
Abstract
Acquired resistance remains a major challenge for therapies targeting oncogene activated pathways. KRAS is the most frequently mutated oncogene in human cancers, yet strategies targeting its downstream signaling kinases have failed to produce durable treatment responses. Here, we developed multiple models of acquired resistance to dual-mechanism ERK/MAPK inhibitors across KRAS-mutant pancreatic, colorectal, and lung cancers, and then probed the long-term events enabling survival against this class of drugs. These studies revealed that resistance emerges secondary to large-scale transcriptional adaptations that are diverse and cell line-specific. Transcriptional reprogramming extends beyond the well-established early response, and instead represents a dynamic, evolved process that is refined to attain a stably resistant phenotype. Mechanistic and translational studies reveal that resistance to dual-mechanism ERK/MAPK inhibition is broadly susceptible to manipulation of the epigenetic machinery, and that Mediator kinase, in particular, can be co-targeted at a bottleneck point to prevent diverse, cell line-specific resistance programs.
Similar content being viewed by others

Introduction
Strategies to durably inhibit the RAF-MEK-ERK mitogen-activated protein kinase (MAPK) signaling network have the potential for broad use in cancers with RAS or RAF family activating mutations, amplifications in downstream kinases, and general dependence on MAPK signaling without genomically conspicuous pathway alterations. While therapies targeting RAF, MEK, and ERK have demonstrated strong preclinical efficacy1,2—and in some cases impressive activity in clinical trials3,4—resistance to first generation ERK/MAPK inhibitors has been problematic, largely secondary to mutational and non-mutational pathway reactivation2,5. Novel classes of dual-mechanism inhibitors have recently been developed to more effectively inhibit ERK/MAPK signaling, predominantly by preventing known adaptations that lead to pathway reactivation, for example by blocking the intrinsic kinase function of the target as well as its ability to be phosphorylated by upstream kinases2,6,7,8,9,10,11. Nevertheless, acquired resistance to dual-mechanism inhibitors has been increasingly reported12,13,14, including studies from our laboratory describing evolved resistance across diverse model systems15. This suggests that signaling events beyond traditional pathway reactivation support the development of resistance to sustained ERK/MAPK suppression.
Recent studies have described dynamic enhancer remodeling in response to kinase inhibition, suggesting that acute chromatin events buttress diverse transcriptional escape programs and that drugs co-targeting the epigenetic machinery can be employed to broadly constrain acquired drug resistance16,17. These findings build off prior work demonstrating the ubiquity of transcriptional changes in drug resistance18,19,20, and the concept of epigenetic remodeling to support adaptive, stable transcriptional programs21,22. However, mechanistic and translational questions remain regarding the combination of targeted kinase inhibitors with epigenetic manipulation. These pertain to (1) the nodes in an oncogenic pathway most susceptible to these combination strategies, (2) the specific chromatin-interacting proteins most targetable in a given genomic and therapeutic context, and (3) the kinetics of acute versus stable epigenetic and transcriptional changes that enable the terminally resistant state.
Here, we developed multiple models of acquired resistance to second generation, dual-mechanism ERK/MAPK inhibitors to characterize transcriptionally-mediated resistance in KRAS-mutant cancers. This oncogenic context was selected given the frequency of KRAS mutations in the most prevalent and lethal solid tumors23,24,25, the refractory nature of KRAS-mutant tumors to single-agent ERK/MAPK inhibition5,26,27,28, and the recent observation that newer ERK/MAPK inhibitors are nonetheless susceptible to evolved resistance in a manner suggestive of transcriptional bypass programs12,13,14,15. Counter to earlier models, we found that transcriptional reprogramming extends beyond the well-established early response, and instead represents a dynamic, evolved process as cell populations refine expression changes to attain a stably resistant phenotype. We delineate the previously hypothesized concept that the intrinsic early transcriptional response can be targeted as a “bottleneck” event and show how this period is broadly vulnerable to pharmacologic perturbation of chromatin-level events in order to block diverse, cell line-specific transcriptional resistance programs. Mechanistic and translational studies reveal that Mediator kinase inhibition antagonizes a conserved early response to ERK/MAPK inhibition, resulting in paralysis of the further transcriptional events necessary for stable resistance. These findings demonstrate that co-targeting of Mediator kinase represents a well-tolerated strategy for preventing resistance to sustained ERK/MAPK inhibition, and furthers our understanding of the kinetics and plasticity underlying drug response.
Results
A revised model of long-term resistance to sustained ERK/MAPK inhibition
We previously assessed long-term ERK/MAPK inhibition across various KRAS-mutant cancers, and consistently observed only transient sensitivity, followed by acquired resistance that develops over several weeks15 (Supplementary Fig. 1a, b). Novel inhibitors of RAF and MEK, despite their proposed mechanisms, nonetheless appear vulnerable to traditional pathway reactivation2,29,30. Alternatively, while dual-mechanism ERK inhibition also activates well-described endogenous feedback events, compensatory activation of MEK and ERK ultimately abates on the timescale at which stable resistance develops (Supplementary Fig. 1c), suggesting that distinct events permit resistance to more sustained ERK/MAPK inhibition.
To confirm these findings in a more translationally relevant system, we utilized a well-credentialed, KrasG12D/Tp53−/− orthotopic, syngeneic mouse model of pancreatic cancer31 to assess resistance to dual-mechanism ERK inhibition (Fig. 1a–c). Tumors demonstrated initial drug sensitivity, yet began to progress within two weeks of treatment initiation (Fig. 1a). Initial drug sensitivity was characterized by ineffective pathway reactivation, evidenced by increased MEK phosphorylation at residues S217/S221 without commensurate recovery of cell proliferation. Conversely, as stable resistance was achieved, phosphorylation of MEK and ERK was relinquished, coupled by cell cycle resumption of tumor proliferation (Fig. 1b, c, Supplementary Fig. 1d). We observed similar findings in additional in vitro models of KRAS-mutant pancreatic and lung cancers (Supplementary Fig. 1e). Notably, these signaling events differ from those induced by first generation, single-mechanism ERK inhibitors, where resistance is characterized by hyperphosphorylation of ERK at residues T202/Y204 (Supplementary Fig. 1f). These findings suggest that while sustained ERK inhibition is susceptible to adaptive resistance, the programs driving ultimate resistance are qualitatively and temporally distinct from traditional pathway reactivation. Moreover, these observations provoke questions regarding the long-term role of MAPK feedback events, which to date have been described on the scale of hours to days2,5.

a FVB/n mice implanted orthotopically with 103 2.1.1syn_Luc cells and treated with either vehicle (20% HpBCD) or SCH772984 (35 mg/kg), with luminescence reported as mean and standard deviation, N = 10 mice per group. Representative micrographs (b) and immunoblots (c) of orthotopic tumors treated with vehicle (20% HpBCD) for 48 h, SCH772984 (35 mg/kg) for 48 h, or SCH772984 (35 mg/kg) for four weeks. The immunoblots are performed with two biologic replicates, and the micrographs are representative of two biologic replicates. d Left, hierarchical clustering of RPPA protein expression changes in MIA PaCa-2 cells treated with SCH772984 (1 µM) relative to DMSO (1:1000), performed in triplicate, with individual proteins annotated by cellular function and clusters indicated by circled numbers. Right, restrictive cubic splines of relative growth rate (total doublings per day) of SCH772984-treated cells compared to DMSO-treated cells (top), as well as selected protein expression changes within MAPK and PI3K/AKT/mTOR signaling pathways (middle) and cell cycle/translation markers (bottom). e Venn diagram depicting total H3K27ac peaks gained or lost (FDR < 0.1) in MIA PaCa-2 cells treated with SCH772984 (1 µM) compared to DMSO (1:1,000) for either one week (early) or eight weeks (stable resistance), performed in duplicate. (f) Scatter plot comparing gene expression changes in MIA PaCa-2 cells treated with SCH772984 (1 µM) compared to DMSO (1:1000) for either one week (early) or eight weeks (stable resistance), performed in triplicate. Each dot represents a single gene, with colored dots representing statistically significant (p < 10−3) gene expression changes at the indicated time points, with statistical significance determined by Wald test using the Benjamini and Hochberg method to correct for multiple hypothesis testing. Error bars represent standard deviations.
To more comprehensively delineate the impact of long-term ERK/MAPK blockade on signaling networks, we applied reverse phase protein array (RPPA)32 to serially profile a KRAS-mutant pancreatic cancer cell line exposed to dual-mechanism ERK inhibition over a time course ranging from one hour to eight weeks (Fig. 1d, Supplementary Table 1). This array probed diverse RTKs and their associated survival pathways, as well as markers of translational control, pro- and anti-apoptotic regulation, cell cycle control, cytoskeletal dynamics, autophagy, transcription factor activation, and histone modifications. Unsupervised clustering revealed three distinct patterns of protein expression characterizing acquired resistance. Cluster 1 was upregulated during an initial period of stunted growth, but then downregulated as stable resistance developed, and was highly enriched for nodes within the MAPK and PI3K/AKT/mTOR signaling pathways. Cluster 2 demonstrated reciprocal downregulation during the initial period of drug sensitivity, followed by a delayed return to baseline expression as resistance developed; this cluster was enriched for markers of cell cycle entry and cap-dependent mRNA translation—which represent an established convergence point of integrated ERK/MAPK and PI3K/AKT/mTOR signaling33—and the asymmetric expression patterns between these clusters suggest that intrinsic feedback events are rendered ineffective in the setting of sustained pathway inhibition. Cluster 3 contained most RTKs and their alternative downstream signaling proteins, which were generally unperturbed by initial drug exposure, and only modestly altered as resistance emerged. Taken together, these findings suggest that pathway reactivation is an endogenous response to ERK/MAPK blockade that may support early survival, but is insufficient to confer stable resistance to sustained inhibition. Furthermore, among diverse alternative signaling pathways, none obviously replaced MAPK signaling to drive growth.
