
Abstract
We have reconstructed baseline δ15N and δ13C of export production at Kingman Reef in the Central Equatorial Pacific (CEP) at sub-decadal resolution, nearly continuously over the last 2000 years. The changes in δ15N reflects the strength of the North Equatorial Counter Current (NECC) relative to the South Equatorial Current (SEC), and to a lesser extent, the North Equatorial Current (NEC). Seasonal to multi-decadal variation in the strength of these currents, through the redistribution of heat, have global climate impacts and influence marine and terrestrial ecosystems. We use modern El Niño-La Nina dynamics and the Tropical Pacific Decadal Variability (TPDV) pattern, which is defined in the CEP, as a framework for analyzing the isotopic data. The CEP δ15N and δ13C records exhibit multi-decadal (50–60 year) variability consistent with TPDV. A large multi-centennial feature in the CEP δ15N data, within age-model uncertainties, is consistent with one of the prolonged dry-pluvial sequences in the American west at the end of the Medieval Climate Anomaly, where low TPDV is correlated with drier conditions. This unique record shows that the strength of the NECC, as reflected in baseline δ15N and δ13C, has at quasi-predictable intervals throughout the late Holocene, toggled the phytoplankton community between prokaryotes and picoplankton versus eukaryotes.
Similar content being viewed by others


Introduction
The redistribution of excess incoming solar radiation (energy) in the tropics by the atmosphere and ocean dictates much of Earth’s climate system. The redistribution of energy as latent and sensible heat drives weather patterns which impacts billions of people, as well as strongly influencing terrestrial and marine ecosystems. Variability in sea surface temperature, which impacts weather and ultimately climate, occurs at sub-seasonal to multi-decadal scales. The tropical Pacific decadal sea surface temperature (SST) anomaly pattern (TPDV) has its highest amplitude in the Central Equatorial Pacific (CEP) and is similar to the pattern of El Niño-Southern Oscillation (ENSO) SST anomalies1. Decades with more La Niña events and fewer El Niño events have lower TPDV and the converse is also true. Interannual (ENSO) and decadal variability (TPDV) reflects and impacts the export of latent and sensible heat from the tropics to extra-tropics which sets up major weather patterns through the ascending and descending limbs of the Hadley Circulation and the position of the jet stream. TPDV is somewhat analogous to the Interdecadal Pacific Oscillation (IPO)2, and the Pacific Decadal Oscillation3. These SST patterns are dynamically connected to each other through tropical-extratropical teleconnections which nearly synchronize the north and south subtropical Pacific4. Together the TPDV and IPO explain a significant portion of global SST variability5. Modulations of the strength, intensity, and pattern of decadal SST (and sea level pressure) variability not only strongly impacts weather and weather extremes but marine ecosystems3,6,7.
By itself the instrumental record is not long enough nor with appropriate geographic coverage to record more than a few multi-decadal oscillations. Due to the lack of records with sufficient fidelity to capture multi-decadal variability the timing and drivers of oscillations before this period are poorly understood. Questions exist on how external forcing (e.g., greenhouse gases and aerosols from the industrial revolution or volcanoes) has influenced the pattern of decadal variability8, and if the variability is intrinsic to the ocean–atmosphere system as an internal periodic oscillation or whether it is stochastically forced quasi-periodic variability9,10. This leads to the question whether or not the recent past is a ‘clean’ analog of the instrumental era11,12. A related question is partitioning natural climate variability, as climate warmed out of the Little Ice Age (LIA) and internal multi-decadal variability from that of anthropogenic induced change13. Paleoclimate reconstructions, particularly those with seasonal to sub-decadal resolution (e.g., tree-rings, reef-building corals, speleothems, some sediment records), gap-fill in time and geography. Multi-decadal variability that is likely related to TPDV/IPO/PDO has been documented in paleo-records from, for example: sediments of the Santa Barbara Basin14, in speleothems that are influenced by the SE Asian and Australian monsoon15,16,17, speleothems that capture the South Pacific Convergence Zone18,19, and in tree-ring drought records from North America20,21 and SE Asia22. To date, however, there are no continuous multi-millennial sub-decadal resolved records from the CEP. This is the core region of the TPDV, and paleo-records are needed to answer if the variability is an intrinsic aspect of Earth’s climate system or if it has changed over longer (geological) timescales with, for example, different insolation patterns or other external forcings.
The Line Islands, a series of atolls and coral islands straddling the equator in the Central Pacific, are uniquely situated to elucidate past variations in sea surface temperature, salinity23,24 and rainfall associated with ENSO and/or the migration of the inter-tropical convergence zone (ITCZ)25,26,27. At 6° N, in the equatorial waveguide, Kingman Reef and nearby Palmyra Atoll are generally bathed by waters of the eastward flowing North Equatorial Counter Current (NECC)28,29,30,31. The NECC is sourced in the western equatorial Pacific from the Mindanao Current, and to a lesser extent the North Equatorial Current, which bring North Pacific Gyre surface water into the low-latitude tropics32. Interannual variations in the strength of NECC transport are associated with ENSO, and have the ability to bring significant amounts of heat from the western equatorial Pacific to the central and eastern Pacific during El Niño events28,30,33. During El Niños when the equatorial trade winds slacken or reverse, water from the North Pacific Gyre expands into the western equatorial Pacific34. To a large extent, SST and nutrients of surface water masses which mix in the CEP co-vary.
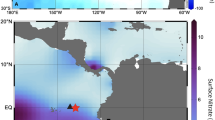
The source waters to the Line Islands from these different regions have highly distinct nitrate (NO3) isotope values. NECC nitrate levels in the western Pacific are low, often below detection limits, and comparable to the subtropical gyres35. In the western equatorial warm pool and the subtropical gyre nitrogen fixation augments intrinsic nitrate leading to low δ15NO3 values (0–5‰). Near the equator rates of nitrogen fixation decrease to the east36,37,38. Conversely, in the eastern Pacific, the NEC and SEC both have higher nitrate concentrations and more positive δ15NO3 values due to advection of waters impacted by denitrification signatures, as well as the fact that residual nitrate which has been upwelled to the surface has undergone progressive Rayleigh fractionation associated with incomplete nutrient utilization as waters are advected away from upwelling locations39,40,41. Consequently, surface δ15N-nitrate values at the equator are very high, δ15N values of 13–18‰ between 155° W and 170° W, before values decline northward into the subtropical gyre39,41. Nitrate isotope values of source waters of the east and west Pacific do not appear to have substantially shifted over the late Holocene42,43,44,45. This sets up a natural water-mass nutrient and 15N-isotope gradient which can be exploited to reconstruct water-mass variability in the CEP (Fig. 1, see also Supplementary Fig. 1).

