
Abstract
Short-grain aromatic Joha rice of Assam is a unique class of specialty rice having tremendous potential in domestic and international markets. The poor yielding ability of Assam's Joha rice demands its systematic characterization for an effective breeding program. This study investigates the morphological, molecular and biochemical profiles of twenty popular Joha (aromatic) rice cultivars indigenous to Assam. Distinctiveness, Uniformity and Stability (DUS) characterization of the cultivars revealed polymorphism in thirty-seven traits, establishing distinctiveness for their utilization in breeding programs. Unweighted Neighbor Joining (UNJ) clustering based on usual Euclidean distances for the polymorphic morphological markers grouped the cultivars into three clusters with eight, eleven, and one genotypes. The Joha rice cultivars showed significant differences for all the quantitative traits except for panicle length. The genotypic and phenotypic coefficients of variability (GCV & PCV) were high for grain yield ha−1 (24.62 & 24.85%) and filled grains panicle−1 (23.69 & 25.02%). Mahalanobis D2 analysis revealed three multi-genotypic and four mono-genotypic clusters of the cultivars. The first five principal components explain 85.87% of the variation among the cultivars for the traits under study; filled grain panicle−1 (0.91) and stem thickness (0.55) positively contributed to the first PC. The cultivars' average polyunsaturated fatty acids were 37.9% oleic acid, 39.22% linoleic acid, and 0.5% linolenic acid. Kon Joha 4 and Ronga Joha contained the highest iron (82.88 mg kg−1) and zinc (47.39 mg kg−1), respectively. Kalijeera, Kunkuni Joha, Kon Joha-5, Manimuni Joha and Kon Joha-2 accorded a strong aroma. PCR amplified 174 alleles with a mean value 2.64 across the 66 polymorphic SSR markers. PIC values ranged from 0.091 to 0.698, with an average of 0.326. The highly informative (PIC > 0.50) markers were RM316, RM283, RM585, RM1388, RM3562, RM171, R1M30, RM118, RM11and RM29 for identification of the twenty aromatic rice cultivars. PCR amplification of 27 SSR markers identified 28 unique alleles (97–362 bp) in 13 Joha rice cultivars, which can help their identification/DNA fingerprinting. The UNJ clustering based on Jaccard's coefficients classified the cultivars into three distinct clusters with eight, ten, and two genotypes. Our study revealed the nutritional richness of these specialty Joha rice cultivars and sufficient scope for yield enhancement through their interbreeding to keep quality intact.
Similar content being viewed by others

Introduction
Globally, rice is planted in approximately 162 million ha and 755 million tons of produce are harvested annually1. Ninety per cent of the production and consumption of the world's rice occur in Asia. India has the world's largest area under rice with 44.0 million ha and is the second-largest producer (96.0 million tonnes in 2010), next only to China, contributing 21.5% of global rice production. The FAO2 estimated a 70 per cent more food requirement for over nine billion people expected to inhabit planet Earth by 2050. As the staple food for most Indians, rice's future demand will increase with the growing population, projected at 1.378 billion by 20303. In Assam, rice occupies approximately 70% of the total cropped area, dominating the state's agriculture4. Rice, the primary agricultural GDP source, plays a significant role in the state economy.
Aromatic rice is a significantly small but important rice subgroup that has gained popularity among consumers. The fragrance of scented rice is a crucial feature that increases its popularity in the international market5. Aromatic rice is considered the best quality and fetches a much higher price than non-aromatic rice6. A unique class of fragrant rice grown as winter rice in Assam is very popular and highly valued due to its quality, known as Joha rice in Assam. Joha rice possesses a superfine kernel, unique aroma, better cooking properties, and excellent palatability7. The aromatic accessions from Assam, Manipur and Sikkim belong to the Indica group8. The aroma (fragrance) of Joha rice cultivars is due to the presence of a non-functional betaine aldehyde dehydrogenase 2 (BADH2), which also lowers grain yield9.
Joha rice is generally a poor yielder, late maturing, tall, and susceptible to lodging. It occupies approximately 5 per cent of the Sali rice area, with an average yield of 1–1.5 per ha7. Modern high-yielding varieties have replaced the Joha rice cultivars, depleting this valuable genetic wealth of aromatic and fine-rice genotypes. However, its aroma and unique cooking quality allowed a global market for Joha rice to enter the European market in 2007.
Furthermore, Assam's non-basmati Joha rice cultivars were granted Geographical Indication (GI) tags in 2017 (http://ipindiaservices.gov.in). Information on genetic variability encompassing morphological, biochemical and molecular characterization of Assam's Joha rice is meagre if present. Genetic diversity studies among the aromatic germplasm of Assam would help their improvement through crossbreeding and identification of pre-breeding materials10 and would preserve information on this unique rice class for the future.
Agro-morphological characterization of rice germplasm is fundamental to providing information for plant breeding programmes11. The nutritional status of rice is becoming increasingly important among consumers because of deficiency disorders, necessitating more knowledge regarding the nutritional composition of different rice varieties in Assam. However, the characterization of rice cultivars in terms of composition and physicochemical aspects is relatively scarce. Rice is a leading food crop with a different nutritional status that helps alleviate poverty12. Proximate analysis is an essential parameter for the routine description of foodstuffs. This analysis comprises six fractions: amylose, crude protein, iron, zinc, and fatty acid. The proximate composition, sensory, gelatinization temperature, and alkali digestion analysis of rice help identify nutritionally complete rice varieties13.
Rice showed a genetically greater extent of diversity. DNA-based molecular markers are essential in assessing genetic variation and elucidating gene relationships between species. Molecular markers have been extensively used in crop improvement research throughout the globe as an appropriate and effective tool for addressing biological parameters in agricultural productivity14. Among PCR-based markers, SSR or microsatellite markers are excellent due to their locus of identity, high polymorphism, and multiallelic nature. Moreover, SSR markers are tandemly interspersed repeats throughout the genome that are amplified through primers that flank these regions. Studies to characterize and improve the aromatic rice of Assam are limited, even though this region stores enormous rice diversity. The present study characterized a panel of popular Joha rice cultivars grown in Assam's Brahmaputra and Barak valleys using morphological, biochemical, and molecular markers for their usage in yield enhancement breeding. Most cultivars are popular among the farmers of the Brahmaputra Valley, except Kalijeera and Harinarayan, which are well-liked in the Barak Valley of Assam. Farmers and consumers have particular preferences for these cultivars because of their unique quality; some are believed to have medicinal properties. Barak Valley farmers consume Kalijeera rice to improve health, increase appetite, and reduce hearing problems. The inherently poor yield of Joha rice is quite discouraging for a high return, even if it fetches a high market price, due to rapidly increasing production costs. Many such known cultivars are already out of cultivation in the farmers' field. This situation demands breeders' immediate attention to mainstream these valuable Joha rice cultivars through intensive research for value-addition and yields to sustain their cultivation (Table 1).
Results
Morphological characterization of the cultivars
A total of sixty-two morphological traits characterized the distinctiveness of the twenty Joha rice cultivars. Among the characteristics, the spectrum of variability in the twenty cultivars revealed twenty-two monomorphic (Supplementary Table S2) and thirty-seven polymorphic markers (Table 2; Fig. 1).

Variations in some morphological characteristics of the Joha rice cultivars: (a) ligule shape—split in all cultivars; (b) basal leaf sheath colour; (c) internode colour; (d) stigma colour; (e) panicle secondary branching; (f) lemma palea colour; (g) panicle awns; (h–k) phenol reaction of the grains.
Cluster analysis of the cultivars based on polymorphic DUS characteristics
Clustering by unweighted neighbour-joining (UNJ) of the usual Euclidean distance matrix (Supplementary Table S3) based on thirty-seven polymorphic traits grouped the twenty indigenous aromatic rice cultivars into two multi-genotypic clusters and one mono-genotypic cluster (Fig. 2); the clusters were further subdivided into sub clusters with an unequal cultivar distribution. The eight cultivars, viz., Local Joha, Keteki Joha, Harinarayan, Kon Joha-5, Kon Joha (Moran), Jeera Joha, Kon Joha-3, and Ronga Joha, belonged to G1. At the same time, G2 had eleven cultivars viz., Kon Joha-1, Manimuni Joha, Kon Joha-4, Kalijeera, Kola Joha, Joha (Bihpuria), Soru Joha (Tinsukia), Kon Joha (Bongaigaon), Joha (Golaghat), Kon Joha (Teok) and Kunkuni Joha, whereas G3 had Kon Joha-2 alone.

Hierarchical horizontal clustering of the 20 Joha rice cultivars using Unweighted Neighbour-Joining (UNJ) method based on usual Euclidean distances estimated from 37 polymorphic morphological markers.
Pooled analysis of variance
The pooled ANOVA over the two years (Supplementary Table S4) revealed that the mean squares due to years were significant for fifteen traits, viz., days to first flowering, flag leaf length, flag leaf breadth, flag leaf area, days to 50% flowering, days to maturity, plant height, productive tillers plant−1, filled grains panicle−1, spikelet fertility, 1000-grain weights, biological yield plant−1, grain yield plant−1, harvest index and grain yield kg ha−1. The cultivar differences for all the traits except for panicle length were also evident from highly significant mean squares. The year x cultivar interaction component was substantial for days to first flowering, days to 50% flowering, days to maturity, filled grain panicle−1, and spikelet fertility.
