
Abstract
Cambodia’s goal to eliminate malaria by 2025 is challenged by persistent transmission in forest and forest fringe areas, where people are exposed to Anopheles mosquito bites during the day and night. Volatile pyrethroid spatial repellents (VPSRs) and insecticide-treated clothing (ITC) could address these gaps. This study evaluated the outdoor application of one passive transfluthrin-based VPSR, four etofenprox-ITCs paired with a picaridin topical repellent, and a combination of VPSR and ITC against wild Anopheles landing in Cambodia. A 7 × 7 Latin-square study was conducted over 49 collection nights in temporary open structures in Mondulkiri Province. All interventions substantially reduced Anopheles landing, with protective efficacy ranging from 61 to 95%. Mathematical modeling showed significant reductions in vectoral capacity, especially with the combined ITC and VPSR and VPSR alone, albeit with decreased effectiveness over time. These interventions have the potential to reduce outdoor and daytime Anopheles biting, offering valuable contributions to malaria elimination efforts in Cambodia and the Greater Mekong Subregion, contingent upon achieving effective coverage and adherence.
Similar content being viewed by others


Introduction
The Greater Mekong Subregion (GMS) aims to achieve elimination of Plasmodium falciparum malaria by 2025, and of all human malaria by 20301,2. From 2000 to 2020, the GMS recorded a 56% decrease in malaria, and an 89% reduction in P. falciparum cases. As of 2022, Cambodia accounts for 2% of all malaria cases and for 1% of P. falciparum cases in the GMS. Operating on an accelerated timeline compared to the remaining GMS countries, Cambodia strives to eliminate P. falciparum malaria by 2024, and all human malaria by 20251.
From 2010 to 2022, confirmed malaria cases in Cambodia declined by 91%. As malaria decreases throughout the country, it has become increasingly confined to malaria transmission foci2. Roughly 0.5 million people in Cambodia live in forest and forest fringe areas characterized by high malaria transmission3,4. These forest transmission foci are predominantly inhabited by ethnic minorities, local populations, and rural mobile and migrant populations working in rubber plantations, mining, and agriculture5,6,7. Under the Malaria Elimination Action Framework (2021–2025), the Cambodia’s National Center for Parasitology, Entomology, and Malaria Control deploys forest packs, often containing insecticide-treated nets (ITNs), insecticide treated hammock nets, and topical repellents to mobile and migrant populations in areas at highest malaria risk2,8,9. Since November 2020, Cambodia has also adopted foci-based innovative strategies as part of its “last mile” strategy towards P. falciparum malaria elimination, including targeted drug administration and intermittent preventive treatment for forest-goers9,10.
Appropriate mosquito bite prevention interventions to reduce forest-going and -dwelling populations’ exposure to forest-based Anopheles are needed based on the spaces and times where, and when these individuals are exposed to Anopheles4. A recent study conducted in northern Cambodia found that while Anopheles densities strongly declined in villages during the dry season, densities remained relatively similar from the dry to the wet season in the forest7. Therefore, forest sites may serve as suitable refuges for Anopheles during the dry season, and consequently, also as a malaria parasite reservoir, since human activity in the forest is particularly extensive during the dry season7. In addition, forest-goers are exposed to vector bites during the day due to Anopheles exhibiting outdoor, daytime and early evening biting11; and during the night, due to low bed net use and open sleeping structures7,12. These characteristics limit the effectiveness of strategies that focus on traditional village- and homestead-centric vector control interventions (indoor residual spraying (IRS), ITNs)6,13.
Additional interventions that target mosquitoes and people outdoors must be available for use alongside IRS and ITNs. This study focuses on evaluating two novel, promising tools: volatile pyrethroid spatial repellents (VPSRs), and insecticide treated clothing (ITC). VPSRs function by preventing human-vector contact primarily through non-contact irritancy, non-contact excito-repellency (the combined effects of both irritation resulting from coming into direct contact with a treated area and the tendency to avoid treated areas due to their repellent properties), spatial repellency, landing inhibition, feeding inhibition, sublethal incapacitation, and pre-/post-prandial (before/after blood feeding) mortality14. ITCs primarily protect humans from mosquito bites through contact irritancy, contact excito-repellency, some short-range non-contact excito-repellency, feeding inhibition, and mortality15,16,17. Synthetic topical repellents like picaridin (and DEET) offer personal defense against mosquito bites through short-range mechanisms such as blocking olfactory attractants18,19, inducing non-contact irritancy, and/or direct contact contact irritancy20,21. Pairing insecticide-treated clothing (ITCs) with a topical repellent may amplify bite prevention22.
VPSRs have been extensively evaluated in Africa23,24,25,26,27,28,29 and increasingly in Southeast Asia30,31,32,33. ITCs, treated with pyrethroids, have been widely evaluated and show promise for their use against Anopheles biting amongst mobile populations and military/ranger personnel34,35,36,37. However, commonly used permethrin formulations have shown poor wash retention and low bite prevention levels on many lighter weight fabrics prompting the development of etofenprox clothing treatment products with superior wash retention and safety profiles38,39. The evidence that permethrin treated clothing provides protection against malaria is unclear16,17,40,41,42 and the WHO currently does not recommend its use for the prevention and reduction of malaria at the community level where malaria transmission is ongoing43.
The study presented here is a component of a multi-staged research program called ‘Project BITE’ (Bite Interruption towards Elimination) (2020–2023)). Project BITE applied a mixed-methods, phased approach to vector control product evaluation44. Prior to the field intervention evaluation, a series of preceding semi-field system (SFS) experiments were conducted in Thailand to measure the primary effects (landing prevention) and the secondary effects (sublethal incapacitation and mortality) of new transfluthrin- and metofluthrin-based VPSRs, and etofenprox-treated clothing, against pyrethroid-susceptible Anopheles minimus (an important vector of malaria in the GMS45) (manuscript under review). In addition to providing insights and estimates of intervention impact on landing inhibition, sublethal incapacitation, and mortality, the data were used to parameterize a mathematical model to predict the potential of the tools to provide community protection46. Vectoral capacity is a measure of the ability of the vector population to transmit a disease, defined as the total number of potentially infectious bites that would eventually arise from all the mosquitoes biting a single infectious human on a single day47. Interventions that reduce vectoral capacity have the potential to confer community-wide protection leading to reduced malaria burden48.