Estimated surface ocean (0–130 m) δ15NO3 in an ocean general circulation model. Arrow vectors denote the main position of the equatorial branch of the South Equatorial Current (SEC), the North Equatorial Current (NEC), and the North Equatorial Counter Current (NECC). Star denotes the position of Kingman Reef and nearby Palmyra Atoll. The surface δ15NO3representation is intended to show the general patterns of the east to west and north to south δ15NO3 gradients, and has been adapted from Somes91. Although some of the details are different, particularly the westward extent of the NEC’s δ15NO3 signature, the model output is sufficiently similar to an observation-based interpolation92 for framing that the western equatorial Pacific has lower δ15NO3 compared to waters advected from the east, and that waters which feed the NECC are primarily derived from the North Pacific Subtropical Gyre.
Deep-sea proteinaceous corals are one of the best bioarchives to reconstruct surface water source and chemistry variations. Kulamanamana haumeaae, a deep-sea colonial zoanthid, is a low-order consumer46, and like other deep-sea corals, feeds primarily on recently exported sinking particulate organic carbon which tracks the carbon and nitrogen isotope values of sinking particulate organic carbon, i.e., export production47. The polyps create a hard, horny protein (gorgonin) skeleton, nearly purely amino acid based, which is resistant to degradation over multi-millennial timescales48,49. These amino acids are from surface production: either directly (source and essential amino acids) from diet or indirectly from diet via biosynthetic synthesis (trophic and non-essential). Bulk proteinaceous coral skeletal isotope values (δ13C, δ15N) are strongly correlated with source (δ15N) and essential (δ13C) amino acid values47,48,50, and provide insight into baseline nitrate variability and the impact of phytoplankton community structure on export production. Here, we present sub-decadal resolved reconstructions, spanning the last 2000 years from Kingman Reef (6° N, 162° W) in the Line Islands complex in the CEP, reflecting the isotopic composition of baseline nitrate (NO3) in surface waters supporting phytoplankton and export production. The changes in δ15N reflects the strength of the North Equatorial Counter Current (NECC) relative to the South Equatorial Current (SEC) and to a lesser extent, the North Equatorial Current (NEC). Corresponding carbon isotope data likely track changes in extant plankton assemblages, in particular variations between diatom and eukaryotic populations versus prokaryotic and/or nitrogen fixing algae. While our records do not resolve individual ENSO events, we will use modern ENSO dynamics as a framework for analyzing past changes.
Results and discussion
Collectively, the live-collected and sub-fossil Kulamanamana specimens capture, nearly continuously, the last 2000 years (Fig. 2). Calibrated age model uncertainties average ± 110 years (95% confidence interval) with the live-collected specimen (2005–1222 CE) having a resolution of 7 years per sample and the sub-fossil specimen (922–13 CE) approximately 5 years per sample. Diagnostics: amino acid mole percent (AA mol%) and carbon:nitrogen ratios (C/N) indicate good preservation of both skeletons and that except for the outermost 1–2 mm of the sub-fossil skeleton, the bulk stable isotope data are not compromised by diagenesis45,49. These specimens’ skeletal compound specific isotope analysis of amino acids (CSIA-AA) nitrogen data show a strong relationship between skeletal bulk and source AA δ15N values, allowing us to interpret bulk values as primarily tracking baseline δ15NO3 (Supplementary File 1).

Comparison of CEP TPDV as reflected by δ15N (blue line, this study), and δ13C (green line, this study), American West Palmer Drought Severity Index for the North American west (33–43° N, 106–119° W: data of Cook20,21), and two eastern Pacific El Niño proxy records: El Junco (1° S, 89° W, 700 m asl) of San Cristobal Island in the Galapagos Archipelago %Sand Index (green line, left axis: data of Conroy60) and the red color of sediment laminae in Laguna Palcacocha (2.8° S, 79.2° W, 4060 m asl) as reported in Moy61. The El Junco record has an age uncertainty that ranges from ± 10 years to ± 195 years and the Laguna Palcacocha may be biased < 100 years (IntCal98 versus SHCal20 framework). More (less) positive δ15N values are interpreted to reflect La Nina-like (El Niño-like) conditions and lower (higher) TPDV. In general, more positive δ13C values are interpreted to reflect a phytoplankton community dominated by eukaryotes including diatoms whereas less positive δ13C values reflect prokaryote and pico-plankton dominated communities. Washington Lake on Teraina Island (4.7° N, 160.4° W, near sea level) wet and dry periods as recorded in plant wax lipid biomarkers and sediment physical properties are noted. When Lake Washington dries out early in the LIA, locally the ITCZ is thought to be south of Teraina and thus south of Kingman Reef and Palmyra Atoll25,26,27. The large, multi-centennial TPDV excursion at the beginning of K1 (1200–1500) is, within age-model uncertainties, coincident with the American West and Sierra megadrought—pluvial-megadrought sequence noted by Stine62 and recorded in tree-ring reconstructions of regional PDSI.
Across the full data-set, the δ15N reconstruction exhibits complex behavior (Fig. 2). We document a long secular multi-centennial increase (~ 75–850 CE), a multi-centennial scale excursion during the end of the Medieval Climate Anomaly (MCA), a post Little Ice Age (LIA) rapid or sharp decrease (1860–2005 CE), and multi-decadal variability in both specimens. Except for the post- LIA and into the modern Industrial Era and its Suess effect51, δ13C variability exceeds that predicted from changes in atmospheric δ13C as reconstructed in ice cores52 or the impact of temperature on carbon isotope systematics in plankton53. In general, δ15N and δ13C values inversely covary, although the carbon isotope amplitude is smaller than that for nitrogen. For all the pre-industrial data 38% of the variance is shared with an evolution of tighter coupling in the older portion of the record. The only exception is the post-LIA and industrial era, in the younger portion of the record (see Supplementary File 1).
Within the ENSO model framework more positive δ15N indicates more frequent if not stronger La Niña events, a cooler CEP and lower TPDV. In contrast, less positive δ15N indicates more frequent/stronger El Niño events, warmer SSTs, and higher TPDV. Local impacts influencing baseline nitrate go hand in hand with the ENSO framework and amplify the baseline δ15N response. Local impacts such as time spans of increased stratification due to warmer temperatures and/or lower salinity due to increased local precipitation, could promote a local increase in nitrogen fixation. Such local effects tend to occur during El Niño events and a stronger influence of the NECC31. Through atmospheric teleconnections, a cooler CEP with low TPDV has been shown in climate models to lead toward a drier western North America54, which is similar to ENSO teleconnection impacts on interannual timescales.
Across the Late Holocene at the Peru Margin, the source of the SEC, sedimentary bulk δ15N has less than half a per mil range, or constant when considering analytical uncertainty (see Supplementary File 1). There is a similar lack of variability in open ocean cores from the eastern tropical Pacific44,45. We also do not observe significant variability in sediment records from the region where the NECC originates (see Supplementary File). Thus, the endmembers of δ15NO3 values in the two main water masses which influence our site have not significantly changed prior to the end of the LIA and into the industrial era.
