
Abstract
The Western honey bee Apis mellifera, which provides about 90% of commercial pollination, is under threat from diverse abiotic and biotic factors. The ectoparasitic mite Varroa destructor vectoring deformed wing virus (DWV) has been identified as the main biotic contributor to honey bee colony losses worldwide, while the role of the microsporidium Nosema ceranae is still controversially discussed. In an attempt to solve this controversy, we statistically analyzed a unique data set on honey bee colony health collected from a cohort of honey bee colonies over 15 years and comprising more than 3000 data sets on mite infestation levels, Nosema spp. infections, and winter losses. Multivariate statistical analysis confirms that V. destructor is the major cause of colony winter losses. Although N. ceranae infections are also statistically significantly correlated with colony losses, determination of the effect size reveals that N. ceranae infections are of no or low biological relevance.
Similar content being viewed by others

Introduction
The basis of human nutrition includes agriculturally grown crops and fruits, many of which are dependent on insect pollination for fruit set, seed production, and yield. Managed and wild insect pollinators, therefore, play a key role in ensuring that mankind is adequately supplied with food1,2,3. As a result, the health and survival of pollinating insects have attracted increasing public and scientific interest and consequently diseases as well as disease-causing pathogens and parasites that threaten pollinating insects have become the focus of many research studies. For pollinating insects, the main focus is on the Western honey bee Apis mellifera L., which is managed by beekeepers for honey production all over the world and provides 90% of the commercial pollination worldwide2. For pathogens and parasites, the focus is on those that threaten the survival of the managed honey bee colonies. The ectoparasitic mite Varroa destructor Anderson and Trueman together with deformed wing virus (DWV) vectored by the mite have been identified as the main contributors to colony losses4,5,6,7. The microsporidium Nosema ceranae Fries (N. ceranae) has also been implicated in regional colony losses8,9,10,11,12. The threat posed by these pathogens is compounded by the fact that honey bee colonies are usually infected by several pathogens simultaneously, with V. destructor (together with DWV) and Nosema spp. being the most widespread and therefore often occurring together7,13,14,15,16.
The mite V. destructor is an ectoparasite of honey bees (Apis mellifera, A. cerana F.) that infests honey bee colonies all over the world (for a recent review on V. destructor please see ref. 17). The life cycle of V. destructor in honey bee colonies is divided into two phases, (i) the dispersal phase in which adult female mites parasitize adult bees and use the bees as a means of transport and (ii) the reproductive phase that takes place in the capped brood cell18. For reproduction, a sexually mature, mated female mite enters a brood cell just before cell capping. When the honey bee larva reaches the prepupal stage, the mother mite begins laying eggs and raising her offspring. For feeding, the mother mite punctures a hole in the cuticle of the developing bee. This hole is then the feeding site for the growing mite family and allows access to the pupa’s fat body, which serves as nutritional resource19. Bees developing from V. destructor parasitized pupae show accelerated behavioral maturation, resulting in a shortened phase as nurse bees20, contribute less to colony productivity, and have a reduced longevity18. Heavily mite-infested colonies are characterized by an increasing rate of emerging bees which are not viable and have crippled wings21. Initially, these symptoms were thought to be caused solely by mite parasitization, but it soon became clear, that V. destructor is an efficient virus vector22,23 and that the crippled wings syndrome was caused by a virus, which was originally isolated as Egypt bee virus (EBV), but then renamed deformed wing virus (DWV)24,25,26,27. We now know that at least four major variants of DWV exist27,28,29,30 and that it is the variant DWV-B that causes most of the symptoms, is more virulent than the DWV-A, and uses V. destructor as biological vector31,32,33,34,35,36. Although V. destructor itself is sufficient to cause considerable damage to the parasitized pupa and the infested colony, it is the mite-vectored viruses, particularly deformed wing virus (DWV), that exacerbate the damage and link mite infestation to colony losses, especially during the winter season5,7,29,34,37,38.
Microsporidia are fungal-related, obligate intracellular parasites that infect many vertebrate and invertebrate host species39. Three microsporidian species infecting the adult Western honey bee A. mellifera are described: Nosema apis, N. neumanni n.sp., and N. ceranae. While N. apis is known as a honey bee-specific pathogen since more than 100 years40, N. neumanni was described as pathogen of A. mellifera in Uganda only recently41. N. ceranae in contrast was originally described as pathogen of the Eastern honey bee Apis cerana42, but obviously switched host several decades ago and by now is even more prevalent than N. apis in many A. mellifera populations43,44,45,46,47. However, a recent study has shown that there is still no general replacement of N. apis by N. ceranae, but replacement rather seems to be a regional phenomenon48, presumably influenced in its dynamics by climatic conditions since N. ceranae spores quickly lose their infectivity when exposed to low temperatures49,50,51.
From a clinical point of view, there is not much difference between N. apis and N. ceranae: Both pathogens follow a seasonal pattern and often cause asymptomatic infections of the midgut epithelium of adult bees48. Both pathogens can also cause diarrhea52, although the factors that cause the transition from asymptomatic to symptomatic infections are poorly understood53,54. Symptomatic outbreaks of Nosema spp.-infections are called nosemosis and can be diagnosed by the characteristic fecal spots visible at the hive entrance and inside the hive52. These fecal spots contain millions of infectious spores and drive the fecal-oral transmission of the disease within the colony, as adult bees cleaning the hive of the spots ingest the spores and become infected26,55. Infection in the individual adult bee host is initiated by germination of the ingested spores in the midgut lumen; germination is followed by extrusion of the polar tube, mechanical piercing of a cell by this polar tube, and injection of the sporoplasm into the cell through the polar tube56,57. The reproductive cycle within the infected host cell takes about 96 h58, goes through several stages (merogony, sporogony) and ends when the newly generated spores are released into the gut lumen by the bursting of the cell. These newly generated spores in the gut lumen are defecated and can infect naïve adult bees when they try to clean the hive from spore-contaminated fecal spots26,55,59.
In 2008, the first study was published suggesting that even asymptomatic N. ceranae infections result in the collapse of honey bee colonies9. Since then, numerous studies have been published demonstrating an association between N. ceranae infections and colony losses, but there are also many studies that failed to confirm this association8,9,10,11,12,60,61,62,63.
