
Abstract
Ciprofloxacin (CIP) and norfloxacin (NOR) belong to the class of emerging contaminants that are frequently detected in the aquatic environment as a binary mixture, responsible for the development of antibiotic-resistant genes and antibiotic-resistant bacteria. This study aims to investigate five different algal species Chlorella vulgaris (Cv), Chlorella pyrenoidosa (Cp), Scenedesmus obliquus (So), Tetradesmus sp (T) and Monoraphidium sp (M) for their tolerance and removal of binary mixture. The effects on biochemical composition in the algal species concerning the binary mixture and its removal efficiency are first reported in this study. The acute toxicity (96 h EC50) values are in the order of So > Cp > T > M > Cv, Chlorella vulgaris is the most sensitive algal species with 17.73 ± 0.24 mg/L and Scenedesmus obliquus is the least sensitive algal species with 39.19 ± 0.79 mg/L. The removal efficiency of the binary mixture was found to be in the order of So > Cp > T > M > Cv, Scenedesmus obliquus removed CIP (52.4%) and NOR (87.5%) with biodegradation as the major contributing removal mechanism. Furthermore, less toxic biotransformed products were detected in Scenedesmus obliquus and the biochemical characterization revealed that the growth-stimulating effect is higher with lipid (35%), carbohydrate (18%), and protein (33%) providing an advantage in the production of valuable biomass.
Similar content being viewed by others


Introduction
The worldwide consumption of antibiotics has increased from 635 to 674 units per 1000 population in developed countries and 1490 units to 1593 units in developing countries1. Studies have reported that 50–80% of the administered antibiotics are excreted through feces and urine as the parent compound and reach the aquatic environment, which has led to the development of antibiotic-resistant genes (ARGs) that have raised concerns in public health globally2. ARG has led to the death of 25,000 infants in European Union countries, 23,000 infants in the USA, and 58,000 infants in India every year3,4. Municipal wastewater is the major source of antibiotic pollution compared to hospital wastewater. The wastewater treatment plants (WWTPs) are not designed to eliminate these pollutants, as the metagenomic analysis has revealed that in WWTPs of 60 countries have a divergent distribution of ARGs5,6,7. India has the third largest active pharmaceutical industry fulfilling 50% of the generic medicine demand globally and is one of the top five pharmaceuticals consuming countries which is predicted to have an increase of 84% by 20308,9. Fluoroquinolone (FQs) belongs to the third largest antibiotics accounting for 17% of the global market and it is an important class of antibiotics that has been extensively used to treat respiratory and urinary infections10,11. Ciprofloxacin (CIP) and norfloxacin (NOR) are the most frequently detected FQs in surface waters exceeding a 50% detection rate globally due to their excessive usage and it has been detected at different concentrations ranging from ng/L to µg/L12.
Various conventional and advanced treatment methods can be employed for the treatment of these pollutants but high operations and maintenance costs restrict their utilization in the municipal WWTPs. The incomplete mineralization of these contaminants generates transformed products that are toxic to the aquatic community and it has become a major concern6. Biodegradation by microorganisms is a good approach in the treatment of these pollutants with the advantage of environmental-friendliness and cost-effectiveness12,13. FQs are not easily biodegradable in conventional WWTPs but when treated with specific microbial species FQs can be biologically treated14. Algae are non-target organisms for antibiotics and are known to treat wastewater effectively removing nutrients, organic pollutants, and heavy metals with the advantage of biomass production6,15. CIP and NOR are tested for phycoremediation and studies have shown the ability of algal species to biodegrade the pollutants individually16,17,18,19. Chlorella vulgaris (Cv), Chlorella pyrenoidosa (Cp), and Scenedesmus obliquus (So) have shown their potential in removing CIP, 85% (Cv), 79% (Cp), and 75% (So) and for NOR, 52% (Cv), 50% (Cp), and 43% (So) with initial concentration CIP (0.029 µg/L) and NOR (0.032 µg/L) separately20. CIP and NOR have been detected in the Indian surface waters as a binary mixture with the maximum detected concentration of 6.5 mg/L (CIP) and 0.52 mg/L (NOR)21.
Individual toxicity assessment studies have reported that an increase in the concentration of pollutants will inhibit the growth of algae and this will have an impact on their removal and biochemical composition16,22. EC50 (concentration at which 50% of the growth rate of algae is inhibited) is the most accepted evaluation for toxicity assessment in algal species23. There are also other algal species such as Tetradesmus sp (T) and Monoraphidium sp (M) that requires much attention in phycoremediation studies for the removal of antibiotics. Tetradesmus sp (T) and Monoraphidium sp (M) have shown good phycoremediation properties and have the potential to produce valuable biomass with good biochemical properties24,25,26,27, but both the species have yet not been tested for their potential in the removal of FQs. Phycoremediation potential can be stunned when the test values are greater than EC50. Therefore, a growth inhibition test must be performed before employing the algae for the removal of pollutants, as toxic pollutants are known to inhibit the growth of algae. These results will contribute to the FQs toxicity data on these selected species and the application of these species in wastewater treatment plants can be identified.
Most of the studies have focused on the individual EC50 assessment and removal of pollutants, without any information regarding its effects on biochemical composition (proteins, carbohydrates, and lipids)16,17,28,29. In the case of a binary mixture (CIP+NOR) acute toxicity assessment, removal, and its biotransformed products have not yet been reported, as the combination of these pollutants exists in the environment. With this in view, this study aims to screen and select the most effective and tolerant algal species among the five different algae representing four different genera (Cv, Cp, So, T, and M), to assess the acute toxicity of binary mixtures and its effects on biochemical composition (proteins, carbohydrates, and lipids) in the selected algal species, and to investigate the removal potential of the binary mixture (CIP+NOR) and the mechanisms adapted by the algal species for their removal, biotransformed products, and its effects on biochemical composition (proteins, carbohydrates, and lipids).