We posit, therefore, that our CEP δ15N record is primarily capable at capturing the frequency of the mixing of source waters in the CEP, as opposed to the ability to precisely apportion the fraction of eastern versus western water. This is because, and although the δ15NO3 NECC, SEC, and NEC source endmembers do not appear to have significantly changed over the last 2000 years, that nitrate uptake by phytoplankton in SEC and NEC waters which are advected westward modifies the residual NO3 along a Rayleigh fractionation pathway leading towards more positive δ15NO3 values55. Thus, the ultimate δ15NO3 value of waters which are entrained into the NECC, or via the SEC directly impinging upon the northern Line Islands during sustained periods of anomalously strong trade winds, is driven by a complicated convolution of productivity changes. Such changes are very likely associated not only with trade-wind strength but also iron availability: through upwelling and entrainment of the Equatorial Under Current and Subantarctic waters56,57, dust deposition58, and recycling of iron59. Additionally, if in the far western Pacific the NECC endmember δ15NO3 changes due to nearly basin-wide changes in δ15NO3, it mainly would change the amplitude of the CEP δ15N variability but leave the spectral character intact, because the general feature of less positive values in the NECC and more positive in the SEC and NEC would remain. Our NECC TPDV δ15N proxy record has its highest multi-decadal variability at 50–65 years (Fig. 3). This frequency range is similar to speleothem based reconstructions of the SPCZ18,19 and the SE Asian and Indo-Pacific monsoon systems15,16,17.

Spectral analysis of the Kingman Reef δ15N data as a function of frequency (year−1). (A) Multi-taper method (MTM) results of the K1 specimen (1220–1880 CE). (B) Multi-taper method results of the K2 specimen (13–858 CE) and (C) Blackman–Tukey results of K1 (solid, bold line) and K2 (dashed line), together. Confidence significance levels for the MTM results were determined using robust red-noise intrinsic to the data. The 95% confidence interval is shown for the Blackman–Tukey results. Data were handled as described in the "Methods" section. The additional arrow highlights the 50–60 year period, consistent with our understanding of TPDV. For clarity, the announcement of the 50–60 year period is only in (A).
Eastern Pacific proxy reconstructions of ‘classic’ El Niño60,61 indicate that the early part of our record, ~ 0–500 (Common Era) CE, is replete with stronger and or more frequent El Niños. Around 500 CE ENSO transitions into more frequent La Niñas (Fig. 2). Our CEP δ15N record, in addition to visually striking, and statistically significant multi-decadal variability, captures this shift in tendency as the trend towards more positive δ15N values indicative of a stronger influence of SEC water. Furthermore, the visual correlations between CEP δ15N and the US American West Palmer Drought Severity Index (PDSI) record are intriguing, albeit we note that with our age-model uncertainties there are nearly as many instances of a mis-match at the < 100 year level as there are matches. The largest multi-centennial feature in the CEP δ15N data, within age-model uncertainties, is consistent with one of the prolonged dry-pluvial sequences in the American west at the end of the MCA20,21,62, i.e., the pluvial between Stine’s two massive droughts. We do not have complete coverage through the MCA to ascertain the decadal variability in the CEP. The observed data and correlations, however, suggest that the CEP was likely in a state of more frequent and possible intense La Niña events with low TPDV leading to a cooler CEP. This is similar to correlations over ENSO timescales. Over their common overlap in time, our data and the tree-ring based North American PDSI reconstruction share common multi-decadal variability (see Supplementary File).
During this time interval, ~ 0 to 700 CE, a speleothem-based reconstruction of the Indo-Australian and SE Asian monsoon systems15 also has what has been interpreted as a decrease in the intensity of monsoon rainfall and a contraction of these systems. Although contrary to a different putative ITCZ reconstruction which includes records across the whole of the tropics17, this is consistent with our record of more La Niña like character, stronger trade-winds, and reduced TPDV. Leaf-wax deuterium isotopes63, a rainfall proxy, from Indonesia implies a stronger Indonesian Monsoon across 0–1600 CE punctuated by periods of weaker monsoon intensity during − 100 to 320 CE (± 200 years, 95%CI) ~ 850 to 1230 (± 200 years, 95% CI). The latter period of weaker Indonesian Monsoon is replicated in speleothem-based data from Thailand16. However, the inter-tropical convergence zone (ITCZ) is not a monolith and is separate from the terrestrial monsoon systems which are more directly anchored to landmasses. Thus, we may anticipate periods when the pan-Pacific or local CEP position of the ITCZ does not exactly reflect the inferred position and intensity of the monsoon-based systems.
Entering the Little Ice Age period, biomarker and sediment physical property reconstructions of the mean position of the ITCZ indicates that for a portion of the LIA, approximately 1420 to 1600 CE, locally the ITCZ was south of Washington Island, which is at 4.7° N within the Line Islands25,26,27 and thus south of Kingman Reef. Baseline δ15NO3 values through this period are consistently less positive than earlier in the record and exhibit smaller amplitude multi-decadal variability. Low amplitude δ15N variability is a characteristic of the LIA, even when the ITCZ moves back north of Washington Island. Low TPDV and a cool CEP is consistent with proxies of ENSO where El Niños are less frequent or intense. This is however separate to the inference of increasing precipitation over the Galapagos Islands in the eastern Pacific, even in the absence of frequent or strong El Niños60,64. Within the LIA, Indonesian and SE Asian monsoon proxies generally include more intense and less intense centennial scale periods16,22,63.
The modern, post-LIA and industrial era δ15N exhibits both multi-decadal variability and a large, nearly 2‰, decrease in values. The rate of change during the latter half of the twentieth century exceeds that of the centennial scale excursion in the MCA by a factor of two to three (0.3‰-decade−1 vs. ~ 0.1‰-decade−1), with comparable rates over 100s of years. Reconstructions of the δ15N in the Mindanao Current from sediment cores43,44 do not have the fidelity to capture this decrease. After the 1976/77 shift in the IPO/PDO, the tropical Pacific experienced multiple strong El Niño events, which in our ENSO framework would lead to a stronger NECC bringing warm, low δ15NO3 water to the CEP. Because of its length, the CEP δ15N expression may miss the shift back to weaker El Niño events post 200065 and thus, the greater impact of cooler more positive δ15NO3 water in the CEP. Missing half of the oscillation would give the appearance of a massive secular δ15NO3 shift. A modification to this interpretation is that there is indeed a change in the δ15NO3 in the western Pacific associated with a post-LIA/industrial era increase in nitrogen fixation. The increase in nitrogen fixation could be specific to the Western Pacific or it could include the North Pacific surface gyre waters from which the NECC is sourced.
Near the Hawaiian Islands in the North Pacific Subtropical Gyre, post-LIA and through the industrial era baseline δ15NO3 decreased due to a phytoplankton community with more nitrogen fixing bacteria in response to increased stratification48,50. Within the gyre, the Hawaiian Islands span a natural south to north less oligotrophic to more oligotrophic gradient48. The post LIA δ15N amplitude of change, 1.2–1.4‰, near French Frigate Shoals is less than near Oahu (~ 2‰) and Cross Seamount (~ 2.5‰). Based on estimates of nitrate not supported by Redfield ratio remineralization, N*66, French Frigate Shoals may be more representative of the gyre as a whole. Gyre surface water expanding into the western Equatorial Pacific during El Niño events and an increase in entrainment of gyre surface waters with low δ15N values into the NEC would, in tandem, work to produce the observed CEP δ15N trend. Such an interpretation would also explain twentieth century long-term δ15N variability at the base of the nitricline in the Western Pacific67,68. These data imply that similarly to the NPSG, regionally in the CEP, due to increased stratification coincident with more frequent El Niño events, has also seen an increase in nitrogen fixation and an implicit change in the phytoplankton community post LIA.