Recent reports have postulated that kinase inhibition induces complex changes to the transcriptional and enhancer landscape permitting a drug-tolerant state, and that early changes (within one week) reflect the necessary adaptations for stable resistance16,17. However, we have consistently observed stunted cell growth beyond this period, and have found that outgrowth is a gradual rather than immediate process during which cell populations display increasing fitness as stable resistance evolves15,34. This suggests that additional transcriptional evolution may be required to permit the terminally resistant state. In fact, our RPPA analysis demonstrated that all histone markers included in our panel underwent dynamic changes throughout the adaptive resistance process (Supplementary Fig. 1g), further supporting longer-term transcriptional reprogramming as a necessary component of the terminally resistant phenotype.
To test this hypothesis, we performed RNA sequencing in parallel with chromatin immunoprecipitation-DNA sequencing (ChIP-seq) for acetylated histone 3, lysine 27 (H3K27ac) in the same treatment-naïve KRAS-mutant pancreatic cancer cell line, as well as at one week of dual-mechanism ERK inhibition and following the development of stable resistance. H3K27ac represents a histone modification that is associated with transcriptional activation and marks active enhancers; thus, changes in H3K27ac density may broadly signify epigenomic remodeling as cells adapt to environmental stress35. To that end, we found that the majority of H3K27ac peaks gained or lost in stable resistance differed from those within the early response, contrasting with previous models suggesting that enhancer remodeling plateaus within 72 h of drug exposure16,17 (Fig. 1e, Supplementary Table S2). And while RNA sequencing revealed a broad early transcriptional response—including well established transcription factors (e.g., EGR1, JUN, FOS), MAPK regulators (e.g., SPRY1/2/4, SPRED1/2), and pro-survival genes in NF-κB/interferon, TGF-β, and alternative tyrosine kinase families16,17 (Supplementary Fig. 1h)—extensive further transcriptional changes were present in stably resistant cells, and the intersection of dysregulated transcripts between these time points was quite limited (Fig. 1f, Supplementary Table 3). This pattern was consistent among even the most highly up- or downregulated transcripts, including a subset of gene expression changes with opposing directionality at the two time points (Supplementary Fig. 1i). Gene set enrichment analysis (GSEA) further confirmed that there were very few enriched pathways shared between the early response and stable resistance (Supplementary Fig. 1j, Supplementary Table 4), and a detailed assessment of RAS/MAPK focused gene sets at these timepoints corresponded with our RPPA analysis suggesting that pathway reactivation was not responsible for resistance (Supplementary Fig. 1j, insert). Taken together, these findings demonstrate that achieving stable resistance requires an adaptive transcriptional process, and that counter to prior models, these gene expression changes extend beyond the intrinsic early response to drug exposure. The fact that H3K27Ac binding also changes substantially between these time points suggests that this transcriptional adaptation may be buttressed by parallel remolding of active enhancers. This model is also consistent with recent work proposing that even treatment-naïve cells upregulating key resistance markers require longer-term transcriptional adaptations to cultivate stable resistance36.
Transcriptionally-mediated resistance programs are broadly vulnerable to manipulation of the epigenetic machinery
To confirm the generalizability of this long-term transcriptional response, we developed and profiled two additional models of evolved resistance to dual-mechanism ERK inhibition in KRAS-mutant lung and colon cancer cells. These cells similarly underwent broad early transcriptional changes followed by extensive further adaptations during the development of stable resistance (Fig. 2a, Supplementary Table 3). Notably, the transcriptional programs giving rise to stable resistance demonstrated limited overlap between cells of different tissue origin (Fig. 2b). Correspondingly, a complete absence of enriched or suppressed GSEA pathways was observed across resistant models (Fig. 2c, Supplementary Table 4). To test whether these differences were due simply to tissue type, we developed stable resistance in an additional KRAS-mutant pancreatic cancer cell line; indeed, the intersection of differentially expressed genes was no greater among the pancreatic cancer lines (Supplementary Fig. 2a, Supplementary Table 3), nor were the associated gene annotations using GSEA (Fig. S2b, Supplementary Table 4). Collectively, these findings demonstrate that diverse, KRAS-mutant cancer cells undergo large-scale transcriptional changes during sustained ERK/MAPK inhibition, yet the transcriptional programs associated with terminal resistance are heterogenous and model-specific.

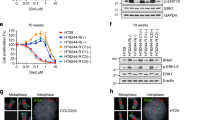
a Scatter plots comparing gene expression changes in SW1573 cells (left) and SW620 cells (right) treated with SCH772984 (1 µM) compared to DMSO (1:1000) for either one week (early) or eight weeks (stable resistance), each performed in triplicate. Each dot represents a single gene, with colored dots representing statistically significant (p < 10−3) gene expression changes at the indicated time points, with statistical significance determined by Wald test using the Benjamini and Hochberg method to correct for multiple hypothesis testing. b Venn diagram depicting differentially expressed genes (p < 10−3) in MIA PaCa-2, SW1573, and SW620 cells treated with SCH772984 (1 µM) relative to DMSO (1:1000) for eight weeks (stable resistance). c Reverse volcano plot depicting gene set enrichment analysis gene sets from the MsigDB Biologic Process Ontology based on the gene expression data from Fig. 2b; Venn diagram depicting enriched gene sets with an FDR < 0.1 (insert). d Schematic representation of the transcriptional modifier pharmacologic screen (left). The heatmap (right) demonstrates the ratio of cell doublings of MIA PaCa-2 cells co-treated with the indicated epigenetic modifier and MAPK inhibitor compared to that MAPK inhibitor alone for three weeks, with each row representing a single biologic replicate. The doubling ratio is plotted, which is calculated as the number of doublings per experimental condition compared to control condition. e Crystal violet staining of 21-day colony growth in MIA PaCa-2 cells treated with the indicated drug combinations, performed in triplicate. f Immunoblot of MIA PaCa-2 cells with indicated genetic modifications (top); the doubling ratio of those same populations treated with Senexin A (1 µM) relative to DMSO (1:1000) for two weeks, either alone (basal growth) or in the presence of SCH772984 (1 µM) (bottom), each condition performed in triplicate. g Eight-point growth inhibition assay of MIA PaCa-2 cells (left), SW620 cells (middle), and SW1573 cells (right) treated with increasing concentrations of SCH772984 in the background of Senexin A (1 µM) or DMSO (1:1000) for four days (top), each condition performed in triplicate; TTP assay of those same cell lines and drug conditions for eight weeks of treatment, each condition performed in triplicate. Error bars represent standard deviations.
Given the obvious challenge of developing strategies to target diverse terminal resistance programs, we instead sought to test whether combining ERK/MAPK inhibition with drugs targeting the epigenetic and transcriptional machinery might broadly perturb heterogeneous responses to drug exposure. To accomplish this, we performed long-term pharmacologic screens assessing a panel of drugs targeting diverse transcriptional processes on the development of resistance to second-generation RAF, MEK, and ERK inhibitors.
In total, we screened 42 combination therapies (Fig. 2d, Supplementary Table 5), revealing, most strikingly, that resistance to sustained ERK inhibition was broadly susceptible to manipulation of the transcriptional machinery, including drugs co-targeting CREBBP/EP300, EZH1/EZH2, BET family bromodomain proteins, and CDK8/19. In keeping with our previous findings of pathway reactivation driving resistance to RAF and MEK inhibition, these targets were generally less vulnerable to combination therapy. We found that only the BET bromodomain inhibitor JQ-1 and the CDK8/19 inhibitor Senexin A delayed resistance to inhibition at all three ERK/MAPK nodes, with the strongest effect in combination with ERK inhibition. The effect of BET bromodomain inhibition on drug response has been broadly reported34,37,38,39. CDK8 and CDK19 are paralogous proteins that reversibly associate with the multiprotein Mediator complex (Mediator kinase)40, and have never, to our knowledge, been implicated in resistance to ERK/MAPK inhibition. CDK8/19 inhibition alone had no effect on basal growth (Supplementary Fig. 2c), nor did it demonstrate short-term synergy with ERK/MAPK inhibition (Supplementary Fig. 2d), suggesting that the impedance of resistance occurred through later-stage inhibition of the adaptive process. Given the profound effect of Mediator kinase inhibition on the activity of drugs targeting all three ERK/MAPK nodes, we chose to characterize this interaction across the KRAS-mutant tissues types for which we had developed models of transcriptionally-mediated resistance.
