
Abstract
Plant meristems require a constant supply of photoassimilates and hormones to the dividing meristematic cells. In the growing root, such supply is delivered by protophloem sieve elements. Due to its preeminent function for the root apical meristem, protophloem is the first tissue to differentiate. This process is regulated by a genetic circuit involving in one side the positive regulators DOF transcription factors, OCTOPUS (OPS) and BREVIX RADIX (BRX), and in the other side the negative regulators CLAVATA3/EMBRYO SURROUNDING REGION RELATED (CLE) peptides and their cognate receptors BARELY ANY MERISTEM (BAM) receptor-like kinases. brx and ops mutants harbor a discontinuous protophloem that can be fully rescued by mutation in BAM3, but is only partially rescued when all three known phloem-specific CLE genes, CLE25/26/45 are simultaneously mutated. Here we identify a CLE gene closely related to CLE45, named CLE33. We show that double mutant cle33cle45 fully suppresses brx and ops protophloem phenotype. CLE33 orthologs are found in basal angiosperms, monocots, and eudicots, and the gene duplication which gave rise to CLE45 in Arabidopsis and other Brassicaceae appears to be a recent event. We thus discovered previously unidentified Arabidopsis CLE gene that is an essential player in protophloem formation.
Similar content being viewed by others

Introduction
In vascular plants, phloem tissues transport sugars and signaling molecules to sink organs for growth and storage1,2. In the growing Arabidopsis root, the root apical meristem is a large sink and two phloem poles, containing functional sieve elements, unload the phloem sap in the meristematic region3. Each pole consists of a protophloem sieve element, flanked by two phloem pole pericycle cells from the outside, and a metaphloem sieve element from the inside, and two companion cells adjacent to these sieve element cells. This complex structure acts as a functional unit4. Remarkably, the delivery of sugars and hormones to the root meristem is uniquely mediated by protophloem sieve elements, while metaphloem sieve elements remain undifferentiated in this region of the root5.
The process of protophloem sieve element formation requires radical cellular modifications, including cell wall reinforcement, enucleation, and sieve plate formation6,7. The genetic control of protophloem development in Arabidopsis has been studied intensively in the last decade4,6,8,9,10,11, including precise transcriptomics analysis of the whole phloem pole4 and developing sieve elements6. Protophloem differentiation is controlled by multiple regulators, such as hormone gradients, DOF transcription factors, and SMXL transcriptional repressors9,12. In addition, two membrane-localized proteins, BREVIS RADIX (BRX) and OCTOPUS (OPS) act as positive regulators of protophloem development8,10. brx and ops loss-of-function mutants display a discontinuous protophloem, characterized by so-called gap cells that fail to differentiate. The interrupted protophloem continuity results in a reduced phloem sap delivery to the root apical meristem and limited root growth8,10. BARELY ANY MERISTEM 3 (BAM3) was identified in a brx suppressor screen with bam3 rescuing both root growth and gap cell phenotypes of brx and ops10. BAM3 encodes for a leucine-rich repeat receptor kinase (LRR-RLK), a cognate receptor of the CLAVATA3/EMBRYO SURROUNDING REGION 45 (CLE45) peptide13. While a higher-order mutant of phloem-expressed CLE genes (CLE25/26/45) have been recently shown to partially rescue the brx and ops gap cell phenotype, this rescue is not to the level of the full rescue achieved in bam314. This phenotypic discrepancy between the receptor and its ligands suggests that additional endogenous CLE peptides, BAM3 ligands, remain to be identified.
CLE peptides are produced from a roughly 100 amino acids pre-propeptide that possess a signal peptide in its N-terminus for apoplastic release, a central variable region, and a conserved 12–13 residues CLE domain in its C-terminus, which is cleaved off to release the mature CLE peptide. Given their small gene size and high sequence variability outside the CLE domain, the genome annotation of CLE genes can be challenging. In Arabidopsis, their discoveries came initially from a mutant screen15,16, following the cloning of CLV317. The first release of the Arabidopsis genome18 allowed the identification of additional members of the CLE family in several iterations19,20,21,22,23. Up to date, 32 CLE genes are annotated encoding for 27 unique peptides. We recently explored the CLE gene family in tomato, and found 37 new CLE genes24, sparking the question whether additional CLE genes remain to be discovered in Arabidopsis.
Here we present the identification of the Arabidopsis CLE33 gene, which was missed in previous genome analyses. We show that this gene is expressed in the developing root phloem and encodes for an active peptide which acts redundantly with CLE45 via BAM3 receptor in inhibiting protophloem differentiation in the protophloem cell lineage and neighboring cells.
Results
CLE33 is a novel Arabidopsis peptide gene
To search for additional Arabidopsis CLE genes, we performed a non-stringent t-BLAST-n analysis using the 32 previously known full-length CLE proteins from A. thaliana as queries. We found one sequence in an unannotated region of chromosome 1 that shows all the characteristics of a CLE gene, including the presence of a functional signal peptide. In other Brassicaceae species, this genetic locus is conserved and is annotated as a CLE45-like homolog (Supplementary Fig. 1a). In A. thaliana, this locus is transcribed, confirming expression of the gene, which we named CLE33 (Fig. 1b and Supplementary Fig. 1b). Based on its full-length protein sequence, we determined that CLE33 is the closest homolog of the well-characterized phloem-expressed CLE45 (Fig. 1a)10,11,13.