Materials and methods
Fluoroquinolones
Stock solutions of norfloxacin (NOR) (CAS No: 70458-96-7) and ciprofloxacin (CIP) (CAS No: 85721-33-1) with > 98% purity (HPLC grade) were prepared by dissolving CIP and NOR in the ultrapure water and stored in an airtight Schott glass container at 4 °C in dark conditions for no longer than 7 days.
Determination of fluoroquinolones concentration
CIP and NOR concentrations were determined according to the USP 28-NF 23 s supplement using 844 UV–VIS compact ion chromatography equipped with an HPLC column (Hichrom-Alltima 5 µm C-18 with dimensions 250 × 4.6 mm)30,31,32. Acetonitrile (15%) and ultrapure water (85%) with pH = 3.2 were used as the mobile phase with a flow rate adjusted to 1 mL/min and 250 µL injection volume19. The wavelength of the UV detector was set for 280 ± 5 nm which corresponds to the maximum wavelength for both the antibiotics and retention time for NOR was 14.5 min and for CIP 16.5 min, respectively. The limit of detection for CIP and NOR in this study was 0.03 µg/L and 0.02 µg/L, respectively. The concentration was estimated using IC-NET 2.3 software integrated with HPLC equipment and upon the completion of the analysis, the consolidated overlay curves of the individual analysis were plotted using the IC-NET software. The biotransformed products were identified using Liquid chromatography-electron spray ionization tandem mass spectrometry. Formic acid (0.1%) and acetonitrile at a volumetric ratio (95:5) is used as mobile phase. Mass spectrometer conditions and the column details are presented elsewhere33.
Algal species and culture conditions
Chlorella vulgaris (Cv) (BDU-GD003) was procured from the National Repository for Microalgae and Cyanobacteria (NRMC), Bharathidasan University, Tamilnadu, India. Chlorella pyrenoidosa (Cp) (NCIM-2738), Scenedesmus obliquus (So) (NCIM-5586), Tetradesmus sp (T) (NCIM-5797), and Monoraphidium sp (M) (NCIM-5792), were procured from the National Collection of Industrial Microorganisms (NCIM), Pune, India. All the procured cultures were sub-cultured in BG-11 medium under sterile conditions in an algal growth chamber with the light intensity of 50 µmol photon/m2/s in a 12 h light and dark cycle at 27 °C until their growth phase was attained.
Determination of algal growth
Cell density (cells/mL) was measured using Neubauer’s improved hemocytometer under 40 × trinocular microscope magnification and the specific growth rate (µ) was calculated using the following Eq. (1)34.
where Ci and Cf are the cell density measured at the initial time (ti) and final time (tf).
Extraction and analysis of chlorophyll-a, chlorophyll-b, and carotenoid
2 mL of algal cells were harvested by centrifuging the cells at 5000 rpm for 10 min from the experimental runs. The supernatant was discarded and resuspended with 5 mL of 90% methanol and incubated at 60 °C for 5 min and then centrifuged again for 10 min. The supernatants were collected in separate tubes and optical densities were measured for 665, 652, and 470 nm wavelength using a UV–visible spectrophotometer, and the concentration of the extracted pigment was calculated using the following Eqs. (2)–(4)16.
Determination of biochemical composition
Protein, carbohydrates, and lipids are the three important biochemical compositions of algae. Samples collected from the experimental runs were subjected to centrifugation to extract the cell pellets from the algal medium35. The harvested cell pellets were subjected to the lowry method to extract the protein from the harvested cell pellets and quantified using bovine serum albumin as standards in a UV–visible spectrophotometer36.The Anthrone method was used to extract the carbohydrates from the harvested cell pellets and quantified using glucose as standards in a UV–visible spectrophotometer36. Lipids were extracted using the sonication method and quantified gravimetrically36,37. Dry cell weight (DCW) was calculated gravimetrically by drying the harvested cell pellets. Lipids, protein, and carbohydrates were expressed on a percentage basis from the harvested biomass using the following Eqs. (5)–(7)35.
Growth inhibition test
Acute toxicity experiments were conducted according to OECD 201 guidelines23. Different concentrations of CIP and NOR ranging between 1 and 100 mg/L were used for individual tests, and CIP+NOR equal proportions of fixed concentrations ratio of (1:1) ranging between 1 and 50 mg/L were spiked into the 100 mL algal medium. The initial cell density for all the experiments was maintained at 100 × 104 cells/mL. Three replicates of test runs (with pollutants) and control runs (without pollutants) were incubated in the algal growth chamber equipped with an orbital shaker (120 rpm) for the growth inhibition test. Percentage inhibition (%I) was calculated using the following Eq. (8)38.
where \(\mu c\) and \(\mu t\) are the cell density measured in the control run and test run.
EC50 is the effective concentration that inhibits 50% of the algal growth. A dose–response curve was plotted using percentage inhibition (%I) calculated for different concentrations, to find EC50 linearising the response data is needed to perform the regression analysis, Probit statistical analysis was performed to linearise the data as per OECD 201 guidelines23 to calculate EC50. The EC50 values of biotransformed products were predicted using ECOSAR software (version 2.2)39.
Risk quotients assessment of binary mixture
Risk quotients (RQs) try to estimate the potential risk for algae caused by the environmental detected maximum concentration. RQ is calculated by the following Eqs. (9) and (10) 40.
where MEC is measured environmental concentration, PNEC is predicted no effect concentration.
When calculated
RQ > 1 indicates that there is an ecotoxicological risk for the algal species.
RQ < 1 indicates there is no risk for the algal species.