Mediator kinase inhibition impedes long-term acquired resistance to ERK/MAPK-targeted therapy
We first validated the interaction of combined ERK/MAPK and Mediator kinase inhibition using three structurally distinct preclinical compounds (Senexin A, Cortistatin A, and CCT251545) in long-term colony forming assays at doses shown to selectively inhibit CDK8/1941,42,43. All three compounds profoundly inhibited clonal outgrowth during sustained treatment with RAF, MEK, and ERK inhibition, and had minimal effect on basal cell growth (Fig. 2e). Most notably, combined ERK and CDK8/19 inhibition completely prevented the emergence of resistant colonies at four weeks, supporting a dominant role for Mediator kinases in transcriptional reprogramming during sustained ERK inhibition.
As our work to this point relied exclusively on pharmacologic inhibition of CDK8/19, we next sought to probe the mechanistic role of each Mediator kinase. CDK8 and CDK19 possess enzymatic activity, but the proteins also serve scaffold functions44, and thus protein depletion and kinase inhibition have distinct cellular effects45,46. To test whether pharmacologic inhibition was in fact specific to Mediator kinase function, we utilized dual CRISPR/Cas9 constructs47 to create knockout derivatives of both CDK8 and CDK19, CDK8 only, CDK19 only, or double-sham control knockouts in three KRAS-mutant cancer cell lines. Each condition was then subjected to treatment with DMSO, ERK inhibition, CDK8/19 inhibition, or combined ERK and CDK8/19 inhibition (Fig. 2f, Supplementary Fig. 2e, f). As expected, in basal growth conditions treatment with CDK8/19 inhibition had a limited effect in all four derivatives. In the presence of ERK inhibition, however, CDK8/19 inhibition profoundly suppressed outgrowth of control cells expressing both CDK8 and CDK19, modestly blocked growth in cells with individual knockout of CDK8 or CDK19, yet had no effect in cells depleted of both CDK8 and CDK19 (Fig. 2f). This confirmed that CDK8 and CDK19 were indeed the targets of kinase inhibition responsible for the long-term impedance of resistance.
Finally, to evaluate the effect of CDK8/19 inhibition on sustained MAPK suppression within a more clinically meaningful timescale, we utilized an established time-to-progression model15,34,48 to test growth up to eight weeks. Despite the diverse transcriptional resistance programs observed across tissue types, CDK8/19 inhibition completely prevented lung and colon cancer cells from developing resistance to ERK inhibition (Fig. 2g, bottom), and markedly delayed the emergence of resistance in pancreatic cancer cells. Of note, combined ERK and CDK8/19 inhibition demonstrated no short-term synergy in any cell line (Fig. 2g, top), and CDK8/19 inhibition alone had a negligible effect on long-term basal growth (Supplementary Fig. 2g), positioning this combination as a promising strategy for preventing long-term acquired resistance in KRAS-mutant cancers.
Antagonization of a conserved response network paralyzes further adaptive potential
Given that KRAS-mutant cancer cells of distinct tissue origin demonstrated distinct stable resistance programs to ERK inhibition, yet that these programs were universally susceptible to CDK8/19 co-inhibition, we next asked whether there might be a common early response to ERK inhibition vulnerable to combination treatment. In fact, we found that in stark contrast to the limited intersection of expression changes seen in stable resistance (Fig. 2b), considerable overlap could be appreciated between cell lines of distinct tissue origin following one week of ERK inhibition. This suggested that ERK inhibition induces a conserved early transcriptional response before cells undergo further heterogeneous adaptations that ultimately establish stable resistance programs. This was further revealed by GSEA (Fig. 3b), which demonstrated substantial overlap of enriched gene annotations between tissue types within the early response, again contrasting with the lack of shared pathways in stable resistance (Fig. 2c). Broadly, GSEA revealed that this initial response resulted in downregulation of major anabolic processes such as DNA replication, cell cycle entry, and protein translation (Supplementary Table 4), which may be necessary for cells to forego growth and replication as unique transcriptional programs are enacted.

a Venn diagram depicting differentially expressed genes (p < 10−3) in MIA PaCa-2, SW1573, and SW620 cells treated with SCH772984 (1 µM) relative to DMSO (1:1000) for one week, performed in triplicate, with statistical significance determined by Wald test using the Benjamini and Hochberg method to correct for multiple hypothesis testing. b Reverse volcano plot depicting gene set enrichment analysis using gene sets from the MsigDB Biologic Process Ontology based on the gene expression data from Fig. 3a; Venn diagram depicting enriched gene sets with an FDR < 0.1 (insert). c In MIA PaCa-2, SW1573, and SW620 cells, odds that genes significantly dysregulated by treatment with SCH772984 for one week (1 µM, p < 0.001 by Wald statistic) are further up- or downregulated by co-treatment with Senexin A (1 µM), relative to genes that are not significantly dysregulated by treatment with ERK inhibition; p-value calculated according to Sheskin method. d In these same cell lines, percent of transcripts significantly dysregulated by SCH772984 (1 µM) for which treatment with Senexin A (1 µM) antagonizes or cooperates with these gene expression changes. e Heatmap (top) depicts differential expression of all genes within the assigned 67 DPGP clusters for MIA PaCa-2 cells treated with SCH772984 (1 µM) relative to DMSO (1:1000) at the indicated time points; restricted cubic splines of the mean differential expression of each cluster at each time point (bottom). f Restricted cubic spline of all genes in representative clusters 1 and 3 treated with SCH772984 (1 µM) alone or in combination with Senexin A (1 µM), with the bolded curves representing mean expression changes of each cluster according to treatment condition; the curves on the top reflect equivalent time points for the two treatment conditions, while on the bottom the SCH772984 alone curve is right-shifted by the indicated time intervals. g Bubble charts reflecting the mean expression changes of each cluster according to treatment condition at one week (left) versus three weeks (right), with the size of each bubble reflecting the number of genes in each cluster. h Bubble charts reflecting the mean expression changes of each cluster for MIA PaCa-2 cells treated with SCH772984 (1 µM) for three weeks compared to SCH772984 (1 µM) and Senexin A (1 µM) for five weeks, with the size of each bubble reflecting the number of genes in each cluster. ****p < 0.0001. Error bars represent standard deviations.
We next asked whether this conserved response to ERK inhibition was particularly vulnerable to combined CDK8/19 inhibition. Specifically, we tested whether CDK8/19 co-inhibition preferentially dysregulated the genes identified within this early conserved response network, or whether CDK8/19 inhibition agnostically caused gene expression changes throughout the transcriptome. We found that genes dysregulated by ERK inhibition were significantly more likely to be further altered by combined CDK8/19 inhibition (Fig. 3c). We next asked whether this interaction occurred with specific directionality. A nonspecific effect should cause limited expression changes with near-random directionality; alternatively, cooperation or antagonism of a transcriptional program should preferentially amplify or dampen expression changes. Strikingly, we found that CDK8/19 co-inhibition broadly antagonized the expression changes within the early conserved response (Fig. 3d, Supplementary Fig. 3a). At this early time point, cells treated with ERK inhibition alone or in combination with CDK8/19 inhibition were phenotypically similar, with equivalent growth inhibition in each treatment condition (Supplementary Fig. 3b). This led us to ask whether the ultimate consequence of Mediator kinase co-inhibition was downstream impairment of further transcriptional adaptations necessary for stable resistance.
To do this, cells treated with ERK inhibition alone or combined with CDK8/19 inhibition were serially profiled by RNA sequencing over the five-week time course during which resistance developed (Supplementary Table 6). Notably, we found that individual transcripts underwent dynamic alterations following the early conserved response, including thousands of genes found to be either significantly upregulated and downregulated at different time points throughout this evolved process (Supplementary Fig. 3c). Notably, transcriptome-wide differences between treatment conditions became increasingly pronounced over time (Supplementary Fig. 3d), further suggesting that the phenotype induced by CDK8/19 co-inhibition was caused by disruption of this downstream evolutionary process.
In order to visualize, quantify, and evaluate patterns of whole-transcriptome evolution over time, we utilized an established Dirichlet process Gaussian process (DPGP) mixture model, which facilitates time series cluster measurement of genomic features49. Applying this model to the genes with the greatest variance over the course of acquired resistance, we identified 67 gene sets reflecting diverse expression trajectories (Fig. 3e; Supplementary Table 7). Like the gene-level analysis between treatment conditions, cluster trajectories between treatment conditions demonstrated overall divergence after the early conserved response (Supplementary Fig. 3e). By quantifying and comparing all cluster-level interval changes, we found that Mediator kinase inhibition exerted its dominant effect following this early response, paralyzing transcriptional reprogramming between weeks one and three of treatment, at which point dually-treated cells resumed a trajectory that mirrored cells treated with ERK inhibition alone (Fig. 3f–h). By visualizing individual clusters and then shifting the timescale of dually-treated cells to “remove” this period of transcriptional stagnation, the trajectory curves superimposed upon one another (Fig. 3f), and we ensured that all cluster trajectories could be realigned with this time shift (Fig. 3g, h). We further extrapolated these findings to the entire transcriptome at both the gene-level and using GSEA (Supplementary Fig. 3f–h). Taken together, these findings indicate that the emergence of resistance coincides with transcriptional escape from the early conserved response, a concept that can be observed at the transcript level, using functional annotated gene sets, and using a DPGP mixture model. Co-targeting Mediator kinase therefore antagonizes the conserved response to ERK inhibition, and then exerts its phenotypic effect by preventing further transcriptional changes necessary to establish stable resistance.
