reshuffled genes (gray). e Box-plots showing RNA-seq expression (in TPM) in control S2 cells for genes carrying sites of Elys_NPC (violet), Elys_nucl (pink), or their combinations (wine-colored). f Box-plots showing RNA-seq expression FC (Elys-KD/control) ranked by the expression level in control S2 cells. g Box-plots showing RNA-seq expression FC (Elys-KD/control) for genes carrying sites of Elys_NPC (violet), Elys_nucl (pink), or their combinations (wine-colored). P-values in (f, g) were estimated in a Wilcoxon signed-rank test. See Fig. 6c legend for the description of box-plot elements (outliers were omitted).
17.1% and 6.8% of genes overlap solely with Elys_nucl or Elys_NPC sites (Fig. 7b; Supplementary Data 9), thus indicating the prevalent positioning of these genes in the nuclear interior or at the NE, respectively. However, 8.2% of genes simultaneously contain various combinations of these sites (including Elys_NPC/nucl sites) (Fig. 7b; Supplementary Data 9). The ecdysone-inducible gene Eip63E (Fig. 7c) belongs to the latter group of genes. They are attached to NPCs by internal sites (which in some cases include enhancers) and are looped out from NPCs by other regions. Such an organization may be associated with the transcriptional memory, inherent to ecdysone-inducible genes35.
Elys_nucl sites are enriched at gene promoters compared to Elys_NPC sites (Supplementary Fig. 14). The metagene profile demonstrates that Elys occupies mostly 5′- and 3′-ends of genes (Fig. 7d). To examine possible mechanism(s) of this enrichment, we built averaged metagene profiles for A/T content, H3K27 acetylation and binding of GAGA factor (GAF). The peak of Elys binding at the 3′-ends of genes exactly coincides with peaks of A/T content and H3K27 acetylation (Supplementary Fig. 15a, b). Thus, Elys enrichment at the 3′-ends of genes may be mediated by either of these features. However, Elys peak at the transcription start sites (TSSs) coincides with the peak of GAF, but not with peaks of A/T content or H3K27 acetylation (Supplementary Fig. 15c). Since GAF may guide the PBAP complex to TSSs59,60, Elys that was shown to associate with the PBAP36, may be recruited to TSSs via this mechanism. Yet, we did not find that Elys and GAF are co-immunoprecipitated from S2 cells (Supplementary Fig. 15d).
Elys-KD in S2 cells causes derepression of genes in LADs
To explore the influence of Elys on gene expression, we performed poly(A+) RNA-seq analysis in the control and Elys-KD S2 cells (Supplementary Fig. 1d; Supplementary Data 9). Unexpectedly, changes in gene expression upon Elys-KD were rather weak. We identified only 175 differentially expressed genes (i.e., genes with fold change (FC) of expression > 1.5 and false discovery rate < 0.05). 131 genes were up-regulated and 44 genes were down-regulated (Supplementary Data 9). Our further analysis was focused on genes bound by NPC-linked or nucleoplasmic Elys. Consistent with low and high levels of histone acetylation at Elys_NPC and Elys_nucl (or Elys_NPC/nucl) sites, respectively, genes carrying only Elys_NPC sites were mostly “silent”, while genes carrying only Elys_nucl sites or various Elys site combinations were actively expressed (Fig. 7e). Elys-KD resulted in slightly increased expression of “silent” and weakly expressed genes, while most active genes did not notably change their expression (Fig. 7f). Accordingly, genes carrying only Elys_NPC sites were slightly up-regulated, while genes carrying Elys_nucl sites or various Elys site combinations were barely affected upon Elys-KD (Fig. 7g). We note that the majority of “silent” or weakly expressed genes, that were up-regulated upon Elys-KD and either contained or not contained Elys_NPC sites, were localized within LADs. Together with our previous results showing that the same effect on gene expression was detected upon Lam-KD19, these data support the idea that the upregulation of weakly expressed genes upon Elys-KD may be caused by the loss of their interactions with the NL. We conclude that Elys-KD faintly affects gene expression.
Discussion
Here we show that Drosophila S2 cells is an appropriate model to analyze the influence of Elys on overall genome architecture, since, unlike in other organisms and some other Drosophila cell types42,43,44,45,46,49, depletion of Elys in S2 cells does not result in a notable reorganization of NE components, including NPCs (Fig. 1a–c), and does not cause the enhanced apoptosis (Supplementary Fig. 3). Moreover, upon Elys depletion in S2 cells, post-mitotic NPC incorporation in the NE occurs prior to telophase (Fig. 1h). Then, what might be a mechanism of rapid post-mitotic NPC incorporation in the absence of Elys? There is an Elys-independent pathway for insertion of NPCs in the NE during interphase, but it is rather slow, at least in mammals48. However, in early Drosophila embryos, the pre-assembled NPCs, associated with membrane stacks of the endoplasmic reticulum (with the annulate lamellae)85, are incorporated into the reforming NE shortly after mitosis86. Moreover, a similar mechanism operates in various mammalian cell lines as well as in the differentiated Drosophila cells87. We hypothesize that, upon Elys-KD, embryonic S2 cells may restore NPCs at the NE at the end of mitosis by employing the same mechanism.
In this work, using the DamID technique, we identified Elys binding sites in late embryos. By comparing them with genomic regions interacting with Nup98 either in the nucleoplasm or at the NPCs28, we identified conservative Elys_NPC and Elys_nucl sites. Elys_NPC sites are largely preserved during cell differentiation, contain non-acetylated nucleosomes, and are located within LADs or, less frequently, within Pc domains (Figs. 2c and 3a, b). On the contrary, Elys_nucl sites contain acetylated chromatin (Figs. 2c, 3c, d) and are colocalized with active gene promoters (Supplementary Fig. 14) and enhancers. Interestingly, both Elys_NPC and Elys_nucl sites bear A/T-rich DNA motifs (Fig. 4) likely determining their recognition by Elys. The prevalence of sequence-specific mode of Elys binding is supported by the fact that the majority (83%) of Elys binding sites in late embryos are shared with Nup98 binding sites in Kc167 cells.
We hypothesize that, in anaphase, Elys recognizes and binds the A/T-rich sequences within decondensing chromatin and maintains binding with them during interphase. In this scenario, whether a particular Elys molecule appears at the NE (within NPC) or in the nucleoplasm depends on whether its A/T-rich binding site is embedded in the non-acetylated or acetylated chromatin, respectively. In the acetylated chromatin, Elys binding with A/T-rich sequences may promote Elys spreading on the neighboring acetylated regions due to Elys ability to associate with the acidic patch of H2A/H2B histone dimer55,56. This ability may explain the reported correlation between Elys and H3K27 acetylation profiles35 (Fig. 3d). Yet, Elys association with the H2A/H2B acidic patch is likely to be blocked in non-acetylated nucleosomal arrays by the non-acetylated N-terminal tail of histone H4 competing for binding with the same interface88. An additional level of complexity in Elys interactions with chromatin is mediated by Elys targeting to some genomic sites by its protein partners. In particular, the colocalization of Elys and GAF peaks at gene promoters (Supplementary Fig. 15c) may suggest the existence of an indirect mechanism for the recruitment of Elys to chromatin via GAF/PBAP complex36,58,59.