Determination of fluoroquinolones removal mechanism in algal cells
A series of experimental runs were performed with five different algal species with control runs (without CIP + NOR) and test runs (with CIP + NOR) in 250 mL Erlenmeyer flasks containing 100 mL of algal medium with an average initial cell density of 100 × 104 cells/mL. CIP (6.5 mg/L) and NOR (0.5 mg/L) were spiked in the test runs. Duplicate experimental runs were incubated in the algal growth chamber equipped with an orbital shaker (120 rpm) with the light intensity of 50 µmol photon/m2/s in a 12-h light and 12-h dark cycle at T = 30 °C ± 1 °C until the growth phase declined to death phase. 5 mL of sample is collected from the experimental runs and centrifuged for 10 min at 2500 rpm for the determination of CIP+NOR total removal is calculated by the following Eq. (11)19.
where Ci is the initial concentration of (CIP+NOR) in the medium and Cf is the residual concentration of (CIP+NOR).
Bioadsorption, bioaccumulation, and biodegradation are the three major removal mechanisms adopted by algae in the removal of antibiotics41. Bioadsorption (Rad) is determined using the high-speed centrifuging method by increasing the rpm from 2500 to 5000 rpm without disrupting the cell wall19,42,43. The collected supernatant was used for the determination of Rad. Bioaccumulation (Rac) is determined using the ultrasonication method19,44. After 30 min of sonication, the sample is centrifuged for 10 min at 2500 rpm and the supernatant was used for the determination of Rac. Biodegradation (Rb) is calculated using the following Eq. (12)19.
Abiotic removal (photodegradation) is determined by keeping the control runs without algae in light and dark conditions for a period of 7 days) and is calculated using the following Eq. (12). Since FQs are known to be photosensitive compounds Eq. (13)17,19,45.
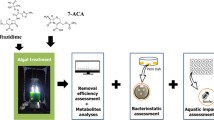
Figure 1 depicts the overall methodology followed in this study.

Methodology followed in this study.
Statistical analysis and plotting
The growth inhibition study was conducted in triplicates and the Probit analysis for EC50 was calculated using SPSS software (version 26). The removal study was conducted in duplicates and the mean values were presented as results. One-way ANOVA coupled with the Dunnett test was conducted to find the statistical significance (p < 0.05) between the control runs and the test runs in the experimental study. The statistical analysis and graph plotting were carried out using origin pro software (version 9.9). The chemical structures were depicted using Chemdraw (version 20.0).
Results and discussion
Acute toxicity and risk quotients assessment of CIP and NOR
Acute toxicity assessment of CIP, NOR, and binary mixture (CIP+NOR), upon the selected algal species was conducted to understand the growth stimulatory and inhibitory effects that a target pollutant can cause individually and in combined form. Figure 2 depicts the dose–response curve for CIP, NOR, and CIP+NOR. The highest growth inhibition for CIP was recorded for Monoraphidium sp (M) 78.6% followed by 67.6% (Cv), 65.1% (Cp), 62.6% (T), and 61% (So). For NOR, the highest growth inhibition of 73.6% was recorded for Chlorella vulgaris (Cv) followed by 72.6% (T), 65.6% (M), 60.4% (Cp), and 56.9% (So). For binary mixture, the highest growth inhibition was observed in Chlorella vulgaris (Cv) 73.33% followed by 72.5% (M), 66.6% (Cp), 63% (T), and 49.8% (So). EC50 values were calculated using the Probit statistical analysis from the experimental runs (CIP, NOR, and CIP + NOR) and tabulated in Table 1. The 96 h EC50 values are in the order of (So) > (T) > (M) > (Cp) > (Cv) for CIP, (So) > (Cp) > (M) > (T) > (Cv) for NOR, and (So) > (Cp) > (T) > (M) > (Cv) for CIP+NOR. The higher the EC50 greater their tolerance towards the specific pollutant. From the EC50 values, the PNEC and RQs were calculated and tabulated in Table 1.

Dose—Growth inhibition (%) response curve for (a) CIP, (b) NOR, (c) CIP+NOR for algal species.
The Commission of the European Communities (EU directive 93/67/EEC) classifies the pollutant in the “harmful” category to the aquatic organism when the calculated EC50 value falls within the range of (10–100 mg/L)46. While categorizing the algal species according to calculated EC50 values and RQ values, results indicate that CIP+NOR is harmful and possess a high risk to the algal species. The individual acute toxicity assessment results also affirm that it poses a threat to the algal species even when they are present in a separate form. Studies conducted for EC50 growth inhibition test for CIP and NOR also affirm that the values fall under the harmful category for Chlorella vulgaris (CIP-96 h EC50 < 100 mg/L)47,48 and (NOR-72 h EC50 < 100 mg/L)49.
The inhibitory effects increased acute toxicity assessment of CIP+NOR increased due to the combined toxicity. A study performed by Magdaleno et al.40 reported that growth inhibition caused by the binary mixture (ciprofloxacin and cephalothin) for the algae Pseudokirchneriella subcapitata is greater than the individual growth inhibition and the results obtained in this study is in line with the reported literature.
The response to the pollutants in algal species is known to vary according to their physiology, morphology, cytology, and phylogenetics characteristics22. The inhibitory effects varied according to the species and their tolerance toward the pollutants, as the binary mixture had a serious impact on their inhibition levels. Scenedesmus obliquus (So) is the least sensitive algal species to the toxic effects of antibiotics (CIP, NOR, and CIP+NOR) whereas, Chlorella vulgaris (Cv) is the most sensitive species among the selected algal species. No toxicity data for Chlorella pyrenoidosa (Cp), Scenedesmus obliquus (So), Tetradesmus sp (T), and Monoraphidium sp (M) have been reported for CIP, NOR, and CIP+NOR. Therefore, these results contribute to the FQs toxicity data on these selected species.
