
Abstract
Passive acoustic monitoring has proven to have many advantages for monitoring efforts and research activities. However, there are considerations to be taken into account regarding the placement of autonomous sound recorders. Detection ranges differ among species and in response to variable conditions such as weather or the location of vocalising animals. It is thus important to the success of a research project to understand, with a certain degree of confidence, the distances at which birds might be detected. In two types of forests in Poland, we played back the vocalisations of 31 species of European forest birds exemplifying different singing characteristics. Based on recordings obtained along a 500-m transect, we estimated the probability of detection and maximum detection distance of each vocalisation. We broadcasted the recording at three heights of singing and repeated playbacks three times during the breeding season to evaluate the effect of vegetation growth. Our results revealed that environmental and meteorological factors had a significant influence on both detection probability and maximum detection distances. This work provides comprehensive measurements of detection distance for 31 bird species and can be used to plan passive acoustic monitoring research in Europe, taking into account species traits and individual characteristics of the study area.
Similar content being viewed by others


Introduction
Estimations of population abundance or the composition of species assemblages at a given spatial and temporal scale are invaluable for ecological research but also serve as the starting point for determining the conservation status of populations. Regardless of the purpose for which such data are collected, abundance estimates should reflect, to the greatest possible extent, the actual status of populations. Until recently, the most reliable way to accomplish this was using human experts, but rapid technological development has provided new tools of automated data collection that in many areas can effectively replace traditional methods of monitoring populations1.
Passive acoustic monitoring (PAM) is based on the deployment of autonomous recording units (ARUs) in the field to record soundscapes. PAM has been shown to be a reliable tool for a wide range of research topics and vocalising taxa2. Compared to traditional monitoring methods, ARUs can be easily deployed, even in remote areas, and can operate without human attention for long periods of time. They are fully programmable at various temporal scales and are cost-effective. Most importantly, however, ARUs can collect data in a fully reproducible manner even over very long periods and large spatial scales3. Furthermore, acoustic data can be stored for future comparisons4.
While PAM has many advantages, a few shortcomings remain. To be directly comparable among different projects, PAM recordings must be collected in a standardised way. Despite exponential growth in the use of PAM in recent years, there are very few practical guides on how to deploy devices to accurately investigate areas of interest. The available suggestions focus on detectability or device settings when recording different species (see5,6,7,8) and some recommend measuring sound detection spaces separately in each habitat9. In general, ARUs are deployed based on the subject and aims of the study or monitoring programme at hand, and recommendations regarding the sampling of a given species/assemblage also tend to reflect these different priorities. For this reason, acoustic data from different projects are often not suitable for comparisons as, for instance, they do not contain information about species’ detection probabilities. The biggest problem seems to be the unknown detection distance, which is species-specific and varies depending on habitat. In addition, issues related to ARU spacing have been poorly investigated, such as data duplication or gaps due to ARUs being too close or too far apart, respectively. Signal detection is affected by a variety of biotic (both interspecific and intraspecific) and abiotic factors that influence sound propagation10. Furthermore, the technical parameters of the ARUs’ microphones only give information on the potential range to be covered, which can be modified by the choice of settings even within a single device.
One of the reasons for the lack of precise guidelines is insufficient knowledge of the distances from which ARUs detect different animals, including birds11. Knowledge about maximum detection distances gives some basic information about the possibility of detecting or missing a particular species and is crucial for planning the spatial distribution of ARUs to ensure that they are independent, i.e., record only different individuals of the monitored species. There have been various proposals on factors to consider when determining the range of an ARU, of which one is the effective detection radius, i.e., the distance beyond which as many vocalisations are undetected as detected12. To determine this, however, we need more data on the probability of detecting various species in different habitats, with more accurate distances for different animals or species. With this information, it is possible to provide more precise guidelines on how to deploy equipment to record particular species or animal groups of interest.
In this study, we conducted an extensive range-testing experiment to evaluate the effect of forest habitats, time of season, and height of sound source on the detection probability of acoustic signals of selected European temperate-forest bird species. Our main objective was to provide practical advice for the planning and interpretation of avian bioacoustics monitoring in temperate forest habitats. We selected a set of bird species with differentiated song characteristics—song duration, frequency characteristics, and amplitude—to evaluate how different vocalisation parameters affect detection ranges. We also analysed how environmental factors affect detection distances to make our results more generally applicable and promote their use in monitoring different forest species.