The δ13C records, while not having as large amplitude changes, are largely consistent with δ15N data within our interpretational framework. In the δ13C time-series, excluding the post-LIA/industrialized era, there are three main long-term δ13C value changes: increase between ~ 200–700 CE and ~ 1250–1500 CE during the MCA, and an approximate plateau from ~ 1500 to 1820 CE during the LIA. All these δ13C changes exceed those of atmospheric CO252 and thus cannot be solely the result of air-sea δ13CO2 exchange and equilibration. Post-LIA and into the industrialized era the near continuous 1.2–1.5‰ δ13C decrease is ~ 60% of the decrease observed in the atmosphere and interpreted to be dominated by the Suess Effect51. The coupling between δ13C and δ15N, i.e., a tendency towards commensurate directional changes in carbon and nitrogen isotope values, is reflected in the similarity of the δ13C data’s spectral analysis (Supplementary Fig. 9). Like δ15N, the δ13C data exhibit consistent multi-decadal variability with 50–60 year periodicity.
We interpret the pre-industrial era δ13C data in a phytoplankton community framework. CSIA-AA δ13C in live-collected and sub-fossil Kulamanamana haumeaae from off Oahu have confirmed the ability of bulk skeletal carbon isotopes in Kulamanamana to capture broad changes in community structure46,50. Different main phytoplankton groups have ranges of isotopic fractionation broadly linked to cell size, growth rate, and other factors, relative to in situ dissolved inorganic carbon53,69,70. Major groups of the equatorial Pacific include Prochlorococcus, Synechococcus, nano-eukaryotes and diatoms. Picoplankton typically have more negative δ13C values compared to larger phytoplankton species. Diatoms, in particular, have characteristically enriched δ13C values linked to cell size and often faster growth rate in nutrient replete conditions69,70,71,72,73. In general, the relative abundance of these taxons leads to more positive δ13C primary and export production values near the equator and more negative values in the oligotrophic gyres73.
In the NPSG this framework can not only explain most δ13C shifts, but also explains coupled δ13C and δ15N changes50. Skeletal δ15N values are generally coupled with δ13C values in both records (K1 r2 = 0.55, K2 r2 = 0.65), consistent with the expectation that coupled atmosphere–ocean dynamic changes associated with the strength of the NECC toggles community composition between a community of prokaryotes and picoplankton (lower δ13C values) versus eukaryotes such as diatoms (more positive δ13C values). Specifically, the periods of lower δ13C values (during 0–200 CE, ~ 1200–1400 CE) suggest greater abundance of smaller celled prokaryotes, coincident with weakened trade winds, and a stronger NECC. Periods of higher δ13C (e.g., during ~ 500–850 CE and 1450–1850 CE), in contrast, represent enhanced larger cell (especially diatom) contributions, which would be expected to be facilitated by increased nutrient transport from the east Pacific during La Niña-like conditions.
There are also some exceptions to the typical tight coupling of δ13C and δ15N values in the record. The clearest exceptions are the ~ 1‰ oscillations in δ15N values found in the K2 record and the first half of the multi-centennial δ15N excursion ~ 1350 to 1500 CE in the K1 record. We hypothesize that oscillation events observable in δ15N during these decoupled periods may be tracking changes in source water δ15N signatures, rather than localized changes in stratification and nitrate concentration which would be expected to influence the overall phytoplankton community, and so also δ13C values. We note that similar “decoupling” behavior between δ13C and δ15N has been documented in other sub-decadal resolved marine isotope records over the last 5000 years45. One possible interpretation of the decoupling between δ13C and δ15N which has previously been proposed is a shift in phytoplankton community composition between nitrate utilizing cyanobacteria versus nitrogen fixing cyanobacteria with a less positive δ15N signature. This would be expected to alter δ15N of export production without substantially changing δ13C values as the phytoplankton composition remains prokaryotic50.
Conclusions
We have determined that TPDV, as reflected by water-mass dynamics in the CEP, has been a near constant fixture in the climate system over at least the last 2000 years. There is at least one period of centennial scale low TPDV which, based on modern teleconnections, is consistent with extreme drought in the American Southwest. We hypothesize that there is a second significant centennial scale period of low TPDV in the data gap between our two individual records. Due to age-model uncertainties in our record we cannot yet clearly determine if the CEP TPDV has a tight, narrow periodicity which could imply an internal oscillation intrinsic to the coupled climate system. Moreover, better chronological control is required to refine the visual correlations we have made with ENSO and drought proxies and to ascertain if there are slight temporal offsets between land-mass monsoon and drought proxies and open ocean CEP sites influenced by the ITCZ. This is regardless of the commonality in the frequency domain of monsoon proxy and terrestrial drought records which exhibit similar multi-decadal variability. Through atmospheric teleconnections CEP variability is expressed in regional drought and pluvial extremes. The general coupling between δ13C and δ15N that we observe in our records is consistent with the framework where physical current (ocean dynamics) and climate regime directly influence phytoplankton community across timescales. In our framework, generally, times of low TPDV promote eukaryote-based phytoplankton communities whereas during high TPDV prokaryote and pico-plankton-based communities, including nitrogen fixers, are selected for.
The centennial period of low TPDV coincident with North American drought provides a natural testbench for models to elucidate not only how TPDV varies with background climate but how the background climate state influences atmospheric teleconnections in models. Modelling studies of future tropical Pacific decadal variability have a wide range of results1,5. Thus, our data provide a new benchmark for estimating, through hindcasting, the quality of models’ TPDV in the near instrumental era realm under known, but different boundary conditions54,74. Although we do not resolve individual El Niño-La Niña events, our record captures the decadal waxing and waning of ENSO such as observed in the instrumental record. Reproduction of ENSO, when compared to instrumental data and where in the surface ocean the signal occurs, is lacking in nearly all of the CMIP5 and CMIP6 models75. The lack of accurate reproduction of ENSO in the instrumental era has corollaries to diverse and inconsistent results in future climate change scenario predictions. A better mechanistic understanding of what conditions lead to different expressions of ENSO diversity and how anthropogenic induced climate effects impacts the background climate, including TPDV, is needed to better predict ENSO variability and diversity in the future.
Methods
Live (K1) and sub-fossil (K2) K. haumeaae specimens were collected using DSRV Pisces IV and V from ∼ 400 m depth offshore Kingman Reef (6° 23′ N, 162° 25′ W) in 2005. Skeletons were first washed with seawater and then fresh water before being air-dried on deck. Cross section disks ∼ 0.7 cm thick were cut from close to the basal attachment, polished, and mounted onto glass plates before being drilled by a Merchanteck micromill45,48,49. Approximately 2–3 mg of proteinaceous coral skeleton was collected at contiguous 0.1 mm resolution along radial transects from the outer edge to the center. The direct center was avoided due to concerns over the host coral parasitized by the K. haumeaae organisms.