Building on the SFS studies, an entomological field intervention evaluation was undertaken to evaluate these VPSR and ITC interventions on wild Anopheles landing using human landing catches (HLCs) in a small-scale, controlled field setting in Cambodia. The study’s secondary objectives were to 1) validate the results from the preceding SFS evaluation by measuring intervention effectiveness against local, wild Anopheles landing, 2) measure the hourly human landing rates (HLR) in the control structures, towards describing the local Anopheles’ evening and nighttime biting trends, 3) confirm the Anopheles species identification from a subset of the HLC-collected specimens, and to 4) predict the reduction of vectoral capacity of these interventions using mathematical modelling. The potential of these tools to provide protection beyond personal protection is investigated by combining data from both the field study in Cambodia and the preceding semi-field studies in Thailand.
Methods
Study site
Field collections took place in the village of Andong Krolong (12.320725, 107.029779), Mondulkiri Province, Cambodia. The study took place from 23 September to 24 November 2022, thus taking place during the rainy season and the early days of the dry season. Temporary, open sleeping structures representative of open sleeping structures used by locals were constructed out of wooden poles and tarpaulin roofs (2 × 2 × 2 m) (Fig. 1). While closely surrounded by forest cover, the open structures were in an area that had been recently cleared, representative of the living conditions of forest workers (Fig. 1).

Temporary, open structures set-up in the field site in Andong Krolong, Mondulkiri Province, Cambodia (left), and front view of the temporary, open structures (right).
Mosquito bite prevention interventions
Six bite prevention interventions were evaluated (Table 1). Interventions included a transfluthrin-based VPSR (BiteBarrier (‘BB’), formerly known as PIRK, PIC Corps), civilian and ranger clothing that were either newly treated with etofenprox or treated with etofenprox and then washed 20 times, as well as one arm with both the VPSR and etofenprox-treated civilian clothing. The treated civilian and ranger clothing was always paired with a 20% picaridin topical repellent (SC Johnson). The picaridin repellent was selected for this study as it has previously been demonstrated to be safe and effective against Southeast Asian vectors of malaria11.
Experimental design and mosquito collections
Seven temporary, open structures located at least 20 m apart were set up. HLCs were carried out in the structures for 12 h, from 18h00 to 06h00, divided into two collection shifts, 18h00–00h00 (shift 1) and 00h00–06h00 (shift 2), where each shift was covered by a single collector per structure (for example, in structure 1, collector 1 worked shift 1, and collector 2 worked shift 2). A fully balanced 7 × 7 Latin-square design was used. Each of the seven study arms (six interventions and one control) was assigned to one structure for seven collection nights, and each pair of collectors rotated through each location on a nightly basis. After each block of seven collection nights, interventions were advanced to the next position, and collectors continued to rotate through structures each night. After 49 nights of collection each collector had tested each intervention in each location seven times. There were 20 unique HLC collector pairs. Some collector pairs worked fewer HLC nights than others as some individual collectors left the study before its completion. Due to cultural perceptions about being in the forest at night, collector pairs remained together in the structure throughout the entire collection night: while one collector worked, the other collector slept underneath the structure, in an untreated hammock net.
HLCs were used to collect mosquitoes landing on the area from knee to ankle of the collector for each collection rotation. For etofenprox-treated clothing interventions, long trousers were not rolled up to the knee in order to estimate the landing protection afforded by the combination of etofenprox with long trousers. The negative control had the area between the knee and the ankle exposed. The total number of mosquitoes caught hourly was recorded. Mosquitoes captured were stored in individually labelled Eppendorf tubes (by treatment and hour of collection), transported in coolers to the base camp every morning, killed by freezing, counted, morphologically sorted, and stored individually with desiccant in Eppendorf tubes for subsequent processing.
Ethical statement
All methods were carried out in accordance with Cambodia’s National Center for Parasitology, Entomology, and Malaria Control guidelines; all HLC collectors were over 18 years of age, and provided written informed consent and were medically supervised, i.e., provided free-of-cost malaria diagnosis and treatment should malaria symptoms (e.g., fever) occur during the study and/or during the two weeks subsequent to the field trial period. Ethics approval for this study protocol was obtained from the Cambodia Ministry of Health’s National Ethics Committee for Health Research (nº296).
Environmental data collection
Temperature (Celsius), % relative humidity, windspeed (m/s) and rainfall occurrence were recorded on an hourly basis during HLC collections, from 18h00 to 06h00. Temperature, % relative humidity, and wind speed data were recorded at the end of each HLC collection hour using a data logger device (HOBO®). Rainfall occurrence was recorded as a binary variable (yes/no) to indicate occurrence or absence of rainfall during each HLC collection hour.
Mosquito morphological and molecular species identification
All Anopheles mosquitoes were sorted to species or species group using morphological identification keys52 in the field. Individual specimens were then packaged in individual, tightly closed, Eppendorf tubes with silica gel and were sent to University of Notre Dame, Indiana, USA, for molecular species confirmation. Approximately 15% of samples were randomly selected across all HLC collections, and sequenced at the ribosomal DNA internal transcribed spacer region 2 (ITS2) and/or cytochrome oxidase subunit 1 (CO1) loci towards species determination12. Conservative molecular species identification was based on matches to GenBank (National Center for Biotechnology Information [NCBI]) and BOLD53 (databases with lower quality matches and an absence of voucher specimens resulted in identifications to higher taxonomic levels).
Data analysis
The change in landing associated with each intervention was estimated as rate ratios (RR), the ratio of the number of Anopheles landing in the intervention compared to that in the control. They were estimated using a mixed-effect negative binomial regression with structure ID, collector pair, and intervention ID as fixed-effects, and with collection date and batch (the location-night) as random effects. The effect of structure location and collector pair on Anopheles catches per collection night was also examined. All RR estimates are presented with 95% confidence intervals (CI). The percentage protective efficacies of each intervention against Anopheles landing were estimated as \(\left(1-RR\right) \times 100\).
The relationship between weather variables and Anopheles densities caught was investigated. Mean nightly temperatures, mean nightly %RH, mean nightly wind speed, and number of rain occurrences per night were plotted against the total number of Anopheles captured per night. All statistical analyses were performed in R (Murray Hill, New Jersey), Version 2023.03.0 + 38654, using the tidyverse packages ‘tidyr’55, ‘dplyr’56, ‘lme4’57, and ‘ggplot2’58.