Materials and methods
Study site
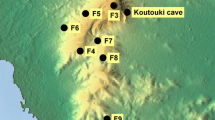
The experiments were carried out in the Zielonka forest near the city of Poznań, in western Poland (N 52.545500, E 17.150639), which is characterised by different types of temperate woodlands. Two transects were designed, separated by ca. 400 m at their beginnings and ca. 900 m at their ends (Fig. 1). The first site was a coniferous forest, mainly consisting of the pine tree Pinus sylvestris. The other was a nearby deciduous forest dominated by common beech Fagus sylvatica and sessile oak Quercus petrea. The average height of trees in the coniferous forest was ca. 22 m, with a 4-m canopy diameter; the mean stand age was 48 years (range 44–53 years). In the deciduous forest, trees were about 25 m in height with a mean canopy diameter of 9 m, and were more differentiated in age (mean 116 years, range 12–257). The pine forest appeared slightly denser due to its smaller canopy size, though both sites are marked as moderately dense forests13. In the beech forest, the leaves gradually developed throughout the research period. The experiments were conducted in 2021. In April the trees had small buds; in May they burst into small leaves that fully developed in June. The coniferous forest needles did not exhibit changes throughout the season, but the underbrush, consisting of taller grasses and ferns, developed in June. Instead, the deciduous forest featured a layer of leaves from the previous year that did not change much throughout the season. The topology of both sites was flat to avoid any influence of the terrain structure on sound propagation. Both sites were located ~ 3 km from the nearest public and tarmac road, so there were no anthropogenic sounds.

Aerial map of the study area. Ten ARUs were placed 50 m apart along two 500-m transects at 4 m height. The speaker at the beginning of each transect was mounted at 3, 6, or 9 m height. The larger scale inset shows a lack of any urban agglomeration close to the site. The map was made using QGIS software and maps provided by the Geoportal service14,15.
Design of transmission experiment
Along each transect, we placed 10 ARUs (Song Meter SM4 with two built-in omnidirectional microphones, Wildlife Acoustics, Maynard, MA, USA), one every 50 m, so that they were located between 50 and 500 m from a loudspeaker (position measured with GPS Garmin GPSMAP 65 s with averaged accuracy ± 3 m). We used built-in omnidirectional microphones with sensitivity − 35 ± 4 dB and signal-to-noise ratio 80 dB, calibrated using sound level calibrator (VOLTCRAFT SLC-100). Each ARU was randomly picked and mounted on a tree trunk at a height of 4 m. Sound files were saved as uncompressed PCM.wav files with a 48 kHz sampling rate and 16-bit resolution. The loudspeaker (JBL XTREME 3, Harman, CA, USA; output power 25W RMS, frequency response range 53.5 Hz–20 kHz) used for broadcasting the recordings was placed at a height of 3, 6, or 9 m at the starting points of both transects. The choice of height for each subsequent broadcast sequence was pseudo-random to prevent, for example, playback from the highest location always occurring earliest in the morning. Broadcasting on the two transects was performed on two consecutive days at the beginning of each month, from April to June, starting one hour after sunrise to avoid significant competition from the dawn chorus. We also measured temperature, humidity, and wind speed during the beginning and end of each broadcast using a Benetech GT8907 (Shenzhen, China) weather meter. During all experimental days, the weather conditions were favourable with no rain and no strong (> 1 m/s) wind. Meteorological data is available in Supplementary Table 1.
Bird vocalisations used for playback
We wanted to examine a wide range of vocalisation characteristics that can affect the probability of detection, such as frequency, amplitude, and duration. For this, we selected songs and territorial calls of 31 European forest bird species (Table 1). The playback began with a series of three pure tones at a frequency of 1000 Hz generated using Avisoft SASLab Pro (version 5.2.14, R. Specht, Berlin, Germany), so that we could easily find the beginning of each playback in the recordings. After this initial sequence, we broadcasted one minute of playback for each species, featuring 2–6 types of vocalisations produced at a species-specific rate. The number of distinctive vocalisations depended on the number of recordings available, and the number of repetitions within the minute-long period was determined by the length of the individual vocalisations. After the songs or territorial calls of each species, we played three pure tones at a frequency of 1000 Hz to mark the end of each species’ playback section. Each pure tone lasted 0.5 s and was separated from the others by ca. 1.5 s of silence. The total duration of the broadcast series was around 45 min.