Among the studies that did not observe a statistically significant association between N. ceranae infection and colony losses is our longitudinal cohort study on Nosema spp. epidemiology and honey bee health in Northeast Germany48,50. In the course of a monitoring project, which was initiated in 2005 and is still ongoing, an extensive data set on colony health was continuously collected from a cohort of honey bee colonies, including data on colony losses during the winter season, prevalence of Nosema spp.-infection in autumn, and V. destructor infestation levels in autumn. We analyzed this unique 15-year data set comprising data on more than 3000 honey bee colonies using diverse uni- and multivariate statistical methods in order to investigate the relationship between colony mortality and the two pathogens that are most commonly blamed for colony losses, V. destructor and N. ceranae. We confirm that V. destructor is the main cause of colony winter losses, even in N. ceranae-infected colonies. However, in nearly mite-free colonies, a statistically significant association was demonstrated between N. ceranae infection in autumn and colony losses in the following winter. However, calculating the effect size of this correlation revealed that it has no biological relevance, since nowadays colonies not affected by V. destructor are rarely found. Thus, although N. ceranae is in principle capable of killing entire colonies, this effect is usually masked by the more dominant deleterious effect of the ubiquitous ectoparasite V. destructor. Our results end a long-standing controversy about the virulence of N. ceranae at the honey bee colony level and whether N. ceranae should be considered a serious threat to honey bee colonies.
Results
Winter mortality
Over the last 15 years, we performed a longitudinal cohort study on Nosema spp. epidemiology and colony health in the honey bee population in Northeast Germany7,48,50. We monitored between 180 and 270 colonies each year throughout the entire study period and continuously collected data on winter mortality as well as on Nosema spp. infection status and Varroa destructor infestation levels in the monitored colonies. The full data set for this study comprises 3502 colonies and is a uniquely solid basis for analyzing both, the dynamics of winter colony losses and the relation between Nosema spp.-infection and V. destructor-infestation in autumn with colony losses in the following winter.
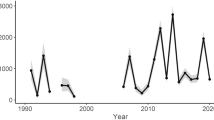
Within the study period, the winter colony loss rate varied from 4.8% as the lowest value in winter 2008/2009 to 26.0% as the highest value in winter 2016/2017. Using a linear model, we show that colony winter mortality increased by about 0.5 % per year in the studied cohort (Fig. 1a), but that this increase between 2005/2006 and 2019/2020 was not statistically significant (p-value of the F-statistic = 0.223, adjusted R2 = 0.043). The mean of the average winter losses was 16.31% ± 6.56% (mean ± SD) and therefore statistically significantly higher (p-value = 0.0023; one-sample t-test) than the empirical threshold for acceptable winter mortality of 10%64.

Honey bee colony losses over winter (a), V. destructor infestation levels in October (b), and N. ceranae infection prevalence in autumn (c) were monitored over the study period from 2005 to 2020. Each data point represents the proportion of dead colonies in spring (n = 3502), the mean mite load per 100 bees in October (n = 3492) or the prevalence of N. ceranae-infected colonies in autumn (n = 3502) per year. Linear regression models were calculated. Their regression lines are shown and their 95% CI (confidence interval) are highlighted in light red.
Long-term dynamics of Varroa destructor infestations and Nosema ceranae infections
It is well proven and generally accepted that the mite V. destructor is the main cause of honey bee colony losses in winter and that mite infestation levels in autumn correlate with colony loss rates in the following winter [for a recent review see ref. 17]. We therefore monitored each colony for its mite infestation level in October over the entire study period 2005–2020 by counting the mites present in a bee sample of about 150 bees and expressing the infestation level as number of mites per 100 bees. The mean mite infestation rate varied between 7.26 ± 2.59 (mean ± SEM) in 2005 and 2.09 ± 1.00 (mean ± SEM) in 2015 and was unchanged throughout the study period according to the used linear model (p-value of the F-statistic = 0.7739, adjusted R2 = 0.006) (Fig. 1b). Hence, no statistically significant increase or decrease in mite infestation levels was observed.
We recently demonstrated that N. ceranae infection prevalence in autumn showed a statistically significant increase between 2005 and 201548. We again analyzed the dynamics of N. ceranae infection prevalence in autumn over the entire study period of meanwhile 15 years using a linear model and demonstrate a steady increase in N. ceranae infection prevalence which was statistically significant (p-value of the F-statistic = 0.021, adjusted R2 = 0.295) and increased by about 0.5 % per year (Fig. 1c).
V. destructor infestation and winter mortality
Although we observed a steady, though not significant, increase in winter losses in the cohort of monitored colonies over the 15-year observation period (Fig. 1a), there was no comparable trend in mean mite infestation rates (Fig. 1b). Nevertheless, a significant relationship between V. destructor parasitization and honey bee colony mortality could be established by comparing the mean mite infestation rates in colonies that died over the winter or survived the winter season. Calculated for the entire data set (n = 3492 colonies), the mean mite infestation rate in autumn was significantly higher (one-sample t-test, p < 0.0001) in colonies that collapsed during winter (13.67% ± 4.84%, mean ± SEM) than in colonies that were still alive in spring (3.27% ± 1.69%, mean ± SEM). Looking at the data for each year individually (Table 1) revealed that this significant overall relationship was true for 13 of the 15 years observed. Only in two winter seasons (2008/2009 and 2015/2016) with very low overall loss rates (4.8% and 7.5%, respectively) mite infestation level was not correlated with winter loss (Fig. 1a, Table 1). There is a simple explanation for this phenomenon: if winter losses are below a biological threshold (natural colony mortality of about 10%64), chances that V. destructor killed the colonies are low.
Classification tree analysis
Our results reconfirmed what is already known from many other studies, namely that V. destructor is the main driver of honey bee winter mortality. However, N. ceranae has also been implicated in colony losses in many studies, although we have not yet been able to confirm this in our previous analyses of our cohort study data: A possible explanation is that these previous analyses were based on univariate analyses that observe the effect of single explanatory variables on colony mortality but are unable to estimate the relative influence of V. destructor and N. ceranae on colony mortality when both are present simultaneously.