Transcriptionally-mediated resistance is driven by distinct terminal mechanisms in different models
Given the magnitude and heterogeneity of gene expression changes observed in each resistant model (Fig. 3a, b), yet the ability of CDK8/19 inhibition to broadly curtail the emergence of resistance (Fig. 2g), we sought to map out a specific resistance mechanism in one model. To date, efforts to characterize the function of individual components of a transcriptional program have been limited by the scalability of candidate-based approaches. We considered it unrealistic to implicate the functional importance of individual genes based simply on expression changes, as some highly dysregulated transcripts were likely secondarily regulated passengers of alterations to the chromatin architecture, vestigially over- or underexpressed following the early response, or simply dysregulated at random.
In order to test the consequence of gene-level transcriptional events, we designed a specialized loss-of-function library of CRISPR/Cas9 constructs targeting the subset of genes most dysregulated throughout the adaptive process in MIA PaCa-2 cells (Fig. 4a, Supplementary Table 8). We then subjected treatment-naïve and evolved resistant cells transduced with this library to five weeks of either ERK inhibition or control conditions to test the effect of gene-loss at various time points. We validated our approach by comparing final and initial sgRNA pools from treatment-naïve parental cells across replicates, focusing on known essential genes, non-essential genes, and non-targeting controls (Fig. 4b). These results revealed that while extensive transcriptional changes were observed as a stably resistant phenotype was established, the vast majority of these genes were not independently necessary for the maintenance of the resistant state, as their knockout failed to re-sensitize resistant cells to ERK inhibition (Fig. 4c, Supplementary Fig. 4a).

a Schematic depicting CRISPR/Cas9 loss-of-function screen. b Replicate-to-replicate comparison of gene-level essentiality phenotypes in MIA PaCa-2 cells treated with DMSO (1:1000) alone. Essential controls are shown in red, non-essential controls in blue, and non-targeting controls in yellow. c Scatter plot of genes included in loss-of-function screen comparing gene expression changes in MIA PaCa-2 cells treated with SCH772984 (1 µM) compared to DMSO (1:1000) for either one week (early) or eight weeks (stable resistance), with loss-of-function gene score indicated by color gradient. Gene score is calculated as the log2 fold change of the fractional representations (time final/time zero) of resistant cells treated with SCH772984 (1 µM) compared to parental cells treated with DMSO (1:1000) at the screen midpoint (19 days). d Gene-level representation of essential phenotypes in evolved resistant MIA PaCa-2 cells treated with SCH772984 (1 µM) at the screen midpoint (19 days), ranked by their mean log2-transformed gene score across duplicates; the insert shows the effect of ABCG2 loss in parental cells treated with DMSO (1:1000, left), resistant cells treated with SCH772984 (1 µM, middle), and resistant cells treated with DMSO (1:1000, right) at all three screen time points (15, 19, and 23 days). e Immunoblot of treatment-naïve parental MIA PaCa-2 cells and three independently evolved resistant derivatives of this same cell line to SCH772984 (1 µM). f Crystal violet staining of MIA Paca-2 cells evolved resistant to SCH772984 (1 µM) with either no alteration, control knockout, or ABCG2 knockout, treated with either SCH772984 (1 µM) or DMSO (1:1000) for one week; representative photograph of all conditions performed in triplicate. g Eight-point growth inhibition assay of parental and evolved resistant MIA PaCA-2 cells treated with increasing concentrations of SCH772984, performed in triplicate; one triplicate set of evolved resistant cells is treated with the ABCG2 inhibitor Elacridar (1 µM) in the background. h GI50 values derived from eight-point dose-response curves of parental and evolved resistant MIA PaCA-2 cells treated with increasing concentrations of the indicated chemotherapies, with and without Elacridar (1 µM). i Genome browser view of ChIP-seq tracks across the ABCG2 gene coding locus depicting H3K27Ac density in MIA PaCa-2 cell treated with DMSO (1:1000, parental), SCH772984 (1 µM) for one week (Early), or SCH772984 (1 µM) for eight weeks (Resistant); known enhancer elements within the ABCG2 coding region are depicted with blue boxes. Error bars represent standard deviations.
Among the subset of genes demonstrating functional importance in the terminally resistant state, the knockout of one gene upregulated in resistant cells —encoding the ATP-binding cassette protein ABCG2 that functions as notorious multidrug transporter with broad polysubstrate specificity50—dramatically sensitized resistant cells to ERK inhibition (Fig. 4c, d). ABCG2-mediated resistance was extensively validated by demonstrating its reproducible upregulation, and subsequently that both gene knockout and pharmacologic inhibition of its ATPase function fully reversed ERK inhibitor resistance, while sensitizing cells to treatment with other known ABCG2 substrates (Fig. 4e–h). ABCG2 loss demonstrated no functional consequence in basal growth nor in resistant cells when drug was removed (Fig. 4d (insert), Supplementary Fig. 4a, b), confirming it as a bona fide driver of drug resistance in this model. To ensure that ABCG2-upregulation was not simply occurring via the outgrowth of small population of ABCG2 high-expressing “persister” cells, we demonstrated that the vast majority of individual cells were capable of developing resistance across a consistent timescale (Supplementary Fig. 4c). The reproducible resistance mechanism observed in MIA PaCa-2 cells (Fig. 4e) was distinct from KRAS-mutant colon and lung cancer models, which evidenced neither ABCG2 upregulation (Supplementary Fig. 4d) nor resensitization to ERK inhibition by pharmacologic ABCG2 inhibition (Supplementary Fig. 4e). Importantly, a region within the gene locus of ABCG2 represented one of the most significantly gained H3K27ac peaks in terminally resistant MIA PaCa-2 cells compared to the treatment-naïve state (ranked #16 of >32,000 peaks by adjusted p-value, and #1 by magnitude; Supplementary Fig. 4f). This region was not significantly gained in the early response (adjusted p value 0.658), and contained the best-known enhancer of ABCG2, approximately 73 kb downstream of its transcriptional start site (Fig. 4i); additionally, two other peaks within the ABCG2 locus were also significantly gained relative to the control or one week-treated samples, both containing other putative ABCG2 promoters/enhancers (Fig. 4i, Supplementary Fig. 4f, g). This further supports the concept that long-term enhancer remodeling and parallel transcriptional adaptations are necessary for stable resistance to second-generation ERK/MAPK inhibition. While ABCG2 upregulation reproducibly drives acquired resistance to ERK inhibition in MIA PaCa-2 cells, differing mechanisms drive resistance in other cell line models, highlighting the notion that strategies targeting transcriptional adaptation are likely to be more broadly effective at blocking resistance than strategies targeting discrete gene expression changes.
Co-targeting Mediator kinases delays resistance to ERK inhibition across translational models of KRAS-mutant cancers
To assess the translational generalizability of combined ERK and Mediator kinase inhibition, we evaluated this strategy across multiple in vitro and in vivo models of KRAS-mutant cancer, including additional eight-week time-to-progression models, patient-derived rectal cancer tumoroids, and a pancreatic cancer mouse model.
We first expanded upon the existing time-to-progression model by testing combined ERK and CDK8/19 inhibition across diverse KRAS-mutant pancreatic, lung, and colorectal cancer cell lines. In all models, cotreatment with CDK8/19 inhibition delayed the emergence of resistance, in many cases completely preventing resistance until the assay was terminated at eight weeks (Fig. 5a, Supplementary Fig. 5a). Again, combined ERK and CDK8/19 inhibition demonstrated no short-term synergy in any cell line (Supplementary Fig. 5b), and CDK8/19 inhibition alone had a negligible effect on long-term basal growth (Supplementary Fig. 5c). We also tested two lines with distinct drug sensitivities based on alternative genomic contexts (one BRAF mutant melanoma (A375) sensitive to RAF inhibition and one KRAS wild type colorectal cancer (LIM1215) sensitive to EGFR inhibition). In these two models, CDK8/19 inhibition had no effect on the emergence of resistance (Fig. 5a, Supplementary Fig. 5a). This suggests that either resistance in these model systems is not driven by transcriptional reprogramming, or that the transcriptional reprogramming occurring in these settings is not CDK8/19-dependent. Either way, it underscores that across KRAS-mutant model systems of pancreatic, lung, and colorectal cancer, resistance to dual-mechanism ERK/MAPK inhibition occurs secondary to large-scale transcriptional adaptations which rely on Mediator kinase. Whether CDK8/19 inhibition serves as an effective combination strategy in other genomic contexts where resistance develops via large-scale, long-term transcriptional adaptations requires further investigation.

a Bar chart depicting the time at which resistance emerged to treatment with either SCH772984 (1 µM) alone or SCH772984 (1 µM) in combination with Senexin A (1 µM). b Oncoplot of three rectal cancer tumoroids based on MSK-IMPACT testing (top), with representative micrographs below of RC-MSK-001 patient tumor (left) and its corresponding tumoroid (right). c TTP assays for rectal cancer tumoroids treated with SCH772984 (100 nM) alone or in combination with Senexin A (1 µM). d Relative viability of rectal cancer tumoroids treated with Senexin A (1 µM) compared to DMSO (1:1000) for 14 days. e Crystal violet staining of 14 day colony growth of 2.1.1syn_Luc cells treated with the indicated drug combinations. Box plot demonstrating whole body luminescence (f) and tumor weights (g) after orthotopic implantation of 103 2.1.1syn_Luc cells into FVB/n mice and treatment with SCH772984 (35 mg/kg) in combination with or without CCT251545 (75 mg/kg) for 21 days, 10 mice per group. g Mean mouse weights throughout treatment described in Fig. 5f, g. Error bars represent standard deviations.