We reason that if Elys links chromatin with NPCs upon mitotic exit and interacts with chromatin in interphase nuclei, it could be required to keep peripheral chromatin attached to the NE during interphase. Indeed, using FISH and chromatin immunostaining we found that several loci analyzed as well as peripheral chromatin as a whole were relocated from the NE to the nuclear interior upon Elys-KD in S2 cells (Fig. 5). This effect resembles the one observed upon Lam-KD in S2 cells19. Our results indicate that NPC-linked Elys is not simply in contact with chromatin, but is bound to it. Moreover, NPC-linked Elys maintains localization of peripheral chromatin at the NE. Elys binding to peripheral chromatin is also supported by the dips in Lam profiles at Elys_NPC sites (Fig. 3a, b), as well as by the peculiar three-dimensional organization of these sites (Fig. 6f, h, j). Taken together, our findings suggest that peripheral chromatin is simultaneously bound both to the NL and, via Elys, to the NPCs (Fig. 5h). As far as we know, our results provide the first evidence that peripheral chromatin in a multicellular organism is maintained at the NE not only due to its interactions with the NL, but also due to its interactions with NPCs.
Apart from the maintenance of genome architecture, NPC-linked Elys binds silent, or weakly expressed genes, whereas nucleoplasmic Elys binds promoters and enhancers of actively expressed genes. Our RNA-seq analysis shows that Elys-KD in S2 cells only weakly affects gene expression. Low-expressed genes become mainly up-regulated, while transcription of active genes is mostly unaltered (Fig. 7e–g). The similarity of effects on gene expression upon Elys-KD and Lam-KD19 implies that the derepression of low-expressed genes upon Elys-KD may be caused by the detachment of genes from the NL, but not by the direct impact of Elys on their transcription. In support of this notion, we found that TADs attached to the NL become decompacted upon both Elys-KD (Fig. 6c) and Lam-KD19. The compactization of TADs, associated with the NL and NPCs, may be caused, in part, by mechanical forces, since our computer modeling has shown that chromatin attachment to the NE is sufficient to make it more condensed19. At the same time, active TADs become more compact after Elys-KD (Fig. 6c), while transcription within them is mostly unchanged (Fig. 7e–g). The decompactization of active TADs may be mediated by the PBAP chromatin remodeling complex associated with Elys36. However, this provides only the potential for active genes to be activated. Of note, Elys binding with promoters and enhancers may be related to the transcriptional memory, i.e., more rapid and strong induction of gene transcription upon repeated treatment of cells with external activation stimuli. In metazoans, this phenomenon was associated with several Nups including Nup98 and Nup15335,89,90.
In summary, comprehensive molecular analysis including DamID, Hi-C, RNA-seq, and FISH in Drosophila cells allowed us to draw several important conclusions: (i) Peripheral chromatin is attached not only to the NL but also, via Elys, to NPCs. Loss of Elys results in bulk chromatin displacement from the NE to the nuclear interior. (ii) Like chromatin associated with the NL, chromatin bound to NPC-linked Elys is mostly inactive. This observation refutes a model postulating that chromatin associated with NPCs constitutes islets of active genes within the repressed chromatin layer lining the NE. (iii) Nucleoplasmic Elys interacts preferentially with active chromatin of enhancers and gene promoters. These interactions lead to local chromatin decondensation possibly caused by Elys recruitment of the PBAP chromatin remodeling complex. However, Elys depletion only weakly affects gene expression. (iv) Since nucleoplasmic and NPC-linked Elys fractions apparently recognize the same AT-rich sequence motifs, their preferential association with active or inactive chromatin may simply reflect chromatin nuclear positioning established by other mechanisms.
Methods
Plasmid construction
To generate pUAST-attB-hsp70-loxP-stop-loxP-Dam-Elys construct, the whole Elys ORF, lacking a start codon, was initially assembled in the pBluescript II vector (Stratagene). To this end, the ∼0.32-kb 5′-fragment and the ∼1.0-kb 3′-fragment of Elys ORF were amplified by PCR from cDNA LD14710 (Drosophila Genomics Resource Center) using primers indicated in Supplementary Data 10. Then, NotI- and NheI-digested 5′-fragment, HindIII- and ApaI-digested 3′-fragment and ∼5.1-kb NheI- and HindIII-digested central fragment of Elys ORF were combined together by cloning into the HindIII and ApaI sites of pBluescript II. Next, the whole Elys ORF was excised by NotI and SpeI sites and cloned by NotI and XbaI sites in pUAST-attB-hsp70-loxP-stop-loxP-Dam-Lam construct91 instead of Lamin Dm0 ORF giving Elys ORF in frame with Dam ORF. The correct nucleotide sequence of the resultant pUAST-attB-hsp70-loxP-stop-loxP-Dam-Elys construct was verified by partial sequencing.
Fly stocks and handling
Fly stocks were maintained under standard conditions at 25 °C. Transgenic strains carrying pUAST-attB-hsp70-loxP-stop-loxP-Dam-Elys were generated by ϕC31-mediated site-specific integration at the attP40 site on the chromosome 2 in the y, w; P{y[+t7.7] = CaryP}attP40; M{3xP3-RFP.attP}ZH-86Fb; M{vas-int.B}ZH-102D line92 as previously described93. Transgenic strains carrying pUAST-attB-hsp70-loxP-stop-loxP-Dam, pUAST-attB-hsp70-loxP-stop-loxP-Dam-Lam, and pUAST-attB-nos-Cre constructs at the same genomic site were described previously65,73,91.
DamID-seq procedure in late embryos
To perform DamID in late embryos, we crossed females expressing Cre-recombinase in the oocytes and zygotes (#766 line from the Bloomington Drosophila Stock Center which was devoid of balancers) with the males carrying pUAST-attB-hsp70-loxP-stop-loxP-Dam or pUAST-attB-hsp70-loxP-stop-loxP-Dam-Lam or pUAST-attB-hsp70-loxP-stop-loxP-Elys constructs. These constructs contain stop-cassette, flanked by loxP sites, which separates basal hsp70 promoter from Dam, Dam-Lam, or Dam-Elys ORFs73. An excision of stop-cassette in early embryos results in Dam-methylation of corresponding genomic sites in late embryos. Approximately two hundred 16–18-h embryos from these crosses (in two biological replicates) were collected, washed with H2O, dechorionated with 2.5% hypochlorite for 3 min, and again washed three times with H2O. Further isolation of genomic DNA, amplification of Dam-methylated genomic fragments, and their subsequent sequencing were performed according to ref. 94. 16 (for Dam-Lam) or 18 (for Dam and Dam-Elys) cycles of PCR amplification (1 min at 94 °C, 1 min at 65 °C, 2 min at 68 °C) was applied for DNA samples. Sequencing on Illumina HiSeq 2500 was performed in the Evrogen facility (www.evrogen.ru) resulting in 27–30 million 100-nt single-end reads per sample (Supplementary Data 11).
Immunostaining of polytene chromosomes
Polytene chromosomes staining was performed according to ref. 95 with some modifications. Chromosomes were stained with anti-Elys antibodies65 (1:400) diluted in PBT containing 0.3% Triton X-100 and 3% Normal Goat Serum (Invitrogen) and counterstained with Dapi. As the secondary, Alexa Fluor 546-conjugated goat anti-rabbit IgG (1:200; Invitrogen) was used. Image capture was performed with a confocal LSM 510 laser scanning microscope (Zeiss).