Mean performances of the cultivars
The cultivars' mean performances (Supplementary Table S5) identified 11 top-ranking cultivars for the observed traits (Table 3). The cultivars were Kola Joha for early flowering/maturing, large grains and high biological yield; Soru Joha (Tinsukia) for broad flag leaf, long decorticated grains, wide decorticated grain length/breadth ratio and increased grain yield ha−1; Ronga Joha for high flag leaf area and high grain yield plant−1; Keteki Joha for reduced height and high productive tillers; Kon Joha-5 for high filled grains on panicles and broad grains; Kon Joha-2 for thick stem wide decorticated grains; Kon Joha (Teok) for long flag leaf; Jeera Joha for high spikelet fertility; Kon Joha-3 for long grains; Local Joha for wide grain length/breadth ratio; and Joha (Bihpuria) for high harvest index. All the Joha rice cultivars are typically low to medium tillering and tall.
Genetic variability parameters
Grain yield (kg ha−1) registered the highest range of variation, followed by flag leaf length, filled grains panicle−1, 1000-grain weights, grain length/breadth ratio, grain yield plant−1, productive tiller plant−1, decorticated grain length, stem thickness, and biological yield plant−1 (Table 4). The magnitude of the genotypic and phenotypic coefficients of variability (Table 4) was high (> 20%) for grain yield kg ha−1and filled grains panicle−1. The variability estimates were moderate (10–20%) for decorticated grain length/breadth ratio,1000-grain weights, grain length/breadth ratio, decorticated grain length, grain length, productive tillers plant−1, flag leaf area and stem thickness. The heritability estimates ranged from 37.36 per cent for grain yield plant−1 to 99.99 per cent for flag leaf length (Table 4). Most traits exhibited high heritability values (82.64 to 99.99%); the exceptions were productive tillers, harvest index and biological yield with average estimates (> 40%), and grain yield plant−1 was low heritable. As a per cent of the mean, the genetic advances were high (> 20%) for grain yield ha−1, filled grain panicle−1, decorticated grain length/breadth ratio, 1000-grain weights, grain length/breadth ratio, decorticated grain length, grain length, flag leaf area, stem thickness and productive tillers plant−1 (Table 4).
Genetic divergence among the twenty Joha rice cultivars
The V statistics and the analysis of dispersion (Supplementary Table S6) showed that the mean differences for the pooled effect of the twenty-two characters between the cultivars were highly significant. Mahalanobis distances (D2) distinguished the twenty Joha rice cultivars into seven clusters, showing eight, five and three cultivars in I, IV and II, respectively and the rest four with single genotypes (Table 5).The average intra- and inter-cluster distances (Table 6) showed cluster I to have the maximum intra-cluster space (331.08), followed by IV (307.84) and II (223.91). The inter-cluster D2 values varied from 353.94 units between clusters V and VI to 7303.31 between IV and VI. The most distantly related cluster pair IV-VI was followed by IV-V (5187.67), II-IV (4709.44), VI-VII (4073.37), V-VII (2737.52), III-IV (2323.71) and I-IV (2281.86).
The cluster mean performances for the various traits showed variations among the groups (Table 7). Cluster III registered the earliest days to first and 50 per cent flowering (108.33 & 114.17) and maturity (146.57) and the highest mean performance for stem thickness (4.59 mm), flag leaf breadth (0.84 cm), grain yield plant−1 (14.83 g) and harvest index (36.40%). The highest values of flag leaf length (54.21 cm), flag leaf area (32.42 cm2), panicle length (27.26 cm) and biological yield plant−1 (36.73 g) were evident in cluster IV. Mean filled grain panicle−1 was the highest (252.07) in the solitary cluster VI, along with grain and decorticated grain breadth (2.87 & 2.33 mm), whereas cluster II registered the shortest height (110.57 cm), the highest number of productive tillers (12.60) and the widest grain length/breadth ratio (3.89). Again, the single cultivar Soru Joha (Tinsukia) in cluster VII had the highest means for flag leaf breadth (0.84 cm), spikelet fertility (90.92%), 1000-grain weights (22.50 g), grain/decorticated grain length (9.04/6.97 mm), decorticated grain length-breadth ratio (3.32) and grain yield ha−1 (3011.67 kg). Only eight of the twenty-three traits contributed to the genetic divergence. The contribution towards the total variation was the maximum for flag leaf length (72.11%), followed by decorticated grain length (13.68%), grain length (6.84%), decorticated grain breadth (3.16%), grain yield ha−1 (1.58%) and grain breadth (1.05%).
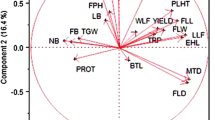
Principal component analysis (PCA) was performed for twenty-two traits of the 20 indigenous Joha rice cultivars (Supplementary Table S7). Principal components (PCs) assume importance when the eigenvalue is greater than one and the PC explains at least 5% of the variation in the data32. Out of twenty-one, only five principal components (PCs) exhibited eigenvalue more than one and explained 85.87% cumulative variability among the traits studied; thus, these five PCs were significant for further explanation. The first five PCs explained 33.23, 25.34, 13.23, 9.11 and 4.95% of the variability among the cultivars for the traits under study. The Scree plot (Fig. 3) showed slight variance after the fifth PC. The traits filled grains panicle−1 (0.91) and stem thickness (0.55) positively contributed to the first PC. In contrast, decorticated grain length (-0.88), grain length (-0.87), grain length/breadth ratio (-0.85), decorticated grain length/breadth ratio (-0.820) and grain yield plant−1 (-0.80) contributed negatively to PC 1. PC 2 accounted for 25.34% of the total variability. The positively related traits were days to 50% flowering (0.88), days to maturity (0.88) and days to first flowering (0.84), while plant height (-0.76), flag leaf length (-0.67) and flag leaf area (-0.65) were negatively related to PC 2. PC 3 contributed 13.23% to the total variability, with grain breadth (0.76), 1000-grain weights (0.60), decorticated grain breadth (0.59), and biological yield plant−1 (0.52) having a positive contribution, while the harvest index (-0.56) and spikelet fertility (-0.55) contributed negatively to PC 3. PC 4 and PC 5 contributed 9.11 and 4.95% of the total variability, respectively. The vector length depends on the character's contribution to the principal component (Fig. 4). Moreover, the vectors' angle reflects the variables' correlation. If the angle between two trait vectors is < 90° (an acute angle), it indicates a positive correlation. The vectors in the first quadrant, viz., days to first/ 50% flowering and maturity, strongly correlated among themselves and loaded on the PC2. At the same time, filled grains panicle−1 loaded on the PC1 had a weak correlation with the above traits. The vectors in the second quadrant, productive tillers, decorticated grain length-breadth ratio, grain length-breadth ratio, decorticated grain length and grain length, were highly correlated variables loaded on PC1. Similarly, the vectors in the third quadrant, grain yield plant−1, panicle length, flag leaf breadth, 1000-grain weights and flag leaf area, were highly correlated variables and loaded on PC1. In the fourth quadrant, stem thickness loaded on PC1 and grain/decorticated grain breadth and plant height correlated to PC2; the latter three vectors were also highly interrelated. If the angle between two traits is > 90° (an obtuse angle), it indicates a negative correlation, while if the grade is equivalent to 90°, it suggests no correlation between the traits. The traits stem thickness, filled grains panicle−1, days to first flowering, grain breadth, plant height and days to 50% flowering were negatively correlated with grain yield plant−1. The cultivar Soru Joha (Tinsukia) projected on the vectors of productive tillers plant−1, decorticated grain length/breadth ratio, grain length/breadth ratio, decorticated grain length, grain length and grain yield plant−1 were close to them, indicating a positive interaction (Fig. 4). Comparing the twenty cultivars, the cultivar Joha (Bihpuria) was superior for flag leaf breadth, 1000-grain weights, biological yield plant−1, flag leaf area, flag leaf length, spikelet fertility, panicle length and grain yield plant−1. Moreover, the cultivars Kon Joha (Teok), Soru Joha (Tinsukia), Ronga Joha, Joha (Golaghat) and Kola Joha also had a positive interaction with those characters. The cultivars with a high positive principal component score for PC 1 (Supplementary Table S8) were Keteki Joha (1.912), Local Joha (1.636), Soru Joha (Tinsukia) (1.273), Ronga Joha (1.219), Kola Joha (0.717) and Joha Bihpuria (0.660).

Scree plot showing Eigen values and percentage of cumulative variability.

Distribution of 20 indigenous Joha rice cultivars and 22 traits across first two components based on PCA.
Biochemical characterization of the Joha rice cultivars
Table 8 shows the biochemical characterization of the twenty Joha rice cultivars based on fatty acid profile, Fe and Zn content, crude protein, amylose, gel consistency, and aroma. A cultivar's mean value was considered desirable for all other biochemical traits except for polyunsaturated fatty acids when it exceeded the cultivars' mean plus the standard deviation. A low mean less than the cultivars' mean minus the standard deviation was desirable for polyunsaturated fatty acids.
Fatty acid profile
Palmitic acid content ranged from 17.98 (Local Joha) to 20.57 per cent (Kon Joha-Teok), averaging 19.01 (Table 8). The stearic acid content was the lowest (0.90%) in Harinarayan and the highest (1.86%) in Kalijeera; the standard was 1.40. The range of oleic acid was from 33.53 per cent in Kunkuni Joha to 41.32 per cent in Joha (Golaghat), with an average of 37.90. The linoleic acid content varied from 36.02 in Joha (Golaghat) to 44.61% in Local Joha, with an average of 39.22. Local Joha recorded the highest linolenic acid content (2.14%), whereas Kola Joha had the lowest estimate (1.06%). The arachidic acid content ranged from 0.28 (Local Joha) to 0.61 per cent (Kon Joha-3), with an average of 0.48.