To close this gap of knowledge, we used our data set comprising for each colony qualitative (yes/no) data on colony winter losses, quantitative data on V. destructor infestation levels and both qualitative (infecting species) and semi-quantitative (categorized spore load) data on Nosema spp.-infection status and performed a classification tree analysis (Fig. 2), a tool of recursive partitioning for multivariate data exploration. This multivariate analysis aimed at identifying the relative impact of V. destructor infestation levels and Nosema spp. infection status (infecting species, spore load category) in autumn on the fate of the colony over winter, hence, whether a colony was prone to collapse over winter or had a realistic chance to survive.

The proportion of colonies represented by each circle (leaf/outcome) is shown above the respective circle. In each circle, the gray parts represent the proportions of overwintered (survived) colonies and the red parts the proportions of collapsed colonies. The number of the collapsed and survived colonies are given next to each circle. The tested attribute that resulted in each of the two nodes is given below the circles (mite infestation rate). Horizontal lines represent the branches/decisions and the numbers within these lines show the mite infestations rates that best splitted the data to obtain the next nodes or leafs. For classification tree analysis, the R packages rpart81 and rattle82 with standard settings were used.
A total of 3492 data sets was available for this analysis. Mite infestation rate was identified as the attribute that best classified our data set and hence became the root node (Fig. 2). Performing a top-down search, two decision points (nodes) describing the fate of the colony (survival or collapse) were identified (Fig. 2). Both were solely based on the attribute “mite infestation level” indicating that the also tested attributes regarding the Nosema spp.-infection status of the colonies (infecting species and spore load category) were not identified by this analysis as decisive factor for colony collapse over winter. The first decision point divided the analyzed colonies into two groups, one comprising 82% (n = 2861) of the colonies with a mite infestation rate in October of less than eight mites per 100 bees and a mortality rate of 9.8%. The remaining 18% of the colonies (n = 631) had an infestation level of eight or more mites per 100 bees and a mortality rate of 45.8%. This group was further subdivided into two groups, again based on the mite infestation level. 12% (n = 430) had an infestation level of eight or more mites but below 21 mites per 100 bees and a mortality rate of 37.2%, whereas the other branch comprised the remaining 6% (n = 201) of the colonies which were characterized by an infestation level of 21 or more mites per 100 bees and a mortality rate of 64.2%. This classification tree showed convincingly that over the study period of 15 years, the strongest link was between colony winter losses and V. destructor infestation level, but not with N. ceranae or N. apis infection.
N. ceranae infection and winter mortality
The results from the classification tree analysis were in accordance with previous studies that did also not reveal any relation between colony winter losses and Nosema spp. infection7,48,50,63,65,66, but contradicted other studies repeatedly showing that N. ceranae infections cause colony losses9,10,12,67,68. A striking feature of our data set is that both winter losses (Fig. 1a) and N. ceranae prevalence (Fig. 1c) each increased by 0.5 % per year over the observation period of 15 years. This coincidence led us to suspect that there was, after all, a link between winter losses and N. ceranae infections that we had previously overlooked7,48,50. Fact is that Nosema spp.-infections usually show a rather low prevalence in autumn26,48, and hence our data set for each autumn comprises only a few colonies infected with Nosema spp. and even fewer colonies infected with N. ceranae or representing a particular spore load category. We, therefore, speculated that the number of Nosema spp. infected colonies per year might be too low to see a relation between winter mortality and Nosema spp. infection. Hence, we decided to generate higher numbers for Nosema infected colonies by ignoring both the species differentiation and the infection categories and summing the numbers for the individual years starting with autumn/winter 2005/2006 and ending with the sum of autumn/winter 2005/2006 up to autumn/winter 2019/2020 (Table 2). Forming these cumulative data subsets and examining time periods instead of annual values, resulted in larger group sizes (n’s) and overall larger numbers of Nosema spp. infections, which made statistical analyses more robust (Table 2). We used a Chi-squared test to calculate the statistical relationship between infection status and winter mortality and indeed, the analysis started to become significant (p-value < 0.05) when more than 11 years (2005/2006 up to 2016/2017 resulting in more than 221 Nosema spp.-infected colonies among the analyzed 3492 colonies) were considered (Table 2, Fig. 3).

Each data point represents the cumulative number of infected colonies summed over the time window indicated on the x-axis. The total numbers per time window are given in Table 2. The y-axis gives the respective p-value of the χ2 test.
To further support and analyze this impact, we performed a Chi-squared test giving Pearson residuals using the data cumulated over the entire study period (2005/2006 to 2019/2020) for Nosema spp. infection (Fig. 4a) and for N. apis, N. ceranae and mixed infections separately (Fig. 4b). The black lines in the associations plots (Fig. 4) represent the expected values for the categories “alive” or “dead” in relation to the infection categories Nosema spp.-negative or -positive (Fig. 4a) or negative and positive for N. apis-, N. ceranae- or co-infections (Fig. 4b). Overrepresented categories are represented by rectangles above the base line, while the rectangles for underrepresented categories are below the base line. Categories with Pearson residuals above 2.0 are shown in blue and appear only for the combinations “dead” and “positive for Nosema spp.-infection” (Fig. 4a) or “dead” and “positive for N. ceranae infection” (Fig. 4b). These results confirmed that the relation between Nosema spp. infection and winter colony losses was statistically significant (p-value = 0.007). However, this was only true for N. ceranae-infections (p-value = 0.002); N. apis- or co-infections did not significantly contribute to winter losses suggesting that N. ceranae is indeed more virulent at colony level than N. apis.

Association plots based on Chi-squared testing show the association between a Nosema spp.-infection and colony mortality and b N. apis-, N. ceranae and co-infections and colony mortality. Overrepresented entries (dead/positive in a or dead/N. ceranae in b) are shown in blue, non-overrepresented entries in gray.
Statistical significance vs. biological relevance
P-values are a measure of statistical significance, but are insufficient to show biological relevance. To analyze the biological relevance of our results, we therefore used the effect size measure Cohen’s ω, which is applicable for two times two and larger contingency tables and is considered a measure of relevance with values above 0.1, 0.3, and 0.5 indicating a small, medium, and large effect size, respectively69. Calculating Cohen’s ω for our data set and the relation between colony losses and Nosema spp.- or N. ceranae-infection revealed an effect size below 0.1, hence, a less than small effect (Fig. 5a). These results indicated that although we showed a statistically significant relationship between colony losses and Nosema spp. infection, particularly N. ceranae infection, these relationships are of minor or no biological relevance supporting the results of the classification tree analysis (Fig. 2) identifying V. destructor as main factor in colony losses.