The hourly human landing rate (HLR) for the control arm across the 49 collection nights was estimated as a proxy for the hourly human biting rate22,59.
Modeling of vectoral capacity
The relative reduction in vectoral capacity was predicted using the method described in Denz et al. (2021)48 and in Fairbanks et al.46. Firstly, the vectoral capacity is calculated for a “baseline scenario” (without the intervention). Parameters used in the calculation for this are given in Suppl Table 1. Biometric parameters for Anopheles dirus were used since this species accounted for most of the local mosquito population. Next, were considered other scenarios where a proportion of the population is protected by an intervention. This proportion is dependent on both the coverage (whether someone has the intervention) and adherence (whether someone uses the intervention).
Fairbanks et al.46 used data from semi-field studies combined with a model to quantify the modes of action of selected vector control tools. The influence of the interventions on the mosquito feeding cycle were described relative to an unprotected human host using four characteristics; the relative reduction in the rate of landing, the relative increase in the rate of preprandial killing and disarming, the proportion of this increase in rate which is due to disarming, and the change in the probability of postprandial mortality. Estimates from the field study are used to update some of these parameters to provide estimates for the wild Anopheles at this field site. For some interventions there are multiple semi-field estimates (from different sites and years), in this case we consider estimations from each semi-field scenario separately, and therefore have multiple estimates for the reduction in vectoral capacity for these interventions.
The relative reduction in the rate of biting parameters estimated from the field study were used. Then, to consider the increase in the rate of preprandial killing and disarming, the relative change in the rate of biting in the field studies compared to semi-field studies is first calculated:
where \({\uppi }_{\text{F}}\) and \({\uppi }_{\text{SF}}\) are the median estimated protective efficacies for the change in the rate of biting in in the field and semi-field studies, respectively. This ratio helps to understand how the reduction in biting in the field studies compares to that in the semi-field studies. The reduction in the rate of disarming and preprandial killing is then calculated as:
where \({\upkappa }_{\text{SF}}\) is the estimated rate from the semi-field data. This scaling method ensures that the proportion of the reduction in biting attributed to disarming or preprandial killing matches that observed in the semi-field studies, while the magnitude of these effects are assumed to scale according to observation under field conditions (with wild Anopheles). The probability a mosquito is killed postprandially and the probability a mosquito is disarmed, given it is disarmed or killed preprandially, are assumed to be the same as in the semi-field conditions.
To incorporate variability and uncertainty, parameter values are sampled from the protective efficacy distribution (reduction in biting) or the from the semi-field study (all other parameters).
Results
Anopheles species identification and human landing rates
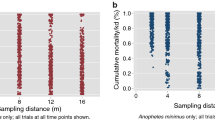
A total of 8,294 Anopheles specimens were collected with HLCs. Of this total, 96% (n = 7951) of specimens were morphologically identified to species or species group. Out of the total morphologically identified, 96% (n = 7621) were identified as An. dirus sl, leaving the remaining 5% of specimens identified to eight different species or species group: Anopheles maculatus sl (n = 234), An. minimus (n = 63), Anopheles philippinensis (n = 10), Anopheles kochi (n = 8), Anopheles aconitus (n = 7), Anopheles barbirostris sl (n = 5), Anopheles baimaii (n = 2), and Anopheles asiaticus (n = 1). Molecular speciation of 15% (n = 1242) specimens were evaluated, and confirmed that the morphological identification was extremely accurate (Table 2).
The estimated mean HLR per hour (control arms) ranged from 8.16 (95%CI 0.21–16.10) Anopheles landings per person per hour (lph) (00h00–01h00) to 2.61 (CI 0.00–5.22) lph (05h00–06h00). Higher landing rates were recorded from 18h00 to 00h00. After 02h00, a decline of the mean hourly HLR was observed until the last HLC collection hour (05h00–06h00) (Fig. 2).

Hourly Anopheles human landing rate (HLR) (Anopheles landings per person, per hour), over the course of 49 collection nights (from control arms).
Intervention protective efficacy against Anopheles landing
All six mosquito bite prevention interventions showed substantial reductions in mosquito landings relative to the control (Ctl) (Fig. 3). The estimated protective efficacies of the interventions against Anopheles landing ranged from 61% (95%CI 48–71%) to 95% (CI 93–96%) (Fig. 4, Table 3). BB alone and the combined interventions (BB + EtoC- 0-Pi) provided the greatest protection against Anopheles landing, with protective efficacies of 94% (CI 91–95%) and 95% (CI 93–96%), respectively. The etofenprox-treated ranger uniform and civilian clothing, unwashed, provided similar protection against landing, 73% (CI 64–80%) and 76% (CI 67–82%), respectively. Both etofenprox-treated ranger uniform and civilian clothing continued to provide substantial protection after 20 washes, 65% (CI 53–74%) and 61% (CI 48–71%), respectively (Fig. 4, Table 3).

Number of Anopheles captured per HLC night by intervention.

Estimated risk ratios (95% CI) for the effect of each intervention on risk of Anopheles landing (plotted on log scale).
Overall, there was variation in the number of Anopheles captured per night by both the structure locations (Suppl Fig. 1), and collector pair (Suppl Fig. 2), justifying the inclusion of location and volunteer pair as covariates in the statistical model. Environmental parameters (temperature, %RH, wind speed and rainfall) were examined against total Anopheles captures per night. Temperatures remained stable, between 22 and 24 °C, and wind speed did not exceed 1.2 m/s; there was no clear relationship between temperatures or wind speed with Anopheles captures. Humidity was highest (100%) during the period of heaviest rainfalls (October), which coincided with lower Anopheles catches.
Relative reduction in vectoral capacity
This parameter was measured by combining field data and the semi-field data generated during the preceding SFS trials in Thailand to predict the potential of the tools to provide community protection.
The relative reduction in vectoral capacity for each intervention coverage is shown in Fig. 5. Overall, all interventions were predicted to have a large impact on the vectoral capacity. The impact increases with coverage and adherence. In line with the reduction in landing observed in the field study, the greatest reduction of vectoral capacity was predicted for the combined EtoC and BB VPSR as well as the BB VPSR used alone. Even at 50% usage large reductions in vectoral capacity are predicted. As interventions age, and consequently dosage of available insecticide deceases, they are less effective at reducing the vectoral capacity (Fig. 5).

