
Abstract
We tested if the personality of captive-raised western ringtail possums (Pseudocheirus occidentalis), and the impact of other variables would influence their survival after release using radiotelemetry. We hypothesised a greater survival for individuals: (i) bold; (ii) habituated in advance to food collected from the release area; (iii) juveniles instead of adults, because more easily tolerated by wild individuals, and (iv) released in new dreys (nests) as they would not have the strong odour of old dreys and would be less attractive to foxes. After 3 months of radio tracking, out of 143 possums released, 79 died: 51 (64.6%) were preyed upon by European red foxes (Vulpes vulpes). Bold or female individuals had higher survival rates than shy or male individuals (survival rate bold: 53%, shy: 41%, p = 0.046, hazard ratio = 0.352, 95% CI HR [0.126, 0.979]; survival rate females: 44%, males: 35%, p = 0.007, hazard ratio = 2.811, 95% CI HR [1.322, 5.976]). Shooting was a more effective fox control strategy to improve survival compared to baiting (p = 0.019, hazard ratio = 0.167, 95% CI HR [0.038, 0.742]). Our results demonstrate that the control of introduced predators is critical for the success of reintroductions of this critically endangered species.
Similar content being viewed by others

Introduction
The introduction of invasive species in Australia (especially foxes, Vulpes vulpes, and cats, Felis catus) has strongly impacted native species’ populations1,2,3. To date, about 100 Australian species of plants, birds, amphibians, mammals and invertebrates are extinct, and mammals are the most impacted4,5.
To support wild populations, many studies have tried to identify the most appropriate conservation actions, including rehabilitation and reintroduction6,7,8,9,10. Several variables can influence the success of releases of animals, such as, for example, health screening, physiological checks, parasites, animal habituation to humans and personality6. Evaluating the personality of animals before their release is considered important for the success of reintroduction, because it can affect survival11,12,13,14.
In vertebrate species, personality varies along an axis, with extremes defined as ‘bold’ and ‘shy’15,16, with bold individuals dispersing and exploring more, and coping better with new and stressful situations. Although this risk-taking may lead to danger or encounters with predators, it may also help them to find additional trophic resources necessary for survival. Conversely, shy individuals are neophobic: dispersing and exploring less, and preferring to adapt to a new, stressful situation, rather than changing the situation15,17,18,19. Personality, by definition ‘a set of multidimensional traits characteristic of an individual, stable across time and situations within the same individual’, influences the animal’s response to new stimuli20. Thus, it is possible to assess the personality of an animal and predict its response by exposing it to a new challenging environment11,22.
A widely adopted method to assess the personality of laboratory and wild animals, including possums, is the open field test (e.g.14). During the test, the animal is placed in a bare enclosed arena and allowed to move freely for a predetermined amount of time. The behaviour inside the arena reflects the individual's reaction to stressful events, and consequently its behavioural patterns in response to, for example, social interactions or the presence of predators23.
A major factor impacting the success of reintroductions of native species in Australia is predation by the European red fox and the cat24,25. The predation rate exerted by these predators is much higher than that of native predators, which is why in many projects animals were released into predator-free sites, such as islands or effectively fenced areas26,27. Predator-free areas, however, are insufficient to sustain all populations of threatened native species, and it is therefore necessary to release animals in areas where predators are present while implementing predator control28.
An environmental variable that is not often considered during reintroductions is acclimation to new food sources present at release sites. During the period of adjustment after their release, the animals must learn how to find adequate food and their gut flora needs to adjust to the change in diet (e.g.29,30). Gut pathogens, phenotypic plasticity of the gastrointestinal system and healthy digestion by microbes or symbionts are all food-linked factors that can influence the outcome of translocations31,32. For example, peninsular bighorn sheep (Ovis canadensis) fed with high-quality food and native vegetation prior to the release did not survive after being released in the harsh environment of Southern California33. Therefore, it is critical to investigate how the source of food impacts the survival of individuals to establish a successful re-introduction or release.
Among the species currently experiencing a decline, is the western ringtail possum, Pseudocheirus occidentalis, a species endemic to the southwest of Western Australia, recently classified as critically endangered5. Because of its limited distribution and recent dramatic population decrease, it is important to increase our knowledge of this species to implement appropriate conservation strategies and improve its conservation outlook4,34. Understanding the factors that most impact the survival of this species could optimise the outcome of translocations and increase the size and genetic diversity of the wild population.
In this study, we tested if the personality of captive-raised western ringtail possums and other environmental factors including acclimation to a novel food source, releasing juveniles instead of adults, releasing animals in a new drey (nest) to avoid attracting foxes with the strong odour of the old dreys and releasing them at first in a predator-proof area would influence survival rates of the animals after their release. We hypothesised higher chances of survival for bold individuals, for animals that were accustomed to the food found at the release sites, for juveniles compared to adults because they would be tolerated by resident wild possums, and for animals released in new dreys compared to those released in old dreys.
The western ringtail possum is particularly prone to fox predation, especially when burdened by radio transmitter collars heavier than 20 g35. Therefore, we were required to release the possums with radio collars weighing less than 20 g and in areas with existing fox control in place, so we did not expect that predation would impact the survival of the released individuals.
Materials and methods
Animals and housing
The sample consisted of 143 western ringtail possums, released in 7 separate cohorts. All possums were orphans raised by volunteer carers from the organization Fostering and Assistance for Wildlife Needing Aid (FAWNA) Inc. When the possums reached a weight of at least 600 g and at least six weeks before being released, they were transferred from the homes of the carers to a pre-release facility at the former Iluka Capel Wetlands site in Capel (Western Australia). Additional information about the pre-release conditions can be found in the Supplementary Material (S1).