Iron and Zn content of the cultivars
The iron content in the cultivars varied from 21.33 (Keteki Joha) to 82.88 mg kg−1 in Kon Joha 4, with an average of 43.57 mg kg−1 (Table 8). Kon Joha 4 was followed by Kon Joha-Teok (63.66 mg kg−1), Kon Joha 2 (62.50 mg kg−1), Soru Joha-Tinsukia (61.09 mg kg−1) and Kon Joha 1 (61.00 mg kg−1).
The zinc content ranged from 12.26 (Keteki Joha) to 47.39 mg kg−1 in Ronga Joha, showing an average of 28.43 mg kg−1. The other cultivars having high grain zinc content were Kon Joha-3 (45.32 mg kg−1), Kon Joha-4 (38.25 mg kg−1) and Jeera Joha (37.95 mg kg−1).
Protein content, amylose content, gel consistency, and aroma score of the cultivars
The cultivars' protein content ranged from 7.51 per cent in Ronga Joha to 10.32 per cent in Kon Joha-1, with an average of 9.09 per cent (Table 8). The amylose content varied from 15.20 in Jeera Joha to 24.40% in Harinarayan, with an average of 19.86. The cultivars exhibited two classes of amylose content—medium (20–25%) and low (10–20%). The lowest and the highest gel consistency in the cultivars were 61.50 in Kon Joha-5 and 140.50 mm in Joha (Bihpuria), respectively, with an average of 100.25. Joha-Golaghat (129.50 mm), Kunkuni Joha (123.00 mm), and Manimuni Joha (121.50 mm) followed Joha (Bihpuria). The cultivars viz., Kalijeera, Kunkuni Joha, Kon Joha-5, Manimuni Joha, and Kon Joha-2 showed a strong aroma, and Joha (Bihpuria), Keteki Joha, Kon Joha-3, Kon Joha (Moran), Joha (Golaghat), Kola Joha, Jeera Joha, Kon Joha (Bongaigaon), Local Joha, Harinarayan, Kon Joha-1, Soru Joha (Tinsukia) and Kon Joha-4 registered a mild aroma. Ronga Joha and Kon Joha (Teok) possess a light scent.
Molecular characterization
Among the seventy-one SSR markers, sixty-six showed polymorphisms. The analysis excluded markers with monomorphic banding patterns. Table 9 summarizes the results on twenty aromatic rice cultivars using the polymorphic SSR loci. Figure 5 depict representative gel pictures of the PCR products.

Representative gel pictures showing the PCR products.
The 66 polymorphic SSR loci amplified a total of 174 alleles (Supplementary Table S9). The allelic richness per locus was 2 to 4, with an average of 2.64 alleles. Among the polymorphic markers, 30 produced two alleles each, 30 produced three alleles each, and 6 generated four alleles. The markers RM283, RM118, RM316, RM29, RM585, and RM26063 amplified the maximum number of alleles. The results revealed that all the markers showed distinct polymorphisms among the cultivars studied, indicating the robust nature of microsatellites revealing polymorphisms.
Among the sixty-six markers, the highest major allele frequency was 0.950 in five (RM237, RM215, RM6641, RM3481 & RM21), followed by 0.900 in nine and 0.875 in two markers (Table 9).The lowest major allele frequency was 0.300 for RM316. The mean value of major allele frequency was 0.727, ranging from 0.300 to 0.950 among the sixty-six markers scored against the twenty Joha rice cultivars. The number of effective alleles ranged from 1.105 (RM237) to 3.922 (RM316), with a mean value of 1.759. Shannon's information index ranged from 0.199 (RM237) to 1.376 (RM316), with a mean value of 0.640. The gene diversity varied from 0.095 to 0.745, with a mean value of 0.377. RM316 showed the highest genetic diversity (0.745), followed by RM283 (0.715) and RM1388 (0.645). A set of four markers, viz., RM271, RM279, RM7434, and RM501, represented heterozygosity among the sixty-six SSRs. All four SSR markers showed the same value, 0.050, with a mean value of 0.003. The rest markers showed no heterozygosity. The polymorphism information content (PIC) values, a reflection of the allelic diversity and frequency among the genotypes, also varied from one locus to another (Table 9). The PIC values ranged from 0.091 to 0.698, with an average of 0.326, indicating that only some SSR markers were highly informative. The highest PIC value of 0.698 was obtained for RM316, followed by RM283 (0.665), RM585 (0.573), RM1388 (0.572), RM3562 (0.559), RM171 (0.559), R1M30 (0.534), RM118 (0.531), RM11 (0.513) and RM29 (0.509), suggesting that these ten markers were highly informative (PIC > 0.50). Among the remaining polymorphic SSR markers, thirty were informative (0.50 < PIC > 0.25), and twenty-six were slightly informative (PIC < 0.25).
An allele observed in only one of the 20 Joha rice cultivars was considered unique. Twenty-seven of the 66 polymorphic SSR loci detected 28 private alleles (97–362 bp) specific to 13 cultivars, with an average of 1.04 per locus. RM3395 amplified a maximum of two unique alleles, followed by the others with single alleles (Table 10). The cultivar Kon Joha-5 had nine unique alleles, followed by Kon Joha-4 (4), Kon Joha-2 (3), Ronga Joha and Joha (Bihpuria), each with two alleles. Jeera Joha, Kon Joha (Moran), Kola Joha, Harinarayan, Kalijeera, Keteki Joha, Kon Joha-3 and Kon Joha (Bongaigaon) showed single private alleles.
Molecular diversity among the aromatic rice cultivars
An unweighted neighbour-joining (UNJ) cluster analysis based on Jaccard's dissimilarity coefficients (Supplementary Table S10) resolved the phylogenetic relationships among the Joha rice cultivars collected from different parts of Assam. The UNJ cluster diagram showed three major clusters (G1, G2 & G3) with additional sub-clusters (Fig. 6); the dendrogram revealed that the cultivars derived from a genetically similar type clustered together. Cluster I comprised eight cultivars, whereas cluster II had ten cultivars, forming the most significant group. Cluster III had only two cultivars (Local Joha & Kunkuni Joha). Cluster I included Kon Joha-3, Jeera Joha, Kon Joha-4, Keteki Joha, Joha (Bihpuria), Joha (Golaghat), Kon Joha-5, and Kon Joha-2. Kon Joha (Bongaigaon), Kon Joha (Teok), Soru Joha (Tinsukia), Ronga Joha, Kola Joha, Kon Joha (Moran), Kalijeera, Kon Joha-1, Manimuni Joha and Harinarayan belonged to cluster II. Cluster I was subdivided into two groups, IA and IB, consisting of six and two cultivars, respectively. Cluster II had two sub-clusters, IIA and IIB, composed of nine and one cultivars, respectively.

Hierarchical horizontal clustering of the 20 Joha rice cultivars using Unweighted Neighbour-Joining (UNJ) method based on Jaccard’s coefficients of similarity estimated from 66 polymorphic SSR markers.
Discussion
Qualitative characteristics are the morphological markers for identifying rice landraces because environmental changes least influence these traits33. The thirty-seven stable morphological characteristics would serve as reliable morphological markers for identifying the twenty Joha rice cultivars, corroborating earlier studies34. A substantial amount of variability within this specialty class of rice was evident from the observed clustering pattern based on the morphological markers, which agrees with Mondal et al.35.
Significant cultivar differences in aromatic rice were also reported in earlier studies [36 &37]. The phenotypic expression of most yield and contributing traits differed significantly in the 2018 and 2019 crops; the 17 June planted crop in 2019 exhibited higher mean performances than that of the 11 July planted crop in 2018. Delaying the sowing time decreased the days to flowering and maturity for most cultivars. A similar observation was reported by Song et al.38 for days to heading reduced in different rice cultivars due to delayed sowing. Nahar et al.39 observed a significant decrease in filled grain production consequent to delayed transplanting, attributed to low temperature at anthesis and primordial spikelet formation. Khalifa40 noted the early date of sowing as the best time for maximizing morpho-physiological traits such as tillering, panicle initiation, chlorophyll content, leaf area index, sink capacity, panicle length, panicle number, and grain yield. Delayed sowing significantly reduces the number of filled grains, panicles, and test weight, finally lowering rice cultivars' grain yield41. Nevertheless, a significant year x cultivar interaction component for days to flowering and maturity, filled grain panicle−1, and spikelet fertility suggested variation in adaptive traits among the Joha rice cultivars. Panwar et al.42 also reported substantial year x cultivar interactions for days to 50% flowering and days to maturity.
The low tillering habit of the aromatic rice cultivars was also supported by Singh et al.43. The considerable genetic variability for yield components observed in the present study was also similar to the findings of Singh et al.43. The variations in the grain characteristics of the Joha rice cultivars were consistent with the findings of Singh et al.43 and Semwal et al.44. The findings of Bajpai and Singh45 further corroborated the present results on grain physical quality characteristics. The phenotypic variations for all the traits except productive tillers plant−1 were mainly determined by the genotypes, indicating the effectiveness of phenotype-based selection. These findings were in line with Karim et al.46. Earlier reports also support the present results on broad sense heritability for flowering/maturity duration, flag leaf and grain characteristics. Chavan et al.47 obtained high heritability for days to 50% flowering (97.5%), plant height (99.5%), filled grains panicle−1 (99.7%), test weight (96.8%) and kernel length (98.7%) in aromatic rice. Debsharma et al.48 also recorded high heritability for days to 50% flowering, ranging from 97.9 to 99.4% in rice genotypes tested over three locations. Similarly, Akshay et al.49 reported high heritability in rice for days to 50% flowering (96.1%), plant height (98.6%), grains panicle−1 (97.8%), 1000-grain weights (99.35), length (98.1%), width (99.4%) and length/breadth ratio (98.7%) of grains. In transplanted aman rice, Faysal et al.50 found high heritability for days to 50% flowering (98.46%), plant height (99.26%), flag leaf length (99.83%), and 1000-grain weights (99.49%).