Effect of CIP+NOR on biochemical composition in algal species
Effect on pigments (chl-a, chl-b, and carotenoids)
Chlorophyll is a major light-harvesting pigment involved in photosynthesis and exposure to pollutants is known to alter the chl-a and chl-b synthesis47,50. During the exposure of algae to environmental stress, carotenoids are known to serve as a photosystem protective mechanism and it plays a crucial role in quenching the singlet oxygen by deactivating the active chlorophyll51. The extracted chl-a, chl-b, and carotenoid pigments from the algal species were plotted to study the binary effects of CIP+NOR upon pigments at different concentrations as shown in Fig. 3. The effect varied according to the sensitivity of the algal species as well as the concentration of the pollutants. It is observed that in the algal species Chlorella pyrenoidosa (Cp) and Scenedesmus obliquus (So), there is an increase in pigment activity at concentrations < 10 mg/L (Fig. 3b,c) and the carotenoids concentration increased up to their corresponding EC50 levels as a response to the stress and then significantly decreased to 11% (So) and 13% (Cp) at 50 mg/L concentration indicating the damage to the algal cells as the concentration increases.

Effect of the binary mixture (CIP+NOR) on chl-a, chl-b, and carotenoids for (a) Cv, (b) Cp, (c) So, (d) T, and (e) M.
In Tetradesmus sp (T) the same activity in pigments was observed up to 5 mg/L (Fig. 3d) and the carotenoid contents decreased up to 27% at 50 mg/L concentration. In Monoraphidium sp (M) increased pigment activity was observed at concentrations < 2.5 mg/L (Fig. 3e) and the carotenoid contents decreased up to 47% at 50 mg/L concentration and for Chlorella vulgaris (Cv), increased chl-a and chl-b activity was observed at 1 mg/L (Fig. 3a), being the most sensitive algal species among the selected algal species. The carotenoid concentration gradually decreased up to 50% at 50 mg/L concentration indicating damage to the cells. The results of the present study are consistent with the studies carried out on Chlamydomonas mexicana16, Chlamydomonas reinhardtii51, and Scenedesmus obliquus42 which affirms that toxic stress can alter pigment activity according to their exposed concentration in the medium and hinder the photosynthetic activity in algal cells.
Effect on protein, carbohydrates, and lipids
After 96 h exposure to the various concentration of CIP+NOR to the selected algal species, the secretion of the protein, lipid, and carbohydrate concentration varied according to the algal species. Figure 4 depicts the effect of different concentrations of the binary mixture (CIP+NOR) on the protein, lipid, and carbohydrates in the selected algal species (Cv, Cp, So, T, and M) to their control runs respectively. The growth-stimulating effects were observed in all the selected algal species within their calculated 96 h EC50 value. For Chlorella vulgaris (Cv) and Chlorella pyrenoidosa (Cp), growth-stimulating effects were observed below their EC50 value (< 25 mg/L). Inhibitory effects were observed when concentration increased beyond the EC50 value. The accumulation of protein has decreased drastically in all the concentrations compared to the control runs. Maximum lipid accumulation for Chlorella vulgaris (375 mg/L) was reported at 1 mg/L and for Chlorella pyrenoidosa (646 mg/L) at 5 mg/L. In our previous study19, when Chlorella vulgaris was exposed to CIP (5 mg/L), growth-stimulating effects were observed with higher accumulation of lipids (465 mg/L), carbohydrates (39 mg/L), and protein (608 mg/L) than reported in this study. The decrease in the biochemical composition in this study is attributed to the binary mixture toxicity.

Effect of the binary mixture (CIP+NOR) on protein, lipids, and carbohydrates for (a) Cv, (b) Cp, (c) So, (d) T, and (e) M.
When Scenedesmus obliquus (So) was exposed to the binary mixture, the lipid concentration increased up to 319 mg/L and then decreased beyond the EC50 concentration (39 mg/L). An increase in protein concentration was observed at 1 mg/L and 2.5 mg/L and then reduced drastically compared to control runs, whereas for carbohydrates the value decreased in all test runs. For Tetradesmus sp (T), there is a gradual decrease in protein and carbohydrates when the CIP+NOR concentration increases but lipid accumulation was higher in all the concentrations. The maximum lipid accumulation of 290 mg/L was observed at 5 mg/L (CIP+NOR). In the Monoraphidium sp (M) species, at 1 mg/L of CIP+NOR concentration, maximum accumulation of lipids (296 mg/L) and proteins (518 mg/L) was observed, whereas for carbohydrates the levels dropped in all the test runs. The reason for this behaviour in algal cells is due to the fact that during photosynthesis chlorophyll pigment performs carbon fixation and stores energy in the form of starch, but when the photosynthetic activity of the algae decreases the carbohydrate production also decreases. The observed results in this present study are consistent with the previous report on antibiotic stress in algae52.
Removal of the binary mixture (CIP+NOR)
A binary mixture of CIP and NOR with concentrations of 6.5 mg/L (CIP) and 0.5 mg/L (NOR) is selected for the removal studies as this binary mixture at these concentrations is detected in the Indian aquatic environment21. The EC50 value is known to increase when the exposure time is increased as algae are known to acclimatize the toxic conditions to survive in the aquatic environment16. From the acute toxicity results, we can conclude that all the selected algal species can tolerate this selected concentration of the binary mixture, growth-stimulating effect in algae can enable the algal species to remove the binary mixture from the algal medium. Figure 5 depicts the experimentation followed for the removal of CIP+NOR.

Experimentation for the removal of CIP+NOR.