a Phylogenetic tree of Arabidopsis thaliana full-length CLE pre-propeptides. The blue line indicates the cluster expressed in the root phloem tissue. b Araport11 RNA-seq-based evidence of transcription from mapping coverage of the light-grown seedlings visualized with JBrowse. The relative position of the predicted CLE33 coding sequence is indicated schematically: signal peptide in green, CLE domain in blue. c Transcriptional fusions showing the promoter activity of CLE33, CLE45 and CLE26 in the root protophloem tissue. Scale bar = 50 μm.
To get insights about the expression pattern of CLE33, we cloned its promoter region to drive a GUS reporter (Supplementary Fig. 1c). CLE33 expression was specifically associated with vasculature in the root meristem, rosette junction and leaves, cotyledons, and inflorescence stem. To precisely determine the expressed tissue, we use the same promoter and two additional phloem-specific CLEs (CLE26 and CLE45) to drive the expression of nuclear fluorescent protein NLS-Citrine (Fig. 1c and Supplementary Fig. 1d). In the root meristem, all three are specifically expressed in protophloem tissue, but with slight differences in the expression onset. CLE45 expression can be observed early in the sieve element precursor cells, immediately after the first periclinal cell division of the sieve element procambium precursor. CLE33 expression starts a few cells later, in the developing protophloem sieve element cells, largely overlapping with the expression domain of CLE45. CLE26 is active in the later stages of sieve element differentiation, when the cell wall thickening has happened, in accordance with the previously published reporters25,26.
CLE33 encodes for a root-active peptide perceived by BAM3
Most CLE peptides induce root growth arrest when overexpressed or when added to the growth media at nanomolar concentrations and are therefore called root-active13,26. To ascertain whether CLE33 peptide could also inhibit primary root growth and which receptors are involved in perceiving this peptide in the root meristem, we applied CLE33 peptide at increasing concentration from 0 to 300 nM in the growth media and assessed the root growth inhibition effect in the wild-type and loss-of-function CLE receptor mutants (Fig. 2a). In the wild type, the root growth was inhibited with 3 nM of CLE33, becoming more pronounced with higher concentrations. As for other known root-active CLEs, this response was fully CLV2/CORYNE-dependent13, even at the highest concentrations. In contrast, bam mutants exhibited a variety of responses. bam2-4 showed similar sensitivity to the wild type, whereas bam1-3 was hypersensitive to CLE33p. This hypersensitivity of around 3-fold was also observed with CLE26 and CLE45 peptides (Supplementary Fig. 2a, b), and to CLE33 peptide in the independent bam1-4 allele (Supplementary Fig. 2d). On the other hand, bam3 mutants were insensitive up to 100 nM of CLE33 peptide, and only showed a response at very high concentration of 300 nM (Fig. 2a and Supplementary Fig. 2c). Only in combination of BAM1 loss-of-function, full insensitiveness was achieved, suggesting that BAM1 plays a role in mediating root growth responses to synthetic CLE33 peptides at high concentration. To further test this finding under closer physiological conditions, we used the XVE-estradiol inducible system to ectopically express CLE33 (Supplementary Fig. 3). While the wild type strongly reacted to the ectopic induction of CLE33, bam3-2 lines remained insensitive even at high estradiol concentration. These results suggest that synthetic and endogenous CLE33 peptides require BAM3 for root growth inhibition effect.

a Dose-dependent root growth inhibition by CLE33p in the wild type and CLE receptor mutants. b Root growth inhibition with synthetic CLE33 (30 nM), CLE45 (30 nM), CLE26 (3 nM) peptides. c Root growth inhibition of CLE33 peptide variants CLE33M2K, CLE33G4P, CLE33S7P (30 nM). a–c Letters indicate different statistical groups (ANOVA, post hoc Tukey test).
BAM3 was shown to be the cognate receptor of CLE45 peptide13, and bam3 mutants present a similar insensitivity to CLE33 and CLE45 peptides (Fig. 2b and Supplementary Fig. 2a). This unique BAM3-dependency is very specific to CLE33 and CLE45 peptides, but not to the closely related CLE26 peptide (Supplementary Fig. 2b). By comparing the amino acid sequences of these three peptides, we noticed that the conserved prolines at the positions 4 and 7 are replaced in CLE33 by glycine and serine, and in CLE45 by arginine and serine (Fig. 2b). To further investigate the importance of these amino acids for receptor specificity, we created and tested variants of CLE26 and CLE33 by swapping residues between these two peptides. First, we observed that all the peptide variants (CLE26P4G, CLE26P7S, CLE26P4G/P7S, CLE33M2K, CLE33G4P, CLE33S7P) had a strong root growth inhibition effect in wild type but largely no effect in crn-10 mutant, which implies that the amino acids substitutions did not affect their ability to suppress root growth (Fig. 2c and Supplementary Fig. 4). However, the BAM3-dependency was strongly relying on the residues at positions 4 and 7. The presence of a proline at position 4 or 7 in CLE33 led to a BAM3 independent response (Fig. 2c), and inversely both prolines needed to be replaced in CLE26 to not inhibit root growth in the bam3 mutant (Supplementary Fig. 4). Taken together, these results suggest that the absence of prolines in position 4 and 7 determine the BAM3 perception specificity of the CLE33 and CLE45 peptides.