Recordings were obtained mostly from The Bird Songs of Europe, North Africa, and the Middle East by Schulze and Dingler17. All recordings were first inspected for quality and filtered up to 500 Hz outside the band of the focal species if possible. The amplitudes of the chosen signals were modified in Avisoft SAS Lab Pro to match the real amplitude at which the species sings or calls (based on literature data, our own measurements, or similar species data). As a final check of the amplitude level, we played signals with a fixed setup of the speaker and recorder, and we measured the amplitude of the reproduced sounds. The speaker and decibel meter were placed 2 m from each other (to avoid taking measurements in the near field) on tripods 1.6-m high. All song sequences were reproduced three times from the speaker, and the maximum amplitude of each vocalisation was noted. Measurements were based on fast-time A-weighting using a CHY 650 Sound Level Meter (Ningbo, China). Measurements were taken outdoors in quiet conditions (ambient noise < 42 dB SPL). The average values for each species are presented in Table 1; they are recalculated and shown as the amplitude SPL dBA at a 1-m distance from the source.
Using principal component analysis, we divided all species into groups based on their peak frequency relative to body mass, in order to compare species with similar characteristics with each other and within groups. The body mass used was the mean weight for the male of each species16. The groups included large birds (with a peak frequency of 431–689 Hz), medium (1895–3618 Hz), and small ones (3058–7020 Hz). All parameters are presented in Table 1.
To inform recommendations on the minimum distances between automatic recording devices, we compared the detection distances of individual species with the average sizes of their territories. As reference values for the latter, we adopted the sizes of the territories found in a natural lowland forest, the Bialowieza Forest18. The values were converted from average densities. We used data from a lime-hornbeam forest to compare with our deciduous transect and a pine-bilberry coniferous forest for the coniferous one.
Acoustic data processing
In total, we analysed 5580 sound files, each a 1-min sample of vocalisations of 1 of 31 species, re-recorded at a given distance, transect, height, and month. We excluded 117 recordings (2.1% of all samples) from the deciduous forest (7 in May, 110 in June) that were marked as errors due to recorder issues. We visually and audibly examined these sound files to determine if at least one of the re-recorded vocalisations could be either seen on the spectrogram or heard in each recording. If so, the detections were marked. This was done by one person (DW) with prior knowledge of which species’ vocalisation to search for in each recording. We used Raven Pro 1.6 (Cornell Lab of Ornithology, Ithaca) at default settings, including Hann window with 512 samples, Jet colour map, brightness, and contrast at 50. Settings were occasionally adjusted slightly to analyse difficult recordings.
To cross-check the initial verification, the recordings were also blindly analysed by PS and TO. We encrypted 310 randomly chosen recordings (5.56% of all samples), and all vocalisations seen in the spectrogram or heard in each recording were noted and identified. In the blind trial, the responses were 95% unanimous (295 samples).