Effect of CIP+NOR on algal growth
Algal cell density and chl-a were monitored every day until the algal species reached the death phase during their growth. Figure 6 compares the growth phase of algal species in the test runs with the control runs. The specific growth rate for control runs was 0.22 d−1 (Cv), 0.15 d−1 (Cp), 0.18 d−1 (So), 0.15 d−1 (T), and 0.14 d−1 (M) whereas for test runs it reached 0.28 d−1 (Cv), 0.2 d−1 (Cp), 0.23 d−1 (So), 0.17 d−1 (T), and 0.19 d−1 (M). The maximum growth rate was recorded for (So) 0.65 d−1 during day 4 of its exponential growth phase and the lowest growth rate was observed in the (Cv) (− 0.01 d−1) during day 6 (Fig. 6a), the negative growth rate indicates the decline in algal cell density due to the presence of the binary mixture, which affirms the acute toxicity assessment of (Cv) being the most sensitive species and (So) being the most tolerant species to the binary mixture of CIP+NOR. Dunnett tests were performed to compare the control runs and test runs in all the experimental runs to find the statistical significance. The generated p-values were 0.04 (Cv), 0.03 (Cp), 0.03 (So), 0.02 (T), and 0.04 (M). which were statistically significant at the (p < 0.05) level.

Effect of the binary mixture (CIP+NOR) on cell density and chl-a for (a) Cv, (b) Cp, (c) So, (d) T, and (e) M.
The growth pattern and the chl-a activity from the results indicate the algal ability to adapt to the stress conditions caused by the binary mixture of CIP+NOR. In all the species, the growth rate and chl-a were inhibited during the 2–3 days of its growth phase due to the acute stress. The cell density and chl-a activity increased after day 3 higher than the control runs affirming the growth-stimulating response of the algal species. For Scenedesmus obliquus (So) the photosynthetic activity was increasing even on day 7 (Fig. 6c), whereas in other species the chl-a activity dropped affirming the inhibitory test results of (So) adapting to the toxic conditions better than other selected species.
Removal mechanisms adapted by algal species
Biotic removal (bioadsorption, bioaccumulation, and biodegradation) and Abiotic removal (photodegradation) are the two major contributions to the removal of CIP+NOR in the medium. Abiotic removal is considered to assess the actual biotic removal by algae in the medium. It has been observed that abiotic removal of NOR (10%) > CIP (1.9%) after 7 days (Fig. 7f) which indicates that NOR is more sensitive towards photodegradation in comparison with CIP45, however, the removal through abiotic conditions is negligible in comparison with the biotic removal.

HPLC overlay detection peaks of binary composition (CIP+NOR) for (a) Cv, (b) Cp, (c) So, (d) T, (e) M, and (f) Abiotic removal.
Bioadsorption, bioaccumulation, and biodegradation are simultaneous events that occur in the algae cells for the assimilation of pollutants53,54. Bioadsorption is an extracellular mechanism that depends on the functional groups (proteins, cellulose, and hemicellulose) followed by bioaccumulation where the stress is counteracted by algae by the generation of reactive oxygen species (ROS) which leads to the biodegradation mechanism in algae53,54. Chl-a is a major light-harvesting pigment, during photosynthesis it releases protons inside the cell which enhances the production of reactive oxygen species (ROS) to counteract the stress caused by the pollutant55. This generated ROS in the algal cell takes part in the degradation of stress-causing pollutants in the algal medium56. The contribution of each mechanism in the removal of a binary mixture (CIP + NOR) is tabulated in Table 2. Figure 7 shows the consolidated overlay peak curves recorded in the HPLC analysis for the determination of removal mechanism adapted by different algal species
The removal efficiency of the binary mixture is in the order of So > Cp > T > M > Cv which is in line with the calculated 96 h EC50 order So > Cp > T > M > Cv. For Chlorella vulgaris (Cv), biodegradation (92%) is the major contributing removal mechanism for CIP, but NOR biodegradation is only 6% and 19% of NOR is accumulated inside the algal cell. The EC50 value indicated that Cv is more sensitive towards CIP compared to NOR, to counteract the stress caused, the reduction of CIP (55%) > NOR (25%) in the algal medium. Biodegradation contribution for the removal of CIP + NOR, in Chlorella pyrenoidosa (Cp) CIP (88%) > NOR (67%), Scenedesmus obliquus (So) CIP (88%) > NOR (87%), Tetradesmus sp (T) CIP (72%) < NOR (82%), and for Monoraphidium sp (M) CIP (87%) > NOR (84%). It is interesting to observe the biodegradation potential of Monoraphidium sp (M) is higher next to Scenedesmus obliquus (So) even though the acute toxicity studies showed that it is sensitive to the binary mixture.
Bioadsorption contribution for NOR is higher in the Chlorella species, 31% (Cv), and 14% (Cp) than in other species. This activity in Chlorella species can be evaluated by the log Kow value of NOR (0.46) which is higher than CIP (0.26), studies have reported that the higher the value greater the adsorption of pollutants onto the cells19,57. The bioadsorption contribution (%) may also vary according to the species assimilation and acclimatizing potential of algal species. In Tetradesmus sp (T), biodegradation contribution for NOR is 82% > CIP (72%) and bioaccumulation for NOR is below the detection limit (BDL). In Scenedesmus obliquus (So), the bioadsorption contribution for CIP is 6% and NOR is 0.39%, and it can be noted that biodegradation contribution for NOR is 87% which is higher than the other species tested and bioaccumulation of NOR is below the deduction limit of the equipment. It is evident from the obtained results that Scenedesmus obliquus is effective in removal of CIP + NOR.
Biotransformed products inside Scenedesmus obliquus
From the experimental reports, it can be inferred that Scenedesmus obliquus performed better in terms of tolerance (96 h EC50) and removal efficiency. The biotransformed products inside the Scenedesmus obliquus are assessed by LC–MS analysis (Fig. 7) and the possible biotransformed pathway is depicted in Fig. 8. The ROS generated inside the algal cells attack the quinolone moiety (carboxyl group), F-atom, and piperazine to breakdown the parent compound into less toxic as a response to the stress induced by the binary mixture. Decomposition of piperazinyl by Cytochrome P450 enzyme and defluorination along with the fragmentation of C5H5N2 from the CIP by the ROS resulted in the formation of 1-cyclopropyl-4-oxo-1,4-dihydroquinoline-3-carboxylic acid (C1) and degradation of the quinoline moiety resulted in the formation of quinolin-4(1H)-one (C2). 1-ethyl-4-oxo-1,4-dihydroquinoline-3-carbaldehyde (N1) and 1-methylquinolin-4(1H)-one (N2) are the two biotransformed products formed as the result of desethylation and defluorination of NOR inside algal cells. The proposed degradation pathway in Fig. 8 is consistent with the literature survey58,59,60.