CLE33 and CLE45 act in concert in protophloem differentiation
It has been previously shown that root protophloem development is suppressed by root-active CLE peptides added to the growth media13. To test the effect of CLE33 on protophloem identity, we evaluated the expression of the protophloem marker pCVP2:NLS-3xVenus upon overnight treatment of CLE33 peptide (Supplementary Fig. 5). The inhibitory effect of CLE33 on protophloem identity was similar to the effect of CLE26 and CLE45, suggesting that CLE33 acts as a negative regulator of protophloem differentiation.
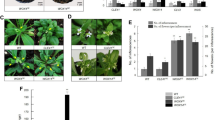
To understand the biological function of CLE33 in planta and its genetic relationship with CLE45, we created CRISPR-Cas9 mediated knock-out mutants in wild type and cle45-2 backgrounds (Supplementary Fig. 6). Alike bam3¸ the single cle33 and double cle33cle45 mutants do not show any reproducible root growth macro-phenotype in our conditions (Fig. 3 and Supplementary Fig. 7). In accordance to previous reports, BAM3 loss-of-function mutants can fully rescue the discontinuous protophloem differentiation of brx and ops8,10, which is not the case for cle4514. We asked whether this phenotypic discrepancy could be attributed to the presence of the remaining CLE33 in protophloem tissue. To explore this further, we crossed the double mutant cle33-3 cle45-2 with brx-3 and ops-2. Mutation in CLE33 alone had no or very little impact on rescuing root growth and protophloem continuity of brx-3 and ops-2 (Fig. 3 and Supplementary Fig. 7). We could reproduce a previously published result, where cle45 partially suppressed the root growth phenotype but failed to significantly reduce the protophloem gap cell frequency14. The combination of cle33 and cle45 could fully rescue the root growth and protophloem discontinuity phenotypes, strongly suggesting that CLE33 and CLE45 act redundantly in this genetic circuit (Fig. 3 and Supplementary Fig. 7). These results taken together support a negative role for CLE33 in protophloem differentiation.

a Root length at 10 days post germination. Letters indicate different statistical group (ANOVA, post hoc Tukey test). b Representative confocal images of calcofluor white stained roots. Red arrows indicate protophloem gap cells. Scale bars correspond to 100 μm. c Gap cell frequency quantification. Letters indicate statistical groups (χ2 test with Benjamini–Hochberg correction).
The protophloem development failure caused by BRX or OPS loss-of-function could be explained by high dosages of the receptor BAM3 or phloem-specific CLE ligands. To test this possibility, we quantified transcript accumulation in brx and ops root tissues. We found, that on the transcriptional level BAM3 is not up-regulated and CLE25, CLE26 and CLE45 even showed significant reduction in their expression (Supplementary Fig. 8a), suggesting that the increased BAM3/CLE33/45- mediated signaling in brx and ops mutant is caused by another mechanism.
While analyzing the protophloem continuity, we came across an additional phenotype of ectopic sieve element-like cells originated from the neighboring cells adjacent to the main protophloem sieve element cell file. These cell files show characteristics of developing sieve elements, including thickening of the cell wall (Fig. 4a). A related phenotype was recently shown in the multiple loss-of-function mutants in phloem CLEs, BAM receptor, and CIK co-receptor mutants, which demonstrated that CLE signaling restricts neighboring cells to differentiate into sieve elements9. This previous analysis was performed at a later stage in phloem development when both protophloem sieve elements and metaphloem sieve elements have differentiated and possess thick cell walls9. In our analyses, we focused on the meristematic zone. We quantified the frequency of the ectopic protophloem differentiation events in our mutants. We found an increased ectopic differentiation in the absence of BAM3 and in the triple mutants cle33 cle45 brx and cle33 cle45 ops, but not in the double mutant combinations (Fig. 4b and Supplementary Fig. 8b). This difference could be due to a weaker phloem identity in the absence of brx and ops, and/or reduced levels of CLE25/CLE26 in these backgrounds (Supplementary Fig. 8a) which are known to repress ectopic sieve element formation9. Nevertheless, the results of our analysis show that CLE33 and CLE45 act redundantly in inhibiting the formation of the ectopic protophloem sieve elements (Fig. 4c and Supplementary Fig. 7).

a Confocal images of calcofluor white stained roots. Blue arrows indicate cell files showing a cell wall reinforcement, sign that they are undergoing a protophloem differentiation process. Scale bars correspond to 50 μm. b Frequency of ectopic protophloem differentiation in roots. Letters indicate statistical groups (χ2 test with Benjamini–Hochberg correction). c Schematic model of autocrine and paracrine signaling mediated by the CLE33/45 peptides via the BAM3 receptor.
It has been recently demonstrated that CLE25, CLE26, and CLE45 have DOF-transcription factor binding sites in their promoter region and are induced by DOF2.29. Our analysis of the CLE33 promoter region revealed similar binding sites for DOF2.2, DOF2.4/PEAR1, DOF3.2, OBP3, DOF5.1/PEAR2, and DOF5.6 (Supplementary Fig. 8c). This result suggests that CLE33 can be an essential player in DOF-CLE genetic circuit.