Next, we tested the combination of ERK and CDK8/19 inhibition in three KRAS-mutant, patient-derived tumoroid models of rectal cancer (Fig. 5b–d). These tumoroids had previously been shown to retain the molecular, histological, and clinical features of the patient tumors from which they were derived, as well as accurately predict patient responses to chemotherapy and radiation therapy51. Like the cell line time-to-progression models, tumoroids developed resistance to ERK inhibition within five weeks of treatment (Fig. 5c). Alternatively, tumoroids treated with combined ERK and CDK8/19 inhibition demonstrated complete growth suppression up to eight weeks (Supplementary Fig. 5d). Again, inhibition of Mediator kinase alone had no effect on tumoroid growth (Fig. 5d).
Finally, we tested the combination of ERK and CDK8/19 inhibition in a well-credentialed, orthotopic, syngeneic mouse model of pancreatic cancer. We first tested three independent CDK8/19 inhibitors in vitro using a cell line derived from the same model. Cell growth was unaffected by each inhibitor alone, yet resistance to ERK inhibition was markedly impeded by co-inhibition of Mediator kinase, with similar efficacy between compounds (Fig. 5e). Of these three inhibitors, we selected CCT251545, a selective inhibitor of CDK8/1942,52 for in vivo studies (Cortistatin A is not yet commercially available, and Senexin A has known limited oral bioavailability). Treatment with ERK inhibition alone dramatically reduced initial tumor growth, yet resistance to treatment developed within two weeks of drug exposure (as in Fig. 1a). In contrast, inhibition of ERK and CDK8/19 delayed the acquisition of resistance (Fig. 5f), resulting in a 34% reduction in tumor weight at study endpoint (Fig. 5g) without additional toxicity (Fig. 5h). As in the cell line and tumoroid models, inhibition of Mediator kinase alone had no effect on in vivo tumor growth or overall toxicity (Supplementary Fig. 5e–g), positioning CDK8/19 co-inhibition as an effective and well-tolerated strategy for preventing resistance to sustained MAPK inhibition.
Discussion
The RAS family of oncoproteins represents the most frequently mutated genes in human cancers53,54, and KRAS, in particular, is altered in a subset of the most common and deadly solid tumors23,24,25. While therapies targeting the ERK/MAPK pathway represent an attractive strategy for these malignancies, early inhibitors were limited by complex negative feedback loops2,5, and newer agents designed to prevent pathway reactivation have also proven susceptible to evolved resistance12,13,14,15. Given these obstacles, the majority of KRAS-mutant cancers continue to be treated with conventional cytotoxic chemotherapies55,56,57,58.
Here, we demonstrate that sustained ERK/MAPK inhibition causes transcriptional reprograming that is broadly vulnerable to pharmacologic perturbation of chromatin binding and modifying factors, including Mediator kinase. Co-targeting CDK8/19 antagonizes a conserved early response to ERK inhibition, which then paralyzes additional transcriptional adaptations necessary to establish stable resistance. Across translational models, inhibition of Mediator kinase alone demonstrated only minimal effect on basal growth, and long-term inhibition of CDK8/19 was well-tolerated while preventing acquired resistance to ERK inhibition. Thus, this work validates a strategy for robust and potentially nontoxic prevention of ERK/MAPK inhibitor resistance, a problem that to this point has proven recalcitrant.
This study builds on prior work demonstrating that repression of early inhibitor-stimulated transcriptional events can broadly improve therapeutic efficacy without a priori knowledge of all potential terminal resistance mechanisms16,17. This strategy differs from “rational” combination approaches that pair oncogene-targeted therapies with drugs inhibiting known resistance mechanisms, which have provided only transient effectiveness in both preclinical studies and patient trials26,27,28. Notably, our own work using CRISPR/Cas9-based screening to identify co-targets for inhibition with second-generation ERK/MAPK inhibitors yielded potent short-term combinations that nonetheless gave rise to resistance; ultimately, three-drug combinations leveraging either the primed apoptotic state or the addition of conventional cytotoxic chemotherapies were required to achieve durable responses in vivo15.
Here, we coopt a strategy to broadly impede a conserved gene expression network in order to block further transcriptional adaptations and circumvent an array of potential resistance mechanisms. The distinct transcriptional programs identified in this study highlight the diversity of resistance mechanisms that may be simultaneously targeted by this approach. Interestingly, prior work in this space has suggested that a transient epigenetic state22 or rare cell transcriptional variability36 leads to resistance in only a small subpopulation of cells. In contrast, findings from this study suggest that a large fraction of the tumor cell population may be capable of transcriptional reprogramming in order to support stable resistance programs, although future work is necessary to elucidate the population dynamics and kinetics of these cell-level events.
Notably, in assessing resistance to multiple levels of MAPK inhibition, we found that inhibition of RAF and MEK were less vulnerable to strategies co-targeting the epigenetic machinery than ERK inhibition. This may be secondary to distinct patterns of resistance; signaling analyses suggest that while even newer inhibitors of RAF and MEK are susceptible to pathway reactivation, second-generation ERK inhibition more sustainably blocks MAPK signaling at its terminal node. Moreover, among the primary ERK/MAPK pathway proteins, ERK uniquely acts on both cytoplasmic and nuclear targets59. Together, this may result in transcriptional reprogramming as the dominant mechanism of resistance for persistent ERK/MAPK blockade, and thus strategies that circumvent broad patterns of resistance may prove highly useful for further clinical development of ERK inhibitors.
While we found that Mediator kinase co-inhibition robustly suppressed resistance to ERK inhibition in our cell line and organoid models, we observed a shorter period of delayed resistance in vivo, which represents one of several limitations to this study that should be acknowledged as we consider the translational application of these findings. It is possible that this reflects the highly aggressive nature of the orthotopic model that we used, and in less aggressive KRAS-mutant models resistance may be delayed in a manner that would be suitable for transitioning to human trials. However, a second hypothesis is that the prevention of resistance in vivo was limited by the oral bioavailability and metabolic stability of CCT251545, which has been broadly problematic for CDK8/19 inhibitors to date. This is supported by the strong effect we observed in vitro using cells derived from the same model with three separate CDK8/19 inhibitors. Currently, the natural product Cortistatin A is considered the most promising preclinical compound in terms of its potency and bioavailability, but it is not yet commercially available, in part due to a complicated synthesis process41,45. Thus, while there is a broad interest in the clinical development of additional potent, bioavailable CDK8/19 inhibitors, significant translational validation in vivo will be needed before this strategy has the potential to move into the clinical space. Furthermore, while we demonstrate that co-inhibition of Mediator kinase impedes that early transcriptional response that ultimately permits adaptive resistance, mechanistic follow-up questions remain regarding the chromatin-level events that determine this response. Future work should specifically address which transcription factors and other DNA-binding proteins causally link the Mediator complex to the transcriptional response to ERK inhibition. Finally, it is worth noting that genetic knockout of CDK8/19 does not phenocopy inhibition of their kinase function in the context of resistance to ERK inhibition. This is a well-known feature of the Mediator kinases, which function as components of larger complexes, and loss of the entire protein can cause destabilization or changes to the complex’s constituents in ways that are produce different effects than kinase inhibition60. Similar phenomena have been observed for other transcriptional CDKs, including by our group61, and while these are noteworthy, they do not diminish the potential pharmacologic opportunities of using CDK inhibitors as combinations therapies.
In sum, this study supports the therapeutic strategy of combining ERK and Mediator kinase inhibition. Our results, which combined diverse cell lines with tumoroids and in vivo models of KRAS-mutant cancers, suggest that Mediator kinase inhibitors may be broadly effective in blocking short- and long-term transcriptional changes required for the emergence of drug resistant cell populations. Additional studies testing novel classes of ERK/MAPK pathway inhibitors in combination with drugs that target epigenetic and transcriptional processes may provide an efficient way of overcoming the rapid resistance that has traditionally been observed with genetically-targeted anticancer therapies.
Methods
Cell lines and reagents
All cell lines were grown at 37 °C in 5% CO2. Human pancreatic, lung, and colorectal cancer cell lines were grown in DMEM/F12, 10% FBS, and 1% penicillin/streptomycin. Mouse cell lines were grown in DMEM, 10% FBS, and 1% penicillin/streptomycin. All cell lines were purchased from American Type Culture Collection (ATCC) or Duke University Cell Culture Facility (CCF). All cell lines were authenticated using Promega PowerPlex 18D kit or were purchased within 6 months from Duke CCF. Drugs were purchased from Selleck Chemicals, except Cortistatin A (a gift from the laboratory of Matthew Shair) and CCT251545 (which was provided by The Institute of Cancer Research).