Bulk δ15N, δ13C, and C/N analyses were performed synchronously on ∼ 0.3 mg raw material using a Carlo Erba 1108 elemental analyzer coupled to a ThermoFinningan Delta Plus XP isotope ratio mass spectrometer. Results are reported in conventional per mil (‰) notation relative to air and VPDB standards for δ15N and δ13C, respectively. Reproducibility based on independent coral replicates is consistent with the 1-sigma sd of the acetanilide standard of < 0.1‰ for both isotopes. Analysis of replicate and standard C/N values indicate a 1-sigma sd reproducibility of 0.2.
Radiocarbon (14C) analyses were performed on ~ 1 mg acid-pretreated sub-samples (n = 6 for K1, n = 7 for K2). The calibrated model for each coral was generated in a Bayesian framework76. Post-1950 ages in K1 were estimated using a surface water F14C reconstruction from nearby Palmyra77. The remaining 14C ages were calibrated using a local reservoir (ΔR) correction of − 134 ± 25 years78 and the Marine20 database79. Average age model uncertainties at the ± 95% confidence interval for each coral is approximately ± 110 years.
Utilizing C/N values the outermost 1–2 mm of K2, the subfossil specimen, was determined to be likely impacted by diagenesis and so was excluded from our interpretation45,49. Data for these samples are included in the supplement for completeness.
Spectral analysis of the bulk δ15N and δ13C time-series was independently performed on each record. Each record was first linearly interpolated to their average resolution: K1 to 7 years and K2 to 5 years. The spectral analysis was performed on detrended, z-score normalized, and simple interpolated versions of the records. Final analysis included passing the data through a 3-point (21 year) or 5-point (25 year) moving average filter for K1 and K2, respectively. Since our focus is natural variability, we excised the post 1880 CE portion of K1. We utilized both a Blackman-Tukey method implemented in the ARAND package80 where we lagged, by one-third, the auto-covariance function and a Multi-Taper Method as implemented in Kspectra (Spectraworks), using 3 tapers, compared against a red-noise AR (1) process with robust noise estimated from the data-set being analyzed. The Blackman-Tukey method is reported against the 95% confidence interval, and the Multi-Taper Method reported with predicted curves at the 90%, 95%, and 99% confidence interval. Spectral signatures were not dependent on data handling (z-score, detrended, simple interpolation).
Compound specific isotope amino acid (CSIA-AA) δ15N analyses were performed utilizing skeletal material from combining adjacent layers to have enough material (~ 6–10 mg) for measurements. Analysis involved established wet chemistry protocols for coral materials (e.g. McMahon et al. 46) where amino acids (AAs) were liberated using standard acid hydrolysis conditions (1 ml of 6 N HCl at 110 °C for 20 h), then spiked with a norleucine (Nor-Leu) internal standard and derivatized81, followed by purification with cation-exchange chromatography and a salt-removal step (p-buffer = KH2PO4 + Na2HPO4 in Milli-Q water, ph 7) before a 3× chloroform rinse and centrifugation before the final conversion to trifluoroacetyl/isopropyl ester derivatives. Derivatized samples were injected in triplicate on a coupled Gas Chromatography-IRMS (ThermoTrace GC, coupled to a Delta + IRMS). Injections were made utilizing concentrations adjusted to produce at least 80 mV IRMS N2 signal intensity for the smallest peaks (typically phenylalanine and isoleucine).
Isotope ratios were measured on 13 AAs: alanine (Ala), glycine (Gly), serine (Ser), valine (Val), threonine (Thr), leucine (Leu), isoleucine (Ile), proline (Pro), phenylalanine (Phe), tyrosine (Tyr), lysine (Lys), glutamine + glutamic acid (Glx), and asparagine + aspartic acid (Asx). A range of verification approaches were simultaneously employed to ensure accurate CSIA-AA data. The Nor-Leu internal standard was used to verify that injections gave expected results. Bracketed standards of l-amino acids were repeatedly analyzed after every triplicate coral skeleton injection, and the average measured value of each external standard AA across the entire run was compared against its authentic value, and any systematic bias/offset used to correct sample AA values82. Lastly, we utilized an internal laboratory reference material (dried/homogenized cyanobacteria) which was run with each sample batch to evaluate accuracy against a long term (> 10 years) internal control, ensuring long-term reproducibility for individual AA isotope values. Analytical reproducibility from triplicate sample injections was, in general, < 1‰ for δ15N CSIA-AA.
From the CSIA-AA data weighted averages of source AAs (Tyr, Lys, and Phe) were calculated using the analytical uncertainty as the weighting function83. The presence of heterotrophic microbial resynthesis was estimated using the CSIA-AA metric ∑V84.
A total of five (0.5–1 mg) samples (K1 n = 2 and K2 n = 3) from the same CSIA-AA composite samples was analyzed for AA molar concentrations. Wet chemical protocols for AA measurements in proteinaceous corals followed established protocols48,49. The AA mole percent (mol%) compositions were quantified using a GC–MS (Agilent 7890 GC coupled to a 5975 MSD) based on single ion monitoring data of the major ion relative to authentic AA external standard calibration curves. Commercial AA standards (Pierce Biochemicals) were used to create concentration series, and response factors from these external standards were used to calculate relative molar concentrations. Reproducibility, as measured by the standard deviation of GC–MS replicates analyses, typically averaged < 5 mol%.
Where possible, records which rely on 14C chronologies and discussed in the manuscript or presented in figures have been updated using 14C-calendar frameworks of IntCal20/SHCal20/Marine2079,85,86, and local estimates of ∆R (www.calib.org). Without the individual varve data, we were unable to update the Laguna Palcacocha ENSO reconstruction61. We do note that the discrete 14C data used as scaffolding by Moy et al., when calibrated using SHCal20, are less than 100 calibrated years different than the (IntCal98) calibrated ages presented in Rodbell for the same core87. The El Junco sand record60, an ENSO proxy, has an age model uncertainty which waxes and wanes from ± 10 years (where 210Pb constrains the upper portion) to ± 195 years, and averages 115 years (95% CI). Spectral comparison with western North America tree-ring based PDSI20,21 was done for discrete sections of the tree-ring reconstruction consistent with the age spans of the two Kulamanamana specimens and including the 95% CI uncertainty in the coral age-models. Annual tree-ring data were filtered with a 21 year moving-average filter prior to spectral analysis via convolution of the 1/3-lagged autocovariance function, or via the Multi-Taper Method.