Personality test
We used an open field test to assess the personality of the possums. The test arena measured 200 × 200 cm and was surrounded by walls 120 cm tall. The floor was marked with tape to subdivide it into 9 equal size squares, each being about three times the size of the test subject. The test subject was introduced into the arena through a hatch and filmed with a GoPro camera for 10 min (Hero4, Woodman Labs, Inc, San Mateo, California). Exploratory behaviours (locomotion, location and time in each square including crossing square, jumping, walking and placing the paws on the wall) and other behavioural patterns (defecation, urination, self-grooming) were blinded analysed by an observer using specialised software (Boris v.8.27.1, Friard & Gamba, Department of Life Sciences and Systems Biology University of Torino, Italy; https://www.boris.unito.it/ ). The test was not repeated to avoid habituation. At the end of the test, the animal was placed into a calico bag and returned to its enclosure. Since ringtail possums are nocturnal, the test was conducted during the day when they are less active, to make capture less stressful for the animals, and at least two hours before sunset to avoid a bias in individual activity.
Anaesthesia and radio-collaring
Animals were anaesthetised by a veterinarian through use of isoflurane gas inhalation to be handled and fitted with radio transmitter collars to ensure minimum stress and optimum collar fit.
During the anaesthesia, animals were weighed and measured (pes, head length, body length, tail length, neck circumference, testes width and length for males) and fitted with a radio-collar. The radio-collars were expandable drop-off collars (custom VHF core, model CCC000052; Lotek, Havelock North, New Zealand) with mortality function (activated after 4 h of inactivity) weighing a maximum of 20 g, as recommended by Yokochi35,36 so as not to affect the survival of the animals. The collars were made of leather with a 10 mm section of elastic that could expand to 20 mm allowing for growth, escape if entangled and it also degraded over time so that collars that could not be retrieved eventually dropped off. For the first release collars weighed around 20 g due to the weight of the battery, which had a life of 5 months. For subsequent releases, lighter collars weighing 13–15 g were used and they were programmed with a duty cycle of 9 h on and 15 h off to extend the battery life and radio-track animals for a minimum of 12 weeks.
In addition to fitting the radio-collar and collecting standard measurements, we microchipped each possum, collected a tissue sample for future genetic analysis and attached a reflective tag (Monel Style #1005–1, to which we added a strip of reflective tape sealed with heat shrink) on the left ear for males and on the right ear for females to facilitate identification after release. A blood sample was collected by the veterinarian from the lateral tail vein and sent to VetPath Laboratory Services (1 Sabre Cres, Jandakot Western Australia) for the analysis of haematocrit (HCT), red blood cells (RBC), mean corpuscular volume (MCV), mean corpuscular haemoglobin concentration (MCHC), mean corpuscular haemoglobin (MCH), white blood cells (WBC), Band cells (BANDS), monocytes (MONO), basophils (BASO), lymphocytes (LYMPH), neutrophils (NEUTRO) and eosinophils (EOSINO). The analysis of these variables served to assess the overall physiological situation of the animals before release, as already used for this and other species (e.g.19,25).
Release site selection
Three release areas were selected during the three years of this project (Table 1). Additional information on area selection protocol and selected sites can be found in Supplementary Material (S2).
All possums were released in their own dreys by hanging each drey containing a possum on tree branches 3–4 m above the ground during the early afternoon, when animals are usually asleep, so that they could make their way out of the dreys at nightfall.
Environmental variables affecting survival
Food source
The first group of 20 possums released in Wyadup was tested to determine if feeding western ringtail possums with food sourced from the release site would increase survival of released animals. Half of the possums (10) were fed with food collected from the area surrounding the pre-release facility, while half of the possums (10) were fed with food collected from the release site. To check if there was a preference in food consumption, we selected four species of trees that were common in the area and known to be eaten by western ringtail possums: peppermint (Agonis flexuosa), marri (Corymbia calophylla), jarrah (Eucalyptus marginata) and flooded gum (Eucalyptus rudis). Branches of these plants were placed in the enclosures’ food holders (described in Supplementary Material, S1). Individuals were weighed once a week to monitor changes in weight.
Juveniles/adults
The first group of possums released in Wyadup experienced territorial conflicts with wild possums: four females were observed with wounds on the body and on the tail. Introduced possums were forced to move away from the release site due to these territorial conflicts and then they may have been preyed upon. Following a suggestion from the carers, in the second release we released 10 juveniles and 10 adults on the same property to test if juveniles would be more easily accepted by wild possums and if this reduced predation rates. Because all the animals involved in the study were orphan pouch young when moved into captivity, we knew the age of the animals. All adults had reached sexual maturity at the time of release.
Dreys
Because predation rate was high in the first two releases, and it was possible for us to smell a possum when walking in the bush during radio-tracking, we supposed that releasing 20 animals 20/30 m apart from each other and in dreys previously utilised in the pre-release facility could attract foxes to the area due to the strong odour of these old dreys. Considering this possibility, in the third and fourth releases in Glenbourne, we released half of the animals (10 each time, 20 in total) in new dreys, spaced approximately 40 m apart from each other, and half of the animals (10 each time, 20 in total) in the dreys previously utilised by the animals in the pre-release facility.
Predator-proof area
Since in the first four releases most of the animals that died were preyed upon by foxes in the first four weeks, we tested if releasing animals in a predator-proof area first and then moving them outside of the predator-proof area would improve survival. So, we released 20 animals in the predator-proof area located at 5566 Caves Road, Gracetown. After four weeks, we trapped 7 possums and released them in the second and third sites located at 110 and 111 Ellenbrook Rd, outside of the predator-proof area. Ten additional possums were released at 110 and 111 Ellenbrook Rd and four more inside the fenced area as control groups.
Radio-tracking
Radio-collared animals were monitored for at least three months after release.
Each individual was located in the following way:
-
Week 1 – once every 24 h, alternating day and night locations.
-
Weeks 2–5 – Every 2nd or 3rd day for survival, either by triangulation or by homing if the collar had shifted to mortality mode. One precise day time and one night location were collected each week for calculation of home ranges.
-
Week 6–12 – Weekly, one day and one night location to monitor movements, dispersal, mortality and to calculate home ranges.