Cell culture maintenance and RNAi
Drosophila melanogaster S2 cell line (from IMG collection) was grown at 25 °C in Schneider’s Drosophila Medium (Gibco) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco), 50 units/ml penicillin, and 50 µg/ml streptomycin. For RNAi treatment of S2 cells, dsRNAs against LacZ (control), Lam, or Elys were prepared as was described previously9 using primers indicated in Supplementary Data 10. Cells were treated with dsRNAs during four days (for FISH experiments with 60D probe), or during three days with an additional treatment for two days (for all other experiments) using previously described protocol96.
Western-blot analysis
Proteins were extracted with 8 M urea, 0.1 M Tris-HCl, pH 7.0, 1% SDS, fractionated by SDS-PAGE (12% acrylamide gel) and transferred to a PVDF membrane (Immobilon-P, Millipore). Blots were developed using alkaline phosphatase-conjugated secondary antibodies (Sigma) and the Immun-Star AP detection system (Bio-Rad). The following antibodies were used for detection: mouse monoclonal anti-Lamin97 (1:2000; ADL67), mouse monoclonal anti-beta Actin (1:5000; ab8224, Abcam), rabbit polyclonal anti-Nup10798 (1:5000), rabbit polyclonal anti-Elys65 (1:3000).
Immunostaining of S2 cells
S2 cells in the growth phase were collected and rinsed two times in PBS. Cells were fixed in 4% formaldehyde (in PBT) for 25 min at room temperature. Fixation was stopped by incubation with 0.25 M glycine (Sigma-Aldrich) for 5 min. Further immunostaining procedure was performed as previously described6. Confocal images were captured using LSM 510 or LSM 710 laser scanning microscope (Zeiss). As the primary, mouse monoclonal anti-Lamin97 (ADL84, 1:500), guinea pig polyclonal anti-LBR99 (1:1000), mouse monoclonal Mab414 (1:300; Abcam ab24609), mouse polyclonal anti-Nup153 (1:100; a gift from D. Kopytova), chicken polyclonal anti-CenpA100 (1:600), rabbit polyclonal α-Tubulin (1:2000; Abcam ab18251) and rabbit polyclonal anti-Elys65 (1:1000) antibodies were used. As the secondary, Alexa Fluor 488-conjugated goat anti-rabbit IgG (Invitrogen), or Alexa Fluor 488-conjugated or 633-conjugated goat anti-mouse IgG (Invitrogen), or Alexa Fluor 633-conjugated goat anti-chicken IgG (Invitrogen) antibodies were used.
TUNEL assay
TUNEL assay was performed using Click-iT Plus TUNEL kit (C10619, Invitrogen). After seeding on the coverslip, fixation in 4% formaldehyde (in PBT) for 25 min at room temperature, stopping the reaction with 0.25 M glycine (Sigma-Aldrich) for 5 min and performing Click-iT Plus reaction according to manufacturer’s instructions, control and Elys-KD S2 cells were immunostained with mouse monoclonal anti-Lamin97 (ADL84, 1:500) and rabbit polyclonal anti-Elys65 (1:1000) antibodies. Confocal images were captured using LSM 710 laser scanning microscope (Zeiss). The percentage of apoptotic cells in control or Elys-KD cells was counted for 10 images per replicate in three biological replicates (Supplementary Data 2).
Flow cytometry
Approximately 1.5 × 106 S2 cells were collected, rinsed twice in PBS, fixed with 70% cold ethanol for 30 min at 4 °C and washed twice in PBS. Cells were incubated in 50 µl of PBS containing 100 µg/ml RNase A for 30 min at 37 °C. Then, cells were incubated in 250 µl of PBS containing 40 µg/ml propidium iodide (Absin) for 10 min at room temperature. After that, the ratio of control and Elys-KD cells in different phases of the cell cycle was analyzed for ~104 single cells (in two biological replicates) using BD Accuri™ C6 Plus Flow Cytometer which has been calibrated on the untreated S2 cells. The gating strategy is presented in Supplementary Fig. 5a.
DNA FISH
Hybridization probes for the 22A and 36C regions were prepared using long-range PCR with primers indicated in Supplementary Data 10. Cosmid clone k99 was used as a hybridization probe for the 60D region. FISH with S2 cells was performed as was described previously19. As the primary antibodies, we used guinea pig polyclonal anti-LBR99 (1:1000), mouse monoclonal anti-Lamin97 (ADL84, 1:500), rabbit polyclonal anti-Elys65 (1:1000), sheep polyclonal anti-DIG-FITC (1:500, Roche). As the secondary antibodies we used Alexa Fluor 633-conjugated goat anti-guinea pig IgG (Invitrogen), Alexa Fluor 488-conjugated or 633-conjugated goat anti-mouse IgG (Invitrogen), Alexa Fluor 488-conjugated goat anti-rabbit IgG (Invitrogen), Alexa Fluor 488-conjugated goat anti-FITC IgG (Invitrogen).
RNA FISH with oligo(dT) probe
S2 cells were fixed with 3.7% formaldehyde in PBS for 10 min. Fixation was stopped by incubation with 0.25 M glycine (Sigma-Aldrich) for 5 min. Next, cells were washed in PBS, permeabilized for 10 min with PBS containing 0.5% Triton X-100, and incubated for 15 min at 37 °C in prehybridization buffer (2× SSC, 20% formamide, 0.2% BSA, 1 mg/ml tRNA). Hybridization was performed overnight at 37 °C in the prehybridization buffer supplemented with 1 µM oligo(dT)40-Cy5. The cells were washed twice for 5 min with 2× SSC, 20% formamide at 42 °C, once with 2× SSC at 42 °C, once with 1× SSC, and once with PBS at room temperature. Next, cells were immunostained with mouse monoclonal anti-Lamin97 (ADL84, 1:500) and rabbit polyclonal anti-Elys65 (1:1000) antibodies. Confocal images were captured using LSM 710 laser scanning microscope (Zeiss).
Measuring distances from FISH signals to the NE
Three-dimensional image stacks were recorded with a confocal LSM 510 or LSM 710 laser scanning microscope (Zeiss). Optical sections at 0.35–0.4 μm intervals along the Z-axis were captured. Images were processed and analyzed using IMARIS 7.4.2 software (Bitplane AG) with a blind experimental setup. Images were thresholded to eliminate non-specific backgrounds. The distances between signals and NE were counted as previously described9. Briefly, NL stained by anti-LBR antibodies (for FISH with 60D and 22A probes) or by anti-Lamin antibodies (for FISH with 22A and 36C probes) was manually outlined by its middle in each plane of the Z-stack, before automatic reconstruction of the nuclear surface and calculation of nuclear volume (Supplementary Data 7) by IMARIS. One measurement point was positioned in the optical section with the brightest FISH signal, at its visually determined center, and another one was placed on the reconstructed nuclear surface at the point of its earliest intersection with the progressively growing sphere from the first measurement point. The distance between the measurement points (the shortest distance between the center of the FISH signal and the middle of the NL) was measured for each nucleus. Data were obtained in two or three replicates with 35-100 FISH signals per replicate (Supplementary Data 5). Distances were normalized on the nuclei radii, which were calculated from the volumes of reconstructed nuclei on the assumption that they have the spherical form.