We note that in the literature there are more records to compare our data against, including paleosalinity and precipitation proxy records from the Makassar Straits63,88,89. These sediment cores’ data, except where constrained by 210Pb, when put into a Bayesian age-model framework, have an approximately ± 200 year age model uncertainty, or twice that of ours. Such uncertainty in the age-model would lead to a less than ideal comparison. We note that these paleosalinity data were differenced against a Galapagos precipitation sand-based proxy record60 to estimate past variations in the Southern Oscillation Index90. Yan et al. only included the analytical uncertainty of the underlying 14C ages, not the calibrated age uncertainties: i.e., the temporal uncertainty is underestimated by a factor of 2–3×, which leads to fundamental questions in the SOI reconstruction itself. Within the framework of documenting the cyclicity in the CEP δ15N data and ascertaining if it is consistent with modern TPDV or not, comparison with all possible paleo-records is not required.
Data availability
In addition to being provided as a Supplementary Datafile, these data are digitally archived at NCEI/NOAA Paleoceanography (https://www.ncei.noaa.gov/access/paleo-search/study/38820).
References
Power, S. et al. Decadal climate variability in the tropical Pacific: Characteristics, causes, predictability, and prospects. Science 374, 9165 (2021).
Power, S., Casey, T., Folland, C., Colman, A. & Mehta, V. Inter-decadal modulation of the impact of ENSO on Australia. Clim. Dyn. 15, 319–324 (1999).
Mantua, N. J., Hare, S. R., Zhang, Y., Wallace, J. M. & Francis, R. C. A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteor. Soc. 78, 1069–1079 (1997).
Linsley, B. K., Wellington, G. M. & Schrag, D. P. Decadal sea surface temperature variability in the subtropical south Pacific from 1726 to 1997 A.D.. Science 290, 1145–1148 (2000).
Di Lorenzo, E. et al. Modes and mechanisms of Pacific decadal-scale variability. Annu. Rev. Mar. Sci. 15, 249–275 (2023).
Martinez, E., Antoine, D., D’Ortenzio, F. & Gentili, B. Climate-driven basin-scale decadal oscillations of oceanic phytoplankton. Science 326, 1253–1256 (2009).
Di Lorenzo, E. et al. North Pacific Gyre oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 35, L08607 (2008).
Mann, M. E., Steinman, B. A., Brouillette, D. J. & Miller, S. K. Multidecadal climate oscillations during the past millennium driven by volcanic forcing. Science 371, 1014–1019 (2021).
Okumura, Y. M. Origins of tropical Pacific decadal variability: Role of stochastic atmospheric forcing from the south Pacific. J. Clim. 26, 9791–9796 (2013).
Sun, T. & Okumura, Y. M. Role of stochastic atmospheric forcing from the south and north Pacific in tropical Pacific decadal variability. J. Clim. 32, 4013–4038 (2019).
Smith, D. M. et al. Role of volcanic and anthropogenic aerosols in the recent global surface warming slowdown. Nat. Clim. Change 6, 936–940 (2016).
Hua, W., Dai, A. & Qin, M. Contributions of internal variability and external forcing to the recent Pacific decadal variations. Geophys. Res. Lett. 45, 7084–7092 (2018).
The PAGES 2k Consortium. Early onset of industrial-era warming across the oceans and continents. Nature 536, 411–418 (2016).
Friddell, J. E., Thunell, R. C., Guilderson, T. P. & Kashgarian, M. Increased northeast Pacific climatic variability during the warm middle Holocene. Geophys. Res. Lett. 30, 1560 (2003).
Denniston, R. F. et al. Expansion and contraction of the Indo-Pacific tropical rain belt over the last three Millennia. Sci. Rep. 6, 34485 (2016).
Tan, L. et al. Rainfall variations in central Indo-Pacific over the past 2,700 years. Proc. Natl. Acad. Sci. U.S.A. 116, 17201–17206 (2019).
Lechleitner, F. A. et al. Tropical rainfall over the last two millennia: Evidence for a low-latitude hydrologic seesaw. Sci. Rep. 7, 45809 (2017).
Maupin, C. R. et al. Persistent decadal-scale rainfall variability in the tropical South Pacific Convergence Zone through the past six centuries. Clim. Past 10, 1319–1332 (2014).
Partin, J. W. et al. Multidecadal rainfall variability in South Pacific Convergence Zone as revealed by stalagmite geochemistry. Geology 41, 1143–1146 (2013).
Cook, E. R. et al. Megadroughts in North America: Placing IPCC projections of hydroclimatic change in a long-term palaeoclimate context: Megadroughts in North America. J. Quat. Sci. 25, 48–61 (2010).
Cook, E. R., Woodhouse, C. A., Eakin, C. M., Meko, D. M. & Stahle, D. W. Long-term aridity changes in the western United States. Science 306, 1015–1018 (2004).
Buckley, B. M., Palakit, K., Duangsathaporn, K., Sanguantham, P. & Prasomsin, P. Decadal scale droughts over northwestern Thailand over the past 448 years: Links to the tropical Pacific and Indian Ocean sectors. Clim. Dyn. 29, 63–71 (2007).
Cobb, K. M., Charles, C. D. & Hunter, D. E. A central tropical Pacific coral demonstrates Pacific, Indian, and Atlantic decadal climate connections. Geophys. Res. Lett. 28, 2209–2212 (2001).
Sanchez, S. C. et al. A continuous record of central tropical Pacific climate since the midnineteenth century reconstructed from fanning and Palmyra Island Corals: A case study in coral data reanalysis. Paleoceanogr. Paleoclimatol. 35, 848 (2020).
Higley, M. C., Conroy, J. L. & Schmitt, S. Last millennium meridional shifts in hydroclimate in the central tropical Pacific. Paleoceanogr. Paleoclimatol. 33, 354–366 (2018).
Sachs, J. P., Mügler, I., Sachse, D., Prebble, M. & Wolhowe, M. Last millennium hydroclimate in the central equatorial North Pacific (5° N, 160° W). Quat. Sci. Rev. 259, 106906 (2021).
Wyman, D. A., Conroy, J. L., Osburn, M. R. & Atwood, A. R. Coeval drying across the central tropical Pacific over the last Millennium. Paleoceanogr. Paleoclimatol. 36, e2021PA004311 (2021).
Wyrtki, K. Teleconnections in the equatorial Pacific ocean. Science 180, 66–68 (1973).
Wyrtki, K. & Kilonsky, B. Mean water and current structure during the Hawaii-to-Tahiti shuttle experiment. J. Phys. Oceanogr. 14, 242–254 (1984).
Webb, D. J. On the role of the North Equatorial Counter current during a strong El Niño. Ocean Sci. 14, 633–660 (2018).
Brainard, R. et al. Chapter 3 Kingman Reef. In Coral Reef Ecosystem Monitoring Report for the Pacific Remote Islands Marine National Monument 2000–2017 89 (Pacific Islands Fisheries Science Center, 2019).
Fine, R. A., Lukas, R., Bingham, F. M., Warner, M. J. & Gammon, R. H. The western equatorial Pacific: A water mass crossroads. J. Geophys. Res. 99, 25063 (1994).
Qiu, B. Large-scale variability in the midlatitude subtropical and subpolar North Pacific Ocean: Observations and causes. J. Phys. Oceanogr. 32, 23 (2002).