Mouse studies
As previously described62, female FVB/n mice were injected orthotopically (into the head of the pancreas) with 1000 luciferase-expressing p53 2.1.1syn_Luc mouse pancreatic tumor-derived cells (KrasG12D/Tp53−/−; provided by Dr. Eric Collisson, UCSF) that had been resuspended in 50% Hanks’ balanced salt solution (Gibco), 50% LDEV-free Matrigel (Corning) at a concentration of 25 cells/mL. Ketamine (80 mg/kg), xylazine (8 mg/kg), and acepromazine (1 mg/kg) were used to anesthetize the mice prior to surgery. A tuberculin syringe with a 30-g needle was inserted into an abdominal incision and used for implantation. The incision was closed using surgical staples. After a one-week recovery, initial IVIS imaging with D-luciferin substrate (Perkin Elmer) was performed using an IVIS Lumina optical imaging system. For each independent experiment, mice were separated into groups of 10, and treated daily with either vehicle (20% HdpCD, IP), SCH772985 (35 mg/kg, IP), CCT251545 (75 mg/kg, OG), or a combination of both SCH772985 (35 mg/kg, IP) and CCT251545 (75 mg/kg, OG). CCT251545 was resuspended in ORA-Plus (Perrigo). IVIS imaging was performed at weekly time points. Mice were sacrificed at the conclusion of the study, with the endpoints defined as 28 days of drug or vehicle treatment, progressively growing tumor nodule >2 cm, or signs of poor health (e.g., weight loss, decreased grooming, or inability to feed). At study endpoint, the pancreas was removed, and tumors were then excised from the pancreas and weighed. Euthanasia was performed by CO2 treatment followed by cervical dislocation. All animal studies were conducted in accordance with the UNC Institutional Animal Care and Use Committee (IACUC).
Short-term growth inhibition assay
As previously described15, cells were seeded into 96-well plates at 5000 cells/well. To generate GI50 curves, cells were treated with vehicle using an eight-log serial dilution of drug. For certain experiments, cells were treated in the background of a constant concentration of a second drug or DMSO (1:1000) to test combinatorial effects. Each treatment condition was represented by at least three replicates. Three days after drug addition, cell viability was measured using Cell Titer Glo (Promega). Relative viability was then calculated by normalizing luminescence values for each treatment condition to control treated wells. Dose-response curves were fit using GraphPad/Prism 6 software.
Long-term time-to-progression (TTP) assay
As previous described34, cells were seeded at 250,000 per plate in 10 cm plates in triplicate. The next day, drugs were added at indicated concentrations. At weekly time points, cells were counted and replated at 50,000–100,000 per plate, and drug/media was replenished. Virtual cell counts were calculated based upon the number of cells plated, the growth rate, and the final cell counts at each week. For most experiments, the TTP assay was terminated at eight weeks. For the double CRISPR/Cas9 knockout of CDK8 and CDK19, the TTP assay was terminated at two weeks. Progression was established when the growth rate exceeded 50% of that of its parental, treatment-naïve derivative.
Western blotting and antibodies
Immunoblotting was performed as previously described15. Membranes were probed with primary antibodies (1:1000 dilution) recognizing KRAS (CST #14429), p-MEK (CST #9121), T-MEK (CST #4694), p-ERK (CST #9101), T-ERK (CST #4695), vinculin (CST #4650), CDK8 (CST #4106), CDK19 (Sigma HPA007053), and ABCG2 (CST #4477). Blots within each subfigure were derived from the same experiment and were processed in parallel.
Immunohistochemistry
Formalin-fixed, paraffin-embedded tissue blocks of mouse orthotopic tumors were sectioned and stained with hematoxylin and eosin, as well as antibodies targeting phospho-MEK (CST #2338), phospho-ERK (CST #4370), and Ki-67 (Sigma SAB5600249). Negative controls were performed on all runs using an equivalent concentration of a subclass-matched immunoglobulin. All immunohistochemistry was performed in biologic duplicate. Images were then digitized and visualized using Aperio Imagescope.
Reverse-phase protein array sample preparation and analysis
MIA PaCa-2 cells were treated with SCH772084 (1 µM) or DMSO (1:1000) for the indicated durations at which point plates were washed twice in ice-cold PBS and then frozen at −80 °C. At the completion of all time points, plates were scraped and RPPA analysis was performed as previously described32. All samples were conducted in biological triplicate and normalized to a DMSO control at each respective time point.
Generation of evolved resistant cell lines
Cells were plated at a density of 250,000 cells in 10 cm plates. The following day, SCH772984 (1 µM) was added. Cells were then split at weekly time points and replated at a density of 50,000, and drug/media was replenished. Lines were considered resistant when the growth rate exceeded 50% of that of its parental, treatment-naïve derivative, and terminal resistance was defined once there was no interval increase in growth rate over a two-week period.
ChIP-Seq
Cells were grown in 15 cm2 plates in either SCH772984 (1 µM) or DMSO (1:1000) in biologic replicate. For the one-week samples, 560,000 and 110,000 cells were plated in the SCH772984 and DMSO conditions, respectively, to achieve 70% confluence at one week. For the terminally resistant cells, evolved resistance was achieved as described above, and then this cell population was plated at 110,000 cells per 15 cm2 plate in biologic replicate and treated for one additional week with SCH772984. At this point, cells were cross-linked using 1% formaldehyde followed by 2.5 M glycine and washed with ice-cold PBS. Cross-linked cells were then scraped in RIPA buffer and snap-frozen. As previously described63, chromatin was sheared and antibody-conjugated (H3K27ac active motif: 39133) Protein A Dynabeads beads (Invitrogen) were used for chromatin extraction. Bound chromatin was eluted, crosslinking was reversed, and DNA was then purified by phenol chloroform extraction. Sequencing libraries were prepared using the KAPA Hyper Prep Kit (KAPA Biosystems). Sequencing was performed on an Illumina NextSeq. For data analysis, peaks were first identified using MACS2 with a configuration suitable to detect narrow peaks as those typically observed in H3K27ac data. A union peakset of all possible acetylation events identified across conditions was then defined. Using this common set, reads in peaks were computed using featureCounts with default parameters. Lastly, to detect differential binding events a negative binomial model using DESeq2 was applied to the counts, followed by a Wald test to compute p-values. Gained peaks represent both regions gaining or increasing acetylation when compared to control samples. Conversely, sites with depleted or decreased signal are referred to as lost peaks.
RNA sequencing and gene set enrichment analysis
RNA was extracted from cell pellets using the RNeasy Kit (Qiagen), and library preparation and sequencing was performed by the Duke Sequencing and Genomic Technologies Shared Resource. RNA-seq data was processed using the TrimGalore toolkit which employs Cutadapt to trim low quality bases and Illumina sequencing adapters from the 3’ end of the reads. Only reads that were 20nt or longer after trimming were kept for further analysis. Reads were mapped to the GRCh37v75 version of the human genome and transcriptome using the STAR RNA-seq alignment tool. Reads were kept for subsequent analysis if they mapped to a single genomic location. Gene counts were compiled using the HTSeq tool. Only genes that had at least 10 reads in any given library were used in subsequent analysis. Normalization and differential expression was carried out using the DESeq2 Bioconductor package with the R statistical programming environment. The false discovery rate was calculated to control for multiple hypothesis testing. Gene set enrichment analysis was performed to identify gene ontology terms and pathways associated with altered gene expression for each of the comparisons performed.
Long-term pharmacologic screens
Cells were plated at 20,000 cells per 10 cm plate in triplicate. The next day, drugs were added at the indicated doses. Inhibitors were selected at doses previously described to achieve target inhibition without significant growth suppression. Media and drug were then exchanged weekly until the control condition for each MAPK inhibitor grew to 90% confluence, at which point cells in all plates in that treatment condition were counted. Population doublings were calculated as Log2(final cell count/initial cells plated), and the average per condition was then compared to the control condition in order to calculate the doubling ratio.
Clonogenic growth assay
Cells were seeded at 2000–10,000 cells per well. The next day, cells were drugged at the indicated doses. At assay completion, plates were rinsed with PBS and fixed and stained with 0.5% (wt/vol) crystal violet in 6.0% (vol/vol) glutaraldehyde solution (Thermo Fisher Scientifics) for 30 min at room temperature. Plates were rinsed in distilled H2O and photographed the following day.
Generation of CRISPR/Cas9 knockout derivatives: CRISPR constructs were cloned into the lentiCRISPR v2 vector as previously described64. After lentivirus production, viral tittering, and transduction15, cells were replated into fresh media in 10 cm plates, and one day later puromycin was added (2 µg/mL). Two days later, puromycin was removed, and cells were considered stably transduced five days later. For double knockout experiments, we also used a modified version of the lentiCRISPR v2 vector (lentiCRISPR v2-Hygro)47, in which the puromycin resistance gene was exchanged for a hygromycin resistance gene. This allowed us to sequentially generate dual-knockout derivatives, and transduction was performed as described above, after the first knockout was considered stable.
Dirichlet Process Gaussian Process (DPGP) mixture model
In order to identify gene sets exhibiting similar gene expression trajectories, we applied a previously published DPGP model49. Briefly, DPGP simultaneously models data clusters using a Dirichlet process and dependencies on data timepoints with Gaussian processes. This joint model allows us to identify large and subtle differences in transcriptional profiles over time. Once DPGP clusters were established, changes in gene expression among each component was defined as the log2 fold change relative to DMSO-treated parental cells at each time point. Mean cluster expression was defined as the average log2 fold change of all cluster components at each time point relative to DMSO-treated parental cells.