a Bar plot showing the effect size Cohen’s ω for Nosema spp.- (blue bar) and N. ceranae-infected (red bar) colonies on colony mortality. Dashed lines indicate the conventional definition of Cohen’s ω with a value for ω between 0.1 and 0.3 as small, between 0.3 and 0.5 as medium and above 0.5 as large effect69. b Bar plot showing the relation between colony mortality, mite infestation level categories (mites per 100 bees) and N. ceranae-infection status (gray bars = not infected with N. ceranae, red bars = infected with N. ceranae). Significance levels, which resulted from p-values of the χ2-tests, are indicated by asterisks (n.s., p ≥ 0.05; significantly different: *0.05 < p < 0.01; **0.01 < p < 0.001; ***0.001 > p).
We therefore reanalyzed the data, but this time we considered the V. destructor infestation rate of the colonies by analyzing the mortality rate of colonies infected and not infected with N. ceranae (Fig. 5b) within the mite infestation categories as defined by the classification tree (Fig. 2). This analysis revealed that N. ceranae infection only contributed significantly to colony mortality when the colonies did not harbor detectable V. destructor mites or very few mites (1-7 per 100 bees) in October. Only in these 149 out of 3474 colonies, N. ceranae was significantly correlated with winter mortality (Fig. 5b).
Discussion
Honey bee colony losses and the quest for abiotic and biotic factors causing them are hot topics in the field of bee research since two decades. While it is widely accepted and unequivocally substantiated by many studies, that the ectoparasitic mite V. destructor and the viruses vectored by the mite play a key role in winter colony losses, the role of N. ceranae infections is less clear. There are studies clearly showing detrimental effects of N. ceranae infection on honey bee colonies8,9,10,11,12. But there are also numerous monitoring studies that fail to observe such an association between N. ceranae infection and colony losses48,50,60,61,62. One possible reason for this could be that in monitoring studies the damage caused by the almost ubiquitous infestation of colonies with V. destructor masks the effects caused by pathogens with rather low prevalence such as N. ceranae. This masking effect is difficult to see through because most observational studies on colony losses include too few colonies and are conducted over too short a time period to observe statistically significant associations for low-prevalence pathogens. Moreover, although most monitoring studies collect data for multiple pathogens, more complex or multivariate analyses on the interaction between pathogens or their joint effect and relative influence on colony mortality are rarely performed. There is one study analyzing the interaction of pathogens and showing, that N. ceranae-infections in spring correlate statistically significantly with an increased prevalence of Ascosphaera apis infections and higher levels of V. destructor infestation in summer70. However, data on the relative impact of individual co-infecting pathogens on colony mortality in the field are lacking, although it is widely accepted that colony collapse is a multifactorial process, often likely involving more than one pathogen13.
Our data on V. destructor and Nosema spp. load and winter colony mortality, collected continuously over 15 years from a relatively stable cohort of about 25 apiaries contributing ten colonies each is a unique resource to study the role of V. destructor and N. ceranae on colony mortality in the field, especially the relative impact of these two parasites on overwintering success of honey bee colonies, most of which were concurrently infested by V. destructor and infected with Nosema spp.
With more than 3000 data sets collected over 15 years, we were able to confirm that big data sets and long study durations are key for robust analyses: Only by summing the data for more than 11 years did a statistically significant association between N. ceranae-, but not N. apis-infection in the autumn and colony losses the following winter become evident. This result indicated that at colony level N. ceranae is indeed more virulent than N. apis confirming previous studies on N. ceranae-induced colony losses8,9,10,11,12. No such relation between N. ceranae-infections and colony losses was observed when the data were analyzed year by year48,50 because the prevalence of Nosema spp.-infections in autumn and the mortality rate among these colonies are usually too low for statistical significance. This in turn explains previous studies that did not support N. ceranae-induced colony losses48,50,60,61,62.
A statistically significant association between N. ceranae infection and colony losses was in accordance with studies that had suggested an increased colony virulence of N. ceranae compared to N. apis and the abilty of N. ceranae to cause the collapse of entire colonies8,9,10,11,12. However, our multivariate data exploration via classification tree analysis had identified V. destructor infestation as the main variable explaining colony losses in our cohort. A result that is also in accordance with many other studies clearly linking mite infestation, but not N. ceranae infection, to winter colony mortality5,7,29,34,37,38. Only with a mite infestation rate in October of less than eight mites per 100 bees, an acceptable and natural winter mortality rate below 10%64 can be reached. In the monitored cohort this was the case for 82% of the colonies over the entire duration of the study, leaving 18% of the cohort contributing to unacceptably elevated colony losses. Since classification tree analysis is the method of choice for determining biologically relevant factors and the classification tree identified mite infestation as the only relevant factor, it was not surprising that the determination of Cohen’s ω confirmed that the biological relevance of N. ceranae infection for colony losses is low despite the statistical significance of this association. This clearly shows that for biological questions the focus should rather not solely lay on statistical significance but more on the effect size and biological relevance.
Hence, our results confirmed that V. destructor is the major cause of colony winter losses, although N. ceranae infections can also have deleterious effects. However, these effects are normally masked by the more severe effects of V. destructor on colony health and therefore only detectable in colonies that are not infested with mites or are infested at low levels. As long as V. destructor infestation is the dominating health problem in honey bee colonies and N. ceranae prevalence is low, N. ceranae can be classified as pathogen causing little concern, because its role in colony losses is marginal. Therefore, we do not yet consider N. ceranae a serious threat to honey bee colonies. However, the situation might change when the prevalence of N. ceranae reaches a critical point. Since the increase in N. ceranae prevalence is continuing (ref. 48 and this study), it might only be a question of time when this point will be reached. Monitoring not only mite infestation levels in colonies but also N. ceranae infection prevalence in honey bee populations is therefore advisable. Hence, we will continue our study and continuously calculate the effect size of N. ceranae infection on colony losses to determine the critical prevalence of N. ceranae in a honey bee population.
