
Abstract
North Africa is a key area for understanding hominin population movements and the expansion of our species. It is home to the earliest currently known Homo sapiens (Jebel Irhoud) and several late Middle Stone Age (MSA) fossils, notably Kébibat, Contrebandiers 1, Dar-es-Soltane II H5 and El Harhoura. Mostly referred to as “Aterian” they fill a gap in the North African fossil record between Jebel Irhoud and Iberomaurusians. We explore morphological continuity in this region by quantifying mandibular shape using 3D (semi)landmark geometric morphometric methods in a comparative framework of late Early and Middle Pleistocene hominins (n = 15), Neanderthals (n = 27) and H. sapiens (n = 145). We discovered a set of mixed features among late MSA fossils that is in line with an accretion of modern traits through time and an ongoing masticatory gracilization process. In Northern Africa, Aterians display similarities to Iberomaurusians and recent humans in the area as well as to the Tighenif and Thomas Quarry hominins, suggesting a greater time depth for regional continuity than previously assumed. The evidence we lay out for a long-term succession of hominins and humans emphasizes North Africa’s role as source area of the earliest H. sapiens.
Similar content being viewed by others


Introduction
For most of the Plio-Pleistocene Africa has been the core continent of hominin evolution. Genetic and fossil evidence suggest that the ancestry of all people living today can be traced back to Africa1,2,3, but little is known about its population dynamics. Situated at a shifting interface between Palaearctic and Afrotropical ecozones, the northern part of the continent played a critical part in H. sapien's expansion and represents a strategic area for encounters between Middle Pleistocene hominins. The redating of the Kabwe4 and Jebel Irhoud fossils to around 300 ka BP5 questions the direct descent of our lineage from a group of African large-brained Middle Pleistocene specimens either classified as H. rhodesiensis or Homo heidelbergensis s.l., namely Kabwe (Broken Hill), Bodo, and Hopefield6,7. With the evidence from Jebel Irhoud3, North African late Early/Middle Pleistocene hominins from Tighenif or Thomas Quarry became potential candidates for the ancestral morphology of Neanderthals and H. sapiens7,8. Yet, regional continuity within Africa has been hotly debated given a sparse fossil record and complex environmental dynamics9,10.
The aim of our study is to “fill a gap” between the Jebel Irhoud and Iberomaurusian humans with the first detailed morphometric analysis of four Moroccan mandibles dated to MIS 6-4 (Fig. 1). As mandibular remains carry phylogenetic signals11 and are numerous in paleoanthropological contexts, these specimens give us insight into people living at the time of the last out-of-Africa dispersal12,13. Three of the studied specimens were associated with the “Aterian”14, a typological variant of the Maghrebian MSA lithic industries that dominates the archaeological record west of the Nile Valley between 14515 and 30 ka BP. As early as 142 ka BP, Aterian material culture documents the emergence of modern behavior14,16 in the form of perforated and ochre-decorated Tricia (Nassarius) shells17, hearths and stone-walled structures18 as well as formal bone tools19,20,21. The three hereafter referred to as Aterian individuals (Supplementary Table S1) were recovered in close vicinity to each other from cave sites in littoral Maghreb, namely Grotte des Contrebandiers, Dar-es-Soltane II and El Harhoura 1. Likewise of late MSA origin, the fossil mandible from Kébibat will be referred to without group affiliation as it lacks any archaeological information.

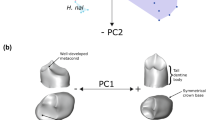
Kébibat and Aterian mandibular specimens.