Guilderson, T. P., Schrag, D. P., Kashgarian, M. & Southon, J. Radiocarbon variability in the western equatorial Pacific inferred from a high-resolution coral record from Nauru Island. J. Geophys. Res. 103, 24641–24650 (1998).
Fiedler, P. C. & Talley, L. D. Hydrography of the eastern tropical Pacific: A review. Prog. Oceanogr. 69, 143–180 (2006).
Yoshikawa, C., Yamanaka, Y. & Nakatsuka, T. Nitrate-nitrogen isotopic patterns in surface waters of the western and central equatorial Pacific. J. Oceanogr. 62, 511–525 (2006).
Lehmann, N. et al. Isotopic evidence for the evolution of subsurface nitrate in the western equatorial Pacific. J. Geophys. Res. Oceans 123, 1684–1707 (2018).
Bonnet, S., Biegala, I. C., Dutrieux, P., Slemons, L. O. & Capone, D. G. Nitrogen fixation in the western equatorial Pacific: Rates, diazotrophic cyanobacterial size class distribution, and biogeochemical significance: N2 fixation in the equatorial Pacific. Glob. Biogeochem. Cycles 23, 1 (2009).
Rafter, P. A., Sigman, D. M., Charles, C. D., Kaiser, J. & Haug, G. H. Subsurface tropical Pacific nitrogen isotopic composition of nitrate: Biogeochemical signals and their transport: Tropical Pacific nitrate isotopes. Glob. Biogeochem. Cycles 26, 979 (2012).
Peters, B. D., Lam, P. J. & Casciotti, K. L. Nitrogen and oxygen isotope measurements of nitrate along the US GEOTRACES Eastern Pacific Zonal transect (GP16) yield insights into nitrate supply, remineralization, and water mass transport. Mar. Chem. 201, 137–150 (2018).
Rafter, P. A. & Sigman, D. M. Spatial distribution and temporal variation of nitrate nitrogen and oxygen isotopes in the upper equatorial Pacific Ocean: Equatorial Pacific nitrate N and O isotopes. Limnol. Oceanogr. 61, 14–31 (2016).
Altabet, M. A. et al. The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific. Deep Sea Res. I Oceanogr. Res. Pap. 46, 655–679 (1999).
Kienast, M. et al. A mid-Holocene transition in the nitrogen dynamics of the western equatorial Pacific: Evidence of a deepening thermocline? Geophys. Res. Lett. 35, L23610 (2008).
Tesdal, J. E., Galbraith, E. D. & Kienast, M. The Marine Sedimentary Nitrogen Isotope Record. https://bg.copernicus.org/preprints/9/4067/2012/bgd-9-4067-2012.pdf, https://doi.org/10.5194/bgd-9-4067-2012 (2012).
Glynn, D. S., McMahon, K. W., Guilderson, T. P. & McCarthy, M. D. Major shifts in nutrient and phytoplankton dynamics in the North Pacific Subtropical Gyre over the last 5000 years revealed by high-resolution proteinaceous deep-sea coral δ15N and δ13C records. Earth Planet. Sci. Lett. 515, 145–153 (2019).
McMahon, K. W., Williams, B., Guilderson, T. P., Glynn, D. S. & McCarthy, M. D. Calibrating amino acid δ13C and δ15N offsets between polyp and protein skeleton to develop proteinaceous deep-sea corals as paleoceanographic archives. Geochim. Cosmochim. Acta 220, 261–275 (2018).
Shen, Y. et al. Amino acid δ13C and δ15N patterns from sediment trap time series and deep-sea corals: Implications for biogeochemical and ecological reconstructions in paleoarchives. Geochim. Cosmochim. Acta 297, 288–307 (2021).
Sherwood, O. A., Guilderson, T. P., Batista, F. C., Schiff, J. T. & McCarthy, M. D. Increasing subtropical North Pacific Ocean nitrogen fixation since the Little Ice Age. Nature 505, 78–81 (2014).
Glynn, D. S., McMahon, K. W., Sherwood, O. A., Guilderson, T. P. & McCarthy, M. D. Investigating preservation of stable isotope ratios in subfossil deep-sea proteinaceous coral skeletons as paleo-recorders of biogeochemical information over multimillennial timescales. Geochim. Cosmochim. Acta 338, 264–277 (2022).
McMahon, K. W., McCarthy, M. D., Sherwood, O. A., Larsen, T. & Guilderson, T. P. Millennial-scale plankton regime shifts in the subtropical North Pacific Ocean. Science 350, 1530–1533 (2015).
Suess, H. E. Radiocarbon concentration in modern wood. Science 122, 415–417 (1955).
Elsig, J. et al. Stable isotope constraints on Holocene carbon cycle changes from an Antarctic ice core. Nature 461, 507–510 (2009).
Rau, G., Riebesell, U. & Wolf-Gladrow, D. A model of photosynthetic 13C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake. Mar. Ecol. Prog. Ser. 133, 275–285 (1996).
Seager, R. et al. Tropical Pacific forcing of North American Medieval megadroughts: Testing the concept with an atmosphere model forced by coral-reconstructed SSTs. J. Clim. 21, 6175–6190 (2008).
Montoya, J. P. & McCarthy, J. J. Isotopic fractionation during nitrate uptake by phytoplankton grown in continuous culture. J. Plankton Res. 17, 439–464 (1995).
Mackey, D. J., O’Sullivan, J. E. O. & Watson, R. J. Iron in the western Pacific: A riverine or hydrothermal source for iron in the Equatorial undercurrent? Deep Sea Res. I Oceanogr. Res. Pap. 49, 877–893 (2002).
Toggweiler, J. R., Druffel, E. R. M., Key, R. M. & Galbraith, E. D. Upwelling in the ocean basins north of the ACC: 2. How cool subantarctic water reaches the surface in the tropics. JGR Oceans 124, 2609–2625 (2019).
Marsay, C. M., Kadko, D., Landing, W. M. & Buck, C. S. Bulk aerosol trace element concentrations and deposition fluxes during the US Geotraces GP15 Pacific Meridional transect. Glob. Biogeochem. Cycles 36, e2021GB007122 (2022).
Rafter, P. A., Sigman, D. M. & Mackey, K. R. M. Recycled iron fuels new production in the eastern equatorial Pacific Ocean. Nat. Commun. 8, 1100 (2017).
Conroy, J. L., Overpeck, J. T., Cole, J. E., Shanahan, T. M. & Steinitz-Kannan, M. Holocene changes in eastern tropical Pacific climate inferred from a Galápagos lake sediment record. Quat. Sci. Rev. 27, 1166–1180 (2008).
Moy, C. M., Seltzer, G. O., Rodbell, D. T. & Anderson, D. M. Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature 420, 162–165 (2002).
Stine, S. Extreme and persistent drought in California and Patagonia during mediaeval time. Nature 369, 546–549 (1994).
Tierney, J. E., Oppo, D. W., Rosenthal, Y., Russell, J. M. & Linsley, B. K. Coordinated hydrological regimes in the Indo-Pacific region during the past two millennia: Currents. Paleoceanography 25, 871 (2010).