Predicted relative reduction in vectoral capacity for Anopheles dirus due to each intervention for a range of coverage levels for each intervention trailed in the field experiments. For interventions with semi-field estimates from multiple sites and years an individual line is plotted for each semi-field scenario. Black lines represent the median estimates, and colored bands represent the 95% CIs.
Although predictions have similar trends when utilizing data from different semi-field experiments, the magnitude of the predicted reduction in vectoral capacity is variable. There were differences in the extent of the modes of action observed, and therefore the parameter estimates, between experiments. These could be due to a number of factors including meteorological differences between locations and times and genetic differences between mosquito colonies.
Discussion
In this field evaluation, the authors measured the impact of outdoor use of a passive, transfluthrin-based spatial repellent and etofenprox-treated clothing (paired with a 20% picaridin topical repellent) in the form of six bite prevention interventions on wild Anopheles landing. The study was designed to estimate landing rates (as a proxy for biting rates) when using the interventions while sitting in a fixed position in open structures similar to the open shelters used by forest-exposed populations in Cambodia.
A wide diversity of Anopheles species was observed in the study site in Mondulkiri (Table 2). Of the 15 Anopheles species collected, five are known vectors of human malaria: An. dirus Form A12,61,62, An. maculatus63,64, An. minimus64,65, An. sawadwongporni64, and An. aconitus7,66. Species An. dirus, An. maculatus, and An. minimus comprise Cambodia’s major malaria vectors5,67,68, and all 15 collected species have been previously identified in Cambodia7,12,64. While HLC collections occurred from early evening (18h00) to early morning (06h00), it is possible that daytime biting would also be observed had collections taken place prior to 18h00. Two other entomological studies in Mondulkiri province, one being in a forested site4, and the other being in both forested and village sites7, included 24-h Anopheles collections demonstrating daytime biting. Daytime biting suggests a further need for additional bite prevention interventions that are protective even when users are awake and outdoors4,7.
This field study found that all of the study interventions, even when washed (for the treated clothing interventions), provided substantial protection against wild Anopheles landing outdoors in open temporary structures (Table 3, Figs. 3, 4). The BB was highly effective against Anopheles landing, preventing 94% (CI 91–95%) of landings (Table 2). Combining this passive VPSR with etofenprox-treated clothing made no substantial difference on the level of protection provided against Anopheles landing. However, the combined intervention—delivered as a forest pack—is intended to protect real-world users both while inside their homes or temporary shelters and also while mobile (outside their homes/shelters), offering “full-time” or 24-h protection. In other words, the combined interventions provide more complete protection as needed based on individuals’ daily activities. However, for this study, the HLC collectors were restricted to the temporary structures.
Etofenprox-treated clothing interventions also provided high levels of protection against mosquito landing, though with slightly lower estimated protective efficacies than the BB intervention. However, while landing rates are often used as a proxy for measuring biting rates22, evidence indicates that landing might not always necessarily lead to biting. When mosquitoes are exposed to pyrethroids (airborne or applied to clothing), they may still be able to detect the host and land, but they are inhibited from biting69. In a lab study, the landing and biting rates of Aedes aegypti were measured while using metofluthrin VPSRs, and observed 74 landings for only eight bites70. Therefore, it cannot be excluded that measuring landing rates may lead to an overestimation of ‘biting’ and an under-estimation of protection. To more accurately estimate the protective efficacy of ITCs, it might be best to allow mosquitoes to feed as a more definitive endpoint of protective efficacy71.
This study only evaluated etofenprox-treated clothing paired with a topical repellent, as the preceding SFS trials demonstrated that the combined ITC and topical repellent intervention was more effective against mosquito landing than the treated clothing alone (Vajda et al.37, unpublished 2020/2021 data (manuscript under review)). In another field study evaluating personal repellent and treated clothing interventions in Lao PDR, permethrin-treated short clothing paired with a topical repellent were found to provide substantially more protection against mosquito landing than the permethrin-treated short clothing alone22. However, the permethrin-treated long clothing provided much higher protection against biting than the treated short clothing, but treated long clothing with a topical repellent was not tested22. Given the challenges with adherence around topical repellents use17,20,72, it would be useful to compare treated long clothing alone, to treated long clothing with a topical repellent. This would help better understand how much more protection against biting is conferred by the addition of a topical repellent.
On the other hand, this study included a negative control with short trousers, but not one with long trousers (due to resource and time constraints). However, because the etofenprox-treated clothing interventions comprised long trousers that were not rolled up to the knees during HLCs, it is possible that the ratios of Anopheles landing in the treated clothing interventions compared to the Anopheles landing in the control slightly overestimate the landing protective efficacy since the negative control did not have the added physical barrier provided by ankle-length trousers73.
Washing the etofenprox-treated clothing interventions 20 times only slightly increased the risk of Anopheles landing. This finding has implications for determining the frequency of retreatment of etofenprox-treated clothing. Efficacy testing of ITCs in controlled field and/or laboratory settings should be regarded as a proxy for real-world field conditions, in which ITCs would face harsher and more variable conditions (e.g., intense sweating during physical labor, textile degradation). Therefore, care should be taken when interpreting controlled testing results, and where possible, test results should be considered alongside results from efficacy testing in real-world field settings as is conducted for other vector control tools such as ITNs74.
Permethrin is the most common insecticide used to treat clothing75. Given the shortcomings of permethrin regarding longevity, efficacy on lightweight fabrics, and higher human toxicity (g/kg), etofenprox formulations for clothing treatment have been developed and registered by the U.S. Environmental Protection Agency76. Etofenprox is a synthetic pyrethroid-like ether insecticide. The structure of etofenprox renders it more stable and with lower mammalian toxicity than other pyrethroid insecticides and functions by attacking the neuronal axon of the mosquito77. As concerns over the spread of pyrethroid resistance grows, and several studies of the impact of pyrethroid resistance on the protective effect treated clothing yield conflicting results38,39,78,79 there is growing need to explore alternative insecticides for clothing treatment.