Statistical analysis
All statistical analyses were conducted in R19. First, to estimate the detection probability of each species’ vocalisation at each distance to the loudspeaker, we built a generalised linear mixed-effects model (GLMM) with binomial error distribution and probit link function using the ‘glmmTMB’ package20. Our response variable was a value indicating whether a given species’ vocalisation could be detected in a recording (1) or not (0). In the full model, we included the month, distance, the height of loudspeaker placement on an ordinal scale, and wind, humidity, temperature, time of day (minutes after sunrise), vocalisation peak frequency, vocalisation duration, species body mass and sound pressure level (SPL) as covariates scaled between 0 and 1. Transect type (deciduous vs coniferous) was included as fixed factor. Species was entered into the model as a random factor and distance was introduced as a random slope. Second, to answer the question of what factors affect the maximum distance at which a given species’ vocalisation can be detected, we built a GLMM with a truncated generalised Poisson distribution and log-link function to assess main effects. Our dependent variable was the maximum distance at which each species’ vocalisation was detected in each trial. Again, we included ordinally scaled month and the height of loudspeaker placement, with wind, humidity, temperature, time of day (minutes after sunrise), vocalisation peak frequency, vocalisation duration, species body mass and sound pressure level (SPL) as scaled covariates. Species identity was also entered into the model as a random factor. Variables were scaled to improve the model fit. We also tested the data for multicollinearity (both environmental variables and species characteristics), however this was not an issue. For both global models, we performed model selection using the ‘MuMIN’ package21. Among 4096 detection distance and 2045 maximum distance models, we identified the best model sets using Akaike’s information criterion (AIC22, Supplementary Table 2), using a threshold of ΔAIC = 2, and calculated the mean of the selected models to create an averaged model. Diagnostics of model residuals were performed with the help of the ‘DHARMa’ package23. To present the detection probability for each species we conducted logistic regression.
Ethics statement
This study was carried out according to Polish law. We obtained all the necessary permissions to carry out the study: permission of the Regional Directorate of Environment Protection (no. WPN-II.6205.8.2021.JM) and permission of the Marshal of the Wielkopolska Region (no. DR-I.7131.1.2.2021).
Results
Detection ranges: general overview
We detected re-recorded species vocalisations in 1692 (30.3%) out of 5580 audio samples.
All of the best models indicated that the probability of detection of the vocalisations of the 31 species was most strongly affected by their body mass and SPL (Tables 2 and S2). As expected, the probability of detection was higher for larger species with low amplitude territorial calls or songs. Peak frequency and call duration did not have significant effect. Weather conditions also affected whether a vocalisation was detected, with temperature and humidity having a strong positive impact and wind a negative one. Furthermore, the probability of detection was lower at greater distances and for higher speaker placement, although the effect of height was minor. Interestingly, the probability of detecting a vocalisation was higher in the pine forest. Seasonal changes had a negative impact, whereas time of day did not significantly affect detectability.
Similarly, SPL and humidity were also the variables with the greatest effect on the maximum detection distance. Maximum distance rose with increasing humidity. Longer-range detections were noted in the pine forest and at lower broadcasting height. Higher peak frequency shortened the distance, while larger and louder species were detectable at longer distances. The time of day was positively correlated with detection distance, with longer distances achieved later in the day. Among meteorological factors, besides major positive effect of humidity, temperature and wind had a negative effect, although the estimated effect sizes were minor. Best models’ results are listed in Supplementary Table 2.
Differences between pine and beech forest
In general, the detection probabilities of vocalisations were significantly lower (p = 0.022) in the deciduous forest than in the coniferous one (754 vocalisation detected (45% of all detections) vs. 938 (55%) (Fig. 2). Modelling of maximum detection distances revealed significantly greater distances in the coniferous forest (p < 0.001).

Fitted detection probability curves for all species at both transects. Curves were fitted with the generalized linear model method with a 95% confidence interval showed as a shaded area.
Season effect
Of all vocalisations in the coniferous and deciduous forests, 18.6% and 14.0%, respectively, were detected in April, while 18.6% and 17.3% were detected in May and 18.3% and 13.2% in June. The detection rate between months was very consistent at the coniferous site and varied in the deciduous transect.
Throughout the season, the general detection probability was higher in coniferous forests for all groups of birds, with the exception of large species in April and May (Fig. 3).

Changes in detection probability during the season in both habitats for species divided into size-related groups based on the principal component analysis. Curves were fitted with the generalized linear model method with a 95% confidence interval showed as a shaded area.
Effect of singing height
In both transects, least detections were achieved at 9 m height. Also, in both environments there were only minor differences between 3 and 6 m (34.2% vs. 34.7%, respectively).
For each group of birds, the probability of detection was higher in the coniferous forest at almost any height, again with the exception of large birds (Fig. 4). Changes in detection probability for different broadcasting heights during the season can be seen in Supplementary Fig. 1.

Changes in detection probability in both habitats for species divided into size-related groups based on the principal component analysis according to broadcasting height. Curves were fitted with the generalized linear model method with a 95% confidence interval showed as a shaded area.