Chromatin visualization by histone H4 or H3K27Ac
Elys-KD or control S2 cells were seeded on coverslips for 30 min. After rinsing with PBS, cells were fixed in 100% methanol for 5 min at room temperature, rinsed with PBS three times, and blocked with PBTX (PBS with 0.1% Tween-20 and 0.3% Triton X-100) containing 3% normal goat serum (Invitrogen) for 1 h at room temperature. The remaining immunostaining procedure was performed as previously described65. Mouse monoclonal anti-histone H4 (1:200; Abcam ab31830), rabbit polyclonal anti-H3K27Ac (1:100; Abcam ab4729), mouse monoclonal anti-Lamin97 (ADL84, 1:150), rabbit polyclonal anti-Elys65 (1:3000) and guinea pig polyclonal anti-LBR99 (1:1000) antibodies were applied. As the secondary, Alexa Fluor 546-conjugated goat anti-rabbit IgG (Invitrogen) or Alexa Fluor 488-conjugated goat anti-mouse IgG (Invitrogen), or Alexa Fluor 633-conjugated goat anti-guinea pig IgG (Invitrogen) antibodies were applied.
ImageJ quantification of chromatin and NE components distribution in the nucleus
Using ImageJ, fluorescence intensities of histone H4, H3K27Ac, Lamin, and LBR staining across the nucleus diameter of the equatorial focal plane of nuclei from Elys-KD or control S2 cells were extracted. Individual profiles were first normalized on the average intensity, then on the diameter of the nucleus (delimited by peaks of LBR, or Lamin fluorescence) and further aligned to determine the averaged profile. Data were obtained for 50 or 170 nuclei from Elys-KD or control S2 cells (in two biological replicates, 25–100 nuclei per replicate) (Supplementary Data 6). For ImageJ quantification of fluorescence intensity of Mab414, Nup153, Lamin, or LBR staining at the NE, the values of fluorescence across the NE were extracted, normalized on the average Dapi staining within the nucleus, and averaged. 75-80 nuclei (in two biological replicates, 35–40 nuclei per replicate) for Mab414 and Lamin staining, or 80 nuclei (in two biological replicates, 40 nuclei per replicate) for Nup153 and LBR staining were analyzed.
Co-immunoprecipitation
Approximately 107 S2 cells per sample were washed in PBS and homogenized in lysis buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1% NP-40 and Complete Protease Inhibitor Cocktail (Roche)). The lysate was incubated for 20 min on ice prior to clearing by centrifugation at 16,000 × g for 10 min at 4 °C. Inputs were taken from the supernatant. 5 μl of rabbit polyclonal anti-Nup10798, 2 μl of rabbit polyclonal anti-Elys65, or 4 μl of rabbit polyclonal anti-GAF101 antibodies per sample were immobilized on Protein G Dynabeads (Thermo Fisher Scientific), incubated with lysates for 90 min at 25 °C, after that beads were washed three times with washing buffer (PBS, 0.1% Tween-20 and 0.3% Triton X-100). The bound proteins were eluted by heating at 95 °C for 10 min in the loading buffer containing SDS and 4 M urea. Control immunoprecipitation experiments using rabbit non-immune serum were performed in parallel.
Hi-C in S2 cells upon Elys-KD
Hi-C experiments were performed in two biological replicates as described previously102, with minor modifications to the fixation and lysis steps. ~107 control and Elys-KD S2 cells per replicate were fixed in PBS containing 2% formaldehyde for 10 min with occasional mixing. The reaction was quenched by the addition of 2 M glycine to a final concentration of 125 mM. Cells were lysed in 1.5 ml isotonic buffer [50 mM Tris−HCl pH 8.0, 150 mM NaCl, 0.5% (v/v) NP-40 substitute, 1% (v/v) Triton X-100, 1× Halt™ Protease Inhibitor Cocktail (Thermo Scientific)] on ice for 15 min. Cells were pelleted by centrifugation at 2500 × g for 5 min, resuspended in 100 μl DpnII buffer (New England Biolabs), and pelleted again. All downstream steps were performed as described previously102. Hi-C libraries were sequenced on the Illumina NovaSeq 6000 at the Evrogen facility (www.evrogen.ru) resulting in 71–98 million 150-nt paired-end reads per sample (Supplementary Data 11).
RNA-seq in S2 cells upon Elys-KD
~107 control and Elys-KD S2 cells (in two biological replicates) were collected, and total RNAs were isolated using Trizol reagent (Invitrogen). Further steps of RNA-seq, such as isolation of poly(A+) RNAs on the oligo(dT) columns, synthesis of cDNAs using random primers, and sequencing of cDNA libraries resulting in 77–88 million 150-nt paired-end reads per sample (Supplementary Data 11), were performed at the GENEWIZ facility (www.genewiz.com).
Analysis of DamID-seq data
Sequencing reads from two biological replicates of Dam, Dam-Lam, or Dam-Elys samples were adapter clipped and uniquely mapped to the dm3/R5 genomic assembly by bowtie2103. Reads were counted by HTSeq-count software104 in the 0.3-kb genomic bins. Read counts were merged between replicates, as they were highly correlated (Supplementary Fig. 16a). The resulting read counts of Dam, Dam-Lam, or Dam-Elys samples were converted to reads per million mapped reads (RPM), normalized to RPM values of the Dam samples and log2 transformed. Next, Lam and Elys domain calling was performed with the two-state or three-state HMM algorithm, respectively (Supplementary Data 3 and 4). The log2 transformed profiles were visualized in the UCSC genome browser using an auto-scale to data view and a smoothing window of 3 pixels.
Analysis of external data
Various datasets were retrieved from GEO NCBI: Nup98_NPC and Nup98_nucl DamID profiles in Kc167 cells – from GSE1930728; Lam DamID profile in Kc167 cells – from GSE203115; Lam DamID profiles in the central brain, Elav-positive neurons, Repo-positive glia, and the fat body – from GSE1094956; H3K27Ac ChIP-chip profile in S2 cells – from GSE2077974; GAF ChIP-seq profile in S2 cells – from GSE40646105; Nup98, Elys and Nup93 ChIP-seq profiles and peaks in S2 cells – from GSE9492235 and GSE13561037; STARR-seq enhancer profiles in S2 cells – from GSE4073983 and GSE4769184. The enrichment regions for Nup98_NPC, Nup98_nucl, Lam, H3K27Ac, and STARR-seq enhancer profiles were identified with two-state (for Lam and STARR-seq enhancers) or three-state (for Nup98_NPC, Nup98_nucl and H3K27Ac) HMM. Less than 900-bp gaps between domains were filled in. For peak calling of ChIP-seq profiles, MACS2106 was applied. Domain/domain or gene/domain intersections were computed in R as a ratio of genome coverage using the GenomicRanges package in Bioconductor107. To perform permutation analysis we invoked BEDTools108 in R to reshuffle 10,000 times various sites of Elys, Nup98, genes, or enhancers, counted the number of intersected sites, and compared them with the original data.