The traits excluding days to first/50 per cent flowering and maturity, flag leaf breadth, and spikelet fertility exhibited high heritability in conjunction with high to moderate genetic advance, indicating the most likely role of additive gene action and effectiveness of simple selection for the traits. High heritability and low genetic advance for days to 50% flowering agreed with Chaurasia et al.51. Plant height registered high heritability and moderate genetic advancement in conformity with the findings of Chaurasia et al.51. Moderate heritability and high genetic advance for productive tillers plant−1 were consistent with Jaiswal et al.52. Filled grains panicle−1 exhibited a high heritability concomitant with high genetic advance, in agreement with the results of Hasib et al.53. High heritability in concurrence with high genetic advance for 1000-grain weights was in accordance with the findings of Nandan et al.54. The grain quality traits viz., grain length, breadth and length-breadth ratio, decorticated grain length, breadth, and length-breadth ratio registered high heritability coupled with the high genetic advance in consonance with the findings of Jaiswal et al.52 for grain length, grain breadth and length-breadth ratio. A low heritability coupled with moderate genetic advance for grain yield plant−1 was in agreement with Adjah et al.55.
Mahalanobis distance-based clustering pattern of the twenty Joha rice cultivars into seven groups confirmed the quantum of diversity present in the indigenous aromatic rice of Assam and offered scope for its exploitation through breeding for yield improvement. Previous studies reported different numbers of clusters in fragrant rice, e.g., six by Allam et al.56 and five groups by Patel et al.57. Prasad et al.58 obtained 20 Euclidean distance-based clusters in 208 Indian aromatic rice accessions, with 57 genotypes in the largest and single genotypes in six groups. Barhate et al.59 used Mahalanobis D2 statistics to classify 45 aromatic rice genotypes into 10 clusters; five had single genotypes. Netam et al.60 classified 40 scented rice genotypes into five groups, the largest having 29 and two with single genotypes. In the present study, cluster pairs viz., IV-VI, IV-V, II-IV, VI-VII, V-VI, and III-IV as widely divergent, and thus, hybridization between parents from these contrasting clusters would likely produce a broad spectrum of variability and transgressive segregations with high heterotic effects, also suggested by Allam et al.56 and Patel et al.57. Flag leaf length had the highest contribution to total divergence, suggesting the scope for grain yield enhancement through crossbreeding among the aromatic rice cultivars. Since aromatic rice cultivars have the poor combining ability, crossbreeding with non-aromatic varieties would decrease aroma and quality61. The rice flag leaf is the main photosynthetic organ crucial in grain yield62. The morphological variation in flag leaves directly affects the population structure, light distribution, and energy utilization and is, therefore, an essential target trait in breeding super high-yield hybrid rice63. Rice breeders emphasize flag leaf characteristics in selecting the ideal rice phenotype. Rice flag leaves are significant functional leaves for grain filling, and their photosynthesis contributes more than half of all carbohydrates in rice seeds64. With its two main components, the flag leaf area is essential in determining its photosynthesis capacity and is influenced by multiple QTLs and their interactions with the environment62. Among the other traits, the aromatic rice cultivars’ decorticated grain and grain length significantly varied. These results agreed with the findings of Allam et al.56 for decorticated grain length and Singh et al.65 for grain length.
The study of many morphological characters in germplasm is vital for the assessment of the differences among populations as well as for the examination of their breeding potential. Plant breeders often measure many variables, some of which may not be of sufficient discriminatory power for germplasm evaluation, characterization and management. In such cases, principal component analysis (PCA) may reveal the patterns and eliminate redundancy in data sets. The PCA, or canonical root analysis, is a multivariate statistical technique to simplify and analyze the relationships among an extensive collection of variables in terms of a relatively small group of components without losing any essential information of the original data set. PCA is a powerful tool to identify the minimum number of components, explain the maximum variability out of the total variability66 and rank genotypes based on PC scores. The cumulative variance of 85.87% by the first five axes with an Eigenvalue > 1.00 indicates that the identified traits significantly influenced the cultivars' phenotype and could effectively be used for selection among them. These results corroborated Lakshmi et al.67. Burman et al.68 reported that four principal components (PCs) exhibited eigenvalues of more than 1.00 and explained 81.62% variability. Ahmed et al.69 showed that the first five components with vector values > 1.00 contributed 82.90% of the total variations in 31 rice germplasm lines. Pachauri et al.70 studied one hundred twenty-four rice germplasm accessions based on nineteen morphological and eleven agronomic traits. From their studies, PC1 expressed 37.12% variability, while PC2, PC3 and PC4 recorded 13.56, 11.04 and 10.76% variability, respectively, and traits such as the number of effective tillers, 100-grain weights were the principal discriminatory traits. PCA helps us identify the characteristics that significantly impact the phenotype of different rice landraces, which is very important in the selection procedure of the breeding program. The biplot analysis showed the relationships between the morphological traits among the tested genotypes. Acute angles were apparent between productive tillers, decorticated grain length/breadth ratio, grain length/breadth ratio, grain yield plant−1, panicle length, flag leaf breadth, 1000-grain weights, and flag leaf area; the selection of these traits would significantly contribute to Joha rice improvement. Increased grain yield is associated with 1000-grain weights71 and long panicle lengths with many filled grains72. The seed size, such as seed length and breadth, also significantly increases the grain yield plant−172. The traits influencing PC 1 were flag leaf breadth, 1000-grain weights, flag leaf area, filled grains panicle−1, stem thickness, decorticated grain length/breadth ratio, grain length/breadth ratio and grain yield plant−1. These results also support the GCV estimates for flag leaf length, filled grains panicle−1, 1000-grain weights, decorticated grain length, stem thickness and grain length/breadth ratio; the first three traits, along with stem thickness, also corroborated Mahalanobis distance-based divergence. The cultivars Soru Joha (Tinsukia), Ronga Joha, Joha (Bihpuria) and Kola Joha had a high principal component score for PC 1. Based on the relationship of traits and cultivars to PC 1, the cultivars Joha (Bihpuria) and Soru Joha (Tinsukia) would serve as parents for the above characteristics for breeding improved Joha rice.
Fatty acids are vital components of food and human health. Fatty acids are the major constituents of the cell membrane structure and play important biological, structural, and functional roles in the human body73. They act as modulators of gene transcription, cytokine precursors, and energy sources in complex interconnected systems74 by producing a vast ATP quantity during their metabolism73. The role of dietary fatty acids in human health is strongly evident in their influence on cardiovascular disease and mental health74. In addition, rice is a dietary consumption; rice fats have unique health benefits75. In the present investigation, oleic, linoleic, and palmitic acids were the primary fatty acids, and stearic, linolenic, and arachidic acids were minor in the aromatic rice cultivars. Palmitic, stearic, and arachidic acids are saturated fatty acids in rice bran, increasing health risks such as atherosclerosis, and associated with a heart attack76. Linoleic acid is absorbed as a predominant unsaturated fatty acid, followed by oleic and linolenic acid. High contents of polyunsaturated fatty acids are desirable for human health, as their consumption minimizes the risk of heart-related diseases77. The mean polyunsaturated fatty acid (PUFA) contents of the aromatic cultivars were 37.9% for oleic acid, 39.22% for linoleic acid, and 0.5% for linolenic acid, whereas the contents of saturated fatty acids (SFAs) were 1.40% for stearic acid and 19.01% for palmitic acid. These estimates were comparable to or even better than the values of 38.4% oleic acid, 34.4% linoleic acid, 2.2% α-linolenic acid of PUFA, and 2.9 and 21.5% of stearic acid and palmitic acid of SFA, respectively78. The present results were also comparable with those reported by Resurreccion and Juliano79.
Similarly, the variations in the fatty acid profile of the present study proved better in having lower maximum limits for SFA and higher maximum limits for linoleic and linolenic acid than those reported by Goffman et al.80, who obtained 13.9–22.1% for palmitic, 1.5–2.7% for stearic, 35.9–49.2% for oleic, 27.3–41.0% for linoleic and 1.0–1.9% for linolenic acid in rice bran. Stearic acid and arachidic acid were present in trace amounts in all the studied aromatic rice cultivars. Comparatively, the fatty acid profile of Local Joha was better than that of the remaining cultivars, as it possessed a high level of linoleic and linolenic acid and low saturated fatty acid content. In general, the fatty acid profile of Joha rice cultivars qualifies for the extraction of quality bran oil for consumption.