The set replacement interval of the BB device in the field must also be based on its residual efficacy. This study only tested new, freshly manufactured BB products towards providing a baseline understanding of the product’s efficacy against local, wild Anopheles, and did not investigate its residual efficacy. Therefore, the BB’s residual efficacy is currently unknown. Similarly to ITCs, transfluthrin-based VPSR efficacy over time is dependent on exposure to UV and other environmental parameters (rainfall, wind), as well as the susceptibility of local mosquitoes to pyrethroids23,24,59,59,70,80. For this field study, insecticide susceptibility testing to pyrethroids, including etofenprox and transfluthrin, was not feasible due to issues with keeping field-caught adult Anopheles alive and other logistical challenges.
This intervention evaluation provides evidence on the efficacy of these bite prevention interventions under controlled field conditions against wild Anopheles landing in Mondulkiri Province, Cambodia and highlights their potential for use in this elimination scenario if effective coverage and adherence of at-risk populations can be achieved. As with all vector control interventions, effectiveness is dependent on local vector bionomics81, vector insecticide susceptibility profiles and resistance mechanisms82,83,84, which uniquely affect intervention functionality19,85,86,87,88,89,90,91. Ecological, and human behavioral factors are also essential components that impact intervention effectiveness51,92. Therefore, malaria control and elimination programs must tailor their strategies and mix of interventions based on generated country-specific evidence and unique circumstances.
To date, the WHO has not established a position statement regarding the applications of VPSRs, ITCs, and topical repellents in public health vector control. However, while refraining from establishing formal recommendations on ITCs and topical repellents applications, the WHO does suggest these interventions for personal protection and considers their use for high-risk groups who do not benefit from other vector control interventions43,93. Recent work on a stochastic transmission model based on time-stratified vector landing data from controlled experiments of transfluthrin-treated eave ribbons (a type of VPSR) found that in addition to Anopheles vector landing reduction (personal protection), transfluthrin-treated ribbons also killed and reduced blood feeding, causing important reductions in vectoral capacity, indicating its potential to offer community protection48. The modeling evaluation conducted for this field study’s interventions’ on vectoral capacity also indicates reductions in vectoral capacity, even at decreased intervention coverage and adherence (Fig. 5). Limitations for the vectoral capacity model are described in detail in the original publication94. Here, parameters describing the bionomics of vectors in the model simulations were derived from averages for An. dirus from the GMS and surrounding areas95. However, these parameters, as well as different species of malaria vectors, can vary across different settings, potentially affecting the generalizability of our results. This model also assumed uniform changes in biting reduction and other endpoints between the SFS and field environments, based on data collected specifically on biting reduction. However, it is possible that other endpoints, such as mortality rates or incapacitation, may exhibit different patterns of change in field conditions compared to those observed in SFS. Still, this modeling assessment corroborates findings from recent SFS experiments23,24,25,26,27,29 and field studies of VPSRs30,96,97. Given this growing body of evidence, insecticidal personal protection interventions are increasingly recognized as having high potential for public health use, but further evidence of epidemiological impact is needed for WHO to establish a policy recommendation for these interventions98.
Conclusion
In Southeast Asia, Anopheles bites occurring outside the protection of the traditional, homestead-centric interventions (IRS, LLINs), constitute important gaps in protection that call for novel bite prevention interventions. This field study is highly encouraging as it demonstrates that this transfluthrin-based BB VPSR and etofenprox ITCs provide substantial protection against Anopheles landing when used outside. Further, the study’s modeling analysis predicted that these interventions would reduce vectoral capacity, even at 50% coverage. Additional studies in other geographic settings are needed to estimate intervention impact on landing of mosquitoes with different bionomics profiles, and effectiveness trials in which the products are distributed and used normally are also required. While further evidence of epidemiological impact is necessary for WHO to establish these tools as effective public health interventions against malaria, this study provides promising results for bite prevention tools that can be used toward addressing gaps in protection of at-risk groups against Anopheles biting. In fact, recently, the U.S. President's Malaria Initiative (PMI) updated its technical guidance, enabling program evaluation of new tools, including spatial repellents, influenced in part by the positive outcomes from Project BITE and other recent and ongoing studies, reflecting an adaptive donor environment.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
World Health Organization. World malaria report 2023 (World Health Organization, 2023).
Sovannaroth, S. et al. Accelerating malaria elimination in Cambodia: An intensified approach for targeting at-risk populations. Malar J. 21, 209 (2022).
Durnez, L. et al. Outdoor malaria transmission in forested villages of Cambodia. Malar J. 12, 329 (2013).
Kunkel, A. et al. Choosing interventions to eliminate forest malaria: Preliminary results of two operational research studies inside Cambodian forests. Malar J. 20, 51 (2021).
Cui, L. et al. Malaria in the greater Mekong Subregion: Heterogeneity and complexity. Acta Tropica. 121, 227–239 (2012).
Canavati, S. E. et al. Targeting high risk forest goers for malaria elimination: A novel approach for investigating forest malaria to inform program intervention in Vietnam. BMC Infect Dis. 20, 757 (2020).
Vantaux, A. et al. Anopheles ecology, genetics and malaria transmission in northern Cambodia. Sci Rep. 11, 6458 (2021).
U.S. President’s Malaria Initiative. U.S. President’s Malaria Initiative Cambodia Malaria Operational Plan FY 2022. 2021.
National Center for Parasitology, Entomology, and Malaria Control. National Treatment Guidelines for Malaria in Cambodia. 2022.
World Health Organization. Countries of the Greater Mekong ready for the “last mile” of malaria elimination, BUlletin #9. 2020.
Van Roey, K. et al. Field evaluation of Picaridin repellents reveals differences in repellent sensitivity between Southeast Asian vectors of malaria and Arboviruses. PLoS Negl Trop Dis 8, e3326 (2014).
St. Laurent, B. et al. Cow-baited tents are highly effective in sampling diverse Anopheles malaria vectors in Cambodia. Malar J. 15, 440 (2016).
Sanann, N. et al. Forest work and its implications for malaria elimination: A qualitative study. Malar J. 18, 376 (2019).
Bibbs, C. S. & Kaufman, P. E. Volatile pyrethroids as a potential mosquito abatement tool: A review of pyrethroid-containing spatial repellents. J. Integr. Pest Manag. https://doi.org/10.1093/jipm/pmx016 (2017).
Kongmee, M. et al. Irritant and repellent responses of Anopheles harrisoni and Anopheles minimus upon exposure to bifenthrin or deltamethrin using an excito-repellency system and a live host. J. Am. Mosq. Control Assoc. 28, 20–29 (2012).