Evolutionary origin of CLE33 and CLE45
The two peptide-encoding genes, CLE33 and CLE45 appear to be functionally redundant (Figs. 3 and 4) and share a high sequence and expression pattern similarity (Fig. 1). Therefore, they could be the result of a gene duplication event. To learn about their evolutionary origin, we performed a phylogenetic analysis across diverse vascular plant species. We identified CLE33 orthologs in the genomes of basal angiosperms, monocots, and eudicots, but not of gymnosperms, suggesting that it emerged ~200 million years ago (Supplementary Fig. 9a). Interestingly, clear CLE45 orthologs were only found in Brassicales species. A multi-sequence alignment revealed that Brassicales CLE45 have an extended variable region with no homology to other CLE33/45-like proteins in the N-terminal part immediately after the signal peptide (Supplementary Fig. 9b). However, when analyzing specifically the CLE domain, we observed that CLE45 is more similar to ancestral CLE33 orthologs (Supplementary Fig. 9c). One possibility for such occurrence is that following a duplication in early Brassicales, one copy quickly drifted genetically in its variable region giving rise to the current CLE45. In such a scenario, Brassicales CLE33 maintained most of the ancestral sequence, except for the residues 2/3/4 in its CLE domain. Strikingly, all the orthologs of both CLE33 and CLE45 do not have any prolines at positions 4 and 7, which we found to be consequential for receptor perception specificity in Arabidopsis (Fig. 2).
Furthermore, we investigated the evolutionary origin of BAM3, the gene encoding for the receptor perceiving these two ligands. We found orthologs present up to the gymnosperms (Supplementary Fig. 10). An early duplication in the angiosperms produced two copies: BAM3 and BAM4. While BAM3 is found in all angiosperm groups, BAM4 is peculiarly absent from the genomes of Brassicaceae species. The loss of BAM4 coincides with the emergence of CLE45 in Brassicales. Yet, BAM4 ligands and their functions remain unknown. Our findings open new directions for further exploration into these receptor-ligand pairs and their potential conserved function throughout plant evolution.
Discussion
In recent years, many additional peptide ligand-encoding genes have been discovered in Arabidopsis and other plant species24,27,28,29,30,31. These belong to previously unknown families, such as the stress-induced CTNIPs/SCREWs peptides27,28, or well-characterized gene families, such as CIF/TWS131 and CLE in our study. It seems that plant genome annotations, including Arabidopsis, still miss many of such peptide-encoding genes. However, advanced methods like ribosome profiling combined with transcriptomics and peptidomics32,33 have shown a great success in identifying a few thousands of such genes in Medicago32 and maize33. In addition, the genome-wide analyses using hidden Markov models34 led to the identification of new peptides24,35. With the aid of such advanced methods, we will be soon able to obtain a full repertoire of plant signaling peptides regulating a myriad of developmental and adaptive responses.
In our study, we performed a non-stringent t-BLAST-n search for CLE genes, which allowed us to identify the previously non-annotated CLE33. Prolines at positions 4 and 7 are among the most conserved amino acids in CLE peptides, and their absence in CLE33/45 makes them exceptions and partially explains why these genes were identified later22. From a structural point of view, the proline residue confers unique conformational changes because of its irregular geometry36. The side chain of the proline is connected to the peptide backbone twice, creating its own secondary structure. Little is known about the contribution of prolines to the interactions with the CLE receptor and co-receptor binding surfaces. However, it was shown that in mature CLE peptides, prolines 4 and 7 undergo hydroxylation and either or both undergo glycosylation with 3, 4, or 6 residues of arabinose37. The hydroxyproline in position 4 in CLE40 peptide was shown to prevent the miscleavage of the precursor peptide38. At the same time, absence or presence of hydroxylation in the mature CLE40 peptide and substitution of the proline in position 4 to alanine (P4A), did not affect the bioactivity of the peptide in root growth assays38. Furthermore, replacing hydroxyprolines in positions 4 and 7 by prolines in CLE9 did not affect binding affinity to BAM1 in vitro39, which could have suggested that proline 4 itself and proline hydroxylation are not essential for peptide–receptor interaction38,39. However, our bioactivity assays with modified CLE26 and CLE33 peptides indicate that prolines at positions 4 and 7 are determinants for receptor specificity. It has been demonstrated that in BAM3, the Q226Y228Y231 residues are crucial for CLE45 binding13, but how the absence of prolines 4 and 7 contribute to the specific binding with BAM3 needs further clarification.