Analysis of RNA-seq data
Sequencing reads from two biological replicates of control and Elys-KD S2 cells were adapter clipped. Low-quality reads with a length of less than 20 nt were filtered out and the remaining reads were uniquely mapped to the dm3/R5 genome assembly. To test whether RNA-seq replicates were similar, sequencing reads were counted per 0.3-kb genomic bins. Then, counts were normalized for sequencing depth and clustering analysis was performed (Supplementary Fig. 16b). Since replicates were highly correlated, they were merged. Reads were counted for genes of FlyBase r5.57 annotation and converted to transcripts per million (TPM) values using the Salmon tool109 (Supplementary Data 9). An analysis of the resulting data was performed in RStudio IDE using the R packages tximport, GenomicRanges, dplyr, and ggplot2. Differentially expressed genes upon Elys-KD were identified by the DESeq2 package110 for R with the cutoff parameters P < 0.05 and TPM FC > 1.5 (Supplementary Data 9).
Analysis of Hi-C data
Sequencing reads from two biological replicates of Hi-C control and Elys-KD experiments were trimmed with Trim Galore and processed with the distiller-nf pipeline (version 0.3.3)111. Parameter “stringency 3” was applied during the trimming procedure. Reads were mapped to chromosomes 2L, 2R, 3L, 3R, 4, X, and M of the dm3/R5 Drosophila genome assembly with the default settings and with an option MAPQ_30 (mapq1 >= 30 and mapq2 >= 30). For the downstream analysis of the interaction matrix, contacts between genomic bins separated by < 1 kb were removed. After that, only chromosomes 2L, 2R, 3L, 3R, and X were considered. For the replicate comparison, Hi-C maps were downsampled with the cooltools package112 to nearly the same total number of contacts across all replicates. Then, the iterative correction (IC)113 was applied, and samples were clustered with the R package HiCRep114 based on the stratum-adjusted correlation coefficient (SCC). Clustering was performed on 4-kb resolution Hi-C maps for bins separated by ≤ 1 Mb. SCCs obtained for each chromosome were averaged and used to build a hierarchical dendrogram (Supplementary Fig. 16c). Next, replicates were merged with the cooler package115, downsampled to nearly the same total counts, and corrected by IC, resulting in the final Hi-C maps.
TAD boundaries were called with the Armatus (version 2.2) software78 in 4-kb resolution Hi-C maps, which were additionally preprocessed. Each interchromosomal matrix was linearly interpolated, the contacts exceeding the range between the 1st and the 99th percentiles were replaced with the corresponding values of these percentiles and, finally, matrices were ln-transformed. Then Armatus was run for autosomes with the parameter γ = 1. For the X chromosome, γ = 0.6 and γ = 0.4 were chosen for control and Elys-KD maps respectively. For the downstream analysis, TADs were divided into 3 groups by their activity. To simultaneously account for the LAD coverage and the proportion of active chromatin (states 1 and 2 according to the 9-state model in S2 cells)74, the Jaccard coefficient between these two metrics was calculated by the formula: Jacc = (X − Y)/(X + Y), where X is the proportion of LADs and Y is the proportion of chromatin states 1 and 2 in the TAD. The most active TADs with a Jaccard coefficient < −0.8 were attributed to group A. TADs of intermediate activity with Jaccard coefficient ≥ −0.8 and ≤ 0.8 were attributed to group B. Finally, inactive TADs with Jaccard coefficient > 0.8 were attributed to group C. ACF was calculated based on the distance-normalized (observed/expected) IC Hi-C maps as the mean value of contacts within a TAD.
Active (A) and inactive (B) compartments77 were identified for 10-kb resolution IC Hi-C maps with the cooltools package. Positive values of the first principal component (PC1) corresponding to the A compartment were selected by correlation with gene expression. Cooltools package112 was used for the saddle-plot generation, which included the following steps. For each chromosome, bins were ranked according to their PC1 values, and 1% of bins with the highest and lowest PC1 values were filtered out. Then, distance-normalized contact frequencies were averaged within quantiles of the PC1, and the resulting values were used for the generation of saddle plots. We applied the same number of PC1 quantiles for both A and B compartments across all chromosomes and experimental conditions to analyze changes of compartmentalization, averaged by chromosomes. Box-plots showing the saddle-plot difference between Elys-KD and control include interactions between the bins falling within the 10 highest and lowest quantiles of PC1.
The insulation score (IS)116 was calculated for 2-kb resolution Hi-C maps. The modified IS (ISm) was calculated as the average distance-normalized contact frequency within the 8-kb square, starting at the second diagonal. Thus, contact frequency within a bin containing the Elys_NPC site and its interactions with the adjacent bins were not taken into account. Averaged ISm-curves were smoothed using linear interpolation.
Generation of averaged profiles
Averaged profiles and A/T profiles were generated in RStudio using the R packages GenomicRanges, dplyr ggplot2, Biostrings, seqinr, and rtracklayer. Metagene profiles were generated using deepTools2 software117 and RStudio. Heatmaps of Lam profiles were generated using R with the genomation package118.
DNA sequence motif analysis
For sequence motif identification, MEME 5.3.3 with the default settings was applied75.
Statistics and reproducibility
The Mann-Whitney (M-W) U-test was applied for P-values estimation upon comparison of two distributions. The Wilcoxon signed-rank test was applied for the estimation of whether log2 FC values were symmetric around zero. P-values for occasional gene/site or site/site overlapping were estimated by in silico performed permutation test with 10,000 random permutations. Upon each iteration, we counted the number of overlap events between two types of genomic sites. P-values were calculated as the ratio of permutations with a greater number of overlap events relative to that occurring in the genome.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Raw and processed DamID-seq, RNA-seq, and Hi-C data were deposited in the NCBI Gene Expression Omnibus (GEO) under the accession numbers GSE219152 (for DamID-seq, RNA-seq) and GSE218886 (for Hi-C). The numerical source data behind the graphs in the paper can be found in Supplementary Data. All other data and materials, including plasmid DNA, that support the findings of this study are available from the corresponding authors upon reasonable request. Uncropped/unedited blot image related to Fig. 1e is presented in Supplementary Fig. 3. The link for UCSC Genome Browser showing Elys DamID profile https://genome.ucsc.edu/cgi-bin/hgTracks?db=dm3&lastVirtModeType=default&lastVirtModeExtraState=&virtModeType=default&virtMode=0&nonVirtPosition=&position=chr2R%3A16166667-16833333&hgsid=2292689452_aXCaEPy5C7QaMaomtYGbtb1oGayN.
Code availability
In-house scripts used in this study are available at Zenodo repository119.
References
Comings, D. E. The rationale for an ordered arrangement of chromatin in the interphase nucleus. Am. J. Hum. Genet. 20, 440–460 (1968).
Gruenbaum, Y. & Foisner, R. Lamins: nuclear intermediate filament proteins with fundamental functions in nuclear mechanics and genome regulation. Annu. Rev. Biochem. 84, 131–164 (2015).
Guelen, L. et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 453, 948–951 (2008).
Peric-Hupkes, D. et al. Molecular maps of the reorganization of genome-nuclear lamina interactions during differentiation. Mol. Cell 38, 603–613 (2010).
van Bemmel, J. G. et al. The insulator protein SU(HW) fine-tunes nuclear lamina interactions of the Drosophila genome. PLoS ONE 5, e15013 (2010).