Nelson, D. B. & Sachs, J. P. Galápagos hydroclimate of the common era from paired microalgal and mangrove biomarker 2H/1H values. Proc. Natl. Acad. Sci. U.S.A. 113, 3476–3481 (2016).
Lübbecke, J. F. & McPhaden, M. J. Assessing the twenty-first-century shift in ENSO variability in terms of the Bjerknes stability index. J. Clim. 27, 2577–2587 (2014).
Gruber, N. & Sarmiento, J. L. Global patterns of marine nitrogen fixation and denitrification. Glob. Biogeochem. Cycles 11, 235–266 (1997).
Williams, B. & Grottoli, A. G. Recent shoaling of the nutricline and thermocline in the western tropical Pacific: Shoaling nutricline in tropical Pacific. Geophys. Res. Lett. 37, 867 (2010).
Williams, B. & Grottoli, A. G. Stable nitrogen and carbon isotope (δ15N and δ13C) variability in shallow tropical Pacific soft coral and black coral taxa and implications for paleoceanographic reconstructions. Geochim. Cosmochim. Acta 74, 5280–5288 (2010).
Laws, E. A., Popp, B. N., Bidigare, J. R. R., Kennicutt, M. C. & Macko, S. A. Dependence of Phytoplankton Carbon Isotopic Composition on Growth Rate and [COzlsq: Theoretical Considerations and Experimental Results] 8.
Bidigare, R. R. et al. Consistent fractionation of 13C in nature and in the laboratory: Growth-rate effects in some haptophyte algae. Glob. Biogeochem. Cycles 11, 279–292 (1997).
Fry, B. 13C/12C fractionation by marine diatoms. Mar. Ecol. Prog. Ser. 134, 283–294 (1996).
Fry, B. & Wainright, S. Diatom sources of 13C-rich carbon in marine food webs. Mar. Ecol. Prog. Ser. 76, 149–157 (1991).
Horii, S., Takahashi, K., Shiozaki, T., Hashihama, F. & Furuya, K. Stable isotopic evidence for the differential contribution of diazotrophs to the epipelagic grazing food chain in the mid-Pacific Ocean: XXXX. Glob. Ecol. Biogeogr. 27, 1467–1480 (2018).
Schmidt, G. A. et al. Using palaeo-climate comparisons to constrain future projections in CMIP5. Clim. Past 10, 221–250 (2014).
Dieppois, B. et al. ENSO diversity shows robust decadal variations that must be captured for accurate future projections. Commun. Earth Environ. 2, 212 (2021).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Druffel-Rodriguez, K. C. et al. Radiocarbon and stable isotopes in Palmyra corals during the past century. Geochim. Cosmochim. Acta 82, 154–162 (2012).
Zaunbrecher, L. K. et al. Coral records of central tropical Pacific radiocarbon variability during the last millennium: Coral 14C during the last Millennium. Paleoceanography 25, 1788 (2010).
Heaton, T. J. et al. MARINE20—The marine radiocarbon age calibration curve (0–55,000 Cal BP). Radiocarbon. https://doi.org/10.1017/RDC.2020.68 (2020).
Howell, P., Pisias, N. G., Ballance, J., Baughman, J. & Ochs, L. ARAND Time-Series Analysis Software (2006).
Silfer, J. A., Engel, M. H., Macko, S. A. & Jumeau, E. J. Stable carbon isotope analysis of amino acid enantiomers by conventional isotope ratio mass spectrometry and combined gas chromatography/isotope ratio mass spectrometry. Anal. Chem. 63, 370–374 (1991).
McCarthy, M. D., Lehman, J. & Kudela, R. Compound-specific amino acid δ15N patterns in marine algae: Tracer potential for cyanobacterial vs eukaryotic organic nitrogen sources in the ocean. Geochim. Cosmochim. Acta 103, 104–120 (2013).
Bevington, P. R. & Robinson, D. K. Data Reduction and Error Analysis for the Physical Sciences (McGraw-Hill, 2003).
McCarthy, M. D., Benner, R., Lee, C. & Fogel, M. L. Amino acid nitrogen isotopic fractionation patterns as indicators of heterotrophy in plankton, particulate, and dissolved organic matter. Geochim. Cosmochim. Acta 71, 4727–4744 (2007).
Reimer, P. J. et al. The INTCAL20 northern hemisphere radiocarbon age calibration curve (0–55 Cal kBP). Radiocarbon. https://doi.org/10.1017/RDC.2020.41 (2020).
Hogg, A. G. et al. SHCal20 southern hemisphere calibration, 0–55,000 years Cal BP. Radiocarbon. https://doi.org/10.1017/RDC.2020.59 (2020).
Rodbell, D. T. An 15,000-year record of El Ni’o-driven alluviation in southwestern Ecuador. Science 283, 516–520 (1999).
Oppo, D. W., Rosenthal, Y. & Linsley, B. K. 2,000-year-long temperature and hydrology reconstructions from the Indo-Pacific warm pool. Nature 460, 1113–1116 (2009).
Newton, A., Thunell, R. & Stott, L. Climate and hydrographic variability in the Indo-Pacific Warm Pool during the last millennium. Geophys. Res. Lett. 33, L19710 (2006).
Yan, H. et al. A record of the southern oscillation index for the past 2000 years from precipitation proxies. Nat. Geosci. 4, 611–614 (2011).
Somes, C. J. et al. Simulating the global distribution of nitrogen isotopes in the ocean: Nitrogen isotopes in the global ocean. Glob. Biogeochem. Cycles 24, 3767 (2010).
Rafter, P. A., Bagnell, A., Marconi, D. & DeVries, T. Global trends in marine nitrate N isotopes from observations and a neural network-based climatology. Biogeosciences 16, 2617–2633 (2019).
Acknowledgements
None of this work would have been possible without the support and assistance of the NOAA/NURP Hawaii Undersea Research Laboratory. Logistical support for the sample collection was provided by the National Geographic Foundation (CRE 7717-04). The authors acknowledge NSF OCE-0551481 in support of preliminary analyses. Danielle Glynn was supported by a Eugene Cota-Robles Fellowship and a National Science Foundation Graduate Research Fellowships Program (NSF GRFP 1339067). Stephanie Christiansen was instrumental in obtaining the CSIA-AA and mole percent data. They thank Kevin Miles of Cabrillo Community College for assisting in sample preparation. They thank Christina Ravelo, Kelton McMahon, and Christopher Maupin for discussions.
Author information
Authors and Affiliations
Contributions
T.P.G. collected samples, designed the experiment, and wrote the majority of the paper. D.S.G. performed analytical work and contributed to writing the paper. M.D.M. supervised analytical work and assisted in writing the paper. All authors contributed to the interpretation of the data.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guilderson, T.P., Glynn, D.S. & McCarthy, M.D. Tropical decadal variability in nutrient supply and phytoplankton community in the Central Equatorial Pacific during the late Holocene. Sci Rep 14, 4161 (2024). https://doi.org/10.1038/s41598-024-54635-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-54635-3
- Springer Nature Limited