Iron and zinc are crucial in numerous metabolic processes in the human body. Inadequacies of zinc and iron in the human diet are associated with growth retardation, physical and cognitive impairment, anaemia, loss of immunity, vulnerability to infection, abnormal pregnancy and neuropsychological disorders81. Although improved rice varieties and crop management practices contributed to a two-fold increase in rice production in the past few decades82, breeding for high-yielding, quality rice is crucial to meet energy needs and ensure nutritional health in developing countries83. Fe and Zn are essential micronutrients in cell development and gene expression84,85. Iron and zinc deficiency is a severe nutritional problem for humans and is particularly prevalent among children and pregnant women, especially in developing countries. As identified in the current study, Joha rice cultivars with very high iron and zinc contents in brown rice can help iron and zinc biofortification through conventional breeding or biotechnology-based approaches. Increasing the iron and zinc content and bioavailability in rice grains can benefit the human population, especially in developing countries. Substantial variations in brown rice's iron and zinc contents agreed with Chowdhury et al.86. A wide variation in iron and zinc contents of dehusked rice grains was evident among the Indian rice cultivars87. The range was between 5.1—441.5 µg/g (mean 67.8 µg/g) for iron and 2.12—39.4 µg/g (mean 23.8 µg/g) for zinc. The brown rice iron and zinc contents varied between 6.2—71.6 ppm and 26.2—67.3 ppm, respectively, in 126 rice accessions88. Vanlalsanga et al.82 also reported iron and zinc content in dehusked rice ranging from 11.42–215.62 ppm and 17.98–75.8 ppm, respectively in northeast Indian rice landraces. Brown rice has higher Zn and Fe contents than polished rice89,90,91. Therefore, emphasis should be given more to pre-breeding for increasing Zn and Fe contents in the polished rice, as the % loss during polishing depends on the degree and duration of polishing92 as well as location and variety93.The variation in protein content was in agreement with Banerjee et al.94, who reported 4.91 to 12.08% protein in 258 diverse rice landraces with a mean of 6.63 percent. Bajpai and Singh45 also noted low to medium amylose content. In aromatic rice, Semwal et al.44 also observed variation in aroma and accordingly classified the genotypes.
The number of alleles per locus (2.64) obtained in the present study was comparable with earlier reports by Shah et al.95, who reported 2.6, 2.75, and 2.3 alleles per locus, respectively. The mean allele number (2.64) obtained in the present study was higher than that of Meti et al.96, who detected 2.08 alleles per locus using 48 traditional indigenous aromatic rice germplasm grown under the eastern part of India through 12 polymorphic SSR loci. Prasad et al.58 obtained 82 alleles amplified by 27 polymorphic SSR markers, averaging 3.04 per locus in 208 aromatic rice genotypes of India. In contrast, the mean alleles (2.64) detected were markedly lower than the average number of alleles reported in previous diversity studies by Rahman et al.97, who obtained an average of 4.4 and 4.18 alleles per locus, respectively. The variability in the number of alleles detected per locus might be due to diverse genotypes and the selection of different SSR primers with scorable alleles. Similarly, Sajib et al.98 reported a significant allele frequency ranging from 0.41 to 0.91; Shah et al.95 noted a range of 0.425 to 0.975 with an average of 0.647, and Kumar et al.99 observed it to vary from 0.510 to 0.970, averaging 0.74. More alleles generated by SSR markers suggest this marker system's usefulness for detecting genetic polymorphisms. Aljumaili et al.100 detected 1.48 effective alleles per SSR locus among 53 rice cultivars.
In contrast, the effective allele number detected in the present study was lower than the average number of effective alleles (5.51) reported by Yelome et al.101 among West African rice accessions. Aljumaili et al.100 reported a similar Shannon's informative index by evaluating fifty-three aromatic rice accessions using 32 SSR markers, and they obtained a mean value of 0.580. The high value of Shannon's information index indicated the presence of high genetic diversity in the rice germplasm under consideration102. In contrast, Shah et al.95 recorded an average gene diversity of 0.448, ranging from 0.049 to 0.664, whereas Kumar et al.99 reported gene diversity ranging from 0.045 to 0.588 with a mean of 0.340. Similarly, the low level of observed heterozygosity, as also reported by Yelome et al.101, could be due to the autogamous mode of reproduction in rice. The ten highly informative markers detected in our study could be used to identify the twenty aromatic rice cultivars. The polymorphism detected in the present study was consistent with the reported mean PIC values in previous works98. However, Nadia et al.103 said an average PIC value of 0.84, markedly higher than the present average PIC value. Sufficient polymorphism by the 66 SSR markers among the twenty indigenous Joha rice cultivars justifies their proper classification and use in the genetic improvement programme based on the extent of genetic variation for desirable alleles. Our study identified 28 unique alleles specific to the 13 Joha rice cultivars. Shamim et al.104 detected 79 private alleles at 28 SSR loci in 16 locally adapted rice varieties and emphasized their importance in rice breeding. Prasad et al.58 considered a genotype-specific SSR locus amplifying a distinct band as unique or less frequent and detected four of 27 polymorphic markers amplified in 13 aromatic rice accessions. The unique SSR alleles represent a rich source of genetic diversity and diagnostic tools in aromatic rice breeding.
The Jaccard’s coefficients of similarity among the 20 Joha rice cultivars ranged from 0.24 between Kon Joha-1 and Manimuni Joha to 0.78 between Kon Joha-5 and Joha-Golaghat, with an average of 0.55, suggesting diverse nature of the genotypes under study. Similar to the present clustering pattern, Meti et al.96 obtained two major clusters for 48 aromatic rice genotypes from Odisha using 12 SSR markers at 49 per cent genetic similarity. Shah et al.95 effectively differentiated the basmati cultivars from non-basmati cultivars based on cluster analysis with 24 microsatellite loci, classifying 40 rice cultivars into three groups. Islam et al.105 used phylogenetic and model-based population structure analyses and classified 113 aromatic rice germplasm into three groups. Thus, SSR markers provided an adequate resolution to discriminate between aromatic rice accessions, and they could serve as a potential tool in identifying and characterizing genetically distant accessions from various sources. The microsatellite assays generated genotype-specific alleles in some of the cultivars evaluated for DNA fingerprints for cultivar identification and differentiation of aromatic rice. DNA fingerprints would be enormously helpful for establishing and defending proprietary rights and maintaining cultivar purity.
Conclusion
Morpho-molecular and biochemical profiling of a panel of Assam’s popular indigenous Joha rice cultivars has been a step forward for exploiting variability in this unique rice class to improve its inherently low-yield potential through breeding. Our study revealed that the Joha rice cultivars are highly diverse in yield and quality traits. Recombination breeding among the trait-specific genotypes such as the early maturing Kola Joha with large grains and high biological yield, Soru Joha (Tinsukia) with high grain yield ha−1, short-statured Keteki Joha with high productive tillers, Kon Joha-5 with more filled grains and Joha (Bihpuria) with high harvest index would provide a broad genetic base for aromatic Joha rice improvement programs. The low to high degree of dissimilarity among the accessions suggests the high molecular level diversity among the aromatic rice cultivars and their possible utilization in breeding programs to develop elite aromatic rice varieties. The unique alleles in 13 Joha cultivars are a rich source of genetic diversity to help marker-based identification/differentiation of aromatic rice cultivars and maintain this high-quality product's integrity to benefit farmers and consumers. The Joha rice cultivar Soru Joha (Tinsukia), with the highest yield (3012 kg ha−1), high spikelet fertility (90.9%), and high Fe content (61.09 mg kg−1), could serve as an immediate resource for mainstreaming. The Joha rice cultivar fatty acid profile qualified to extract quality bran oil for consumption. Our study opened the scope for value addition through nutritional profiling and yield enhancement through crossbreeding within this specialty rice class without compromising inherent quality characteristics. High-yielding nutri-rich Joha rice would encourage farmers’ adoption of wide-scale cultivation and increase farm income. At the same time, these valuable rice germplasm need to be collected, preserved, characterized, genetically enhanced, and documented in the context of intellectual property rights (IPR). Studying the medicinal properties of the Joha rice cultivars is another vital area of research.
Methods
Phenotypic characterization
The experiments were carried out during the Sali season of 2018 and 2019 at the Instructional-cum-Research (ICR) Farm, Assam Agricultural University. All molecular work, including DNA extraction, PCR, and gel electrophoresis, was performed in the Mutation Breeding Section-I Laboratory of Nuclear Agriculture and Biotechnology Division (NA&BTD), Bhabha Atomic Research Centre, Trombay. The field experimental site is located at 26°45/north latitude and 94°12/east longitude and has an elevation of 86.6 m above the mean sea level. The soils of the experimental site belong to the order Inceptisols with sandy loam texture and pH 4.8. The status of organic carbon, available nitrogen and phosphorus was medium, and available potassium was low. The growing situation was shallow land with a maximum water depth of 30 cm during the peak monsoon.
Twenty indigenous scented (Joha) rice cultivars collected from different agro-climatic zones of Assam (Table 1) were grown in a randomized complete block design with three replications. The seedlings' age was 30 days at transplanting in the main field. Each genotype constituted ten rows of 2.5 m long spaced 20 cm apart with one seedling per hill. A fertilizer dose of 60 kg N, 20 kg P2O5, and 40 kg K2O was applied as per the Sali rice recommendation for Assam. The standard agronomic practices recommended for the state of Assam were adopted in both experiments. Observations were recorded according to the National Test Guidelines for DUS test in rice developed by the Directorate of Rice Research, Hyderabad15. The yield-attributing traits were based on five random plants per replication, while days to flowering and maturity were recorded per plot. Additional data were recorded on flag leaf length, breadth and area, days to first flowering, plant height (cm), spikelet fertility (%),biological yield plant−1 (g), harvest index (%), grain yield plant−1 (g), grain yield ha−1 (kg), protein content (%), iron (Fe) and zinc (Zn) content (mg kg−1), fatty acid profile in rice bran.