Remarkable is that in an early report on colony collapse due to N. ceranae10 it was explicitly pointed out that mites were absent in all samples indicating a very low number or even the total absence of V. destructor in these collapsed colonies due to efficient mite control. The absence of V. destructor and concomitant presence of N. ceranae was the most convincing argument for N. ceranae being the cause of colony collapse in the reported case. Moreover, for experimentally demonstrating N. ceranae-induced colony collapse, the colonies needed to be tightly controlled for V. destructor infestation9. These studies indirectly corroborate our results which indicated that N. ceranae-induced colony collapse only becomes evident in (nearly) mite-free colonies. Hence, one can say that the more efficient the mite control is, the more likely it is that N. ceranae induced damages become detectable. But again, as long as the prevalence of N. ceranae is low, this does not pose a serious threat because N. ceranae is simply not a highly virulent pathogen. With these results we end a long-standing controversy about whether N. ceranae is capable of killing entire honey bee colonies—yes, it is under certain circumstances—and should be considered a serious threat for honey bees in general—no, rather not as long as V. destructor infestations are the dominating health problem.
Methods
Bee samples, field survey
The data set of this study comprises data and samples which were collected from autumn 2005 to spring 2020 in the course of a 15 year longitudinal cohort-study on Nosema spp. epidemiology and honey bee health (Supplementary Data 1)7,48,50. Honey bee samples were collected in autumn and colonies were checked for their survival in spring of the respective overwintering period (week 36 to week 14 of the following year) from about 23 apiaries which were located in Northeast Germany (Fig. 6). Briefly, apiaries participated with ten so called “monitoring colonies” each. Apiaries or monitoring colonies that dropped out during the study period were substituted by adequate replacement. Hence, more than half of the apiaries (14 of ~23) participated for more than 9 years and 5 of them even for the entire duration of the study, i.e., 15 years. From at least 19 apiaries, samples were provided over a time period of consecutive 5–11 years (Fig. 6). This resulted in an annual mean of 23.4 ± 2.26 (mean ± SD) apiaries with 9.77 ± 1.25 (mean ± SD) colonies each, giving an overall count of n = 3502 sampled monitoring colonies which provide the basis of our analyses.

Northeast Germany is shown in gray with differently sized circles in different shades of blue showing the locations of the apiaries. The size and color of the circles represent the number of years for which data are available for each apiary (light blue, 12–15 years; gray-blue, 9–11 years; royal blue, 5–8 years; dark blue, 1–4 years). The map of Northeast Germany was sourced from the Database of Global Administrative Areas (http://GADM.org)86.
Sampling of bees for subsequent pathogen analysis was performed between calendar week 36 and 38 (late September/beginning of October)48,50. About 300 in-hive honey bees were sampled from a super above the queen excluder71 from each monitoring colony. Bee samples were stored at −20 °C until further analysis. At least 150 bees were used for determining V. destructor infestation levels, 20 bees were used for determining the Nosema spp. infection status and the remaining bees were stored as retention sample in case analyses had to be repeated.
Determination of mite infestation levels
For determining the mite infestation level of a colony, V. destructor mites were washed from about 150 sampled bees following a standard protocol using a detergent solution72. Briefly, the frozen bees were covered with soap and water in a jar. Subsequently, the jar was shaken for 20 s and emptied into two stacked sieves with a white nylon cloth between them. All mites were rinsed with plenty of water under high pressure through the upper sieve which has larger apertures (3–4 mm) not allowing bees to pass. All mites were collected on the cloth in the second sieve, which has smaller apertures (<0.5 mm) that no mite fits through. To gain the mite infestation level of an individual colony in %, the number of counted mites was divided by the number of sampled and washed bees multiplied by 100. The mite infestation levels for each winter season (mean percentage of mites per year; Table 1) for the three categories “all colonies”, “survived colonies”, and “collapsed colonies” were calculated on the basis of the individual colony values by calculating the mean (±SD) over all colonies of the respective category in the respective winter season. The mean (±SEM) mite infestation levels for the entire study period for each of the three categories were calculated on the basis of these annual means (Table 1). Unfortunately, mite infestation rate could not be determined in one apiary (ten colonies) in the first year (2005/2006) because of an insufficient number of honey bees available. Therefore, the dataset for mite infestation level had to be reduced from 3502 to 3492 (Table 1).
Diagnosis of Nosema spp. infection and molecular species differentiation
Diagnosis of Nosema spp. infections was performed in accordance with the “Manual of Standards for Diagnostics and Vaccines” published by the Office International des Epizooties (OIE), the World Organization for Animal Health7,48,50,73. In short, per colony 20 pooled bee abdomens were homogenized in 4 ml tap water (200 µl/bee) and microscopically examined for the presence of spores. The moderate sample size of 20 bees also given in the OIE manual is considered sufficient when the study unit is the honey bee colony74,75. This sample size allows to microscopically determine detectable levels of infection above 15% with 96% probability of detection76,77. This level of infection in a colony was considered biologically relevant9. Infection levels were determined by counting the number of Nosema spp. spores per view field (three technical replicates each) in a hemocytometer (Neubauer-improved, VWR, Darmstadt, Germany) using an inverse microscope (VWR, Darmstadt, Germany) with ×100 magnification. Since the view field represents 0.1 µl, the detection threshold is 1 spore in 0.1 µl, hence, 40,000 spores per pooled sample or 2000 spores per bee on average. For classification of the infection levels, standard categories were used73: 0 (no spores), 1 (1–10 spores), 2 (11–100 spores), and 3 (more than 100 spores).
Nosema spp.-positive samples were subjected to further molecular species differentiation either via PCR-amplification of a conserved region of the 16S rRNA gene followed by RFLP (restriction fragment length polymorphism) analysis of this amplicon43,50 or via a species-specific duplex PCR-protocol taking advantage of species-specific sequence differences in the highly conserved gene coding for the DNA-dependent RNA polymerase II largest subunit78. Molecular differentiation enabled the distinction between single infections with either N. apis or N. ceranae, or co-infections where both infections are present at the same time (Table 3).
Statistics
Data were curated, transformed, and presented in spreadsheets for the analysis with the statistic software R79 using the package openxlsx80 (Tables 1, 2, and 3). The statistical analysis of mean percentage of Varroa infestation in survived colonies and collapsed colonies was determined by the one-sample t-test for the entire observation period as well as for each winter season individually (Table 1).
For the dynamics of winter losses, mean mite infestation levels, and N. ceranae infection prevalence, we performed linear regression models (using R base package stats) and calculated its regression line, its slope, the adjusted R2 and the F-statistic (Fig. 1a–c).