Loss-of-function CRISPR/Cas9 screens
Criteria for library selection included the 200 genes most up- or downregulated at early drug exposure and at stable resistance, with priority based on the differential expression level and significance across two biological replicate conditions, each performed in technical triplicate (all 716 selected genes meeting these criteria had an absolute log2 fold change of >0.5 and a p < 1.0 × 10−5 in both models; Table S9). Also included were 100 control genes selected for their general essentiality or dispensability65, 50 internal control genes which exhibited minimal expression changes during the adaptive process, and 50 non-targeting control guides. The library was cloned into an established lentiviral system as previously described15,66, and all sgRNA sequences (five guides per genes and non-targeting controls) were selected from a reputable genome-wide library67. Drug-naïve and resistant cells were then separately transduced with library virus at a multiplicity of infection of 0.2 at 1000x coverage, and then these populations underwent seven days of puromycin selection. Following puromycin selection, each population was divided and treated with either SCH772984 (1 µM) or DMSO (1:1000). Each condition was conducted in biologically independent replicates and carried out at >1000x coverage for five weeks, and cells were split once they reached 80% confluence. At each split, excess cells beyond those needed to maintain 1000x coverage were pelleted and stored at −80 °C. Genomic DNA from these pellets was extracted with the DNeasy Blood & Tissue Kit (Qiagen). Amplification of the sgRNA barcodes and indexing of each sample was performed via two-step PCR as previously described15,64. To determine differences in sgRNA composition between samples, deep sequencing was performed using the Illumina Nextseq platform (single-ended 75 bp). As previously described15,64, barcoded reads were converted to guide-level counts and the fractional representation of each sgRNA construct was found by dividing the count of each sgRNA in a sample by the sum of all sgRNA counts in that sample. Construct-level depletion scores were collapsed to gene-level depletion scores by taking the average depletion score across five sgRNA constructs. All depletion/enrichment effects were reported as log2 ratios.
Resistance potential
MIA PaCa-2 cells were plated in six-well plates at a density of 500 cells per well. The following day, cells were treated with increasing concentrations of SCH772984, with all conditions performed in technical triplicate. Following two weeks of treatment, plates were rinsed with PBS and fixed and stained with 0.5% (wt/vol) crystal violet in 6.0% (vol/vol) glutaraldehyde solution (Thermo Fisher Scientifics) for 30 min at room temperature. Plates were rinsed in distilled H2O and photographed the following day. Colonies were hand-counted using scanned images at 400x, and resistance potential was defined as the number of colonies in drug-treated conditions relative to DMSO-treated cell.
Long-term tumoroid assay
Colorectal cancer tumoroids, generated and maintained as previously described51, we treated with DMSO alone (1:1000), Senexin A (1 µM), SCH772084 (1 µM), or a combination of both SCH772084 (1 µM) and Senexin A (1 µM). As previously described51, tumoroids were passaged for up to eight weeks, with counts performed at weekly time points. These patient-derived tumoroids complied with all relevant ethical regulations including the Declaration of Helsinki, and the protocol was reviewed and approved by the Memorial Sloan Kettering Cancer Center Institutional Review Board (IRB). Informed consent was obtained from all human participants.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Request for raw data files can be provided by KCW pending scientific review and a completed material transfer agreement. Requests for the data should be submitted to: kris.wood@duke.edu. Data uploaded to GEO can be accessed via the following accession numbers: GSE237177 GSE234378.
References
Ryan, M. B., Der, C. J., Wang-Gillam, A. & Cox, A. D. Targeting RAS-mutant cancers: is ERK the key? Trends Cancer 1, 183–198 (2015).
Samatar, A. A. & Poulikakos, P. I. Targeting RAS-ERK signalling in cancer: promises and challenges. Nat. Rev. Drug Discov. 13, 928–942 (2014).
Chapman, P. B. et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N. Engl. J. Med. 364, 2507–2516 (2011).
Flaherty, K. T. et al. Improved survival with MEK inhibition in BRAF-mutated melanoma. N. Engl. J. Med. 367, 107–114 (2012).
Pratilas, C. A. & Solit, D. B. Targeting the mitogen-activated protein kinase pathway: physiological feedback and drug response. Clin. Cancer Res. 16, 3329–3334 (2010).
Peng, S. B. et al. Inhibition of RAF Isoforms and Active Dimers by LY3009120 Leads to Anti-tumor Activities in RAS or BRAF Mutant Cancers. Cancer Cell 28, 384–398 (2015).
Hatzivassiliou, G. et al. Mechanism of MEK inhibition determines efficacy in mutant KRAS- versus BRAF-driven cancers. Nature 501, 232–236 (2013).
Morris, E. J. et al. Discovery of a novel ERK inhibitor with activity in models of acquired resistance to BRAF and MEK inhibitors. Cancer Discov. 3, 742–750 (2013).
Martinez-Garcia, M. et al. First-in-human, phase I dose-escalation study of the safety, pharmacokinetics, and pharmacodynamics of RO5126766, a first-in-class dual MEK/RAF inhibitor in patients with solid tumors. Clin. Cancer Res. 18, 4806–4819 (2012).
Germann, U. A. et al. Targeting the MAPK signaling pathway in cancer: promising preclinical activity with the novel selective ERK1/2 Inhibitor BVD-523 (Ulixertinib). Mol. Cancer Ther. 16, 2351–2363 (2017).
Zhang, C. et al. RAF inhibitors that evade paradoxical MAPK pathway activation. Nature 526, 583–586 (2015).
Vakana, E. et al. LY3009120, a panRAF inhibitor, has significant anti-tumor activity in BRAF and KRAS mutant preclinical models of colorectal cancer. Oncotarget 8, 9251–9266 (2017).
Lito, P. et al. Disruption of CRAF-mediated MEK activation is required for effective MEK inhibition in KRAS mutant tumors. Cancer Cell 25, 697–710 (2014).
Jaiswal, B. S. et al. ERK mutations and amplification confer resistance to ERK-inhibitor therapy. Clin. Cancer Res. 24, 4044–4055 (2018).
Anderson, G. R. et al. A landscape of therapeutic cooperativity in KRAS mutant cancers reveals principles for controlling tumor evolution. Cell Rep. 20, 999–1015 (2017).
Rusan, M. et al. Suppression of adaptive responses to targeted cancer therapy by transcriptional repression. Cancer Discov. 8, 59–73 (2018).
Zawistowski, J. S. et al. Enhancer remodeling during adaptive bypass to MEK inhibition is attenuated by pharmacologic targeting of the P-TEFb Complex. Cancer Discov. 7, 302–321 (2017).
Obenauf, A. C. et al. Therapy-induced tumour secretomes promote resistance and tumour progression. Nature 520, 368–372 (2015).
Blakely, C. M. et al. NF-kappaB-activating complex engaged in response to EGFR oncogene inhibition drives tumor cell survival and residual disease in lung cancer. Cell Rep. 11, 98–110 (2015).
Duncan, J. S. et al. Dynamic reprogramming of the kinome in response to targeted MEK inhibition in triple-negative breast cancer. Cell 149, 307–321 (2012).
Brown, R., Curry, E., Magnani, L., Wilhelm-Benartzi, C. S. & Borley, J. Poised epigenetic states and acquired drug resistance in cancer. Nat. Rev. Cancer 14, 747–753 (2014).
Sharma, S. V. et al. A chromatin-mediated reversible drug-tolerant state in cancer cell subpopulations. Cell 141, 69–80 (2010).
Cancer Genome Atlas Research Network Electronic address, a. a. d. h. e. & cancer genome Atlas Research, N. Integrated genomic characterization of pancreatic ductal adenocarcinoma. Cancer Cell 32, 185–203.e113 (2017).
Cancer Genome Atlas, N. Comprehensive molecular characterization of human colon and rectal cancer. Nature 487, 330–337 (2012).
Cancer Genome Atlas Research, N. Comprehensive molecular profiling of lung adenocarcinoma. Nature 511, 543–550 (2014).
Ebi, H., Faber, A. C., Engelman, J. A. & Yano, S. Not just gRASping at flaws: finding vulnerabilities to develop novel therapies for treating KRAS mutant cancers. Cancer Sci. 105, 499–505 (2014).
Al-Lazikani, B., Banerji, U. & Workman, P. Combinatorial drug therapy for cancer in the post-genomic era. Nat. Biotechnol. 30, 679–692 (2012).
Singh, H., Longo, D. L. & Chabner, B. A. Improving prospects for targeting RAS. J. Clin. Oncol. 33, 3650–3659 (2015).
Chandarlapaty, S. Negative feedback and adaptive resistance to the targeted therapy of cancer. Cancer Discov. 2, 311–319 (2012).
Lito, P., Rosen, N. & Solit, D. B. Tumor adaptation and resistance to RAF inhibitors. Nat. Med. 19, 1401–1409 (2013).
Collisson, E. A. et al. A central role for RAF->MEK->ERK signaling in the genesis of pancreatic ductal adenocarcinoma. Cancer Discov. 2, 685–693 (2012).
Pierobon, M. et al. Enrichment of PI3K-AKT-mTOR pathway activation in hepatic metastases from breast cancer. Clin. Cancer Res. 23, 4919–4928 (2017).