Day locations were collected between 9:00 and 13:00 h for the releases conducted in Wyadup, and between 13:00 and 18:00 for all other sites. Night locations were collected between 20:00 and 1:00. For each location, we recorded the GPS coordinates, the height above the ground where the possum was located, the height of the vegetation at that point, the species of plant the animal was on and if there were other western ringtail possums or brushtail possums (Trichosurus vulpecula) within 5–6 m radius.
After three months, animals that still had their collars on were captured to remove the collars using wire cage traps (22 × 22 × 45 cm, Sheffield Wire Products, Welshpool Western Australia) baited with fruit and strategically placed on branches37. Each possum was weighed and the pouches of females were inspected for the presence of pouch young.
Mortality and survival
When a collar was detected on mortality mode indicating that a possum had died or that it had dropped its collar, we homed on the signal to retrieve either the body or the collar. Retrieved bodies were necropsied when possible. Conditions of carcasses and collars were observed to spot signs of wounds or teeth marks. If found, we used a sterile swab to collect traces of predator saliva from the collar or the carcass. The presence of predators’ DNA was detected through melt curve analysis38 by Helix Molecular Solutions (PO Box 155 Leederville, Western Australia 6153). The test detects the presence of DNA from fox, cat, dog (Canis lupus familiaris) or chuditch (Dasyurus geoffroii, a native predator).
Statistical analyses
Three possums were excluded from the analyses: one individual was found with a severe injury during the radio-tracking and was brought back into care; two other possums disappeared immediately after being released and no data could be collected from these possums. So, the total sample consisted of 140 individuals. We used Principal Component Analysis (PCA) with VARIMAX rotation to analyse the behaviours recorded during the open field test and during the radio-tracking.
We developed a personality index to assess personality. This was calculated by summing the individual values for the first two factors from the PCA on the whole sample (F1 and F2) for each animal. We ranked animals from the highest to the lowest value: a high value of the sum (F1 + F2) indicated a bold animal.
Since there is a lack of knowledge of behaviours related to the personalities of western ringtail possums, we correlated the factors of the PCA to other variables that have been found to be linked to boldness or shyness in other species39. We used the Pearson correlation when the variables had a normal distribution, or a Spearman rank correlation coefficient when the normality of the distribution of the variables was violated.
Because some animals died within few days after release, and some of the blood samples could not be analysed, we did not have all the information for all the variables for all the animals, so we ran different PCAs (with VARIMAX rotation) for each set of complete data. We run a PCA on animals for which we had the home range and core areas, a PCA on animals for which we had the HCT value and another PCA on animals for which we had the physiological variables measured in the blood samples.
Home range area and core area of the individuals (respectively 95 and 50 percentiles) were calculated with the kernel density estimator through QGis (Geographic Information System, version 3.32, QGIS Association. http://www.qgis.org). We removed the data for the first three weeks from the calculation, because during this time the animals were dispersing and had not yet established their home ranges.
To evaluate dispersal from the point of release, we calculated the distance between the release point and the farthest location recorded for each animal. The effect of gender on dispersal was tested using a one-way ANOVA to identify if males dispersed more than females from the release site as observed in similar species40.
The effect of personality on home range, core areas and height in the vegetation where the animal was located was analysed using a one-way ANOVA.
The analyses on survival involved 128 possums; because 11 animals lost their collar (e.g. the collars were found on branches or on the ground) and one had a collar whose frequency had a strong interference, probably due to a source of radio waves in the area, we had no information about the fate of those animals, so they were excluded from the analyses and we run another PCA on the remaining animals. The personality index was calculated as described above by summing the individual scores of the resulting factors of the PCA (F1 and F2) and ranking them from high (bold) to low (shy), following a similar procedure used in dogs19.
Since we tested a different variable in each release, we ran four different Chi-square tests of independence to check whether survival was increased by:
-
Feeding possums with food collected from the release site;
-
Releasing juveniles or adults;
-
Releasing animals in new, clean dreys;
-
Releasing animals in a predator-proof area for four weeks and then moving them outside.
A Cox Regression survival analysis was used to assess the variables hypothesised to influence survival in the whole sample (personality, sex, the different releases, average height on the vegetation, maximum distance from release point, season, home range 50th percentile, home range 90th percentile). A backward stepwise regression was run to exclude irrelevant variables and improve the strength of the model. The criteria to remove a variable was a probability of F > 0.100. The remaining variables were analysed using the Cox Regression survival analysis.
All the analyses were performed using the IBM SPSS software (Version 28.0. Armonk, NY: IBM Corp; https://www.ibm.com/spss ).
Animal ethics
The Animal Ethics Committee of The University of Western Australia approved all the procedures described in this article (RA/3/100/1610). We conducted our fieldwork following The Australian Code of Practice for the Care and Use of Animals for Scientific Purposes endorsed by the National Health and Medical Research Council of Australia (NHMRC, 2004), and following the Department of Biodiversity, Conservation and Attractions’ (DBCA) Standard Operating Procedures41.
This study was reported in accordance with ARRIVE guidelines.
Results
Personality test
In all the PCAs performed, the behaviours were clustered in a similar fashion. We extrapolated two factors for each PCA (Supplementary Material, Tables S1, S2, S3 and S4). The cumulative variance for all possums was 62.46% in the first PCA (N = 140), 63.71% in the second PCA (80 possums with home range), 62.39% in the third PCA (138 animals for which we have measured the HCT) and 62.42% in the fourth PCA (116 animals for which we had data for the haemogram and the white blood cell counts). Correlations of 0.31 and above were considered significant. The first factor (F1) was positively correlated with the number of squares crossed, jumping, time spent walking and placing the paws on the wall, all measures of activity during the open field test. The first factor was defined as ‘Activity’: possums with a high score were very active during the open field test. The second factor (F2) was positively correlated with time spent in the centre and side squares, and negatively correlated with the time spent in a corner square and scratching the wall. The second factor was defined as ‘Bravery’: possums with a high individual score with this factor appeared to be less scared as they spent more time in the central square where they would be more exposed to potential threats; they also scratched the walls less.
We determined that the two factors were related to bold personalities.