Possible biotransformation pathway of CIP+NOR in Scenedesmus obliquus.
ECOSAR is employed to predict acute toxicity (96 h EC50) for biotransformed products, C1 (866 mg/L), C2 (156 mg/L), N1 (185 mg/L), and N2 (71 mg/L). The biotransformed products were less toxic compared to the parent compounds due to the loss of fluorine atom and piperazine ring from the compounds, as these substituents contribute to the antibacterial activity in Fluoroquinolone61. The presence of harmless biotransformed products inside Scenedesmus obliquus indicates the algal potential to treat these toxic compounds in wastewater matrices without the production of toxic by-products.
Biochemical composition assessment in all algal species
In all the experimental runs the accumulation of biomass (dry cell weight) was higher in the test runs than in the control runs. The biochemical composition (carbohydrates, lipids, and proteins) assessment was tabulated in Table 3. It can be noted from the table that Scenedesmus obliquus (So) has a higher lipid (35%), carbohydrate (18%), and protein (33%) accumulation compared to the control runs affirming the growth-stimulating response in the (So) species. In algal species (Cv, Cp, T, and M), being the sensitive species to the binary mixture the protein production decreased in comparison with the control runs. Algal cells convert the carbon source and CO2 into glyceraldehydes-3-phosphate (G3P) during photosynthesis by the action of chl-a activity62. G3P is responsible for the production of protein, carbohydrates, and lipid accumulation inside the cell, especially protein and lipids are the results of the generation of acetyl-CoA and pyruvate from G3P, when a toxic pollutant is accumulated inside algae cells, the cells counteract the stress conditions by increasing the production of lipids as a defense mechanism63. The increased production of lipids than the protein in all the algal species is due to the result of the stress induced by CIP+NOR.
Comparing the biochemical composition of biomass in all the algal species. (So) has higher lipid accumulation with good removal efficiency and it can also tolerate the stress caused by the binary mixture. Scenedesmus obliquus is the most suitable candidate among these tested algal species in the treatment of antibiotic-polluted water with the advantage of valuable biomass production.
Conclusion
In this experimental study, inhibitory tests were performed for Chlorella vulgaris (Cv), Chlorella pyrenoidosa (Cp, Scenedesmus obliquus (So), and Tetradesmus sp (T), and Monoraphidium (M) by spiking the algal medium with the binary mixture (CIP+NOR). Acute toxicity assessment revealed that Scenedesmus obliquus is the least sensitive algal species with 49% inhibition at 50 mg/L. Furthermore, removal studies revealed that Scenedesmus obliquus performed better in removing both pollutants with biodegradation as the major removal mechanism. LC–MS results revealed the biotransformed products are less toxic than the parent compound. Biochemical composition (protein, carbohydrates, and lipids) analysis revealed growth stimulating effect was observed higher in (So), this effect contributes an advantage in the utilization of biomass after the treatment for the production of value-added bioproducts.
Scenedesmus obliquus cannot be employed when the binary mixture concentration in the medium is higher than its tolerance level. However, there is a need to optimize and evaluate the performance of Scenedesmus obliquus in a pilot-scale study (real wastewater conditions). In addition, the formation of biotransformed products in the presence of other pollutants in the real wastewater need to be investigated.
Data availability
All the data generated or analyzed during this study are included in this article.
References
Khouja, T., Mitsantisuk, K., Tadrous, M. & Suda, K. J. Global consumption of antimicrobials: Impact of the WHO global action plan on antimicrobial resistance and 2019 coronavirus pandemic (COVID-19). J. Antimicrob. Chemother. 77, 1491–1499 (2022).
Danner, M., Robertson, A., Behrends, V. & Reiss, J. Antibiotic pollution in surface fresh waters: Occurrence and effects. Sci. Total Environ. 664, 793–804 (2019).
Laxminarayan, R. et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis https://doi.org/10.1016/S1473-3099(13)70318-9 (2013).
Laxminarayan, R. & Chaudhury, R. R. Antibiotic resistance in India: Drivers and opportunities for action. PLoS Med. https://doi.org/10.1371/journal.pmed.1001974 (2016).
Hendriksen, R. S. et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 10, 1124 (2019).
Ricky, R. & Shanthakumar, S. Phycoremediation integrated approach for the removal of pharmaceuticals and personal care products from wastewater: A review. J. Environ. Manag. 302, 113998 (2022).
Kümmerer, K. Antibiotics in the aquatic environment: A review—Part I. Chemosphere 75, 417–434 (2009).
Cherian, J. J. et al. India’s road to independence in manufacturing active pharmaceutical ingredients: Focus on essential medicines. Economies 9, 71 (2021).
Bhagat, C., Kumar, M., Tyagi, V. K. & Mohapatra, P. K. Proclivities for prevalence and treatment of antibiotics in the ambient water: A review. npj Clean Water 3, 42 (2020).
Gou, Y. et al. Degradation of fluoroquinolones in homogeneous and heterogeneous photo-Fenton processes: A review. Chemosphere 270, 129481 (2021).
Van Doorslaer, X., Dewulf, J., Van Langenhove, H. & Demeestere, K. Fluoroquinolone antibiotics: An emerging class of environmental micropollutants. Sci. Total Environ. 500–501, 250–269 (2014).
Yang, Q. et al. Antibiotics: An overview on the environmental occurrence, toxicity, degradation, and removal methods. Bioengineered 12, 7376–7416 (2021).