To describe and visualize which variable(s) has (have) the largest share in honey bee colony mortality, we took advantage of classification tree analysis (decision trees), a tool of recursive partitioning for multivariate data exploration. We constructed a tree (Fig. 2) with default settings using rpart81 and rattle82. In the construction of the tree, the binary target variable was colony mortality over winter (yes/no) and the tested attributes on each colony were the mite infestation levels and the Nosema infection status, i.e. the infecting Nosema species (n = 3151 non-infected, n = 128 for N. apis, n = 195 for N. ceranae, and n = 18 for mixed infections) and the respective spore load categories (n = 3151 for infection category 0, n = 140 for category 1, n = 141 for category 2, and n = 60 to category 3). In the obtained tree, each node represents an attribute, each branch represents a decision, and each leaf represents the outcome of the decision.
To understand the role of N. ceranae infection in more detail, we looked at the Nosema spp. infection frequencies using contingency tables (Table 2) and performed chi-squared calculation (χ2-calculation). The results were plotted in a dot plot (Fig. 3) and two-way association plots (Fig. 4) created by vcd83,84,85. The obtained results of the significance statistic were further assessed by strength statistic using the effect size index Cohen’s ω69, which is an index for the biological effect size between two categorical variables (Fig. 5a). Cohen’s ω is calculated as follows:
p1i = the proportion in cell i posited by the alternate hypothesis and reflects the effect for that cell; p0i = the proportion in cell i posited by the null hypothesis; m = number of cells [69 p. 216 formula 7.2.1]. For the interpretation of Cohen’s ω in terms of effect size, there is a framework of conventional definition saying that 0.1 ≤ ω < 0.3 is a small, 0.3 ≤ ω < 0.5 is a medium and ω ≥ 0.5 is a large effect size [69 chapter 7.3 p. 227]. To obtain the relation of colony mortalities depending on the two factors Nosema-infection and Varroa-infestation, chi-squared tests were performed and visualized by a bar plot (Fig. 5b).
The map section of Northeast Germany (Fig. 6) showing the boundaries of the Northeastern Federal States of Germany was sourced from the Database of Global Administrative Areas (http://GADM.org)86. Data on the location of the apiaries and the duration of participation were inserted by the use of the following R packages: rnaturalearth87, raster88, ggplot289, sf90, sp91,92, rgeos93, and reshape94.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data generated or analyzed during this study are included in this published article (and its supplementary information files). The numerical data for the analyses and graphs are provided in Supplementary Data 1.
References
Aizen, M. A., Garibaldi, L. A., Cunningham, S. A. & Klein, A. M. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann. Bot. 103, 1579–1588 (2009).
Aizen, M. A., Garibaldi, L. A., Cunningham, S. A. & Klein, A. M. Long-term global trends in crop yield and production reveal no current pollination shortage but increasing pollinator dependency. Curr. Biol. 18, 1572–1575 (2008).
Garibaldi, L. A. et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339, 1608–1611 (2013).
Dainat, B., Evans, J. D., Chen, Y. P., Gauthier, L. & Neumann, P. Predictive markers of honey bee colony collapse. PLoS ONE 7, e32151 (2012).
Dainat, B. & Neumann, P. Clinical signs of deformed wing virus infection are predictive markers for honey bee colony losses. J. Invertebr. Pathol. 112, 278–280 (2013).
van Dooremalen, C. et al. Winter survival of individual honey bees and honey bee colonies depends on level of Varroa destructor infestation. PLoS ONE 7, e36285 (2012).
Genersch, E. et al. The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41, 332–352 (2010).
Fries, I., Martin-Hernandez, R., Meana, A., Garcia-Palencia, P. & Higes, M. Natural infections of Nosema ceranae in European honey bees. J. Apic. Res. 45, 230–233 (2006).
Higes, M. et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 10, 2659–2669 (2008).
Higes, M. et al. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 1, 110–113 (2009).
Botías, C., Martín-Hernández, R., Barrios, L., Meana, A. & Higes, M. Nosema spp. infection and its negative effects on honey bees (Apis mellifera iberiensis) at the colony level. Vet. Res. 44, 25 (2013).
Martin-Hernandez, R. et al. Outcome of colonization of Apis mellifera by Nosema ceranae. Appl. Environ. Microbiol. 73, 6331–6338 (2007).
Cornman, R. S. et al. Pathogen webs in collapsing honey bee colonies. PLoS ONE 7, e43562 (2012).
Tentcheva, D. et al. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 70, 7185–7191 (2004).
Berenyi, O., Bakonyi, T., Derakhshifar, I., Köglberger, H. & Nowotny, N. Occurence of six honeybee viruses in diseased Austrian apiaries. Appl. Environ. Microbiol. 72, 2414–2420 (2006).
Cox-Foster, D. L. et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318, 283–287 (2007).
Traynor, K. S. et al. Varroa destructor: a complex parasite, crippling honey bees worldwide. Trends Parasitol. 36, 592–606 (2020).
Rosenkranz, P., Aumeier, P. & Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 103, S96–S119 (2010).
Ramsey, S. D. et al. Varroa destructor feeds primarily on honey bee fatbody tissue and not hemolymph. Proc. Natl Acad. Sci. USA 116, 1792–1801 (2019).
Zanni, V., Değirmenci, L., Annoscia, D., Scheiner, R. & Nazzi, F. The reduced brood nursing by mite-infested honey bees depends on their accelerated behavioral maturation. J. Insect. Physiol. 109, 47–54 (2018).
De Jong, D., Morse, R. A. & Eickwort, G. C. Mite pests of honey bees. Ann. Rev. Entomol. 27, 229–252 (1982).
Ball, B. V. The association of Varroa jacobsoni with virus diseases of honey bees. Exp. Appl. Acarol. 19, 607–613 (1983).
Ball, B. V. Varroa jacobsoni as a virus vector. In Present status of varroatosis in Europe and progress in the varroa mite control. (ed. Cavalloro R) (1989).
Bailey, L., Carpenter, J. M. & Woods, R. D. Egypt bee virus and Australian isolates of Kashmir bee virus. J. Gen. Virol. 43, 641–647 (1979).
Bowen-Walker, P. L., Martin, S. J. & Gunn, A. The transmission of deformed wing virus between honeybees (Apis mellifera L.) by the ectoparasitic mite Varroa jacobsoni Oud. J. Invertebr. Pathol. 73, 101–106 (1999).