Pindyurin, A. V. et al. The large fraction of heterochromatin in Drosophila neurons is bound by both B-type lamin and HP1a. Epigenetics Chromatin 11, 65 (2018).
Ikegami, K., Egelhofer, T. A., Strome, S. & Lieb, J. D. Caenorhabditis elegans chromosome arms are anchored to the nuclear membrane via discontinuous association with LEM-2. Genome Biol. 11, R120 (2010).
Handoko, L. et al. CTCF-mediated functional chromatin interactome in pluripotent cells. Nat. Genet. 43, 630–638 (2011).
Shevelyov, Y. Y. et al. The B-type lamin is required for somatic repression of testis-specific gene clusters. Proc. Natl Acad. Sci. USA 106, 3282–3287 (2009).
Mattout, A. et al. An EDMD mutation in C. elegans lamin blocks muscle-specific gene relocation and compromises muscle integrity. Curr. Biol. 21, 1603–1614 (2011).
Zullo, J. M. et al. DNA sequence-dependent compartmentalization and silencing of chromatin at the nuclear lamina. Cell 149, 1474–1487 (2012).
Solovei, I. et al. LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate differentiation. Cell 152, 584–598 (2013).
Kohwi, M., Lupton, J. R., Lai, S. L., Miller, M. R. & Doe, C. Q. Developmentally regulated subnuclear genome reorganization restricts neural progenitor competence in Drosophila. Cell 152, 97–108 (2013).
Poleshko, A. et al. The human protein PRR14 tethers heterochromatin to the nuclear lamina during interphase and mitotic exit. Cell Rep. 5, 292–301 (2013).
Harr, J. C. et al. Directed targeting of chromatin to the nuclear lamina is mediated by chromatin state and A-type lamins. J. Cell Biol. 208, 33–52 (2015).
Lukášová, E., Kovarík, A., Bacíková, A., Falk, M. & Kozubek, S. Loss of lamin B receptor is necessary to induce cellular senescence. Biochem. J. 474, 281–300 (2017).
Zhu, Y. et al. Comprehensive characterization of neutrophil genome topology. Genes Dev. 31, 141–153 (2017).
Zheng, X. et al. Lamins organize the global three-dimensional genome from the nuclear periphery. Mol. Cell 71, 802–815.e7 (2018).
Ulianov, S. V. et al. Nuclear lamina integrity is required for proper spatial organization of chromatin in Drosophila. Nat. Commun. 10, 1176 (2019).
Shevelyov, Y. Y. & Ulianov, S. V. The nuclear lamina as an organizer of chromosome architecture. Cells 8, 136 (2019).
Brickner, J. H. & Walter, P. Gene recruitment of the activated INO1 locus to the nuclear membrane. PLoS Biol. 2, e342 (2004).
Casolari, J. M. et al. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell 117, 427–439 (2004).
Cabal, G. G. et al. SAGA interacting factors confine sub-diffusion of transcribed genes to the nuclear envelope. Nature 441, 770–773 (2006).
Taddei, A. et al. Nuclear pore association confers optimal expression levels for an inducible yeast gene. Nature 441, 774–778 (2006).
Brown, C. R., Kennedy, C. J., Delmar, V. A., Forbes, D. J. & Silver, P. A. Global histone acetylation induces functional genomic reorganization at mammalian nuclear pore complexes. Genes Dev. 22, 627–639 (2008).
Ahmed, S. et al. DNA zip codes control an ancient mechanism for gene targeting to the nuclear periphery. Nat. Cell Biol. 12, 111–118 (2010).
Capelson, M. et al. Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell 140, 372–383 (2010).
Kalverda, B., Pickersgill, H., Shloma, V. V. & Fornerod, M. Nucleoporins directly stimulate expression of developmental and cell-cycle genes inside the nucleoplasm. Cell 140, 360–371 (2010).
Vaquerizas, J. M. et al. Nuclear pore proteins nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS Genet. 6, e1000846 (2010).
Brickner, D. G. et al. Transcription factor binding to a DNA zip code controls interchromosomal clustering at the nuclear periphery. Dev. Cell 22, 1234–1246 (2012).
Liang, Y., Franks, T. M., Marchetto, M. C., Gage, F. H. & Hetzer, M. W. Dynamic association of NUP98 with the human genome. PLoS Genet. 9, e1003308 (2013).
Pascual-Garcia, P., Jeong, J. & Capelson, M. Nucleoporin Nup98 associates with Trx/MLL and NSL histone-modifying complexes and regulates Hox gene expression. Cell Rep. 9, 433–442 (2014).
Jacinto, F. V., Benner, C. & Hetzer, M. W. The nucleoporin Nup153 regulates embryonic stem cell pluripotency through gene silencing. Genes Dev. 29, 1224–1238 (2015).
Ibarra, A., Benner, C., Tyagi, S., Cool, J. & Hetzer, M. W. Nucleoporin-mediated regulation of cell identity genes. Genes Dev. 30, 2253–2258 (2016).
Pascual-Garcia, P. et al. Metazoan nuclear pores provide a scaffold for poised genes and mediate induced enhancer-promoter contacts. Mol. Cell 66, 63–76 (2017).
Kuhn, T. M., Pascual-Garcia, P., Gozalo, A., Little, S. C. & Capelson, M. Chromatin targeting of nuclear pore proteins induces chromatin decondensation. J. Cell Biol. 218, 2945–2961 (2019).
Gozalo, A. et al. Core components of the nuclear pore bind distinct states of chromatin and contribute to Polycomb repression. Mol. Cell 77, 67–81.e7 (2020).
Sharma, R. et al. Differential spatial and structural organization of the X chromosome underlies dosage compensation in C. elegans. Genes Dev. 28, 2591–2596 (2014).
Breuer, M. & Ohkura, H. A negative loop within the nuclear pore complex controls global chromatin organization. Genes Dev. 29, 1789–1794 (2015).
Shevelyov, Y. Y. The role of nucleoporin Elys in nuclear pore complex assembly and regulation of genome architecture. Int. J. Mol. Sci. 21, 9475 (2020).
Kimura, N. et al. Identification of a novel transcription factor, ELYS, expressed predominantly in mouse foetal haematopoietic tissues. Genes Cells 7, 435–446 (2002).
Rasala, B. A., Orjalo, A. V., Shen, Z., Briggs, S. & Forbes, D. J. ELYS is a dual nucleoporin/kinetochore protein required for nuclear pore assembly and proper cell division. Proc. Natl Acad. Sci. USA 103, 17801–17806 (2006).
Fernandez, A. G. & Piano, F. MEL-28 is downstream of the Ran cycle and is required for nuclear-envelope function and chromatin maintenance. Curr. Biol. 16, 1757–1763 (2006).
Galy, V., Askjaer, P., Franz, C., López-Iglesias, C. & Mattaj, I. W. MEL-28, a novel nuclear-envelope and kinetochore protein essential for zygotic nuclear-envelope assembly in C. elegans. Curr. Biol. 16, 1748–1756 (2006).
Franz, C. et al. MEL-28/ELYS is required for the recruitment of nucleoporins to chromatin and postmitotic nuclear pore complex assembly. EMBO Rep. 8, 165–172 (2007).