Species effect
The effects of SPL and peak frequency were consistent across species. For some smaller species, the detection probability dropped rapidly at a certain distance (Fig. 5). The species with the highest probability of detection was the common cuckoo Cuculus canorus, while the species with the lowest was the long-tailed tit Aegithalos caudatus.

Predicted probability of detection of each species at different distances from the speaker. Curves were fitted with generalized linear model method. Each species assigned to a size group based on the principal component analysis is showed as a separate curve.
The common cuckoo and stock dove Columba oenas were the only two species whose vocalisations were detected at a distance of 500 m. Vocalisations of most of the studied species were usually not visually detectable on spectrograms at a distance of 150 m, although they were still audible on recordings even at 200 m. Table 3 presents the detection probability for all species based on logistic regression. The measurements incorporate all of the observations from this study, and in many cases they are shorter than the extreme distances we noted in exceptional cases during the study. Detection probability for each species can be seen in the Supplementary Fig. 2.
Discussion
In this study, we estimated the maximum detection distances that could be obtained from sound propagation experiments using territorial vocalisations of 31 European bird species. We studied the effect of physical properties of the songs/calls, weather, and habitat variables on the detection probability at different distances from a speaker. Our study represents a move towards the standardisation of acoustic surveys, which is important for planning research or monitoring using the PAM approach.
Species
Our results revealed that body mass and SPL are the best predictors of detection probability. The higher the SPL and the larger the species, the longer the distance at which the sound can be heard. According to Brenowitz, only 1/3 of the variation in SPL is attributable to changes in body weight. Therefore, the simplistic assumption that body mass can predict SPL—and thus detection distance—is incorrect, and such approximations must be performed with caution. There are indeed species of similar body mass that relevantly differ in detection probability, such as the common cuckoo and Eurasian sparrowhawk Accipiter nisus. Hence, vocal parameters and body mass should be considered collectively when evaluating detection distance. However, song frequency can, to a certain degree, be predicted by body mass24 and this information could be useful in estimating characteristics of lesser-known species for the purpose of research planning. This factor is highly species-specific, as forest birds generally produce more pure tones, and the frequency of their songs is less variable than in species living in other habitats25. In the forest, the sound frequency for optimal communication is around 1.6–2.5 kHz25, because higher frequencies attenuate at shorter distances26,27. Other effects, such as reverberation, also predominantly influence high frequencies (above 8 kHz), while frequencies below 2 kHz are less affected28,10. Most of the small bird species in our study are characterised by a frequency range oscillating from about 1500 to 8500 Hz, which translates to a maximum detection distance of 150 m. Instead, species characterised by the 100–2500 Hz frequency range were detected at much further distances. This type of information is useful for planning PAM surveys based on the species of interest, as it can be used to set expectations for the detectability and maximum distance possible in optimal environmental conditions. Knowing at least one of the vocal parameters might be helpful in those estimations.
Even though the height effect was substantial for both detection probability and maximum distance, the overall effect sizes were low (Table 2). In terms of PAM planning, the height of singing is not something that can be set by the researcher. Different species prefer different song posts, which are determined only by the opportunities given by the habitat structure. However, knowledge about a species' singing behaviour and environmental characteristics should help in the decision-making stage and in the proper interpretation of the data obtained. For example, the monitoring of low-amplitude species that sing from higher positions above the ground will require the deployment of ARUs at higher densities.
Type of forest
We found that forest type had a major effect on the maximum detection distance and detection probability. Species were detected at farther distances in the coniferous forest than in the deciduous, and more detections were recorded there (55.4% vs. 44.6%). Under differing conditions, the same acoustic signal can experience different distortions; in a forest habitat, the most important types of distortions are amplitude fluctuations and reverberation, the strength of which depends on the forest type28,10. The coniferous forest in our study is characterised by a higher homogeneity of smaller-crowned, more densely packed trees than the deciduous forest, which might be one of the reasons for the consistent difference between the two transects. However, as sound propagation cannot be predicted based on vegetation structure9 and forest types differ in terms of structural properties, the type of forest is only a minor consideration in PAM planning—researchers are much more likely to choose an area of interest based on the species that inhabit it rather than its inherently higher or lower probability of detection.