Banks, S. D., Murray, N., Wilder-Smith, A. & Logan, J. G. Insecticide-treated clothes for the control of vector-borne diseases: A review on effectiveness and safety. Med Vet Entomol. 28, 14–25 (2014).
Maia, M. F., Kliner, M., Richardson, M., Lengeler, C. & Moore, S. J. Mosquito repellents for malaria prevention. Cochrane Database Syst. Rev. https://doi.org/10.1002/14651858.CD011595.pub2 (2018).
Dogan, E. B., Ayres, J. W. & Rossignol, P. A. Behavioural mode of action of deet: Inhibition of lactic acid attraction. Med. Vet. Entomol. 13, 97–100 (1999).
Afify, A., Betz, J. F., Riabinina, O., Lahondère, C. & Potter, C. J. Commonly used insect repellents hide human Odors from Anopheles mosquitoes. Curr. Biol. 29, 3669-3680.e5 (2019).
Wilson, A. L., Chen-Hussey, V., Logan, J. G. & Lindsay, S. W. Are topical insect repellents effective against malaria in endemic populations? A systematic review and meta-analysis. Malar J. 13, 446 (2014).
Okumu, F. O., Titus, E., Mbeyela, E., Killeen, G. F. & Moore, S. J. Limitation of using synthetic human odours to test mosquito repellents. Malar J. 8, 150 (2009).
Tangena, J.-A.A. et al. Field evaluation of personal protection methods against outdoor-biting mosquitoes in Lao PDR. Parasit Vectors 11, 661 (2018).
Ogoma, S. B. et al. A low technology emanator treated with the volatile pyrethroid transfluthrin confers long term protection against outdoor biting vectors of lymphatic filariasis, arboviruses and malaria. PLoS Negl. Trop Dis. 11, e0005455 (2017).
Masalu, J. P. et al. Efficacy and user acceptability of transfluthrin-treated sisal and hessian decorations for protecting against mosquito bites in outdoor bars. Parasit Vectors 10, 197 (2017).
Stevenson, J. C. et al. Controlled release spatial repellent devices (CRDs) as novel tools against malaria transmission: A semi-field study in Macha. Zambia. Malar J. 17, 437 (2018).
Tambwe, M. M. et al. Semi-field evaluation of freestanding transfluthrin passive emanators and the BG sentinel trap as a “push-pull control strategy” against Aedes aegypti mosquitoes. Parasites Vectors 13, 392 (2020).
Sangoro, O. P. et al. Evaluation of personal protection afforded by repellent-treated sandals against mosquito bites in south-eastern Tanzania. Malar J. 19, 148 (2020).
Tambwe, M. M., Moore, S., Hofer, L., Kibondo, U. A. & Saddler, A. Transfluthrin eave-positioned targeted insecticide (EPTI) reduces human landing rate (HLR) of pyrethroid resistant and susceptible malaria vectors in a semi-field simulated peridomestic space. Malar J. 20, 357 (2021).
Burton, T. A. et al. Semi-field evaluation of a volatile transfluthrin-based intervention reveals efficacy as a spatial repellent and evidence of other modes of action. PLoS ONE 18, e0285501 (2023).
Sukkanon, C. et al. Field evaluation of a spatial repellent emanation vest for personal protection against outdoor biting mosquitoes. J. Med. Entomol. 58, 756–66 (2021).
Charlwood, J. D. et al. Effects of the spatial repellent metofluthrin on landing rates of outdoor biting anophelines in Cambodia, Southeast Asia: Metofluthrin in Cambodia. Med. Vet. Entomol. 30, 229–234 (2016).
Ponlawat, A. et al. Semi-field evaluation of metofluthrin-impregnated nets on host-seeking Aedes aegypti and Anopheles dirus. J. Am. Mosq. Control Assoc. 32, 130–138 (2016).
Yan, C., Hii, J., Ngoen-Klan, R., Saeung, M. & Chareonviriyaphap, T. Semi-field evaluation of human landing catches versus human double net trap for estimating human biting rate of Anopheles minimus and Anopheles harrisoni in Thailand. PeerJ. 10, e13865 (2022).
Pennetier, C. et al. New protective battle-dress impregnated against mosquito vector bites. Parasites Vectors 3, 81 (2010).
Most, B. et al. Long-lasting permethrin-impregnated clothing: Protective efficacy against malaria in hyperendemic foci, and laundering, wearing, and weathering effects on residual bioactivity after worst-case use in the rain forests of French Guiana. Parasitol Res. 116, 677–684 (2017).
Williams, Y. A. et al. Expanding the vector control toolbox for malaria elimination: A systematic review of the evidence. Adv Parasitol. 99, 345–379 (2018).
Vajda, É. A. et al. A semi-field evaluation in Thailand of the use of human landing catches (HLC) versus human-baited double net trap (HDN) for assessing the impact of a volatile pyrethroid spatial repellent and pyrethroid-treated clothing on Anopheles minimus landing. Malar J. 22, 202 (2023).
Gordon, Scott, W. Personal protection tools from the deployed warfighter research program (DWFP) [Internet]. 2013. Available from: https://endmalaria.org/sites/default/files/4_Scott%20Gordon_DWFP_Outdoor%20WS.pdf
Combating the Aedes aegypti [Internet]. perimiterinsectguard.com/eto/. Available from: https://www.perimeterinsectguard.com/eto/
Eamsila, C., Frances, SP. EVALUATION OF PERMETHRIN.TREATED MILITARY UNIFORMS FOR PERSONAL PROTECTION AGAINST MALARIA IN NORTHEASTERN THAILAND’.
Rowland, M. et al. Permethrin-treated chaddars and top-sheets: Appropriate technology for protection against malaria in Afghanistan and other complex emergencies. Transact. R. Soc. Trop. Med. Hyg. 93, 465–472 (1999).
Kittayapong, P. et al. Mitigating diseases transmitted by Aedes mosquitoes: A cluster-randomised trial of permethrin-impregnated school uniforms. PLoS Negl Trop Dis 11, e0005197 (2017).
World Health Organization. WHO guidelines for malaria (World Health Organization, 2023).
Vontas, J. et al. Framework for rapid assessment and adoption of new vector control tools. Trends Parasitol. 30, 191–204 (2014).
Garros, C., Van Bortel, W., Trung, H. D., Coosemans, M. & Manguin, S. Review of the minimus complex of Anopheles, main malaria vector in Southeast Asia: From taxonomic issues to vector control strategies. Trop. Med. Int. Health. 11, 102–114 (2006).