The emergence of highly specialized sugar-conducting sieve elements was a crucial step in the evolution of land plants. This invention facilitated the separation of photosynthetic organs that need to compete for light and grow towards the sun, and water-absorbing organs that grow deep into the soil. Primitive sieve element files can be found in brown algae species and mosses40, often showing only partial protoplast degradation and enucleation. All non-flowering vascular plants, including gymnosperms, possess sieve cells, which have sieve areas and not sieve plates on both end and side walls41. In gymnosperms, the conducting sieve cells lack adjacent companion cells and are shaped like elongated spindle-like structures that are axially and laterally connected by sieve areas with narrow sieve pores, which leads to higher flow resistance and slower transport rates. Angiosperms develop highly efficient isolated phloem sieve elements with sieve plates on the end walls, which facilitate the fast transport of photoassimilates41,42. Here we identified a new molecular player in root protophloem formation, a small signaling peptide gene CLE33. In the Arabidopsis root, phloem-specific DOF transcription factors induce a set of positive regulators of phloem development, and CLEs, which downregulate DOF expression in a feedback loop9,12. With DOF binding sites in its upstream regulatory sequence, CLE33 is likely involved in this genetic circuit. We demonstrated that CLE33 acts in concert with CLE45 to counteract sieve element differentiation mediated by BRX/OPS. Such a mechanism can potentially have a role in defining the right timing for differentiation of sieve element depending on endogenous cues and environmental conditions. Furthermore, we show that CLE33 acts redundantly with CLE45 to repress the protophloem neighboring cells from differentiating into sieve elements. These results suggest that CLE33/45 act both as autocrine and paracrine signals, whereas the two other phloem CLE genes (CLE25 and CLE26) have a limited role as autocrine signals in the BRX/OPS-BAM3 module regulating protophloem differentiation. Another player in this pathway, RECEPTOR PROTEIN LIKE KINASE 2 (RPK2) was recently shown to be involved in protophloem differentiation43. The loss-of-function rpk2 mutant was able partially rescue the protophloem gap phenotypes and root growth of brx, ops or cvp2cvl1 mutants43. It was shown that RPK2 interacts with BAM1 genetically and the two proteins form heteromeric complexes44, but the possible interaction of RPK2 with BAM3 is not yet explored.
The ability of protophloem sieve element surrounding cells to develop into sieve element cells is important for the developmental plasticity of this tissue43. Laser-ablation of developing protophloem triggers the differentiation of the neighboring cells43, bypassing the original interrupted protophloem to maintain the supply of sugars and hormones to the meristem. It seems, that the right balance between suppressing and keeping the potential for differentiation to sieve elements in case of failure is essential for the protophloem function. It remains to be answered why maintaining only one single functional sieve element file has such a strict genetic control.
In higher vascular plants, the CLAVATA pathway was largely extended both through the number of CLE peptide genes, as well as a new components of the receptor complexes, including duplications of CLV1/BAM- like RLKs and the appearance of receptor-like protein CLAVATA2 and pseudo-kinase CORYNE45. With CLE33 orthologs found in basal angiosperms, it is possible that during plant evolution, phloem-expressed CLEs were recruited to shape the unique angiosperm phloem tissue that functions efficiently in the fast translocation of photoassimilates.
Methods
Search of CLE genes in Arabidopsis thaliana
A t-BLAST-n search in the A. thaliana genome in the EnsemblPlant database was conducted using the 32 previously known full-length CLE protein from A. thaliana as queries. To get more hits, the e value threshold was set to 10 instead of the original setting of 10−1. Candidate sequences that were not annotated as CLE genes were manually compared to the queries. Only sequences similar to the CLE domain were further analyzed for the presence of a signal peptide, proof of expression, and not being part of another described protein. After filtering, one single sequence in an unannotated region of chromosome 1 remained and possessed all the characteristics of a CLE gene: CLE33.
Plant material
All mutants described in this study are in the Arabidopsis thaliana Columbia-0 (Col-0) background: clv2-113 crn-1013, bam1-3 (SALK_015302), bam1-4 (SALK_107290), bam2-4 (SAIL_1053_E09), bam3-2 (SALK_044433)13, bam3-3 (SALK_118860), bam1-4 bam3-3, bam2-4 bam3-2, cle45-246, brx-347, ops-2 (SALK_139316)8.
Generation of CRISPR-Cas9 cle33
The loss-of-function mutants in CLE33 were generated by CRISPR-Cas9 base gene editing. Design of guide-RNAs with low off-targets was performed using CCTop (10.1371/journal.pone.0124633). Generation of the gRNA was achieved by primer dimer annealing (C67xC68; C69xC70) and cloned into the destination vector pAGM5526148 by BsaI cut-ligation. A. thaliana Col-0 wild-type and cle45-2 were transformed by a floral dip in Agrobacterium tumefaciens suspension. Transgenic seeds were screened for DsRed fluorescence. The frameshift-generating mutations in CLE33 were searched in the non-transgenic progeny. Higher-order mutants were obtained by crossing.
Plasmid generation
Genes and promoter regions were amplified by Phusion PCR with the primers indicated in Supplementary Table 1. Plasmids were constructed as indicated in Supplementary Table 2 by Golden Gate modular assembly49.
Phylogenetic and synteny analysis
CLE33/CLE45 and BAM3 homologs were searched by BLASTP in the Phytozome13 and PLAZA Gymnosperm databases. No hints were found in Carica papaya, and a t-BLAST-n was performed to find the CLE33/45 homologs. Tomato CLE sequences were retrieved from24. Full-length protein sequences were aligned using either MUSCLE and manually curated in MEGA-X50 for Fig. 1a, or MAFFT with DASH enabled51 for Supplementary Figs. S9 and S10. The alignments were used to produce trees with 1000 bootstrap replicates with IQTREE52, and visualized with iTOL53. The synteny analysis of the CLE33 in the Brassicaceae was performed with the Genome Context Viewer54. Multi-sequence alignment profiles were composed with alignmentviewer.org55. Sequence conservation logos were generated by WebLogo56.