While early H. sapiens from South Africa display a rather large variation22,23, the Jebel Irhoud and Aterian humans share a robust craniofacial/mandibular morphology with late Early/Middle Pleistocene hominins from North Africa24,25,26, in particular a small number of archaic features. Broad braincases and faces with well defined superstructures in the Jebel Irhoud, Contrebandiers and Dar-es-Soltane II 5 crania were reported as intermediate between preceding (Tighenif, Kébibat) and subsequent groups (Iberomau-rusians)24,26. A similar pattern of progressive evolution was proposed for the associated mandibles (Tighenif, Thomas Quarry, Sidi Abderrhaman, Kébibat, Jebel Irhoud, Contrebandiers)27, but challenged by the presence of a modern-looking face and high cranial capacity in the Jebel Irhoud humans3 that contrast the geologically younger but more archaic morphology of Kébibat27 (140 ka) and the smaller cranial capacity of the older Salé cranium (400 ka)28. As indicated above, the massive Iberomaurusian crania have been sometimes compared to Aterian and Jebel Irhoud skulls from the same area24,26. However, the apparent archaeological hiatus at the Middle/Later Stone Age transition29,30 renders population continuity between Aterians and Iberomaurusians an unresolved case.
Geological context of late MSA sites in Morocco and description of associated fossils
In 1933, M. Allenda, who supervised communal works in Kébibat, a suburb of Rabat, discovered a fossil mandible in a pile of cobbles intended for roadworks31. Thereupon he located the reburied remains of this adolescent individual (15–16 yrs) in the coastal quarry Mifsud-Giudice, consisting of 23 cranial fragments and an incomplete left maxilla, heavily damaged by a detonation and without any associated archaeological material. The geologic provenance of this so-called “Rabat Man” could be tracked via an imprint of the palate and its molars in the calcarenite limestone. The associated sedimentary unit 2 from sequence 2 was dated by Infrared Stimulated Luminescence (IRSL) to 137 ± 7 ka BP32, which is in line with previous dating attempts of an underlying marine shell layer33. The Kébibat human remains (Fig. 1) have been attributed to a population featuring a mixture of archaic and modern traits8,27,28, in accord with the mosaic character of human morphology at that time11.
After J. Roche had conducted some test excavations in the Smuggler's Cave (Grotte des Contrebandiers) between 1955 and 1957 a human mandible (Fig. 1)—hereafter referred to as Contrebandiers 1—was recovered in layer 9, OSL-dated to 111–92 ka BP34. This layer corresponds to the lower part of layer 4 in the central area of recent excavations35 and exposed another human calvaria just before the end of Roche’s excavations in 1975. The resumption of Roche’s excavations has been directed by M. A. El Hajraoui and H. Dibble (deceased 2018) since 200618. Due to its high robusticity in bone and dentition Contrebandiers 1, consisting of an incomplete corpus and three ramus fragments18,36, was initially thought to originate from an Acheulian deposit37. Accordingly, it displays an inferior transverse torus, a prognathic and vertical symphysis as well as a mental foramen situated under P4/M1. In contrast, the anteroposterior decreasing corpus height represents a H. sapiens characteristic.
Near Rabat, a partial cranium with the left half of the face (H5), an adolescent mandibular corpus (H4) and an infant calvaria (H3) were excavated by A. Debénath in 1975 in the cave of Dar-es-Soltane II24. The specimens came from an archaeologically sterile marine sand deposit (unit 7), immediately overlain by an Aterian MSA (Middle Stone Age) layer38. Based on OSL, an age possibly beyond 100 ka was proposed for Unit 739. Amino acid racemization on Patella shells calibrated by C14/AMS rather point to a maximum age of 85–75 ka BP40. The morphology of individual H5 has been interpreted as intermediate between Jebel Irhoud and Iberomaurusians24,41, exhibiting extraordinary breadth dimensions, marked brow ridges and a high robusticity. Some facial structures were even described as Neanderthal-like. Yet, the associated left hemimandible, hereafter referred to as Dar-es-Soltane II 5, lacks a retromolar gap and its large mental foramen is under P4 (Fig. 1). It belongs to a mature individual and is one of the largest specimens studied. The ramus is wide with an expanded gonial area showing medial pterygoid tubercles, but also a marked masseteric fossa. Despite its unpreserved inferior symphysis, a curved mental profile is discernible.