The sum of the individual scores for F1 and F2 of the first PCA (N = 140) was normally distributed. The personality typology was classified using percentiles: 35 possums with a F1-F2 sum that fell in the 25th percentile were defined as ‘Shy’, 71 possums with a F1-F2 sum that fell in between the 25th and the 75th percentile were defined as ‘intermediate’ and 34 possums with a F1-F2 sum that fell in between the 75th and the 100th percentile were defined as ‘Bold’ (Table 2).
The results of the PCAs used to link behaviours to bold personalities are described below.
Sixty individuals died or went missing during the first three weeks of radio-tracking, so a total of 80 individuals were included in the calculation of the core area and home range. The average core area was 0.072 ha (95% CI [± 0.013 ha]) and the average home range was 0.270 ha (95% CI [± 0.035 ha]). Females and males had similar mean values for core areas and home range areas (core areas xf = 0.074 ha, xm = 0.070 ha; home range areas xf = 0.279 ha, xm = 0.259 ha).
The second factor of PCA (F2: ‘Bravery’) was correlated with the core area (Spearman rank correlation, N = 80, two-tailed p-value = 0.014, rho = 0.274).
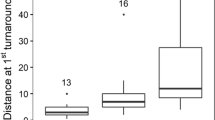
Females seemed to disperse, on average, more than males from the release site (average dispersal females: 664 m, average dispersal males: 318 m), even if the difference was not significant (one-way ANOVA: F1,138: 1.410, p = 0.237).
The sample size (N), mean, median and standard deviations of HCT, RBC, MCHC, MCH, WBC, BANDS BASO, NEUTRO and EOSINO are summarised in the Supplementary Material (Table S5).
There was a positive correlation between F1 (‘Activity’) and haematocrit (Pearson correlation, N = 138, r = 0.175, p = 0.040): animals with a higher F1 value had a higher haematocrit level.
We found a positive correlation between F1 and mean corpuscular volume and between F2 (‘Bravery’) and red and white blood cells; a negative correlation between F1 and mean corpuscular haemoglobin concentration and between F2 and mean corpuscular haemoglobin; a positive correlation between F1 and band cells, monocytes and basophils; and a negative correlation between F1 and lymphocytes (Supplementary Material, Table S6).
Survival and ecology
Baiting as a strategy to control foxes was not effective in this study: no fox ate the poisoned eggs laid for them in the first release at Wyadup and the survival rate of possums was 5%. In the second release, after the shooter eliminated four foxes, the survival rate of possums increased to 20%. In the releases at Gracetown, the survival rates increased to 25% in 2021 after the shooter eliminated 40 foxes and to 64% in 2022, after the shooter eliminated > 80 foxes. During the last release in Yalgorup National Park in which 15 of the 29 possums received fox awareness training before the release the survival rate was 44% (Table 3).
There was a difference, but not significant, in the average home range and core area between bold (mean core area: 0.087 ha, mean home range: 0.312 ha), intermediate (mean core area: 0.076 ha, mean home range: 0.258 ha) and shy possums (mean core area: 0.055 ha, mean home range: 0.252 ha; one-way ANOVA on core areas: N = 79, F2,77: 1.741, p = 0.182; one-way ANOVA on home ranges: N = 79, F2,77: 0.986, p = 0.378). The difference remained not significant when we compared only bold and shy individuals (one-way ANOVA on core areas: N = 46, F1,45: 3.568, p = 0.065; one-way ANOVA on home ranges: N = 46, F1,45: 1.585, p = 0.215). Similarly, there was a difference in the maximum distance from the release point among the three personalities, but the difference was not significant, even removing the intermediates from the test (one-way ANOVA between bold, shy and intermediate on dispersal: N = 135, F2,133: 1.544, p = 0.217; one-way ANOVA between bold and shy on dispersal: N = 66, F1,65: 1.084, p = 0.302;). There was also no difference in the average height of animals in the vegetation, which was about 5.5 m for bold, intermediate and shy animals (one-way ANOVA between bold, shy and intermediate on average height: N = 46, F2,133: 0.021, p = 0.979; one-way ANOVA between bold, shy and intermediate on average height: N = 66, F1,65: 0.039, p = 0.843: Supplementary Material, Table S7).
The first two factors of the PCA made on the possums whose fate we knew at the end of the three months of radio-tracking (N = 128) explained 62.46% of the total variance. The variables were grouped in the same way as the PCAs described above, so the definition of the factors remained the same: F1 was defined as ‘Activity’ and F2 was defined as ‘Bravery’. The behavioural categories of this new PCA and relative loadings are summarised in Supplementary Material, Table S8.
Similarly to the PCAs used to determine personality of all the possums (see section "Personality test"), the sum of the individual scores of both F1 and F2 of the PCA made on the 128 possums whose fate we knew was calculated and it was normally distributed. The percentiles were utilised to classify the personality: 32 possums with a F1-F2 sum that fell in the 25th percentile were defined as ‘Shy’, 64 possums with a F1-F2 sum that fell in between the 25th and the 75th percentile were defined as ‘intermediate’, 32 possums with a F1-F2 sum that fell in between the 75th and the 100th percentile were defined as ‘Bold’ (Table 4).
At the end of the radio-tracking across all seven releases combined, 79 possums died and 49 survived (32 females, 17 males). Fifty-eight died by predation (51 by foxes, four by carpet pythons, Morelia spilota; two by cats; one by a bird of prey) and two by drowning (in a dam and a stream respectively), while the cause of death of the remaining 19 could not be determined.