Root growth assay
Seeds were placed onto 1% sugar half-strength MS plates supplemented or not with the indicated concentration of peptides (Genescript, >75% purity, dissolved in water) or estradiol (stock solution was diluted in DMSO) or corresponding solvent, and kept in dark at 4 °C for 48 h before being transferred in a growth chamber at 22 °C with 16-h-light/8-h-dark cycles. Seedlings were grown for 7 days for root growth inhibition assays by peptides, or 10 days for the genetic suppression of brx-3/ops-2 root growth phenotypes. After scanning the plates with an Epson scanner at high resolution (600dpi), root length was measured using the single-neurite-tracer tool in Fiji.
Promoter activity analysis
The promoter region of CLE33 (876 bp from start codon) was cloned to drive the expression of the GUS reporter system. Transgenic seedlings were incubated at 37 °C in GUS staining buffer (0.5 mg/ml X-Gluc, 100 mM Phosphate buffer, 10 mM EDTA, 1 mM Potassium Ferricyanide, 1 mM Potassium Ferrocyanide, 0.1% Triton X-100) and cleared in 70% ethanol. Images were taken using Zeiss Axioplan microscope and Leica MDG36 stereomicroscope.
For cell-specific expression analysis, upstream regions of CLE26 (1615bp), CLE45 (2513 bp), and CLE33 (876 bp) were cloned and used to drive the expression of nuclear-localized fusion consisting of H2B or NLS and Citrine. For peptide treatment, homozygous transgenic pCVP2:NLS-3xVenus seedlings were transferred to the fresh plates containing no peptide, or 100 nM of CLE33p or CLE45p, 22 h before fixation.
Confocal imaging and sample preparation
Seedlings were fixed with 4% paraformaldehyde (Sigma) in PBS for 1 h minimum. After washing with PBS, samples were stained overnight with a 0.2% Calcofluor White dissolved in a ClearSee solution57, and cleared with ClearSee for at least 48 h. Imaging was performed using a Leica TCS SP5 laser-scanning confocal microscope with the following settings: calcofluor-stained cell wall (excitation at 405 nm, detection at 415–500 nm), Venus or Citrine (excitation at 514 nm, detection at 522–574 nm or 524–600 nm).
Subcellular localization in Nicotiana benthamiana
N. bethamiana leaves were transiently transformed by infiltration with A. tumefaciens carrying a multi-cassette plasmid expressing pUbi:CLE33-mCherry and p35s:Venus. After 3 days, leaf disks were sequentially imaged with a Leica TCS SP5 laser-scanning confocal microscope for Venus (emission at 514 nm, detection from 524 to 551 nm) and mCherry (emission at 561 nm, detection from 571 to 650 nm).
qRT-PCR analysis
Five-day-old seedlings were transferred into new plates containing 1 µM Estradiol or the equivalent amount of solvent (DMSO). After 16 hours, seedlings were shock frozen in liquid nitrogen. Frozen samples were grinded in a mill with metal beads. RNA was extracted using the MagMAX Plant RNA Isolation kit (Applied Biosystems). The remaining DNA was eliminated by a 2 M LiCl precipitation. cDNA synthesis was performed using SensiFAST cDNA synthesis kit (Meridian). Quantitative PCRs were performed with Fast Start Universal SYBR-green Master (Roche), with primers from Supplementary Table 1. The thermal cycler (Mic qPCR Cycler, biomolecular systems) conditions were: 95 °C 2 min, 45 cycles of 95 °C 15 s, 58 °C 10 s, 60 °C 50 s, followed by a dissociation curve analysis. Expression values were normalized to Actin.
Statistics and reproducibility
We performed statistical analyses using R v4.0.2 within the Rstudio interface. We used log transformation for root length and distance QC-protophloem cell identity. Statistical significance was determined by ANOVA followed by a post hoc Tukey test for multi-comparison. For gap cell frequency, we employed a χ2 test with a Benjamini–Hochberg P value correction. In general, the sample size was chosen based on the variability observed in preliminary experiments. For assays involving seedlings grown on plates, multiple plates per conditions were used to limit a plate effect variability. We performed root growth assay and root protophloem analysis at least twice with similar results. The expression pattern of CLE33 was derived from more than a dozen of T1 transcriptional reporter lines. Expression analysis by qPCR was performed once with four technical replicates.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Supplementary datasets supporting this study have been submitted to Dryad and can be found using this link: https://doi.org/10.5061/dryad.x69p8cznw. These datasets include the numerical source data for graphs and charts that can be found in the Excel file, original confocal images in lif and lsm formats, and gene alignments in fas format. All additional data are available from the corresponding author upon reasonable request.
References
Milne, R. J., Grof, C. P. L. & Patrick, J. W. Mechanisms of phloem unloading: shaped by cellular pathways, their conductances and sink function. Curr. Opin. Plant Biol. 43, 8–15 (2018).
Braun, D. M. Phloem loading and unloading of sucrose: what a long, strange trip from source to sink. Annu. Rev. Plant Biol. 73, 553–584 (2022).