Maia, A. S. et al. Degradation of fluoroquinolone antibiotics and identification of metabolites/transformation products by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 1333, 87–98 (2014).
Jia, A., Wan, Y., Xiao, Y. & Hu, J. Occurrence and fate of quinolone and fluoroquinolone antibiotics in a municipal sewage treatment plant. Water Res. 46, 387–394 (2012).
Abinandan, S. & Shanthakumar, S. Challenges and opportunities in application of microalgae (Chlorophyta) for wastewater treatment: A review. Renew. Sustain. Energy Rev. 52, 123–132 (2015).
Xiong, J. Q., Kurade, M. B., Kim, J. R., Roh, H. S. & Jeon, B. H. Ciprofloxacin toxicity and its co-metabolic removal by a freshwater microalga Chlamydomonas mexicana. J. Hazard. Mater. 323, 212–219 (2017).
Zhang, J., Fu, D. & Wu, J. Photodegradation of Norfloxacin in aqueous solution containing algae. J. Environ. Sci. 24, 743–749 (2012).
Pan, L.-J. et al. Study of ciprofloxacin biodegradation by a Thermus sp. isolated from pharmaceutical sludge. J. Hazard. Mater. 343, 59–67 (2018).
Ricky, R., Chiampo, F. & Shanthakumar, S. Efficacy of ciprofloxacin and amoxicillin removal and the effect on the biochemical composition of Chlorella vulgaris. Bioengineering 9, 134 (2022).
Hena, S., Gutierrez, L. & Croué, J. P. Removal of pharmaceutical and personal care products (PPCPs) from wastewater using microalgae: A review. J. Hazard. Mater. 403, 124041 (2021).
Sengar, A. & Vijayanandan, A. Human health and ecological risk assessment of 98 pharmaceuticals and personal care products (PPCPs) detected in Indian surface and wastewaters. Sci. Total Environ. 807, 150677 (2022).
Välitalo, P., Kruglova, A., Mikola, A. & Vahala, R. Toxicological impacts of antibiotics on aquatic micro-organisms: A mini-review. Int. J. Hyg. Environ. Health 220, 558–569 (2017).
OECD. Test No. 201: Alga, Growth Inhibition Test (OECD Publishing, 2011). https://doi.org/10.1787/9789264069923-en.
Dommange, X., Tanguy, P. A. & Jolicoeur, M. Feasibility of lipid mechanical extraction from viable Monoraphidium minutum. Microalgae Biotechnol. 1, 12–19 (2015).
He, Q., Yang, H., Wu, L. & Hu, C. Effect of light intensity on physiological changes, carbon allocation and neutral lipid accumulation in oleaginous microalgae. Bioresour. Technol. 191, 219–228 (2015).
Ravi Kiran, B. & Venkata Mohan, S. Phycoremediation potential of Tetradesmus sp. SVMIICT4 in treating dairy wastewater using flat-panel photobioreactor. Bioresour. Technol. 345, 126446 (2022).
Bibi, F. et al. Production of lipids biosynthesis from Tetradesmus nygaardii microalgae as a feedstock for biodiesel production. Fuel 326, 124985 (2022).
Hom-Diaz, A., Norvill, Z. N., Blánquez, P., Vicent, T. & Guieysse, B. Ciprofloxacin removal during secondary domestic wastewater treatment in high rate algal ponds. Chemosphere https://doi.org/10.1016/j.chemosphere.2017.03.125 (2017).
Hom-Diaz, A. et al. Insights into removal of antibiotics by selected microalgae (Chlamydomonas reinhardtii, Chlorella sorokiniana, Dunaliella tertiolecta and Pseudokirchneriella subcapitata). Algal Res. 61, 102560 (2022).
Raju, C. B. V. N., Sharma, H. K., Rao, S. C. & Rao, G. RP-HPLC method for analysis of related substances in amoxicillin drug substance. Acta Chromatogr. 21, 57–70 (2009).
Pistos, C., Tsantili-Kakoulidou, A. & Koupparis, M. Investigation of the retention/pH profile of zwitterionic fluoroquinolones in reversed-phase and ion-interaction high performance liquid chromatography. J. Pharm. Biomed. Anal. 39, 438–443 (2005).
Meena, N. S., Sahni, Y. P., Shrman, K., Singh, A. K. & Kumar, A. Detection of norfloxacin in muscle, liver and kidney of broiler chicken. Indian J. Anim. Res. 54, 739–743 (2020).
Anjali, R. & Shanthakumar, S. Simultaneous degradation of amoxicillin, ciprofloxacin and acetaminophen in a mixture by ozonation: Kinetics and mechanisms pathway. J. Clean. Prod. 378, 134509 (2022).
Turner, M. Microalgae—Biotechnology and Microbiology. Journal of Experimental Marine Biology and Ecology Vol. 183 (Cambridge University Press, 1994).
Ricky, R., Shanthakumar, S. & Gothandam, K. M. A pilot-scale study of the integrated phycoremediation-photolytic ozonation based municipal solid waste leachate treatment process. J. Environ. Manag. 323, 116237 (2022).
Chen, Y. & Vaidyanathan, S. Simultaneous assay of pigments, carbohydrates, proteins and lipids in microalgae. Anal. Chim. Acta 776, 31–40 (2013).
Bligh, E. G. & Dyer, W. J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917 (1959).
Krishna Moorthy, A., Govindarajan Rathi, B., Shukla, S. P., Kumar, K. & Shree Bharti, V. Acute toxicity of textile dye methylene blue on growth and metabolism of selected freshwater microalgae. Environ. Toxicol. Pharmacol. 82, 103552 (2021).
Wang, J. & Wang, S. Toxicity changes of wastewater during various advanced oxidation processes treatment: An overview. J. Clean. Prod. 315, 128202 (2021).
Magdaleno, A., Saenz, M. E., Juárez, A. B. & Moretton, J. Effects of six antibiotics and their binary mixtures on growth of Pseudokirchneriella subcapitata. Ecotoxicol. Environ. Saf. 113, 72–78 (2015).