The backward stepwise regression excluded season, the average height of the animal on the vegetation, area of the core home range (50th percentile) and maximum distance from the release point as relevant variables. Then, the new model included: personality (p = 0.020), sex (p = 0.029), home range (95th percentile) (p = 0.066) and the different releases (p = 0.039). The Cox Regression run on these variables showed that bold animals had a higher survival rate than shy individuals (survival rate bold: 53%, shy: 41%, p = 0.046, hazard ratio = 0.352, 95% CI HR [0.126, 0.979], Fig. 1a) and females had a higher survival rate than males (survival rate females 44%, males: 35%, p = 0.007, hazard ratio = 2.811, 95% CI HR [1.322, 5.976], Fig. 1b). The possums released in the fifth and sixth groups in Gracetown in 2022 and in the seventh group in Yalgorup in 2022 had higher survival rates compared to the possums released in the first group in Wyadup in 2019 (respectively: p = 0.019, hazard ratio = 0.167, 95% CI HR [0.038, 0.742]; p = 0.016, hazard ratio = 0.130, 95% CI HR [0.025, 0.686] Fig. 1c).

Cumulative survival for western ringtail possums (Pseudocheirus occidentalis) after release: a) shy (in orange) compared to bold (in red); b) males (in blue) compared to females (in pink); and c) first release (in bright red) compared to fifth and sixth release (in azure) and seventh release (in green), during which animals received an fox awaress training (see the information about the release sites in the Supplementary Materials section, S2).. A vertical drop in the curves indicates an event of death.
Our results did not identify any effects of environmental factors on the possums’ survival (food sources: N = 25, χ2 = 1.053, df = 1, p = 0.305; juveniles vs adults: N = 18, χ2 = 0.055, df = 1, p = 0.814; old dreys vs new dreys: N = 39, χ2 = 0.208, df = 1, p = 0.648; pre-release in a predator-proof area: N = 34, χ2 = 0.273, df = 1, p = 0.601).
Discussion
Our results demonstrated that the personality of western ringtail possums can be evaluated with an open field test and support the hypothesis that bold individuals would have greater survival rates than shy individuals.
Our findings show that the open field test is a valid method to assess personality in the western ringtail possum, as already demonstrated in many species, including other marsupials14,42 and confirm the wide use of the open field test to assess personality, and the possibility of adapting this test from one species to another.
The PCA analysis of behavioural data collected in the open field test identified two main factors, that we defined as ‘Activity’ and ‘Bravery’. Personality is a complex set of multidimensional traits and cannot be reduced to only part of the behaviours exhibited by the individual20. We calculated the sum of the individual scores on both F1 and F2 to combine the different aspects of the personality of the animals and consequently the best way to assess their personality. Clearly, this approach to assess personality was shown to be valid since the behaviours clustered in the same way in all the PCAs performed in the study.
Not surprisingly, we found that physiological variables measured in the blood samples were related to personality: HCT was positively correlated with the factor ‘Activity’. HCT is higher in animals or species that require greater storage of blood oxygen or more resistance, or in situations where the oxygen demand is increased like migration or physical exertion. Exercise, movement or dehydration can cause a subsequent increase in HCT43,44. This may be the case for the western ringtail possum, which is an endemic species of Western Australia, where water is often scarce and even the foliage, which is the possums' main source of sustenance including water, has a low water content. Similarly, dehydration might explain the positive correlation between the factor ‘Activity’ (F1) and MCHC, as shown in humans (e.g.45).
Individuals can be adversely affected by stress depending on their personality16,19,46,47,48,49. Shy individuals do not cope well with stress, and they often develop stress-related diseases. This is reflected in the physiological state of the individual50. The positive correlation between the F1 and white blood cell counts (WBC) suggests that the individuals with a low individual score for F1 could be more vulnerable to infections and stress. Similarly, F2 and WBC were positively correlated, suggesting that individuals with a low score for F2 may have leukopenia, a condition where the number of white blood cells is not sufficient to cope with a possible infection51.
The values of the physiological variables measured in the blood samples fall within the range of values for the species available in the literature, suggesting that the health status of the animals was not compromised52.
Survival
Our results showed that, for captive-raised western ringtail possums, bold individuals are more likely to survive when released into the wild, if environmental variables, and in particular the presence of introduced predators, are not too unfavourable for survival. Being shy increases the risk of death by 35% compared to being bold. Similar results have been found in other projects on different species of mammals including marsupials (for example 11,53) and indeed a similar result was obtained in the brushtail possums, a closely related species54. An increase in exploratory behaviour in bold individuals would allow them to discover more or better trophic resources by increasing foraging opportunities, as previously showed in common brushtail possums54. In our study, the height at which the animal was located on the vegetation was not different between bold and shy individuals, which suggests that both groups use the canopy for feeding in a similar manner. Furthermore, we were not able to relate boldness to the home range or core area size, although the literature indicates that personality influences the use of resources, and consequently the use of space and dispersal (e.g.55,56). Surprisingly, in our study shy individuals dispersed more than bold, although not significantly: when experiencing territorial conflicts, shy possums probably moved and tried to find a different area with less competition, even if that led them to establish home ranges in sub-optimal habitats57. Bold individuals, instead, could have opted to stay and defend their newly established territories, as already reported in other species58.
Our study demonstrated that males had a risk rate of death 280% greater than females. The effect of sex on survival has already been described in marsupials. In reintroduced endangered eastern quolls (Dasyurus viverrinus), females were more likely to survive than males even in a predator-proof sanctuary27. In that study females also dispersed less than males, possibly because of the absence of resident quolls, so the competition was lower than in our study, where possums had to deal with already established wild individuals. We do not have enough data to suggest if females have greater survival rates only in marsupials, since in eutherian mammals males have higher survival rates59,60.
Despite our findings, releasing only bold and female individuals would not be ethical or ecologically viable. Genetic and behavioural diversity are essential for wild populations to cope with different contexts and problems, especially in a species where fragmentation has been identified as a major threat in the species’ recovery plan4. Behavioural diversity has also been found to be related to animal welfare61. Lastly, personality has a low heritability (h2 < 0.2), which allows inter-generational behavioral fluctuations that promote adaptation to environmental changes62. Behavioural personalities do have a genetic component, but also a responsiveness to environmental variation. For these reasons, reintroduction projects should consider personality only to better match the individuals to the characteristics of the release site: bold individuals are better suited to more challenging environments.