Oparka, K. J., Duckett, C. M., Prior, D. A. M. & Fisher, D. B. Real-time imaging of phloem unloading in the root tip of Arabidopsis. Plant J. 6, 759–766 (1994).
Otero, S. et al. A root phloem pole cell atlas reveals common transcriptional states in protophloem-adjacent cells. Nat. Plants 8, 954–970 (2022).
Graeff, M. & Hardtke, C. S. Metaphloem development in the Arabidopsis root tip. Development 148, dev199766 (2021).
Roszak, P. et al. Cell-by-cell dissection of phloem development links a maturation gradient to cell specialization. Science 374, eaba5531 (2021).
Esau, K., Cheadle, V. I. & Gifford, E. M. Comparative structure and possible trends of specialization of the phloem. Am. J. Bot. 40, 9–19 (1953).
Truernit, E., Bauby, H., Belcram, K., Barthélémy, J. & Palauqui, J. C. OCTOPUS, a polarly localised membrane-associated protein, regulates phloem differentiation entry in Arabidopsis thaliana. Development 139, 1306–1315 (2012).
Qian, P. et al. A Dof-CLE circuit controls phloem organization. Nat. Plants 8, 817–827 (2022).
Depuydt, S. et al. Suppression of Arabidopsis protophloem differentiation and root meristem growth by CLE45 requires the receptor-like kinase BAM3. Proc. Natl Acad. Sci. USA 110, 7074–7079 (2013).
Rodriguez-Villalon, A. et al. Molecular genetic framework for protophloem formation. Proc. Natl Acad. Sci. USA 111, 11551–11556 (2014).
Miyashima, S. et al. Mobile PEAR transcription factors integrate positional cues to prime cambial growth. Nature 565, 490–494 (2019).
Hazak, O. et al. Perception of root-active CLE peptides requires CORYNE function in the phloem vasculature. EMBO Rep. 18, 1367–1381 (2017).
Hu, C. et al. A CLE-BAM-CIK signalling module controls root protophloem differentiation in Arabidopsis. N. Phytol. 233, 282–296 (2022).
Ottoline Leyser, H. M. & Furner, I. J. ‘Characterisation of three shoot apical meristem mutants of Arabidopsis thaliana’. Development 116, 397 (1992).
Clark, S. E., Running, M. P. & Meyerowitz, E. M. CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development 121, 2057–2067 (1995).
Fletcher, J. C., Brand, U., Running, M. P., Simon, R. & Meyerowitz, E. M. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283, 1911–1914 (1999).
I. The Arabidopsis Genome. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408, 796–815 (2000).
Sawa, S., Kinoshita, A., Betsuyaku, S. & Fukuda, H. A large family of genes that share homology with CLE domain in Arabidopsis and rice. Plant Signal Behav. 3, 337–339 (2008).
Cock, J. M., McCormick, S. & Large, A. Family of genes that share homology with CLAVATA3. Plant Physiol. 126, 939–942 (2001).
Sharma, V. K., Ramirez, J. & Fletcher, J. C. The ArabidopsisCLV3-like (CLE) genes are expressed in diverse tissues and encode secreted proteins. Plant Mol. Biol. 51, 415–425 (2003).
Strabala, T. J. et al. Gain-of-function phenotypes of many CLAVATA3/ESR genes, including four new family members, correlate with tandem variations in the conserved CLAVATA3/ESR domain. Plant Physiol. 140, 1331–1344 (2006).
Haas, B. J. et al. Full-length messenger RNA sequences greatly improve genome annotation. Genome Biol. 3, research0029.0021 (2002).
Carbonnel, S., Falquet, L. & Hazak, O. Deeper genomic insights into tomato CLE genes repertoire identify new active peptides. BMC Genomics 23, 756 (2022).
Anne, P. et al. CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis. Development 145, dev162354 (2018).
Jun, J. et al. Comprehensive analysis of CLE polypeptide signaling gene expression and overexpression activity in Arabidopsis. Plant Physiol. 154, 1721–1736 (2010).
Rhodes, J. et al. Perception of a conserved family of plant signalling peptides by the receptor kinase HSL3. eLife 11, e74687 (2022).
Liu, Z. et al. Phytocytokine signalling reopens stomata in plant immunity and water loss. Nature 605, 332–339 (2022).
Ghorbani, S. et al. Expanding the repertoire of secretory peptides controlling root development with comparative genome analysis and functional assays. J. Exp. Bot. 66, 5257–5269 (2015).
de Bang, T. C. et al. Genome-wide identification of Medicago peptides involved in macronutrient responses and nodulation. Plant Physiol. 175, 1669–1689 (2017).
Okuda, S. et al. Molecular mechanism for the recognition of sequence-divergent CIF peptides by the plant receptor kinases GSO1/SGN3 and GSO2. Proc. Natl Acad. Sci. USA 117, 2693–2703 (2020).
Patel, N. et al. Diverse peptide hormones affecting root growth identified in the Medicago truncatula secreted peptidome. Mol. Cell Proteom. 17, 160–174 (2018).
Liang, Y., Zhu, W., Chen, S., Qian, J. & Li, L. Genome-wide identification and characterization of small peptides in maize. Front. Plant Sci. 12, 695439 (2021).