Gillespie, P. J., Khoudoli, G. A., Stewart, G., Swedlow, J. R. & Blow, J. J. ELYS/MEL-28 chromatin association coordinates nuclear pore complex assembly and replication licensing. Curr. Biol. 17, 1657–1662 (2007).
Rasala, B. A., Ramos, C., Harel, A. & Forbes, D. J. Capture of AT-rich chromatin by ELYS recruits POM121 and NDC1 to initiate nuclear pore assembly. Mol. Biol. Cell 19, 3982–3996 (2008).
Doucet, C. M., Talamas, J. A. & Hetzer, M. W. Cell cycle-dependent differences in nuclear pore complex assembly in metazoa. Cell 141, 1030–1041 (2010).
Mehta, S. J. K., Kumar, V. & Mishra, R. K. Drosophila ELYS regulates Dorsal dynamics during development. J. Biol. Chem. 295, 2421–2437 (2020).
Sabinina, V. J. et al. Three-dimensional superresolution fluorescence microscopy maps the variable molecular architecture of the nuclear pore complex. Mol. Biol. Cell 32, 1523–1533 (2021).
Bilokapic, S. & Schwartz, T. U. Molecular basis for Nup37 and ELY5/ELYS recruitment to the nuclear pore complex. Proc. Natl Acad. Sci. USA 109, 15241–15246 (2012).
Gómez-Saldivar, G. et al. Identification of conserved MEL-28/ELYS domains with essential roles in nuclear assembly and chromosome segregation. PLoS Genet. 12, e1006131 (2016).
Inoue, A. & Zhang, Y. Nucleosome assembly is required for nuclear pore complex assembly in mouse zygotes. Nat. Struct. Mol. Biol. 21, 609–616 (2014).
Zierhut, C., Jenness, C., Kimura, H. & Funabiki, H. Nucleosomal regulation of chromatin composition and nuclear assembly revealed by histone depletion. Nat. Struct. Mol. Biol. 21, 617–625 (2014).
Kobayashi, W. et al. Structural and biochemical analyses of the nuclear pore complex component ELYS identify residues responsible for nucleosome binding. Commun. Biol. 2, 163 (2019).
Skrajna, A. et al. Comprehensive nucleosome interactome screen establishes fundamental principles of nucleosome binding. Nucleic Acids Res. 48, 9415–9432 (2020).
Mohrmann, L. et al. Differential targeting of two distinct SWI/SNF-related Drosophila chromatin-remodeling complexes. Mol. Cell. Biol. 24, 3077–3088 (2004).
Nakayama, T., Shimojima, T. & Hirose, S. The PBAP remodeling complex is required for histone H3.3 replacement at chromatin boundaries and for boundary functions. Development 139, 4582–4590 (2012).
Judd, J., Duarte, F. M. & Lis, J. T. Pioneer-like factor GAF cooperates with PBAP (SWI/SNF) and NURF (ISWI) to regulate transcription. Genes Dev. 35, 147–156 (2021).
Richards, L., Lord, C. L., Benton, M. L., Capra, J. A. & Nordman, J. T. Nucleoporins facilitate ORC loading onto chromatin. Cell Rep. 41, 111590 (2022).
Dixon, J. R. et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 485, 376–380 (2012).
Hou, C., Li, L., Qin, Z. S. & Corces, V. G. Gene density, transcription, and insulators contribute to the partition of the Drosophila genome into physical domains. Mol. Cell 48, 471–484 (2012).
Nora, E. P. et al. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature 485, 381–385 (2012).
Sexton, T. et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell 148, 458–472 (2012).
Ilyin, A. A. et al. Piwi interacts with chromatin at nuclear pores and promiscuously binds nuclear transcripts in Drosophila ovarian somatic cells. Nucleic Acids Res. 45, 7666–7680 (2017).
Lenz-Böhme, B. et al. Insertional mutation of the Drosophila nuclear lamin Dm0 gene results in defective nuclear envelopes, clustering of nuclear pore complexes, and accumulation of annulate lamellae. J. Cell Biol. 137, 1001–1016 (1997).
Lau, C. K. et al. Transportin regulates major mitotic assembly events: from spindle to nuclear pore assembly. Mol. Biol. Cell 20, 4043–4058 (2009).
Rotem, A. et al. Importin beta regulates the seeding of chromatin with initiation sites for nuclear pore assembly. Mol. Biol. Cell 20, 4031–4042 (2009).
Zykova, T. Y., Levitsky, V. G., Belyaeva, E. S. & Zhimulev, I. F. Polytene chromosomes - a portrait of functional organization of the Drosophila genome. Curr. Genom. 19, 179–191 (2018).
Shevelyov, Y. Y. & Nurminsky, D. I. The nuclear lamina as a gene-silencing hub. Curr. Issues Mol. Biol. 14, 27–38 (2012).
van Steensel, B. & Henikoff, S. Identification of in vivo DNA targets of chromatin proteins using tethered dam methyltransferase. Nat. Biotechnol. 18, 424–428 (2000).
Filion, G. J. et al. Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell 143, 212–224 (2010).
Laktionov, P. P., White-Cooper, H., Maksimov, D. A. & Beliakin, S. N. Transcription factor comr acts as a direct activator in the genetic program controlling spermatogenesis in D. melanogaster. Mol. Biol. 48, 153–165 (2014).
Kharchenko, P. V. et al. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. Nature 471, 480–485 (2011).
Bailey, T. L., Johnson, J., Grant, C. E. & Noble, W. S. The MEME suite. Nucleic Acids Res. 43, W39–W49 (2015).
Vogel, M. J., Peric-Hupkes, D. & van Steensel, B. Detection of in vivo protein-DNA interactions using DamID in mammalian cells. Nat. Protoc. 2, 1467–1478 (2007).
Lieberman-Aiden, E. et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326, 289–293 (2009).
Filippova, D., Patro, R., Duggal, G. & Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms Mol. Biol. 9, 14 (2014).
Shevelyov, Y. Y., Ulianov, S. V., Gelfand, M. S., Belyakin, S. N. & Razin, S. V. Dosage compensation in Drosophila: its canonical and non-canonical mechanisms. Int. J. Mol. Sci. 23, 10976 (2022).
Ulianov, S. V. et al. Active chromatin and transcription play a key role in chromosome partitioning into topologically associating domains. Genome Res. 26, 70–84 (2016).
Pott, S. & Lieb, J. D. What are super-enhancers? Nat. Genet. 47, 8–12 (2015).
Tyagi, S. et al. High-precision mapping of nuclear pore-chromatin interactions reveals new principles of genome organization at the nuclear envelope. eLife 12, RP87462 (2023).
Arnold, C. D. et al. Genome-wide quantitative enhancer activity maps identified by STARR-seq. Science 339, 1074–1077 (2013).
Shlyueva, D. et al. Hormone-responsive enhancer-activity maps reveal predictive motifs, indirect repression, and targeting of closed chromatin. Mol. Cell 54, 180–192 (2014).
Cordes, V. C., Reidenbach, S. & Franke, W. W. Cytoplasmic annulate lamellae in cultured cells: composition, distribution, and mitotic behavior. Cell Tissue Res. 284, 177–191 (1996).
Hampoelz, B. et al. Pre-assembled nuclear pores insert into the nuclear envelope during early development. Cell 166, 664–678 (2016).