Limitations
In this study, we were able to determine ranges for audible and spectrogram-visible detections (Fig. 6) that should be considered when planning an analysis of recordings. Depending on the type of detection intended (automatic or manual), such differences in detectability can be an important factor in PAM planning. At a distance of 150 m, we could hear all 31 species, but only 64.52% of them were visible on the spectrogram. Vocalisations of most small birds were still visible at 100 m, and for large species this threshold was about 300–400 m, although they could be heard at much greater distances. This difference is the result of two factors: the effective range of the recorder, which gathered distant sounds that were not loud enough to be visually represented, and masking by other birds, which was likely high in this study due to the short duration (1 min) of the recordings. Masking should not be as much of a problem in long-term acoustic monitoring as there is more likely to eventually be a sound window for the species of interest to fit in. However, in our study, the time regime was kept deliberately short to facilitate analysis. Here, the greatest difference in the maximum distance of visual and audio detection was obtained for the European robin, Erithacus rubecula (29%). This species was frequently masked in audio recordings from greater distances and its vocalisations were difficult to detect visually. Instead, the mean value for all species was 16%.

Comparison of species detection distances. The dashed line is the mean distance (m) at which the species could be heard, while the solid line is the mean distance (m) at which species could be detected visually on a spectrogram.
Researchers should keep in mind that the estimations presented in our study will vary (most likely be shorter) depending on the habitat in question29. Depending on the time of day and environmental factors that influence sound propagation, the maximum communication distance of a bird song can vary by about several dozen metres throughout the day24.
For more detailed guides, species could be organised into groups of families or genera according to the similarity of their vocalisations, since certain call traits, such as frequency, determine the range at which they can be detected11. This methodology could potentially be used for the species whose detection distances have not yet been estimated. If a species of interest is not listed in this work, we suggest comparing its sound parameters with a species from this study that is close to its vocal values.
Recommendations
We suggest the following simple guidelines for planning research with the use of ARUs. If the research is focused on a single species, we recommend measuring its song amplitude directly and performing a field test similar to the one presented here. This will greatly facilitate later interpretation of the data. For more comprehensive studies, we developed the following advice for obtaining the most effective results.
PAM planning will depend on researcher’s approach. Either individual should not be recorded on two devices at the same time, and it can be treated as a spatial point within the recorder’s range, or the individual can be recorded on multiple ARUs while singing from different points at different time, and it should be treated as a point cloud. The second approach will result in deploying the recorders closer together, as varied detectability on each device can provide additional data, e.g. on spatial distribution.
If the species of interest is listed in this study, we propose using Table 3 as a reference to estimate the placement of ARUs. To properly determine the minimum distance between ARUs, researchers should choose a maximum detectability range adjusted to the potential territory size and territory spacing of the chosen species. Decisions about the monitoring setup will then reflect the goals at hand. A first consideration in planning where to deploy ARUs is the size and density of the bird population, as having devices in close proximity might result in recordings of the same individual on several ARUs. For studies of species that possess large home ranges, though, sparsely distributed ARUs may not generate results if a bird of interest will not call or sing in their proximity, away from its central territory. However, this approach could still provide information on whether the species is present there. With a limited number of devices, the decision on where to deploy ARUs will reflect a trade-off between the size of the potential area of a species’ occurrence and the area that can be effectively covered by the devices.
Regarding this point, we have developed a simple formula based on territory size and detectability range to follow while planning PAM for various species (Fig. 7). For research on birds whose territories are smaller than their maximum detectability range, we propose separating recorders with a distance equal to triple the range (3d). This type of monitoring scheme would provide recordings of separate individuals in close proximity to the recorder, while possibly gathering some distant sounds of other individuals as well, particularly if their home ranges partially overlap or are close by. As large species with larger detection ranges often occupy large territories, deciding where to deploy devices will depend on whether the species is territorial and the size of its territories. For birds with a territory larger than their detection range, we propose quadrupling the detection distance for the placement of ARUs (4d). This would ensure the independence of data for territorial birds that occupy areas of, e.g., double the size of their detectability. For those that move throughout very large distances, such as the common cuckoo, it increases the probability that the bird will be recorded on any device. The estimated distance can be adjusted according to a species’ potential range and vocal characteristics.