Xiong, J. Q., Kurade, M. B. & Jeon, B. H. Biodegradation of levofloxacin by an acclimated freshwater microalga, Chlorella vulgaris. Chem. Eng. J. 313, 1251–1257 (2017).
Xiong, J.-Q. et al. Biodegradation of carbamazepine using freshwater microalgae Chlamydomonas mexicana and Scenedesmus obliquus and the determination of its metabolic fate. Bioresour. Technol. 205, 183–190 (2016).
Bai, X. & Acharya, K. Removal of trimethoprim, sulfamethoxazole, and triclosan by the green alga Nannochloris sp. J. Hazard. Mater. 315, 70–75 (2016).
Ding, T. et al. Toxicity, degradation and metabolic fate of ibuprofen on freshwater diatom Navicula sp. J. Hazard. Mater. 330, 127–134 (2017).
Babić, S., Periša, M. & Škorić, I. Photolytic degradation of norfloxacin, enrofloxacin and ciprofloxacin in various aqueous media. Chemosphere 91, 1635–1642 (2013).
Ortiz de García, S. A., Pinto Pinto, G., García-Encina, P. A. & Irusta-Mata, R. Ecotoxicity and environmental risk assessment of pharmaceuticals and personal care products in aquatic environments and wastewater treatment plants. Ecotoxicology 23, 1517–1533 (2014).
Nie, X., Wang, X., Chen, J., Zitko, V. & An, T. Response of the freshwater alga Chlorella vulgaris to trichloroisocyanuric acid and ciprofloxacin. Environ. Toxicol. Chem. 27, 168–173 (2008).
Geiger, E., Hornek-Gausterer, R. & Saçan, M. T. Single and mixture toxicity of pharmaceuticals and chlorophenols to freshwater algae Chlorella vulgaris. Ecotoxicol. Environ. Saf. 129, 189–198 (2016).
Eguchi, K. et al. Evaluation of antimicrobial agents for veterinary use in the ecotoxicity test using microalgae. Chemosphere 57, 1733–1738 (2004).
Tang, D. Y. Y. et al. Triphasic partitioning of mixed Scenedesmus and Desmodesmus for nutrients’ extraction and chlorophyll composition prediction for algae bloom. Environ. Technol. (United Kingdom) https://doi.org/10.1080/09593330.2022.2150094 (2022).
Wan, L., Wu, Y., Ding, H. & Zhang, W. Toxicity, biodegradation, and metabolic fate of organophosphorus pesticide trichlorfon on the freshwater algae Chlamydomonas reinhardtii. J. Agric. Food Chem. 68, 1645–1653 (2020).
Xiong, J. Q. et al. Toxicity of sulfamethazine and sulfamethoxazole and their removal by a green microalga, Scenedesmus obliquus. Chemosphere 218, 551–558 (2019).
Yu, Y. et al. Investigation of the removal mechanism of antibiotic ceftazidime by green algae and subsequent microbic impact assessment. Sci. Rep. 7, 4168 (2017).
Lau, Z. L. et al. A review on the diverse interactions between microalgae and nanomaterials: Growth variation, photosynthetic performance and toxicity. Bioresour. Technol. 351, 127048 (2022).
Liu, X., Wu, F. & Deng, N. Photoproduction of hydroxyl radicals in aqueous solution with algae under high-pressure mercury lamp. Environ. Sci. Technol. 38, 296–299 (2004).
Bai, X. & Acharya, K. Algae-mediated removal of selected pharmaceutical and personal care products (PPCPs) from Lake Mead water. Sci. Total Environ. 581–582, 734–740 (2017).
Rogers, H. R. Sources, behaviour and fate of organic contaminants during sewage treatment and in sewage sludges. Sci. Total Environ. 185, 3–26 (1996).
Feng, N. X. et al. Co-metabolic degradation of the antibiotic ciprofloxacin by the enriched bacterial consortium XG and its bacterial community composition. Sci. Total Environ. 665, 41–51 (2019).
Xu, D. et al. Enhanced molecular oxygen activation on (001) facets of Zn-doped BiOCl nanosheets for ciprofloxacin degradation. Adv. Mater. Interfaces 7, 2000548 (2020).
Zhao, R. et al. Efficient enzymatic degradation used as pre-stage treatment for norfloxacin removal by activated sludge. Bioprocess Biosyst. Eng. 40, 1261–1270 (2017).
Amorim, C. L., Moreira, I. S., Maia, A. S., Tiritan, M. E. & Castro, P. M. L. Biodegradation of ofloxacin, norfloxacin, and ciprofloxacin as single and mixed substrates by Labrys portucalensis F11. Appl. Microbiol. Biotechnol. 98, 3181–3190 (2014).
Debnath, C. et al. Microalgae: Sustainable resource of carbohydrates in third-generation biofuel production. Renew. Sustain. Energy Rev. 150, 111464 (2021).
Alishah Aratboni, H., Rafiei, N., Garcia-Granados, R., Alemzadeh, A. & Morones-Ramírez, J. R. Biomass and lipid induction strategies in microalgae for biofuel production and other applications. Microb. Cell Fact. 18, 178 (2019).
Acknowledgements
The authors would like to thank Vellore Institute of Technology (VIT) for supporting this work.
Author information
Authors and Affiliations
Contributions
R.R.: Formal analysis, Investigation, Figures, Writing—original and revised draft. S.S.: Methodology, Supervision, Conceptualization, Writing—Review & Editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ricky, R., Shanthakumar, S. An investigation on removal of ciprofloxacin and norfloxacin by phycoremediation with an emphasis on acute toxicity and biochemical composition. Sci Rep 13, 13911 (2023). https://doi.org/10.1038/s41598-023-41144-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-41144-y
- Springer Nature Limited