Home ranges, territorial conflicts and dispersal
The home ranges of the possums involved in this study were similar to those previously reported by Yokochi et al.36. However, in our study females had a slightly larger home ranges then males, in contrast to reports for this and other species of possums, as well as other marsupials24,36. Dimorphism in western ringtail possum is not very pronounced: males on average are only slightly larger than females, and this may explain why home ranges were similar. Many characteristics can influence the size of the home range in the western ringtail possum, like habitat quality, population density and competition for resources36. During the radio-tracking, we found females with wounds and scars suggesting that they had been in fights, but not males. These observations suggest that the wild females of this species are territorial, and defended their territories against the released females, while tolerating males probably because in the wild males move between the territories of different females in search of mates, as described in other marsupials63. The difference in dispersal between females and males that we observed, seems to support this hypothesis, even if the difference failed to reach significance. Introduced females were chased away by resident wild females to areas with a lower density of individuals. In the present study, many possums, after moving away from the release site, established home ranges in sub-optimal habitat, where competition was probably lower57. Males, on the other hand, did not disperse so much as they probably experienced fewer territorial conflicts. As shown in other species of marsupials by the overlapping of home ranges, male western ringtail possums are likely to tolerate other males63.
Our results for the home ranges of the females are similar to those found by Yokochi et al.36, confirming that the species has small home ranges, but not for males, reported to have larger home ranges than females. It is possible that the number of locations collected for the males was not sufficient to accurately determine the home range, since the males died more frequently and often earlier. Some males, however, even though they survived, had extremely small home range values: within the 17 males that survived, five had a home range value smaller than 0.07 ha, and three of them smaller than 0.035 ha. This could suggest that the difference in home range size between our study and the literature could be due to the permanence in captivity of the animals before the release, and not to a bias in our data collection.
Effective predator control is essential
Contrary to our initial expectation, fox predation strongly impacted the survival of released individuals, and effective fox control increased the survival of the released possums.
The difference in survival between the various releases in our study can be attributed to differences in effectiveness of fox control. For the first release, the site was baited with 1080 poisoned eggs, but the foxes ignored the bait and consumed most of the released possums instead. In the second and third releases, the shooter killed four and 40 foxes respectively. Although the predation rate was reduced and survival increased, predation rate was still high, so there was no difference in survival between both the second and third release and the first one. In the fifth and sixth releases, the elimination of 120 foxes reduced fox predation rate to 28% and increased survival to 64–65%. Predation has led many reintroductions to fail (e.g.24,25), so effective fox management is critical to improve the survival of released western ringtail possums and to provide management agencies with the opportunity to pursue successful conservation projects. During the last release, providing fox awareness training to half of the cohort increased drastically the survival of trained individuals64, explaining the increased survival despite the absence of a professional shooter eliminating foxes. On the contrary of what is reported in literature, WBC did not directly affect survival in the possums released in this study24. In addition, our data did not demonstrate any effect on survival of the other variables evaluated in the study (different sources of food, releasing juveniles instead of adults, releasing animals in new and less smelly dreys, releasing animals in a predator-proof area). The presence of a predator as efficient as the fox might have limited the influence of other environmental variables on survival. However, during the releases and especially in the release in the predator-proof area, the deaths of some individuals was attributed to unknown causes that were unrelated to predation. Had they been in an area with foxes present, foxes might have scavenged on these animals resulting in ‘false positive’ DNA tests; in other words, our DNA data only shows that foxes interfered with the possums, but not that they actively killed all of them. At the same time, we are quite certain that many possums were killed by foxes due to observing them alive and well one day and finding them dead with clear signs of fox predation the next. It has been suggested that fox predation can have a compensatory effect where foxes might prey on animals that were going to die anyway for other causes65, therefore masking the impact of some variables.
In conclusion, we demonstrated that bold individuals had higher survival rate than shy individuals, and that females had higher survival rates than males. Our results strongly stressed the importance of an effective fox control strategy to improve the survival rate of the individuals. These findings will be useful in future reintroduction projects and in management strategies for the western ringtail possums and other species.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Bibliographys
Bradshaw, C. J. A. Little left to lose: deforestation and forest degradation in Australia since European colonization. J. Plant Ecol. 5, 109–120 (2012).
Commonwealth of Australia. Threatened species strategy. (2015).
Kearney, S. G. et al. The threats to Australia’s imperilled species and implications for a national conservation response. Pac. Conserv. Biol. 25, 231 (2019).
Woinarski, J. C. Z., Burbidge, A. & Harrison, P. L. The Action Plan for Australian Mammals 2012 (CSIRO Publishing, 2014).
IUCN. The IUCN Red List of Threatened Species. IUCN, Gland and Cambridge. https://www.iucnredlist.org/en (2022).
Guy, A. J., Curnoe, D. & Banks, P. B. A survey of current mammal rehabilitation and release practices. Biodivers. Conserv. 22, 825–837 (2013).
Bencini, R. & Chambers, B. Bunbury Outer Ring Road: Monitoring of Arboreal Mammals after Road Construction Interim Progress Report 2, 2013. https://doi.org/10.13140/2.1.2255.0085 (2013).
Jourdan, J. et al. Reintroduction of freshwater macroinvertebrates: challenges and opportunities. Biol. Rev. 94, 368–387 (2019).
Auster, R. E., Barr, S. W. & Brazier, R. E. Improving engagement in managing reintroduction conflicts: learning from beaver reintroduction. J. Environ. Plan. Manage. 64, 1713–1734 (2021).
Armstrong, D. P., Hayward, M. W., Moro, D. & Seddon, P. J. Advances in Reintroduction Biology of Australian and New Zealand Fauna (CSIRO Publishing, 2015).
Haage, M., Maran, T., Bergvall, U. A., Elmhagen, B. & Angerbjörn, A. The influence of spatiotemporal conditions and personality on survival in reintroductions–evolutionary implications. Oecologia 183, 45–56 (2017).
Santicchia, F. et al. Habitat-dependent effects of personality on survival and reproduction in red squirrels. Behav. Ecol. Sociobiol. 72, 134 (2018).