PAM planning formula based on the size of a species’ territory and its detection range. Grey circles are species’ home ranges with radii (t); coloured circles show detection ranges with radii (d), with the recorders placed at their centres. The distance between recorders is marked as D.
In Table 3 we present mean radii of detectability and territory size for the species examined in this study. However, the home ranges of certain species might vary as some inhabit large areas or are not territorial. For example, the common cuckoo travels a distance of several kilometres throughout the day while the wood pigeon Columba palumbus may defend only the area in close proximity to its nest. Therefore, PAM of species that are difficult to assess in terms of their presence and area usage will need to be flexible and adapted to a given situation and its research needs.
For any species not included here, as well as those that are, researchers should gather information about the frequency and amplitude at which the species of interest is vocalising and, based on that information, estimate its detection radius. Because birds differ in vocal characteristics both between and within species, a reasonable range should be determined based on the broadcast signals (usually songs). This estimate should be compared to the potential size of the species’ territory to determine how much space will need to be covered with ARUs in order to collect information about all individuals in the area.
Finally, the area of interest should also be analysed, as its structure, vegetation cover, and topography may influence the assessed distances in both positive and negative ways. The present study was conducted in neutral, flat terrain; in hillier or more variable terrain we suggest reducing the estimated detection radius to ensure that any negative impact on detection will be mitigated.
Data availability
The recordings and datasets analysed during the current study are available from the corresponding author on reasonable request (Dominika Winiarska, email: dmwiniarska1@gmail.com).
References
Digby, A., Towsey, M., Bell, B. D. & Teal, P. D. A practical comparison of manual and autonomous methods for acoustic monitoring. Methods Ecol. Evol. 4, 675–683. https://doi.org/10.1111/2041-210X.12060 (2013).
Smith, D. G., Truskinger, A., Roe, P. & Watson, D. M. Do acoustically detectable species reflect overall diversity? A case study from Australia’s arid zone. Remote Sens. Ecol. Conserv. 6, 286–300. https://zslpublications.onlinelibrary.wiley.com/doi/10.1002/rse2.173 (2020).
Gibb, R., Browning, E., Glover-Kapfer, P. & Jones, K. E. Emerging opportunities and challenges for passive acoustics in ecological assessment and monitoring. Methods Ecol. Evol. 10, 169–185. https://doi.org/10.1111/2041-210X.13101 (2019).
Bardeli, R. et al. Detecting bird sounds in a complex acoustic environment and application to bioacoustic monitoring. Pattern Recogn. Lett. 31, 1524–1534. https://doi.org/10.1016/j.patrec.2009.09.014 (2010).
Darras, K. Sound detection spaces [WWW document]. Sound detection spaces. http://wwwuser.gwdg.de/~ecosound/index.html (2015) (Accessed 23 Mar 2023).
MacLaren, A. R., Crump, P. S., Royle, J. A. & Forstner, M. R. J. Observer-free experimental evaluation of habitat and distance effects on the detection of anuran and bird vocalizations. Ecol. Evol. 8, 12991–13003. https://doi.org/10.1002/ece3.4752 (2018).
Yip, D. A., Leston, L., Bayne, E. M., Sólymos, P. & Grover, A. Experimentally derived detection distances from audio recordings and human observers enable integrated analysis of point count data. Avian Conserv. Ecol. https://doi.org/10.5751/ACE-00997-120111 (2017).
Yip, D. A. et al. Sound level measurements from audio recordings provide objective distance estimates for distance sampling wildlife populations. Remote Sens. Ecol. Conserv. 6, 301–315. https://doi.org/10.1002/rse2.118 (2020).
Darras, K., Pütz, P., Fahrurrozi, Rembold, K. & Tscharntke, T. Measuring sound detection spaces for acoustic animal sampling and monitoring. Biol. Conserv. 201, 29–37. https://doi.org/10.1016/j.biocon.2016.06.021 (2016).
Richards, D. G. & Wiley, R. H. Reverberations and amplitude fluctuations in the propagation of sound in a forest: Implications for animal communication. Am. Nat. 115, 381–399. https://doi.org/10.1086/283568 (1980).