Estimation of total protein
Nitrogen was estimated in the samples of polished rice of the selected accessions by the modified Micro-Kjeldahl method16. The percentage of nitrogen was multiplied by the conversion factor of 5.9517 to estimate the total protein content. About 0.5 g of rice flour was digested at 400 °C in the presence of concentrated H2SO4 and a mixture of K2SO4 and CuSO4, followed by distillation using 4% boric acid and 40% NaOH solution. The distilled samples were titrated against the 0.1 N sulphuric acid until the first pink colour appeared at the last point. The titer value was used to calculate the per cent nitrogen.
Estimation of iron (Fe) and zinc (Zn)
The seeds harvested from the 20 selected cultivars were used for the zinc and iron estimation. The samples were accurately weighed (0.5 g each) and placed in a 250 ml digestion tube with nitric acid. To each sample, 5 ml of 65% HNO3 was added and boiled gently over a digester (90 °C) for 1–2 h. or until obtaining a clear solution. Subsequently, 2.5 ml of 65% HNO3 was added, and the tubes were further heated until total digestion18. During digestion, the tubes' inner walls were washed with 2 ml of deionized water to avoid loss of the samples. The samples were then filtered using Whatman No. 42 (2.5 μm particle retention) filter papers, and the final volume was made up to 25 ml by adding sufficient deionized water. Fe and Zn standard solutions were prepared in deionized water. The signal of the blank solution was recorded in duplicate. The signals of the standard solutions (in duplicate) were taken using the lamp corresponding to each element. The calibration curves for Fe and Zn were prepared after subtracting the blank from the recorded signals. The Fe/Zn solution was absorbed using the respective elements. The concentrations of Fe and Zn were calculated from the Fe and Zn calibration curves, respectively.
Fatty acid profiling in rice bran
Fatty acid estimation of each rice cultivar was done in duplicates by Gas Chromatography technique (Shimadzu, Kyoto, Japan) at the Nuclear Agriculture and Biotechnology Division of Bhabha Atomic Research Centre, Trombay. After the hulling process, decorticated grains (brown rice) of all 20 rice cultivars were used for milling up to 5% (approx.) by using a McGill No. 2 miller (Rapsilver Supply Co. Inc., Brookshire, TX). After milling every sample, their bran was collected into small, stripped polythene and adequately labeled. The rice bran of each rice cultivar was stored at 4 °C to prevent the harmful activity of the lipase enzyme. About 200 mg of rice bran of each genotype was taken in a 50 ml glass test tube. One ml each of methanol (analytical grade) and 0.5 M sodium methoxide (analytical grade) was added to the tube. The tubes were shaken thoroughly by vortex and kept for 20 min at room temperature. Then, all the tubes were kept in a water bath at 500 °C for 1 h and then for 10 min at room temperature for cooling. Two ml each of HPLC-grade petroleum ether and deionized water were added to each tube, vortexed properly, and kept for one hour of phase separation. The supernatant was extracted from each tube using a 1 ml micro-pipette and taken in 1.8 ml clear GC vials for analyzing the samples by gas chromatography (GC SOLUTION, Shimadzu, Kyoto, Japan). The fatty acid concentration was recorded by normalizing peak areas using GC SOLUTION software (Shimadzu, Kyoto, Japan) and converted to a percentage. For further analysis, fatty acid proportional contents were arcsine transformed according to Sokal and Rohlf19.
Genomic DNA extraction and purification
Cultivars' seeds were grown in a growth chamber, maintaining a temperature of 30 °C, 10 h of light, and 85% relative humidity. The leaves were harvested in liquid nitrogen for DNA isolation at the three-leaf stage,. The genomic DNA was isolated by the cetyl trimethyl ammonium bromide (CTAB) method20. The concentration and quality of genomic DNA were determined by measuring the absorbance at 260 and 280 nm. The samples showing a 260/280 ratio exceeding 1.8 were good-quality DNA free from protein contamination. The quality of the DNA fragment was also confirmed by 0.8% agarose gel electrophoresis using 1XTBE buffer at 100 V for 90 min.
Primer selection and PCR amplification
Seventy-one SSR markers of genome-wide distribution (Supplementary Table S1), selected from various published literature and the Gramene database (www.gramene.org), were used for genotyping the cultivars.PCR was performed in a 25 μL mixture containing 1 μL (25 ng/μL) template DNA, 2.5 μL of 10x PCR buffer with 25 mM MgCl2, 1.0 μL 5 mM of each forward and reverse SSR primer, 1.0 μL 10 mM dNTPs and 0.2 μL Taq DNA polymerase. PCRs were performed in a thermal cycler (Eppendorf, Hamburg, Germany). The amplification profile consisted of initial denaturation for 2 min at 95 °C, 35–40 cycles of denaturation at 95 °C, annealing at 50–60 °C, and extension at 72 °C. After that, the final extension was carried out at 72 °C for 7 min. PCR products of SSR markers were resolved on a capillary electrophoresis system (Qiagen Pvt. Ltd., Hamburg, Germany). Only intense bands were scored based on their product size determined from a ladder of known molecular weights. The presence of a product in a particular genotype was designated as '1', and the absence as '0'.
Research involving plants
The aromatic rice cultivars (Oryza sativa L.) used in this study are maintained in the Department of Plant Breeding and Genetics, Assam Agricultural University, Jorhat-13, Assam, India. This study complied with institutional and national guidelines for experimental research involving plants.
Data analysis
A pooled ANOVA was performed for the traits over the two years, considering replication, genotype, and environment as fixed effects21 in MS Excel 2007. Genetic parameters were based on the formulae given by Burton22 for GCV and PCV, Hanson, Robinson, and Comstock23 for heritability, and Allard24 for expected genetic advances in MS Excel 2007. Mahalanobis D2 analysis25 was performed in Windostat version 9.2 (http://www.windostat.org). The principal component analysis was computed using the ‘FACTOEXTRA’ package of R statistical software26. Also, the biplot analysis was carried out using the ‘GGplot2’ package of R statistical software27. Usual Euclidian distances between the cultivars were determined from the standardized data matrix in DARwin version 6.0.02128 and represented through cluster analysis using the unweighted neighbour-joining (UNJ) method by feeding the distance matrix as input data. Genetic relatedness among the genotypes was computed by using Jaccard's coefficient of similarity29, and a dendrogram was constructed illustrating the genetic relationship among the rice genotypes using the UNJ method as proposed by Gascuel30, which uses a criterion of weighted average in DARwin 6.0.02128. The number of different alleles amplified per locus (Na), major allele frequency (MAF), number of effective alleles (Ne), Shannon's information index (I), observed heterozygosity (Ho), expected heterozygosity (He), and polymorphism information content (PIC) values were calculated using GenAlEx version 6.531.
Data availability
The supplementary information file includes all data generated or analyzed during this investigation.
References
FAO. Production/Yield quantities of rice, paddy in world + (Total) (2020). 22/12/2020. Visited on 05/01/2021. http://www.fao.org/faostat/en/#data/QC/visualize.
FAO. Food Security and agricultural mitigation in developing countries: Options for capturing synergies. Rome, Italy (2009). www.fao.org/docrep/012/i1318e/i1318e00.pdf.
Goyal, S. K. & Singh, J. P. Demand versus supply of food grains in India: Implications to food security.13th International Farm Management Congress, Wageningen, The Netherlands, July 7–12, 2002 (2002) (No. 1026-2016-82014).
Singh, S. P. et al. Maternal exposure to second hand cigarette smoke primes the lung for induction of phosphodiesterase-4D5 isozyme and exacerbated Th2 responses: Rolipram attenuates the airway hyper reactivity and muscarinic receptor expression but not lung inflammation and atopy. J. Immunol. 183(3), 2115–2121 (2009).
Nayak, A. R., Chaudhury, D. & Reddy, J. N. Genetic variability, heritability and genetic advance in scented rice. Indian J. Agric. Sci. 46(12), 45–47 (2002).
Hien, N. L., Ahmad, W. S., Oikawa, Y. & Hirata, Y. Genetic diversity of morphological responses and the relationships among Asia aromatic rice (Oryza sativa L.) Cultivar. Tropics 16(4), 343–355 (2007).
Das, A., Kesari, V. & Rangan, L. Aromatic joha rice of Assam: A review. Agric. Rev. 31(1), 1–10 (2010).
Roy, S. et al. Genetic diversity and population structure in aromatic and quality rice (Oryza sativa L.) landraces from North-Eastern India. PLoS ONE 10(6), e0129607 (2015).
Bradbury, L. M. T., Fitzgerald, T. L., Henry, R. J., Jin, Q. & Waters, D. L. E. The gene for fragrance in rice. Plant Biotechnol. J. 3, 363–370 (2005).
Travis, A. J. et al. Assessing the genetic diversity of rice originating from Bangladesh, Assam and West Bengal. Rice 8, 35. https://doi.org/10.1186/s12284-015-0068-z (2015).
Lin, M. S. Genetic base of japonica rice varieties released in Taiwan. Euphytica 56(1), 43–46 (1991).
Otegbayo, B. O., Osamuel, F. & Fashakin, J. B. Effect of parboiling on physicochemical qualities of two local rice varieties in Nigeria. J. Food Technol. Afr. 6(4), 130–132 (2001).
Dong, M. H., Sang, D. Z., Wang, P., Wang, X. M. & Yang, J. C. Changes in cooking and nutrition qualities of grains at different positions in a rice panicle under different nitrogen levels. Rice Sci. 14, 141–148 (2007).
Jones, N., Ougham, H. & Thomas, H. Genome mapping, molecular markers and marker-assisted selection in crop plants. New Phytol. 137, 165–177 (1997).