Moiron, M., Laskowski, K. L. & Niemelä, P. T. Individual differences in behaviour explain variation in survival: a meta-analysis. Ecol. Lett. 23, 399–408 (2020).
Mella, V. S. A., Gillies, C., McArthur, C., Webb, E. & Herbert, C. Factors influencing the success of hand-reared juvenile brushtail possums. Wildlife Res. 50, 96–107 (2022).
Wilson, D. S., Clark, A. B., Coleman, K. & Dearstyne, T. Shyness and boldness in humans and other animals. Trends Ecol. Evolut. 9, 442–446 (1994).
Koolhaas, J. M. et al. Coping styles in animals: current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 23, 925–935 (1999).
Kurvers, R. H. J. M. et al. Personality differences explain leadership in barnacle geese. Anim. Behav. 78, 447–453 (2009).
Thomson, J. S., Watts, P. C., Pottinger, T. G. & Sneddon, L. U. Physiological and genetic correlates of boldness: characterising the mechanisms of behavioural variation in rainbow trout Oncorhynchus mykiss. Hormones Behav. 59, 67–74 (2011).
Corsetti, S. et al. Bold personality makes domestic dogs entering a shelter less vulnerable to diseases. PLoS ONE 13, e0193794 (2018).
Sih, A., Bell, A. M., Johnson, J. C. & Ziemba, R. E. Behavioral syndromes: an integrative overview. Quart. Rev. Biol. 79, 241–277 (2004).
Moscicki, M. K. & Hurd, P. L. Sex, boldness and stress experience affect convict cichlid, Amatitlania nigrofasciata, open field behaviour. Anim. Behav. 107, 105–114 (2015).
Corsetti, S., Pimpolari, L. & Natoli, E. How different personalities affect the reaction to adoption of dogs adopted from a shelter. Animals 11, 1816 (2021).
Reeder, D. M. & Kramer, K. M. Stress in free-ranging mammals: integrating physiology, ecology and natural history. J. Mammal. 86, 225–235 (2005).
Clarke, J. Translocation Outcomes for the Western Ringtail Possum (Pseudocheirus occidentalis) in the Presence of the Common Brushtail Possum (Trichosurus vulpecula): Health, Survivorship and Habitat Use. PhD Thesis, Murdoch University (2011).
McCutcheon, H., Clarke, J., de Tores, P. & Warren, K. Health status and translocation success of wild and rehabilitated possums. in National Wildlife Rehabilitation Conference. Fremantle, Western Australia (2007).
Legge, S. et al. Havens for threatened Australian mammals: the contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators. Wildl. Res. 45, 627–644 (2018).
Wilson, B. A. et al. Adapting reintroduction tactics in successive trials increases the likelihood of establishment for an endangered carnivore in a fenced sanctuary. PLoS ONE 15, e0234455 (2020).
Evans, M. J. et al. The ‘Goldilocks Zone’ of predation: the level of fox control needed to select predator resistance in a reintroduced mammal in Australia. Biodivers. Conserv. 30, 1731–1752 (2021).
Britt, A., Welch, C. & Katz, A. Can small, isolated primate populations be effectively reinforced through the release of individuals from a captive population?. Biol. Conserv. 115, 319–327 (2004).
Brightsmith, D. et al. The use of hand-raised psittacines for reintroduction: a case study of scarlet macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 121, 465–472 (2005).
Kohl, K. D. & Dearing, M. D. Experience matters: prior exposure to plant toxins enhances diversity of gut microbes in herbivores. Ecol. Lett. 15, 1008–1015 (2012).
Tarszisz, E., Dickman, C. R. & Munn, A. J. Physiology in conservation translocations. Conserv. Physiol. 2, 054 (2014).
Ostermann, S. D., Deforge, J. R. & Edge, W. D. Captive breeding and reintroduction evaluation criteria: a case study of peninsular bighorn sheep. Conserv. Biol. 15, 749–760 (2001).
Wayne, A. F. et al. Ngwayir (Pseudocheirus Occidentalis) Declines in the Upper Warren, the Issue in Brief. (2012).
Yokochi, K. A major road and an artificial waterway are barriers to the rapidly declining western ringtail possum, Pseudocheirus occidentalis. PhD Thesis, The University of Western Australia (2015).
Yokochi, K., Chambers, B. K. & Bencini, R. An artificial waterway and road restrict movements and alter home ranges of endangered arboreal marsupial. J. Mammal. 96, 1284–1294 (2015).
Corsetti, S., Yokochi, K., Webb, E., Urso, A. & Bencini, R. Practical, effective, and safer: placing traps above ground is an improved capture method for a critically endangered arboreal folivore, the ngwayir (Pseudocheirus occidentalis). Animal Welfare, accepted (2024).
Berry, O. & Sarre, S. D. Gel-free species identification using melt-curve analysis. Mol. Ecol. Notes 7, 1–4 (2007).
Wat, K. K. Y., Herath, A. P. H. M., Rus, A. I., Banks, P. B. & Mcarthur, C. Space use by animals on the urban fringe: interactive effects of sex and personality. Behav. Ecol. 31, 330–339 (2020).
Giffney, R. A. et al. Age of road-killed common brushtail possums (Trichosurus vulpecula) and common ringtail possums (Pseudocheirus peregrinus) in an urban environment. Aust. Mammalogy 31, 137–142 (2009).
Department of Biodiversity, Conservation and Attractions. Standard Operating Procedure Cage Traps for Live Capture of Terrestrial Vertebrates. Department of Biodiversity, Conservation and Attractions. https://www.dbca.wa.gov.au/science/animal-ethics-and-use-animals (2023).
Luna, D. et al. Do domestic pigs acquire a positive perception of humans through observational social learning?. Animals 11, 127 (2021).
Schuler, B. et al. Optimal hematocrit for maximal exercise performance in acute and chronic erythropoietin-treated mice. Proc. Natl. Acad. Sci. 107, 419–423 (2010).