Bailey, L. & Ball, B. V. Honey Bee Pathology. (Academic Press, 1991).
de Miranda, J. R. et al. Cold case: The disappearance of Egypt bee virus, a fourth distinct master strain of deformed wing virus linked to honeybee mortality in 1970’s Egypt. Virol. J. 19, 12 (2022).
Mordecai, G. J., Wilfert, L., Martin, S. J., Jones, I. M. & Schroeder, D. C. Diversity in a honey bee pathogen: first report of a third master variant of the Deformed Wing Virus quasispecies. Int. Soc. Micro. Ecol. 10, 1264–1273 (2016).
Martin, S. J. et al. Global honey bee viral landscape altered by a parasitic mite. Science 336, 1304–1306 (2012).
Ongus, J. R. et al. Complete sequence of a picorna-like virus of the genus Iflavirus replicating in the mite Varroa destructor. J. Gen. Virol. 85, 3747–3755 (2004).
Gisder, S., Möckel, N., Eisenhardt, D. & Genersch, E. In vivo evolution of viral virulence: switching of deformed wing virus between hosts results in virulence changes and sequence shifts. Environ. Microbiol. 20, 4612–4628 (2018).
Gisder, S. & Genersch, E. Direct evidence for infection of Varroa destructor mites with the bee-pathogenic deformed wing virus variant B, but not variant A, via fluorescence in situ hybridization analysis. J. Virol. 95, e01786–01720 (2021).
Posada-Florez, F. et al. Deformed wing virus type A, a major honey bee pathogen, is vectored by the mite Varroa destructor in a nonpropagative manner. Sci. Rep. 9, 12445 (2019).
McMahon, D. P. et al. Elevated virulence of an emerging viral genotype as a driver of honeybee loss. Proc. R. Soc. B 283, 20160811 (2016).
Gisder, S., Aumeier, P. & Genersch, E. Deformed wing virus: replication and viral load in mites (Varroa destructor). J. Gen. Virol. 90, 463–467 (2009).
Yue, C. & Genersch, E. RT-PCR analysis of Deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J. Gen. Virol. 86, 3419–3424 (2005).
Martin, S. J. The role of Varroa and viral pathogens in the collapse of honeybee colonies: a modelling approach. J. Appl. Ecol. 38, 1082–1093 (2001).
Martin, S. J., Hogarth, A., van Breda, J. & Perrett, J. A scientific note on Varroa jacobsoni Oudemans and the collapse of Apis mellifera colonies in the United Kingdom. Apidologie 29, 369–370 (1998).
Keeling, P. J. & Fast, N. M. Microsporidia: Biology and evolution of highly reduced intracellular parasites. Annu. Rev. Microbiol. 56, 93–116 (2002).
Zander, E. Tierische Parasiten als Krankheitserreger bei der Biene. Münchener Bienenztg. 31, 196–204 (1909).
Chemurot, M., De Smet, L., Brunain, M., De Rycke, R. & de Graaf, D. C. Nosema neumanni n. sp. (Microsporidia, Nosematidae), a newmicrosporidian parasite of honeybees, Apis mellifera in Uganda. Eur. J. Protistol. 61, 13–19 (2017).
Fries, I., Feng, F., daSilva, A., Slemenda, S. B. & Pieniazek, N. J. Nosema ceranae n sp (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 32, 356–365 (1996).
Klee, J. et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the Western honey bee, Apis mellifera. J. Invertebr. Pathol. 96, 1–10 (2007).
Paxton, R. J., Klee, J., Korpela, S. & Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 38, 558–565 (2007).
Botías, C. et al. The growing prevalence of Nosema ceranae in honey bees in Spain, an emerging problem for the last decade. Res. Vet. Sci. 93, 150–155 (2012).
Chauzat, M. P. et al. Presence of Nosema ceranae in French honey bee colonies. J. Apic. Res. 46, 127–128 (2007).
Higes, M., Martín-Hernández, R. & Meana, A. Nosema ceranae in Europe: an emergent type C nosemosis. Apidologie 41, 375–392 (2010).
Gisder, S., Schüler, V., Horchler, L. L., Groth, D. & Genersch, E. Long-term temporal trends of Nosema spp. infection prevalence in Northeast Germany: Continuous spread of Nosema ceranae, an emerging pathogen of honey bees (Apis mellifera), but no general replacement of Nosema apis. Front. Cell Infect. Microbiol. 7, 301 (2017).
Fenoy, S., Rueda, C., Higes, M., Martín-Hernandez, R. & del Aguila, C. High-level resistance of Nosema ceranae, a parasite of the honeybee, to temperature and desiccation. Appl. Environ. Microbiol. 75, 6886–6889 (2009).
Gisder, S. et al. Five-year cohort study of Nosema spp. in Germany: does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 76, 3032–3038 (2010).
Martin-Hernandez, R. et al. Effect of temperature on the biotic potential of honeybee microsporidia. Appl. Environ. Microbiol. 75, 2554–2557 (2009).
Horchler, L., Gisder, S., Boecking, O. & Genersch, E. Diagnostic value of faecal spots on and in honey bee (Apis mellifera) hives. Berl. Münch Tier.ärztl Wochenschr. 132, 41–48 (2019).
Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 103, S73–S79 (2010).
Fries, I. Nosema apis - A parasite in the honey bee colony. Bee World 74, 5–19 (1993).
Bailey, L. Nosema apis and dysentery of the honey bee. J. Apic. Res. 6, 121–125 (1967).
Bigliardi, E. & Sacchi, L. Cell biology and invasion of the microsporidia. Microbes Infect. 3, 373–379 (2001).
Franzen, C. How do microsporidia invade cells? Folia Parasitol. 52, 36–40 (2005).
Gisder, S., Möckel, N., Linde, A. & Genersch, E. A cell culture model for Nosema ceranae and Nosema apis allows new insights into the life cycle of these important honey bee pathogenic microsporidia. Environ. Microbiol. 13, 404–413 (2011).
Bailey, L. The infection of the ventriculus of the adult honeybee by Nosema apis (Zander). Parasitology 45, 86–94 (1955).