Ren, H. et al. Postmitotic annulate lamellae assembly contributes to nuclear envelope reconstitution in daughter cells. J. Biol. Chem. 294, 10383–10391 (2019).
Kalashnikova, A. A., Porter-Goff, M. E., Muthurajan, U. M., Luger, K. & Hansen, J. C. The role of the nucleosome acidic patch in modulating higher order chromatin structure. J. R. Soc. Interface 10, 20121022 (2013).
Light, W. H. et al. A conserved role for human Nup98 in altering chromatin structure and promoting epigenetic transcriptional memory. PLoS Biol. 11, e1001524 (2013).
Kadota, S. et al. Nucleoporin 153 links nuclear pore complex to chromatin architecture by mediating CTCF and cohesin binding. Nat. Commun. 11, 2606 (2020).
Ilyin, A. A. et al. Comparison of genome architecture at two stages of male germline cell differentiation in Drosophila. Nucleic Acids Res. 50, 3203–3225 (2022).
Markstein, M., Pitsouli, C., Villalta, C., Celniker, S. E. & Perrimon, N. Exploiting position effects and the gypsy retrovirus insulator to engineer precisely expressed transgenes. Nat. Genet. 40, 476–483 (2008).
Bischof, J., Maeda, R. K., Hediger, M., Karch, F. & Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc. Natl Acad. Sci. USA 104, 3312–3317 (2007).
Pindyurin, A. V. Genome-wide cell type-specific mapping of in vivo chromatin protein binding using an FLP-inducible DamID system in Drosophila. Methods Mol. Biol. 1654, 99–124 (2017).
Lavrov, S., Déjardin, J. & Cavalli, G. Combined immunostaining and FISH analysis of polytene chromosomes. Methods Mol. Biol. 247, 289–303 (2004).
Clemens, J. C. et al. Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc. Natl Acad. Sci. USA 97, 6499–6503 (2000).
Stuurman, N., Maus, N. & Fisher, P. A. Interphase phosphorylation of the Drosophila nuclear lamin: site-mapping using a monoclonal antibody. J. Cell Sci. 108, 3137–3144 (1995).
Katsani, K. R., Karess, R. E., Dostatni, N. & Doye, V. In vivo dynamics of Drosophila nuclear envelope components. Mol. Biol. Cell 19, 3652–3666 (2008).
Wagner, N., Weber, D., Seitz, S. & Krohne, G. The lamin B receptor of Drosophila melanogaster. J. Cell Sci. 117, 2015–2028 (2004).
Blower, M. D. & Karpen, G. H. The role of Drosophila CID in kinetochore formation, cell-cycle progression and heterochromatin interactions. Nat. Cell Biol. 3, 730–739 (2001).
Erokhin, M. et al. Transcriptional read-through is not sufficient to induce an epigenetic switch in the silencing activity of Polycomb response elements. Proc. Natl Acad. Sci. USA 112, 14930–14935 (2015).
Ulianov, S. V. et al. Order and stochasticity in the folding of individual Drosophila genomes. Nat. Commun. 12, 41 (2021).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Anders, S., Pyl, P. T. & Huber, W. HTSeq – a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Fuda, N. J. et al. GAGA factor maintains nucleosome-free regions and has a role in RNA polymerase II recruitment to promoters. PLoS Genet. 11, e1005108 (2015).
Feng, J., Liu, T., Qin, B., Zhang, Y. & Liu, X. S. Identifying ChIP-seq enrichment using MACS. Nat. Protoc. 7, 1728–1740 (2012).
Lawrence, M. et al. Software for computing and annotating genomic ranges. PLoS Comput. Biol. 9, e1003118 (2013).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 14, 417–419 (2017).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Goloborodko, A., Venev, S., Abdennur N., azkalot1 & Di Tommaso, P. A modular Hi-C mapping pipeline. Version v0.3.3rc3. Zenodo https://doi.org/10.5281/zenodo.3350926 (2019).
Open2C et al.Cooltools: enabling high-resolution Hi-C analysis in Python. PLoS Comput. Biol. 20, e1012067 (2024).
Imakaev, M. et al. Iterative correction of Hi-C data reveals hallmarks of chromosome organization. Nat. Methods 9, 999–1003 (2012).
Yang, T. et al. HiCRep: assessing the reproducibility of Hi-C data using a stratum-adjusted correlation coefficient. Genome Res. 27, 1939–1949 (2017).
Abdennur, N. & Mirny, L. A. Cooler: scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics 36, 311–316 (2020).
Crane, E. et al. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature 523, 240–244 (2015).
Ramírez, F. et al. deepTools2: a next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 44, W160–W165 (2016).
Akalin, A., Franke, V., Vlahoviček, K., Mason, C. E. & Schübeler, D. Genomation: a toolkit to summarize, annotate and visualize genomic intervals. Bioinformatics 31, 1127–1129 (2015).
Ilin, A. Foriin/Elys_embryos_2022: R scripts used for analysis of data in Doronin et al. 2023. Zenodo https://doi.org/10.5281/zenodo.7808899 (2023).
Acknowledgements
We thank Paul Fisher (Stony Brook University) for anti-Lamin antibodies, Georg Krohne (University of Würzburg) for anti-LBR antibodies, Gary Karpen (University of California, Berkeley) for anti-CenpA antibodies, Valerie Doye (Institut Jacques Monod) for anti-Nup107 antibodies, Daria Kopytova (Institute of Molecular Biology, RAS) for anti-Nup153 antibodies, Maksim Erokhin (Institute of Gene Biology, RAS) for anti-GAF antibodies. The main part of this work was supported by the Russian Science Foundation (23-14-00049 to Y.Y.S.). Hi-C experiments were supported by the Russian Science Foundation (21-64-00001 to S.V.R.). Hi-C data analysis was supported by the Russian Foundation for Basic Research (21-34-70051 to E.E.K.). Flow cytometry experiments were supported by the state task of the NRC “Kurchatov Institute”.
Author information
Authors and Affiliations
Contributions
Y.Y.S. conceived the project. S.A.D. generated the construct for DamID and carried out DamID in embryos, performed RNAi experiments, Western-blot analysis, co-immunoprecipitation, immunostaining, TUNEL, and FISH experiments. M.A.S. (supervised by S.V.U.) performed Hi-C experiments. E.A.M. maintained cell cultures and performed some immunostaining experiments. O.M.O. performed fly crossing. S.A.L. performed immunostaining of polytene chromosomes. A.A.F. carried out early FISH experiments. A.Y.I. carried out FISH experiments with a 36C probe. R.A.S. performed flow cytometry. A.A.I. (supervised by Y.Y.S.) analyzed various datasets (DamID, RNA-seq, and external data). A.D.K. (supervised by E.E.K.) analyzed Hi-C data. V.V.N. analyzed FISH results. Y.Y.S. wrote the draft of the manuscript and prepared figures. S.V.U., E.E.K., and S.V.R. made significant contributions to the final version with the input of all other authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Mengtan Xing.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Doronin, S.A., Ilyin, A.A., Kononkova, A.D. et al. Nucleoporin Elys attaches peripheral chromatin to the nuclear pores in interphase nuclei. Commun Biol 7, 783 (2024). https://doi.org/10.1038/s42003-024-06495-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-024-06495-w
- Springer Nature Limited