Bury, A., Niedojadlo, J., Sadowska, E. T., Bauchinger, U. & Cichoń, M. Contrasting response of haematological variables between long-term training and short exercise bouts in zebra finches (Taeniopygia guttata). J. Exp. Biol. 222, jeb193227 (2019).
Berda-Haddad, Y. et al. Increased mean corpuscular haemoglobin concentration: artefact or pathological condition?. Int. J. Lab. Hematol. 39, 32–41 (2017).
Sgoifo, A., Carnevali, L. & Grippo, A. J. The socially stressed heart Insights from studies in rodents. Neurosci. Biobehav. Rev. 39, 51–60 (2014).
Carnevali, L. et al. Vagal withdrawal and susceptibility to cardiac arrhythmias in rats with high trait aggressiveness. PLoS ONE 8, e68316 (2013).
Fleshner, M., Laudenslager, M. L., Simons, L. & Maier, S. F. Reduced serum antibodies associated with social defeat in rats. Physiol. Behav. 45, 1183–1187 (1989).
Capitanio, J. P., Mendoza, S. P. & Baroncelli, S. The relationship of personality dimensions in adult male rhesus macaques to progression of simian immunodeficiency virus disease. Brain Behav. Immun. 13, 138–154 (1999).
Sgoifo, A., Koolhaas, J., Alleva, E., Musso, E. & Parmigiani, S. Social stress. Acute and long-term effects on physiology and behavior. Physiol. Behav. 73, 253–254 (2001).
Nola, M. & Dotlić, S. The Hematopoietic and Lymphoid Systems. in Pathology Secrets (Third Edition) (ed. Damjanov, I.) 161–202 (Mosby, Philadelphia, 2009). https://doi.org/10.1016/B978-0-323-05594-9.00009-X.
Clarke, J. et al. Hematologic and serum biochemical reference ranges and an assessment of exposure to infectious diseases prior to translocation of the threatened western ringtail possum (Pseudocheirus occidentalis). J. Wildl. Dis. 49, 831–840 (2013).
Amin, B., Verbeek, L., Haigh, A., Griffin, L. L. & Ciuti, S. Risk-taking neonates do not pay a survival cost in a free-ranging large mammal, the fallow deer (Dama dama). R. Soc. open sci. 9, 220578 (2022).
Herath, A. P. H. M., Wat, K. K. Y., Banks, P. B. & McArthur, C. Animal personality drives individual dietary specialisation across multiple dimensions in a mammalian herbivore. Function. Ecol. 35, 2253–2265 (2021).
Luna, Á., Palma, A., Sanz-Aguilar, A., Tella, J. L. & Carrete, M. Personality-dependent breeding dispersal in rural but not urban burrowing owls. Sci. Rep. 9, 2886 (2019).
Wauters, L. A. et al. Personality traits, sex and food abundance shape space use in an arboreal mammal. Oecologia 196, 65–76 (2021).
Bonte, D. et al. Costs of dispersal. Biol. Rev. 87, 290–312 (2012).
Wilson, D. S., Coleman, K., Clark, A. B. & Biederman, L. Shy-bold continuum in pumpkinseed sunfish (Lepomis gibbosus): an ecological study of a psychological trait. J. Comparative Psychol. 107, 250–260 (1993).
Moehrenschlager, A. & Macdonald, D. W. Movement and survival parameters of translocated and resident swift foxes Vulpes velox. Anim. Conserv. 6, 199–206 (2003).
Maran, T., Põdra, M., Põlma, M. & Macdonald, D. W. The survival of captive-born animals in restoration programmes – Case study of the endangered European mink Mustela lutreola. Biol. Conserv. 142, 1685–1692 (2009).
Miller, L. J., Vicino, G. A., Sheftel, J. & Lauderdale, L. K. Behavioral diversity as a potential indicator of positive animal welfare. Animals 10, 1211 (2020).
Dochtermann, N. A., Schwab, T. & Sih, A. The contribution of additive genetic variation to personality variation: heritability of personality. Proc. R. Soc. B. 282, 20142201 (2015).
Leiner, N. O. & Silva, W. R. Territoriality in females of the slender opossum (Marmosops paulensis) in the Atlantic forest of Brazil. J. Tropic. Ecol. 25, 671–675 (2009).
Corsetti, S., Chambers, B., Blache, D. & Bencini, R. Learning as a key to cope with introduced predators: training the western ringtail possum to be scared of foxes. Communications Biology (Under review).
Norrdahl, K. & Korpimäki, E. Effects of predator removal on vertebrate prey populations: birds of prey and small mammals. Oecologia 103, 241–248 (1995).
Acknowledgements
We wish to thank the South West NRM, formerly South West Catchments Council (SWCC) for providing funds for the project. A special thank goes to the owners of the private properties that allowed us to release the animals: Gail Taylor, Alan Taylor and Kerry Warburton. We also wish to thank the arborists from Naturaliste Tree Care who climbed the trees to release the possums. We are grateful to Jaya Vaughan, Jane Putland, Georgia Driver, Alyse Archer and Daniel Lupardo for their help during radio tracking. We also want to thank the FAWNA association for providing the animals for the study, and for their help in the early stages of the project. Sara Corsetti was supported by an Australian Government Research Training Program (RTP) Scholarship and the University Postgraduate Award for International Students (UPAIS) Scholarship. She was also recipient of Holsworth Wildlife Research Endowment awards from the Ecological Society of Australia.
Author information
Authors and Affiliations
Contributions
Study conception and design: RB, BC, SC; data collection: SC, RB; analysis and interpretation of results: SC, RB, DB; writing – original draft preparation: SC; writing – review and editing: SC, RB, BC, DB; funding acquisition: RB, BC; supervision: RB. All authors reviewed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Corsetti, S., Chambers, B., Blache, D. et al. Captive-raised western ringtail possum (Pseudocheirus occidentalis) with a bold personality have higher survival rates when released into the wild. Sci Rep 14, 20110 (2024). https://doi.org/10.1038/s41598-024-67726-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-67726-y
- Springer Nature Limited