Fairbanks, E. L. et al. Inference for entomological semi-field experiments: Fitting a mathematical model assessing personal and community protection of vector-control interventions. Comput. Biol. Med. 168, 107716 (2024).
Garrett-Jones, C. & Shidrawi, G. R. Malaria vectorial capacity of a population of Anopheles gambiae: An exercise in epidemiological entomology. Bull. World Health Organ. 40(4), 531 (1969).
Denz, A. et al. Predicting the impact of outdoor vector control interventions on malaria transmission intensity from semi-field studies. Parasit. Vectors 14, 64 (2021).
Miyamoto, J. Degradation, metabolism and toxicity of synthetic pyrethroids. Environ. Health Perspect. 14, 15–28 (1976).
Ogoma, S. B. et al. The mode of action of spatial repellents and their impact on vectorial capacity of Anopheles gambiae sensu stricto. PLoS ONE 9, e110433 (2014).
Chen, I. et al. Applying the COM-B behaviour change model to a pilot study delivering volatile pyrethroid spatial repellents and insecticide-treated clothing to forest-exposed populations in Mondulkiri Province, Cambodia. Malar J. 22, 251 (2023).
Panthusiri, P. Illustrated keys to the mosquitoes of Thailand IV. Anopheles. Southeast Asian J. Trop. Med. Public Health 37, 2 (2006).
www.boldsystems.org. Barcode of Life Data Systems Handbook: A web-based bioinformatics platform supporting the DNA barcoding of animal, plant, and fungal species. 2023.
R Core Team. R: A language and environment for statistical computing. Vienna, Austria; 2022.
Hadley Wickham, Davis Vaughan and Maximilian Girlich. tidyr: Tidy Messy Data. R package version 1.3.0. [Internet]. 2023. Available from: https://CRAN.R-project.org/package=tidyr
Hadley Wickham, Romain François, Lionel Henry and Kirill Müller. dplyr: A Grammar of Data Manipulation. R package version 1.0.8. [Internet]. 2022. Available from: https://CRAN.R-project.org/package=dplyr
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. https://doi.org/10.18637/jss.v067.i01 (2015).
Wickham, H. ggplot2: Elegant graphics for data analysis (Springer-Verlag, 2016).
Tambwe, M. M. et al. Human landing catches provide a useful measure of protective efficacy for the evaluation of volatile pyrethroid spatial repellents. Parasit. Vectors 16, 90 (2023).
Fassler, J., Cooper, P., BLAST Glossary. 2011 Jul 14. In: BLAST® Help [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2008.
Peyton, E. L. & Harrison, B. A. Anopheles (Celia) dirus, a new species of the Leucosphyrus group from Thailand 11 (Culicidae, 1979).
Manguin, S., Garros, C., Dusfour, I., Harbach, R. E. & Coosemans, M. Bionomics, taxonomy, and distribution of the major malaria vector taxa of Anopheles subgenus Cellia in Southeast Asia: An updated review. Infect. Genet. Evolut. 8, 489–503 (2008).
Rahman, W. A., Hassan, A. A., Adanan, C. R. & Rashid Mohd, R. The prevalence of Plasmodium falciparum and P. vivax in relation to Anopheles maculatus densities in a Malaysian village. Acta Trop. 55, 231–5 (1993).
Sinka, M. E. et al. The dominant Anopheles vectors of human malaria in the Asia-Pacific region: Occurrence data, distribution maps and bionomic précis. Parasit. Vectors 4, 89 (2011).
Tananchai, C., Manguin, S., Bangs, M. J. & Chareonviriyaphap, T. Malaria vectors and species complexes in Thailand: Implications for vector control. Trends Parasitol. 35, 544–558 (2019).
Chen, B. et al. Population genetics of the malaria vector Anopheles aconitus in China and Southeast Asia. Infect. Genet. Evolut. 12, 1958–1967 (2012).
Vantaux, A. et al. Contribution to malaria transmission of symptomatic and asymptomatic parasite carriers in Cambodia. J. Infect. Dis. 217, 1561–1568 (2018).
Zhang, C. et al. Survey of malaria vectors on the Cambodia, Thailand and China-Laos Borders. Malar J. 21, 399 (2022).
Lucas, J. R. et al. Laboratory and field trials of metofluthrin (sumione®) emanators for reducing mosquito biting outdoors1. J. Am. Mosq. Control Assoc. 23, 47–54 (2007).
Mmbando, A. S. et al. Small-scale field evaluation of push-pull system against early- and outdoor-biting malaria mosquitoes in an area of high pyrethroid resistance in Tanzania. Wellcome Open Res. 2, 112 (2017).
Swai, J. K. et al. CDC light traps underestimate the protective efficacy of an indoor spatial repellent against bites from wild Anopheles arabiensis mosquitoes in Tanzania. Malar J. 22, 141 (2023).
Sluydts, V. et al. Efficacy of topical mosquito repellent (picaridin) plus long-lasting insecticidal nets versus long-lasting insecticidal nets alone for control of malaria: A cluster randomised controlled trial. Lancet Infect. Dis. 16, 1169–1177 (2016).
Harbach, R. E., Tang, D. B. Relative repellency of two formulations of n,n-diethyl-3-methylbenzamide(deet) and permethrin-treatedclothing against culex si?Iens and Aedes Vigilax in Thailand’. 1990;
World Health Organization. Guidelines for monitoring the durability of long-lasting insecticidal mosquito nets under operational conditions. 2011;44.
Goodyer, L. I. et al. Expert review of the evidence base for arthropod bite avoidance. J. Travel Med. 17, 182–192 (2010).
U.S Environmental Protection Agency. Notice of pesticide registration [Internet]. 2016. Available from: https://www3.epa.gov/pesticides/chem_search/ppls/082392-00003-20160822.pdf
Lissenden, N. et al. Review and meta-analysis of the evidence for choosing between specific pyrethroids for programmatic purposes. Insects 12, 826 (2021).
The Assistant Secretary of Defense, (Health Affairs). Updated Policy for Prevention of Arthropod-Borne Diseases Among Department of Defense Personnel Deployed to Endemic Areas [Internet]. Health Affairs Policy: 07-007; 2007. Available from: https://apps.dtic.mil/sti/pdfs/ADA512584.pdf
Estep, A. S. et al. Failure of permethrin-treated military uniforms to protect against a laboratory-maintained knockdown-resistant strain of Aedes aegypti. J. Am. Mosq. Control Assoc. 36, 127–130 (2020).