She, Q. B. et al. 4E-BP1 is a key effector of the oncogenic activation of the AKT and ERK signaling pathways that integrates their function in tumors. Cancer Cell 18, 39–51 (2010).
Singleton, K. R. et al. Melanoma therapeutic strategies that select against resistance by exploiting MYC-Driven evolutionary convergence. Cell Rep. 21, 2796–2812 (2017).
Creyghton, M. P. et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl Acad. Sci. USA 107, 21931–21936 (2010).
Shaffer, S. M. et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature 546, 431–435 (2017).
Leonard, B. et al. BET inhibition overcomes receptor tyrosine kinase-mediated cetuximab resistance in HNSCC. Cancer Res. 78, 4331–4343 (2018).
Zhao, B., Cheng, X. & Zhou, X. The BET-bromodomain inhibitor JQ1 mitigates vemurafenib drug resistance in melanoma. Melanoma Res. 28, 521–526 (2018).
Esteve-Arenys, A. et al. The BET bromodomain inhibitor CPI203 overcomes resistance to ABT-199 (venetoclax) by downregulation of BFL-1/A1 in in vitro and in vivo models of MYC+/BCL2+ double hit lymphoma. Oncogene 37, 1830–1844 (2018).
Luyties, O. & Taatjes, D. J. The Mediator kinase module: an interface between cell signaling and transcription. Trends Biochem. Sci. 47, 314–327 (2022).
Pelish, H. E. et al. Mediator kinase inhibition further activates super-enhancer-associated genes in AML. Nature 526, 273–276 (2015).
Dale, T. et al. A selective chemical probe for exploring the role of CDK8 and CDK19 in human disease. Nat. Chem. Biol. 11, 973–980 (2015).
Porter, D. C. et al. Cyclin-dependent kinase 8 mediates chemotherapy-induced tumor-promoting paracrine activities. Proc. Natl Acad. Sci. USA 109, 13799–13804 (2012).
Steinparzer, I. et al. Transcriptional responses to IFN-gamma require mediator kinase-dependent pause release and mechanistically distinct CDK8 and CDK19 functions. Mol. Cell 76, 485–499.e488 (2019).
Poss, Z. C. et al. Identification of mediator kinase substrates in human cells using cortistatin A and quantitative phosphoproteomics. Cell Rep. 15, 436–450 (2016).
Audetat, K. A. et al. A kinase-independent role for cyclin-dependent kinase 19 in p53 response. Mol. Cell Biol. 37. https://doi.org/10.1128/MCB.00626-16 (2017).
Chirieleison, S. M. et al. Nucleotide-binding oligomerization domain (NOD) signaling defects and cell death susceptibility cannot be uncoupled in X-linked inhibitor of apoptosis (XIAP)-driven inflammatory disease. J. Biol. Chem. 292, 9666–9679 (2017).
Misale, S. et al. Vertical suppression of the EGFR pathway prevents onset of resistance in colorectal cancers. Nat. Commun. 6, 8305 (2015).
McDowell, I. C. et al. Clustering gene expression time series data using an infinite Gaussian process mixture model. PLoS Comput. Biol. 14, e1005896 (2018).
Jackson, S. M. et al. Structural basis of small-molecule inhibition of human multidrug transporter ABCG2. Nat. Struct. Mol. Biol. 25, 333–340 (2018).
Ganesh, K. et al. A rectal cancer organoid platform to study individual responses to chemoradiation. Nat. Med. 25, 1607–1614 (2019).
Mallinger, A. et al. Discovery of potent, orally bioavailable, small-molecule inhibitors of WNT signaling from a cell-based pathway screen. J. Med. Chem. 58, 1717–1735 (2015).
Papke, B. & Der, C. J. Drugging RAS: Know the enemy. Science 355, 1158–1163 (2017).
Cox, A. D., Fesik, S. W., Kimmelman, A. C., Luo, J. & Der, C. J. Drugging the undruggable RAS: mission possible? Nat. Rev. Drug Discov. 13, 828–851 (2014).
Tempero, M. A. et al. Pancreatic adenocarcinoma, Version 2.2017, NCCN clinical practice guidelines in oncology. J. Natl Compr. Cancer Netw. 15, 1028–1061 (2017).
Ettinger, D. S. et al. Non-small cell lung cancer, Version 5.2017, NCCN clinical practice guidelines in oncology. J. Natl Compr. Cancer Netw. 15, 504–535 (2017).
Benson, A. B. 3rd et al. Colon Cancer, Version 1.2017, NCCN clinical practice guidelines in oncology. J. Natl Compr. Cancer Netw. 15, 370–398 (2017).
Benson, A. B. 3rd et al. Rectal Cancer, Version 2.2018, NCCN clinical practice guidelines in oncology. J. Natl Compr. Cancer Netw. 16, 874–901 (2018).
Plotnikov, A., Zehorai, E., Procaccia, S. & Seger, R. The MAPK cascades: signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Biophys. Acta 1813, 1619–1633 (2011).
Vogel, C. & Marcotte, E. M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 13, 227–232 (2012).
Ang, H. X. et al. Cooperative regulation of coupled oncoprotein synthesis and stability in triple-negative breast cancer by EGFR and CDK12/13. Proc. Natl Acad. Sci. USA 120, e2221448120 (2023).
Waters, A. M. et al. Targeting p130Cas- and microtubule-dependent MYC regulation sensitizes pancreatic cancer to ERK MAPK inhibition. Cell Rep. 35, 109291 (2021).
Levandowski, C. B. et al. The Delta40p53 isoform inhibits p53-dependent eRNA transcription and enables regulation by signal-specific transcription factors during p53 activation. PLoS Biol. 19, e3001364 (2021).
Lin, K. H. et al. Systematic dissection of the metabolic-apoptotic interface in AML Reveals Heme Biosynthesis to Be a Regulator of Drug Sensitivity. Cell Metab. 29, 1217–1231.e1217 (2019).
Hart, T. et al. High-resolution CRISPR screens reveal fitness genes and genotype-specific cancer liabilities. Cell 163, 1515–1526 (2015).
Shalem, O. et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science 343, 84–87 (2014).
Wang, T., Wei, J. J., Sabatini, D. M. & Lander, E. S. Genetic screens in human cells using the CRISPR-Cas9 system. Science 343, 80–84 (2014).
Acknowledgements
National Institutes of Health F32CA180569 (D.P.N.), National Institutes of Health P30CA008748 (D.P.N.), National Institutes of Health R01CA207083 (KCW), National Institutes of Health R01CA263593 (K.C.W.), National Institutes of Health R01CA175747 (C.J.D.), National Institutes of Health U01CA199235 (C.J.D.), National Institutes of Health P01CA203657 (C.J.D.), National Institutes of Health R35CA232113 (C.J.D.), National Institutes of Health P30CA008748 (J.J.S.), National Institutes of Health R37CA248289 (J.J.S.), Department of Defense Lung Cancer Research Program W81XWH2110362 (K.C.W.), Duke University School of Medicine/Duke Cancer Institute start-up funds (K.C.W.), American Cancer Society PF 18-061 (A.M.W.)
Author information
Authors and Affiliations
Contributions
Conceptualization: D.P.N., C.A.M., E.F.P., J.J.S., T.E.R., C.J.D., D.J.T., and K.C.W. Methodology: D.P.N., C.A.M., E.F.P., J.J.S., T.E.R., C.J.D., D.J.T., and K.C.W. Investigation: C.A.M., A.M.W., A.B., A.L., J.C.R., C.G.C.-S., A.E.S., C.W., M.C., C.B.W., D.E.K., and M.P. Visualization: D.P.N., A.L. Supervision: K.C.W., E.F.P., J.J.S., T.E.R., C.J.D., D.J.T. Writing—original draft: D.P.N., K.C.W. Writing—review & editing: All authors.
Corresponding author
Ethics declarations
Competing interests
K.C.W. is a founder, consultant, and equity holder at Tavros Therapeutics and Celldom and has performed consulting work for Guidepoint Global, Bantam Pharmaceuticals, and Apple Tree Partners. C.J.D. is an advisory board member for Deciphera Pharmaceuticals, Mirati Therapeutics, Reactive Biosciences, Revolution Medicines and SHY Therapeutics, has received research funding support from Deciphera Pharmaceuticals, Mirati Therapeutics, Reactive Biosciences, Revolution Medicines and SpringWorks Therapeutics, and has consulted for Day One Biotherapeutics, Eli Lilly, Jazz Therapeutics, Ribometrix, Sanofi, and Turning Point Therapeutics. The remaining authors declare no competing interests. JJS received travel support for fellow education from Intuitive Surgical (August 2015). He also served as a clinical advisor for Guardant Health (March 2019) and as a clinical advisor for Foundation Medicine (April 2022). He served as a consultant and speaker for Johnson and Johnson (May 2022). And he serves as a clinical advisor and consultant for GlaxoSmithKline (2023-24).
Ethical approval
All co-authors have fulfilled authorship criteria as determined by Nature portfolio.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nussbaum, D.P., Martz, C.A., Waters, A.M. et al. Mediator kinase inhibition impedes transcriptional plasticity and prevents resistance to ERK/MAPK-targeted therapy in KRAS-mutant cancers. npj Precis. Onc. 8, 124 (2024). https://doi.org/10.1038/s41698-024-00615-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41698-024-00615-9
- Springer Nature Limited