Darras, K., Furnas, B., Fitriawan, I., Mulyani, Y. & Tscharntke, T. Estimating bird detection distances in sound recordings for standardizing detection ranges and distance sampling. Methods Ecol. Evol. https://doi.org/10.1111/2041-210X.13031 (2018).
Buckland, S. T. Advanced Distance Sampling (Oxford University Press, 2004).
Bank Danych o Lasach. http://www.bdl.lasy.gov.pl (2023). (Accessed 23 Mar 2023).
Geoportal.gov.pl. Główny Urząd Geodezji i Kartografii. Geoportal.gov.pl. (2023). (Accessed 23 Mar 2023).
QGIS.org. QGIS Geographic Information System. QGIS Association. (2023).
Birds of the World. Edited by S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg. Cornell Laboratory of Ornithology, Ithaca, NY, USA. https://birdsoftheworld.org/bow/home (2022).
Schultze, A. & Dingler, K.-H. The Bird Songs of Europe, North Africa and the Middle East. Ample (2007).
Wesołowski, T. et al. Long-Term changes in breeding bird community of a primeval temperate forest: 45 years of censuses in the Białowieża National Park (Poland). Acta Ornithol. https://doi.org/10.3161/00016454AO2022.57.1.005 (2022).
R Core Team. R: A language and environment for statistical computing. https://www.R-project.org/. (R Foundation for Statistical Computing, 2022).
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. The R Journal, 9(2), 378–400. https://journal.r-project.org/archive/2017/RJ-2017-066/RJ-2017-066.pdf (2017).
Barton, K. Mu-MIn: Multi-model inference. R Package Version 0.12.2/r18. https://cran.r-project.org/web/packages/MuMIn/index.html (2009).
Burnham, K. P. & Anderson, D. R. Model Selection and Inference: A Practical Information-Theoretic Approach 2nd edn. (Springer, 2002).
Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R package version 0.4.6. http://florianhartig.github.io/DHARMa/ (2022).
Brenowitz, E. A. The active space of red-winged blackbird song. J. Comp. Physiol. A 147, 511–522. https://doi.org/10.1007/BF00612017 (1982).
Mikula, P. et al. A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecol. Lett. 24, 477–486. https://doi.org/10.1111/ele.13662 (2021).
Morton, E. S. Ecological sources of selection on avian sounds. Am. Nat. 109, 17–34. https://doi.org/10.1086/282971 (1975).
Marten, K. & Marler, P. Sound transmission and its significance for animal vocalization-I. Temperate habitats. Behav. Ecol. Sociobiol. 2, 271–290. https://doi.org/10.1007/BF00299740 (1977).
Wiley, R. H. & Richards, D. G. Adaptations for acoustic communication in birds: Sound transmission and signal detection. In Acoustic Communication in Birds 131–181 (Elsevier, 1982). https://doi.org/10.1016/B978-0-08-092416-8.50014-0.
Priyadarshani, N., Castro, I. & Marsland, S. The impact of environmental factors in birdsong acquisition using automated recorders. Ecol. Evol. 8, 5016–5033. https://doi.org/10.1002/ece3.3889 (2018).
Acknowledgements
We would like to thank Katarzyna Łosak for help in the fieldwork, Grzegorz Neubauer and Michał Budka for their comments and suggestions on the first draft of the manuscript. We gratefully acknowledge the Forest Experimental Institution in Murowana Goślina, Poznań University of Life Sciences, Poland for allowing us to conduct our experiments on the chosen terrain.
Author information
Authors and Affiliations
Contributions
D.W., T.O., and P.S. designed the research; D.W., T.O., P.S. conducted the experiments; D.W., T.O., P.S. conducted the analyses and interpreted the data; D.W. led the writing of the manuscript. All authors critically contributed to the drafts and gave their final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Winiarska, D., Szymański, P. & Osiejuk, T.S. Detection ranges of forest bird vocalisations: guidelines for passive acoustic monitoring. Sci Rep 14, 894 (2024). https://doi.org/10.1038/s41598-024-51297-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-51297-z
- Springer Nature Limited