Andrés, M., Lorenz, L. M., Mbeleya, E. & Moore, S. J. Modified mosquito landing boxes dispensing transfluthrin provide effective protection against Anopheles arabiensis mosquitoes under simulated outdoor conditions in a semi-field system. Malar J. 14, 255 (2015).
Trung, H. D. et al. Behavioural heterogeneity of Anopheles species in ecologically different localities in Southeast Asia: A challenge for vector control. Trop. Med. Int. Health 10, 251–262 (2005).
Machani, M. G. et al. Behavioral responses of pyrethroid resistant and susceptible Anopheles gambiae mosquitoes to insecticide treated bed net. PLoS ONE 17, e0266420 (2022).
Hemingway, J. & Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 45, 371–391 (2000).
Santolamazza, F. et al. Distribution of knock-down resistance mutations in Anopheles gambiae molecular forms in west and west-central Africa. Malar J. 7, 74 (2008).
Githeko, A. K., Mbogo, C. M., Atieli, F. A. & Juma, F. O. Sampling Anopheles arabiensis, A. gambiae sensu lato and A. funestus (Diptera: Culicidae) with CDC light-traps near a rice irrigation area and a sugarcane belt in western Kenya. Bull. Entomol. Res. 84, 319–24 (1994).
Chandre, F. et al. Modifications of pyrethroid effects associated with kdr mutation in Anopheles gambiae. Med. Vet. Entomol. 14, 81–88 (2000).
Wagman, J. M., Achee, N. L. & Grieco, J. P. Insensitivity to the spatial repellent action of transfluthrin in Aedes aegypti: A heritable trait associated with decreased insecticide susceptibility. PLoS Negl. Trop. Dis. 9, e0003726 (2015).
Agramonte, N. M., Bloomquist, J. R. & Bernier, U. R. Pyrethroid resistance alters the blood-feeding behavior in Puerto Rican Aedes aegypti mosquitoes exposed to treated fabric. PLoS Negl. Trop. Dis. 11, e0005954 (2017).
Bowman, N. M. et al. Pyrethroid insecticides maintain repellent effect on knock-down resistant populations of Aedes aegypti mosquitoes. PLoS ONE 13, e0196410 (2018).
Sherrard-Smith, E. et al. Mosquito feeding behavior and how it influences residual malaria transmission across Africa. Proc. Natl. Acad. Sci. USA 116, 15086–15095 (2019).
Afify, A. & Potter, C. J. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar J. 19, 127 (2020).
Liverani, M., Charlwood, J. D., Lawford, H. & Yeung, S. Field assessment of a novel spatial repellent for malaria control: A feasibility and acceptability study in Mondulkiri, Cambodia. Malar J. 16, 412 (2017).
World Health Organization. Data requirements and methods to support the evaluation of new vector control products. 2017;7.
Chitnis, N., Smith, T. & Steketee, R. A mathematical model for the dynamics of malaria in mosquitoes feeding on a heterogeneous host population. J. Biol. Dyn. 2, 259–285 (2008).
Wang, Y., Chitnis, N. & Fairbanks, E. L. Optimizing malaria vector control in the Greater Mekong Subregion: A systematic review and mathematical modelling study to identify desirable intervention characteristics. Parasit. Vectors 17, 162 (2024).
Dia, I. et al. Comparisons of human-landing catches and odor-baited entry traps for sampling malaria vectors in Senegal. J. Med. Entomol. 42(2), 104–109 (2005).
Charlwood, J. D., Rowland, M., Protopopoff, N. & Le Clair, C. The Furvela tent-trap Mk 1.1 for the collection of outdoor biting mosquitoes. PeerJ. 5, e3848 (2017).
Achee, N. L. et al. Spatial repellents: The current roadmap to global recommendation of spatial repellents for public health use. Curr. Res. Parasitol. Vector-Borne Dis. 3, 100107 (2023).
Briët, O. J. T. et al. Models of effectiveness of interventions against malaria transmitted by Anopheles albimanus. Malar J. 18, 263 (2019).
Acknowledgements
This study was funded by the Australia Department of Foreign Affairs and Trade (DFAT) through the Innovative Vector Control Consortium (IVCC), Liverpool, United Kingdom. We would also like to acknowledge and thank all the project teams, entomological collectors and technicians, local authorities, and fieldworkers who provided unwavering support throughout the study, even during the COVID-19 pandemic.
Author information
Authors and Affiliations
Contributions
E.A.V. substantially contributed to study conceptualization and design, led writing of protocol, SOPs, and data collection tools, led data curation, analysis, and synthesis, and led manuscript writing and review process. A.R. substantially contributed to protocol writing, data analysis, and manuscript review. D.D. led all fieldwork activities and contributed to manuscript review. E.F. led the modeling component and write-up in the manuscript, and also substantially contributed to manuscript review. N.C. supported the modeling component of the paper and contributed to manuscript review. J.H. substantially contributed to study design, protocol, and SOP writing, fieldwork activities, and manuscript review. S.J.M. substantially contributed to study conceptualization, protocol writing, data synthesis, and manuscript review. J.R. contributed to manuscript review. M.M. contributed contributed to manuscript review. S.S. contributed to study conceptualization and facilitated study rollout in Cambodia. P.K. provided support to fieldwork planning and activities. D.J.M. substantially contributed to study conceptualization, design, and protocol writing, and manuscript review. A.T. substantially contributed to study conceptualization, design, and protocol writing, and manuscript review. N.F.L. substantially contributed to study conceptualization, protocol writing, fieldwork, and manuscript review.
Corresponding author
Ethics declarations
Competing interests
J.H.R. and M.M. are employed by IVCC. While they took part in the manuscript review process, they were never involved with the study design, fieldwork, data analysis and interpretation processes. All other authors has no competing interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vajda, É.A., Ross, A., Doum, D. et al. Field evaluation of a volatile pyrethroid spatial repellent and etofenprox treated clothing for outdoor protection against forest malaria vectors in Cambodia. Sci Rep 14, 17348 (2024). https://doi.org/10.1038/s41598-024-67470-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-67470-3
- Springer Nature Limited