Shobha Rani, N., Shobha Rao, L. V., Viraktamath, B. C. & Mishra, B. National Guidelines for the Conduct of Tests for Distinctiveness, Uniformity and Stability. Directorate of Rice Research pp 6–13 (2004).
Johri, R. P., Singh, S. P., Srivastava, K. N., Gupta, H. O. & Lodha, M. L. Chemical and Biological Evaluation of Nutritional Quality of Food Grains: A Laboratory Manual (New Delhi Publications, 2000).
Juliano, B. O. Polysaccharides, proteins, and lipids of rice. In Rice Chemistry and Technology (ed. Juliano, B. O.) 59–174 (American Association of Chemists, 1985).
Zheljazkov, V. D. & Nielsen, N. E. Effect of heavy metals on peppermint and cornmint. Plant Soil 178, 59–66 (1998).
Sokal, R. R. & Rohlf, F. J. Biometry: The Principles and Practice of Statistics in Biological Research 3rd edn. (Freeman, 1995).
Murray, M. G. & Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 8, 4321–4325 (1980).
Singh, R. K. & Chaudhary, B. D. Biometrical Methods in Quantitative Genetic Analysis (Kalyani Publishers, 1985).
Burton, G. V. Quantitive inheritance in pearl millet. Agron. J. 50, 503 (1952).
Hanson, G. H., Robinson, H. F. & Comstock, R. E. Biometric studies in segregating population of Korean lespedesa. Agron. J. 48, 268–272 (1956).
Allard, R. W. Principles of Plant Breeding 1st edn. (Wiley, 1960).
Mahalanobis, P. C. On the generalised distance in statistics. Proc. Natl. Inst. Sci. India 2, 49–55 (1936).
Kassambara, A. & Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. (2020).
McGill, R., Tukey, J.W. & Larsen, W.A. Variations of box plots. The American Statistician. 32 12-16. R Package Version 1.0.7 (1978).
Perrier, X., & Jacquemoud-Collet, J. P. DARwin software: Dissimilarity analysis and representation for windows. http://darwin.cirad.fr/Darwin (2006).
Jaccard, P. Nouvelle recherchés sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 44, 223–270 (1980).
Gascuel, O. BIONJ: an improved version of the NJ algorithm based on a simple model of sequence data. Mol. Biol. Evol. 14(7), 685–695 (1997).
Peakall, R. O. D. & Smouse, P. E. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6(1), 288–295 (2006).
Wander, M. M. & Bollero, G. A. Soil quality assessment of tillage impacts on Illinois. Soil Sci. Soc. Am. J. 63, 961–971 (1999).
Raut, V. M. Qualitative genetics of Soyabean: A review. Soybean Res. 1, 1–28 (2003).
Sinha, A. K. & Mishra, P. K. Morphology based multivariate analysis of phenotypic diversity of landraces of rice of Bankura district of West Bengal. J. Crop Weed. 9(2), 115–121 (2013).
Mondal, B., Singh, S. P. & Joshi, D. C. DUS characterization of rice (Oryza sativa L.) using morphological descriptors and quality parameters. Outlook Agric. 43(2), 131–137 (2014).
Gangashetty, P. I., Salimath, P. M. & Hanamaratti, N. G. Genetic variability studies in genetically diverse non-basmati local aromatic genotypes of rice (Oryza sativa L.). Rice Genom. Genet. 4(2), 4–8 (2013).
Verma, P. K., Chaurasia, A. K., Bara, B. M. & Deepshikha, M. Evaluation of aromatic short grain rice cultivars and elite lines for yield and quality parameters. Int. J. Res. Eng. Technol. 3(6), 103–106 (2014).
Song, Y., Oh, N. & Hwang, C. Growth and response of yield characters of dry seeded rice at different seeding dates in plain area. RDA J. Agric. Sci. 38(2), 20–31 (1996).
Nahar, K., Hasanuzzaman, M. & Majumder, R. R. Effect of low temperature stress in transplanted aman rice varieties mediated by different transplanting dates. Acad. J. Plant Sci. 2(3), 132–138 (2009).
Khalifa, A. A. B. A. Physiological evaluation of some hybrid rice varieties under different sowing dates. Aust. J. Crop Sci. 3(3), 178–183 (2009).
Patel, A. R., Patel, M. L., Patel, R. K. & Mote, B. M. Effect of different sowing date on phenology, growth and yield of rice: A review. Plant Arch. 19(1), 12–16 (2019).
Panwar, L. L., Joshi, V. N. & Ali, M. Genotype x environment interaction in scented rice. Oryza 45(2), 103–109 (2008).
Singh, B., Mishra, M. K. & Naik, R. K. Genetic diversity among some traditional aromatic rice (Oryza sativa L.) varieties of Orissa. Ind. J. Agric. Res. 44(2), 141–145 (2010).
Semwal, D. P., Pandey, A., Bhandari, D. C., Dhariwal, O. P. & Sharma, S. K. Variability study in seed morphology and uses of indigenous rice landraces (Oryza sativa L.) collected from West Bengal, India. AJCS. 8(3), 460–467 (2014).
Bajpai, A. & Singh, Y. Study of quality characteristics of some small and medium grained aromatic rices of Uttar Pradesh and Uttarakhand. Agric. Sci. Digest. 30(4), 241–245 (2010).
Karim, D., Sarkar, U., Siddique, M. N. A., Khaleque, M. M. A. & Hasant, M. Z. Variability and genetic parameter analysis in aromatic rice. Int. J. Sustain. Crop Prod. 2(5), 15–18 (2007).
Chavan, B. R., Dalvi, V. V., Kunkerkar, R. L., Mane, A. V. & Gokhale, N. B. Studies on genetic variability for yield and yield contributing traits in aromatic rice (Oryza sativa L.). Pharma Innov. J. 11(2), 1732–1735 (2022).
Debsharma, S. K. et al. Harnessing on genetic variability and diversity of rice (Oryza sativa L.) genotypes based on quantitative and qualitative traits for desirable crossing materials. Genes (Basel) 14(1), 10. https://doi.org/10.3390/genes14010010 (2023).
Akshay, M., Chandra, B. S., Devi, K. R. & Hari, Y. Genetic variability studies for yield and its attributes, quality and nutritional traits in rice (Oryza sativa L.). Pharma Innov. J. 11(5), 167–172 (2022).
Faysal, A. S. M. et al. Genetic variability, character association, and path coefficient analysis in transplant Aman rice genotypes. Plants 11, 2952. https://doi.org/10.3390/plants11212952 (2022).
Chaurasia, A. K., Rai, P. K. & Kumar, A. Estimation of genetic variability, heritability and genetic advance in aromatic fine grain rice. Roman. J. Biol. Plant Biol. 57(1), 71–76 (2012).
Jaiswal, H. K., Shrivastava, A. K. & Dey, A. Variability and association studies in indigenous aromatic rice. Oryza 44(4), 351–353 (2007).
Hasib, K. M., Ganguli, P. K. & Kole, P. C. Evaluation of the performance of advanced generation lines of mutant x Basmati crosses of scented rice. J. Interacad. 8(1), 7–10 (2004).
Nandan, R. & Singh, S. K. Character association and path analysis in rice (Oryza sativa L.) genotypes. World J. Agric. Sci. 6(2), 201–206 (2010).
Adjah, K. L., Abe, A., Adetimirin, V. O. & Asante, M. D. Genetic variability, heritability and correlations for milling and grain appearance qualities in some accessions of rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 26(6), 1309–1317 (2020).
Allam, C. R., Jaiswal, H. K. & Qamar, A. Divergence analysis for yield and quality traits in some indigenous basmati rice genotypes (Oryza sativa L). Int. J. Appl. Biol. Pharm. Technol. 5(4), 257–263 (2014).
Patel, N. B., Shrivastava, R., Kumar, V. & Dhirhi, N. Genetic diversity among some traditional aromatic rice (Oryza sativa L.) landraces of Chhattisgarh. Electron. J. Plant Breed. 6(1), 204–209 (2015).
Prasad, G. S. V. et al. Assessment of diversity of Indian aromatic rice germplasm collections for morphological, agronomical, quality traits and molecular characters to identify a core set for crop improvement. CABI Agric Biosci 1, 13. https://doi.org/10.1186/s43170-020-00013-8 (2020).
Barhate, K. K., Jadhav, M. S. & Bhavsar, V. V. Genetic diversity analysis in aromatic lines of rice (Oryza sativa L.). J. Pharmacogn. Phytochem. 10(3), 367–370 (2021).
Netam, V., Sinha, S. K., Tigga, K., Singh, V. K. & Chouksey, N. Genetic diversity analysis by D2 analysis in fine scented genotypes of rice (Oryza sativa L.). Int. J. Curr. Microbiol. Appl. Sci. 10(11), 48–55 (2021).
Bourgis, F. et al. Characterization of the major fragrance gene from an aromatic japonica rice and analysis of its diversity in Asian cultivated rice. Theor. Appl. Genet. 117, 353–368 (2008).
Wu, J. et al. Genetic architecture of flag leaf length and width in rice (Oryza sativa L.) revealed by association mapping. Genes Genom. 39, 341–352 (2017).
Yuan, L. P. Hybrid rice breeding for super high yield. Hybrid Rice 12, 1–6 (1997).
Li, Z. K., Pinson, S. R. M. & Stansel, J. W. Genetic dissection of the source-sink relationship affecting fecundity and yield in rice (Oryza sativa L.). Mol. Breed. 4, 419–426 (1998).