Guimarães-Cestaro, L. et al. Nosema ceranae (Microsporidia: Nosematidae) does not cause collapse of colonies of Africanized Apis mellifera (Hymenoptera: Apidae) in tropical climate. Sociobiology 67, 408–416 (2020).
Fernández, J. M. et al. Asymptomatic presence of Nosema spp. in Spanish commercial apiaries. J. Invertebr. Pathol. 111, 106–110 (2012).
Stevanovic, J. et al. Dominance of Nosema ceranae in honey bees in the Balkan countries in the absence of symptoms of colony collapse disorder. Apidologie 42, 49–58 (2011).
Guzman-Novoa, E. et al. Nosema ceranae has parasitized Africanized honey bees in Mexico since at least 2004. J. Apic. Res. 50, 167–169 (2011).
Jacques, A. et al. A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLoS ONE 12, e0172591 (2017).
Invernizzi, C. et al. Presence of Nosema ceranae in honeybees (Apis mellifera) in Uruguay. J. Invertebr. Pathol. 101, 150–153 (2009).
Williams, G. R., Shutler, D. & Rogers, R. E. L. Effects at Nearctic north-temperate latitudes of indoor versus outdoor overwintering on the microsporidium Nosema ceranae and Western honey bees (Apis mellifera). J. Invertebr. Pathol. 104, 4–7 (2010).
Higes, M., Garcia-Palencia, P., Martin-Hernandez, R. & Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 94, 211–217 (2007).
Cepero, A. et al. Holistic screening of collapsing honey bee colonies in Spain: a case study. BMC Res. Notes 7, 649 (2014).
Cohen, J. Statistical Power Analysis for the Behavioral Sciences. 2nd edn (Lawrence Erlbaum Associates, Hillsdale, 1988).
Hedtke, K., Jensen, P. M., Jensen, A. B. & Genersch, E. Evidence for emerging parasites and pathogens influencing outbreaks of stress-related diseases like chalkbrood. J. Invertebr. Pathol. 108, 167–173 (2011).
Fries, I. et al. Standard methods for Nosema research. J. Apicult. Res. https://doi.org/10.3896/IBRA.3891.3852.3891.3814 (2013).
Dietemann, V. et al. Standard methods for Varroa research. J. Apic. Res. 52, 28 (2013).
Anonymous. Nosemosis of honey bees (Infection of honey bees with Nosema spp.). OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (OIE World Organisation for Animal Health, 2021).
Doull, K. & Cellier, K. A survey of incidence of Nosema disease (Nosema apis Zander) of the honey bee in South Australia. J. Insect Pathol. 3, 280–288 (1961).
Doull, K. M. The effects of time of day and method of sampling on the determination of Nosema disease in beehives. J. Invertebr. Pathol. 7, 1–4 (1965).
Fries, I., Ekbohm, G. & Villumstad, E. Nosema apis, sampling techniques and honey yield. J. Apic. Res. 23, 102–105 (1984).
Pirk, C. W. W. et al. Statistical guidelines for Apis mellifera research. J. Apic. Res. 52, 1–24 (2013).
Gisder, S. & Genersch, E. Molecular differentiation of Nosema apis and Nosema ceranae based on species–specific sequence differences in a protein coding gene. J. Invertebr. Pathol. 113, 1–6 (2013).
R Development Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org (2021).
Schauberger, P. et al. openxlsx: Read, Write and Edit xlsx Files (Version 4.2.5). https://CRAN.R-project.org/package=openxlsx (2021).
Therneau, T. & Atkinson, B. rpart: Recursive Partitioning and Regression Trees (Version 4.1-15). https://CRAN.R-project.org/package=rpart (2019).
Williams, G. J. Data Mining with Rattle and R: The art of excavating data for knowledge discovery. (Springer, 2011).
Meyer, D., Zeileis, A. & Hornik, K. Residual-based shadings for visualizing (conditional) independence. J. Comput. Graph Stat. 16, 507–525 (2007).
Meyer, D., Zeileis, A. & Hornik, K. vcd: Visualizing Categorical Data (Version 1.4-8). https://CRAN.R-project.org/package=vcd (2020).
Meyer, D., Zeileis, A. & Hornik, K. The strucplot framework: visualizing multi-way contingency tables with vcd. J. Stat. Softw. 17, 1–48 (2006).
Hijmans, R. J. et al. GADM database of Global Administrative Areas https://gadm.org (2016).
South A. rnaturalearth: World Map Data from Natural Earth (Version 0.1.0). https://CRAN.R-project.org/package=rnaturalearth (2017).
Hijmans, R. J. raster: Geographic Data Analysis and Modeling (Version 3.5-15). https://CRAN.R-project.org/package=raster (2022).
Wickham, H. ggplot2: Elegant graphics for data analysis. (Springer, 2016).
Pebesma, E. J. Simple features for R: standardized support for spatial vector data. R. J. 10, 439–446 (2018).
Bivand, R. S., Pebesma, E. J. & Gomez-Rubio, V. Applied spatial data analysis with R. 2nd edn. (Springer, 2013).
Pebesma, E. J. & Bivand, R. S. Classes and methods for spatial data in R. R. N. 5, 9–13 (2005).
Bivand, R. S. & Rundel, C. rgeos: Interface to Geometry Engine - Open Source (“GEOS”) (Version 0.5-9). https://CRAN.R-project.org/package=rgeos (2021).
Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 21, 1–20 (2007).
Acknowledgements
This study was supported by funds of the Ministries responsible for Agriculture of the German Federal States of Brandenburg (MLUK), Saxony-Anhalt (MWL), Saxony (SMEKUL), and Berlin (SENUMVK), by funds of the Federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE), as well as by funds of the German Research Foundation (GRK 2046).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
E.G. conceived and designed the study and the experiments. S.G., V.S., and L.H. carried out the experiments and the microscopic and molecular diagnosis of Nosema spp. S.G., V.S., Y.-C.L., and D.G. performed the statistical analysis. E.G. supervised all work, S.G. supervised the laboratory experiments, and D.G. supervised the statistical analysis. V.S., S.G., D.G., and E.G. interpreted the results and wrote the paper. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schüler, V., Liu, YC., Gisder, S. et al. Significant, but not biologically relevant: Nosema ceranae infections and winter losses of honey bee colonies. Commun Biol 6, 229 (2023). https://doi.org/10.1038/s42003-023-04587-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-023-04587-7
- Springer Nature Limited