Eddy, S. R. Accelerated profile HMM searches. PLoS computational Biol. 7, e1002195 (2011).
Gancheva, M. S. et al. Polymorphism of CLE gene sequences in potato. Vavilovskii Zh . Genet Selektsii 25, 746–753 (2021).
Morgan, A. A. & Rubenstein, E. Proline: the distribution, frequency, positioning, and common functional roles of proline and polyproline sequences in the human proteome. PLoS ONE 8, e53785 (2013).
Shinohara, H., Moriyama, Y., Ohyama, K. & Matsubayashi, Y. Biochemical mapping of a ligand-binding domain within Arabidopsis BAM1 reveals diversified ligand recognition mechanisms of plant LRR-RKs. Plant J. 70, 845–854 (2012).
Stührwohldt, N., Ehinger, A., Thellmann, K. & Schaller, A. Processing and formation of bioactive CLE40 peptide are controlled by posttranslational proline hydroxylation. Plant Physiol. 184, 1573–1584 (2020).
Roman, A.-O. et al. HSL1 and BAM1/2 impact epidermal cell development by sensing distinct signaling peptides. Nat. Commun. 13, 876 (2022).
Esau, K. The phloem. (Gebruder Borntraeger Verlagsbuchhandlung, Science Publishers, Germany, 1969).
Simpson, M. G. “4—Evolution and diversity of vascular plants”. In Plant Systematics, Third Edition. (ed. Simpson, M. G.) 75–130 (Academic Press, 2019).
Liesche, J. & Schulz, A. Phloem transport in gymnosperms: a question of pressure and resistance. Curr. Opin. Plant Biol. 43, 36–42 (2018).
Gujas, B. et al. A reservoir of pluripotent phloem cells safeguards the linear developmental trajectory of protophloem sieve elements. Curr. Biol. 30, 755–766.e754 (2020).
Shimizu, N. et al. BAM 1 and RECEPTOR-LIKE PROTEIN KINASE 2 constitute a signaling pathway and modulate CLE peptide-triggered growth inhibition in Arabidopsis root. N. Phytol. 208, 1104–1113 (2015).
Miwa, H., Tamaki, T., Fukuda, H. & Sawa, S. Evolution of CLE signaling: origins of the CLV1 and SOL2/CRN receptor diversity. Plant Signal Behav. 4, 477–481 (2009).
Yamaguchi, Y. L. et al. A collection of mutants for CLE-peptide-encoding genes in arabidopsis generated by CRISPR/Cas9-mediated gene targeting. Plant Cell Physiol. 58, 1848–1856 (2017).
Rodrigues, A. et al. The short-rooted phenotype of the brevis radix mutant partly reflects root abscisic acid hypersensitivity. Plant Physiol. 149, 1917–1928 (2009).
Grützner, R. et al. High-efficiency genome editing in plants mediated by a Cas9 gene containing multiple introns. Plant Commun. 2, 100135 (2021).
Binder, A. et al. A modular plasmid assembly kit for multigene expression, gene silencing and silencing rescue in plants. PLoS ONE 9, e88218 (2014).
Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549 (2018).
Katoh, K., Rozewicki, J. & Yamada, K. D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinforma. 20, 1160–1166 (2017).
Trifinopoulos, J., Nguyen, L.-T., von Haeseler, A. & Minh, B. Q. W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 44, W232–W235 (2016).
Letunic, I. & Bork, P. Interactive tree of life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–W296 (2021).
Cleary, A. & Farmer, A. Genome Context Viewer: visual exploration of multiple annotated genomes using microsynteny. Bioinformatics 34, 1562–1564 (2017).
Reguant, R. et al. AlignmentViewer: sequence analysis of large protein families. F1000Res. 9, (2020).
Crooks, G. E., Hon, G., Chandonia, J. M. & Brenner, S. E. WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190 (2004).
Kurihara, D., Mizuta, Y., Sato, Y. & Higashiyama, T. ClearSee: a rapid optical clearing reagent for whole-plant fluorescence imaging. Development 142, 4168–4179 (2015).
Acknowledgements
We thank Dr. Bojan Gujas for sharing transgenic line (pCVP2:NLS-Venus) and technical advice for phloem microscopy. We thank the Bioimage Core Facility at the University of Fribourg and Adolphe Merkel Imaging Facility. This work was funded by Ambizione SNSF Grant (PZ00P3_179745) to O.H., COST SNSF Grant (IZCOZ0_189892) to O.H. and additional funding provided by the University of Fribourg to O.H.
Author information
Authors and Affiliations
Contributions
S.Car. and O.H. designed research; S.Car. and S.Cor. performed research; S.Car analyzed the data and prepared figures; O.H. wrote the first draft; O.H., S.Car., and S.Cor. together edited the manuscript; O.H. supervised project; O.H. acquired funding.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
This manuscript has been previously reviewed at another Nature Portfolio journal. Communications Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: David Favero.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carbonnel, S., Cornelis, S. & Hazak, O. The CLE33 peptide represses phloem differentiation via autocrine and paracrine signaling in Arabidopsis. Commun Biol 6, 588 (2023). https://doi.org/10.1038/s42003-023-04972-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-023-04972-2
- Springer Nature Limited