Singh, Y., Pani, D. R., Pradhan, S. K., Bajpai, A. & Singh, U. S. Divergence analysis for quality traits in some indigenous Basmati rice genotypes. Crop. Improv. 45(4), 263–267 (2008).
Anderson, T. W. An Introduction to Multivariate Analysis (Wiley, 1972).
Lakshmi, M., Shanmuganathan, M., Jeyaprakash, P. & Ramesh, T. Genetic variability and diversity analysis in selected rice (Oryza sativa L.) varieties. Electron. J. Plant Breed. 13(3), 959–966 (2022).
Burman, M., Nair, S. K. & Sarawgi, A. K. Principal component analysis for yield and its attributing traits in aromatic landraces of rice (Oryza sativa L). Int. J. Bio-resour. Stress Manag. 12(4), 303–308 (2021).
Ahmed, M. S. U., Khalequzzaman, M., Bashar, M. K. & Shamsuddin, A. K. M. Agro-morphological, physico-chemical and molecular characterization of rice germplasm with similar names of Bangladesh. Rice Sci. 23(4), 211–218 (2016).
Pachauri, A. K., Sarawgi, A. K., Bhandarkar, S. & Ojha, G. C. Agro-morphological characterization and morphological based genetic diversity analysis of Rice (Oryza sativa L.) germplasm. J. Pharmacogn. Phytochem. 6(6), 75–80 (2017).
Blu, R. O. Analysis of factors affecting spikelet sterility in flooded rice under field conditions in Chile. Arch. Agron. Soil Sci. 53, 183–192 (2007).
Li, R., Li, M., Ashraf, U., Liu, S. & Zhang, J. Exploring the relationships between yield and yield-related traits for rice varieties released in China from 1978 to 2017. Front. Plant Sci. 10, 543 (2019).
Nagy, K. & Tiuca, I.-D. Importance of fatty acids in physiopathology of human body. Fatty Acids https://doi.org/10.5772/67407 (2017).
Glick, N. R. & Fischer, M. H. The role of essential fatty acids in human health. J. Evid. Based Complement. Altern. Med. 18(4), 268–289. https://doi.org/10.1177/2156587213488788 (2013).
Jennings, B. H. & Akoh, C. A. Effectiveness of natural versus synthetic antioxidants in a rice bran oil-based structured lipid. Food Chem. 114(4), 1456–1461 (2009).
Oluremi, O. I., Solomon, A. O. & Saheed, A. A. Fatty acids, metal composition and physico-chemical parameters of Igbemo Ekiti rice bran oil. J. Environ. Chem. Ecot. 5(3), 39–46 (2013).
Law, M. Dietary fat and adult diseases and the implications for childhood nutrition: An epidemiologic approach. Am. J. Clin. Nutr. 72(5), 1291–1296 (2000).
Sayre, R. N. & Saunders, R. M. Rice bran and rice bran oil. Lipid Technol. 2, 72 (1990).
Resurreccion, A. P. & Juliano, B. O. Fatty acid composition of rice oils. J. Sci. Food Agric. 26(4), 437–439 (1975).
Goffman, F. D., Pinson, S. & Bergman, C. Genetic diversity for lipid content and fatty acid profile in rice bran. J. Am. Oil Chem. Soc. 80(5), 485–490 (2003).
Tako, E. Addressing Iron and Zinc Deficiency Across the Globe. https://www.ars.usda.gov/oc/utm/ (7/12/2023)
Vanlalsanga, S., Singh, P. & Singh, Y. T. Rice of northeast India harbor rich genetic diversity as measured by SSR markers and Zn/Fe content. BMC Genet. 20, 79. https://doi.org/10.1186/s12863-019-0780-6 (2019).
Bouis, H. & Welch, R. Biofortification—a sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Sci. 50, 20–32 (2010).
Hambridge, M. Human zinc deficiency. J. Nutr. 130(5), 1344S-S1349. https://doi.org/10.1093/jn/130.5.1344S (2000).
Zhang, C. Essential functions of iron-requiring proteins in DNA replication, repair and cell cycle control. Plant Cell 5(10), 750–760 (2014).
Chowdhury, B. D., Nath, A. & Dasgupta, T. Evaluation of some popular rice genotypes with special emphasis on zinc, iron and protein content. Int. J. Sci. Res. Publ. 6(7), 609–619 (2016).
Brar, B., Jain, S., Singh, R. & Jain, R. K. Genetic diversity for iron and zinc contents in a collection of 220 rice (Oryza sativa L.) genotypes. Indian J. Genet. 71(1), 67–73 (2011).
Anuradha, K. et al. Mapping QTLs and candidate genes for iron and zinc concentrations in unpolished rice of Madhukar x Swarna RILs. Gene 508, 233–240. https://doi.org/10.1016/j.gene.2012.07.054 (2012).
Martínez, C.P., Borrero, J., Taboada, R., Viana, J. L., Neves, P., Narvaez, L., Puldon, V., Adames, A. & Vargas, A. Rice cultivars with enhanced iron and zinc content to improve human nutrition. 28th International Rice Research Conference, 8–12 November (2010), Hanoi, Vietnam OP10: Quality Grain, Health, and Nutrition.
Raza, Q. et al. Genetic diversity in traditional genotypes for grain iron, zinc and β-carotene contents reveal potential for breeding micronutrient dense rice. J. Exp. Biol. Agric. Sci. 7(2), 194–203 (2019).
Sanjeeva Rao, D. et al. Zinc biofortified rice varieties: challenges, possibilities, and progress in India. Front. Nutr. https://doi.org/10.3389/fnut.2020.00026 (2020).
Krupa, K. N., Ningaraj, D., Shashidhar, H. E. & VijaykumaraSwamy, H. V. Determination of effect of different levels of polishing on zinc content in rice grains. Int. J. Pure Appl. Biosci. 5(1), 694–702. https://doi.org/10.18782/2320-7051.2588 (2017).
Chandel, G. et al. Effects of different nitrogen fertilizer levels and native soil properties on rice grain Fe, Zn and Protein Contents. Rice Sci. 17(3), 213–227 (2010).
Banerjee, S., Chandel, G., Mandal, N., Meena, M. & Saluja, T. Assessment of nutritive value in milled rice grain of some Indian rice landraces and their molecular characterization. Bangladesh J. Agril. Res. 36(3), 369–380 (2011).
Shah, S. M., Naveed, S. A. & Arif, M. Genetic diversity in Basmati and non-Basmati rice varieties based on microsatellite markers. Pak. J. Bot. 45(51), 423–431 (2013).
Meti, N., Samal, K. C., Bastia, D. N. & Rout, G. R. Genetic diversity analysis in aromatic rice genotypes using microsatellite based simple sequence repeats (SSR) marker. Afr. J. Biotechnol. 12(27), 4238–4250 (2013).
Rahman, M. M., Rasaul, M. G., Hossain, M. A., Iftekharuddaula, K. M. & Hasegawa, H. Molecular characterization and genetic diversity analysis of rice using SSR markers. J. Crop Improv. 26, 244–257 (2012).
Sajib, A. M. et al. SSR marker-based molecular characterization and genetic diversity analysis of aromatic landraces of rice (Oryza sativa L.). J. Bio Sci. Biotech. 1(2), 107–116 (2012).
Kumar, V., Kumar, S., Chakrabarty, S. K., Mahapatra, T. & Dadlani, N. Molecular characterization of farmers’ varieties of rice (Oryza sativa). Indian J. Agric. Sci. 85(1), 118–124 (2015).
Aljumaili, S. J. et al. Genetic diversity of aromatic rice germplasm revealed by SSR markers. Biomed. Res. Int. 2018, 1–11. https://doi.org/10.1155/2018/7658032 (2018).
Yelome, O. I. et al. Analysis of population structure and genetic diversity reveals gene flow and geographic patterns in cultivated rice (O. sativa and O. glaberrima) in West Africa. Euphytica 11, 214–215. https://doi.org/10.1007/s10681-018-2285-1 (2018).
Kibria, K., Islam, M. M. & Begum, S. N. Screening of aromatic rice lines by phenotypic and molecular markers. Bangladesh J. Bot. 37(2), 141–147 (2008).
Nadia, I., Mohiuddin, A. K. M., Sultana, S. & Ferdous, J. Diversity analysis of indica rice accessions using morphological and SSR markers. Ann. Biol. Res. 5(11), 20–31 (2014).
Shamim, M. Z., Manzar, H., Sharma, V. K. & Kumar, P. Microsatellite marker-based characterization and divergence analysis among rice varieties. Indian J. Biotechnol. 15, 182–189 (2016).
Islam, M. Z. et al. Variability assessment of aromatic rice germplasm by pheno-genomic traits and population structure analysis. Sci. Rep. 8(1), 9911 (2018).
Acknowledgements
The authors thankfully acknowledge the financial support from the Assam Agricultural University, Assam and the Board of Research in Nuclear Science (BRNS), Department of Atomic Energy, Government of India, Mumbai.
Author information
Authors and Affiliations
Contributions
D.S., N.S.B., B.K.D. and R.D. conceived and designed the study; D.B. accomplished the field/laboratory works under the supervision of D.S., N.S.B., B.K.D. and R.D.; D.B. collected data; D.S. and D.B. analysed and interpreted the results; D.B prepared the original draft and all the authors reviewed and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bordoloi, D., Sarma, D., Sarma Barua, N. et al. Morpho-molecular and nutritional profiling for yield improvement and value addition of indigenous aromatic Joha rice of Assam. Sci Rep 14, 3509 (2024). https://doi.org/10.1038/s41598-023-42874-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-42874-9
- Springer Nature Limited