In the second archaeological layer (see Fig. 7 in18) of the Zouhrah Cave (El Harhoura 1), a large and robust mandibular corpus (Fig. 1) and a large isolated canine were unearthed during a salvage excavation in 1977/197825,42. Burnt sandstone structures in the underlying layer 1 date to 32,150 ± 4800 BP by ƴ TL43. The relatively young C14 date of 25,580 ± 130 BP on Helix shells44 from layer 1 or 2 has been suspected to result from secondary colonization by these gastropods40. Unpublished Uranium–Thorium dates of a spelothem that caps layer 2 rather indicate that the El Harhoura mandible has a minimum age of approximately 66 ka BP. Despite such relatively recent age and the presence of a human chin, it displays a uniform corpus height, a prognathic and U-shaped dental arcade as well as an inferior transverse torus.
Information on North African late Early/Middle Pleistocene sites in the current study
Between 1969 and 2008, the sandstone quarries of Thomas I and III near Casablanca (Fig. 2) yielded several craniofacial and dental remains of fossil hominins associated with Acheulian lithics and abundant fauna45. Thomas I yielded an eponymous left corpus fragment46,47, a complete jaw (hereafter referred to as Thomas Gh10717) and a juvenile right corpus fragment. Geochronology, biostratigraphy and ESR dating of tooth enamel point towards an early Middle Pleistocene age of around 700–600 ka BP45,48,49,50.

Map with North African sites mentioned in the text. 1 Sidi Abderrahman, 2 Thomas Quarries, 3 Grotte des Contrebandiers, 4 El Harhoura II, 5 Dar-es-Soltane, 6 Kébibat, 7 Salé, 8 Jebel Irhoud, 9 Taforalt, 10 Tighenif, 11 Afalou Bou Rhummel. Topographic map taken from https://www.naturalearthdata.com/ and edited in PDF-XChange Editor v. 8 (PDF-XChange Co. Ltd, Bolney).
From 1954 to 1956 the Tighenif quarry east of Mascara in Algeria (Fig. 2), previously known as Ternifine or Palikao51, yielded two nearly complete jaws (Tighenif 1 + 3) and a hemimandible (Tighenif 2). Geological and micromammalian evidence indicate a short site formation process characterized by a seasonal lake in an open environment52. Biostratigraphy and paleomagnetism initially placed the site close to the lower Middle Pleistocene boundary around 700 ka BP, its lithic assemblage was assigned to the late Lower Acheulean. After biostratigraphic revision, a late Early Pleistocene age within the Jaramillo Event around 1 Ma has been suggested53. This seems to be confirmed by a reevaluation of the faunal association54,55,56. For data analysis, we group the Tighenif fossils with the African Middle Pleistocene.
Objectives and working hypotheses
Not all of the early H. sapiens mandibles fall into the morphological range of recent modern humans3,11, making it seem likely that the emergence of diagnostic H. sapiens features follows an accretional pattern. If modern mandibular morphology emerged by such a shift in the frequency of modern traits through time, we expect a mix between archaic and modern traits in all four late MSA mandibles from Morocco (Fig. 1). Aterian remains have been described as less archaic than the earliest currently known H. sapiens from Jebel Irhoud, but not as modern as Iberomaurusian or European Upper Paleolithic groups23,24,41. In this regard, Aterian mandibular shape and size might fill a gap in the human fossil record that currently exists between early and later H. sapiens3,11. The isolated Kébibat hominin has been classified as a morphological link between archaic and modern populations in Africa8. The occurence of this non-modern specimen well after the emergence of the first H. sapiens3,5, reflects a non-linear evolution towards H. sapiens and may indicate rapid shifts in the frequency of certain traits.
Apart from the Jebel Irhoud and Aterian humans, the Maghreb provided fossils of late Early/Middle Pleistocene hominins (Fig. 2), offering unique insights into early phases of H. sapiens evolution. Genetic evidence suggests regional continuity in the Maghreb since the Epipaleolithic57,58, but biological exchanges with the Eastern Mediterranean Basin and the Sahelian zone to the south are documented as well57,59.Craniofacial studies question a progressive evolution towards H. sapiens3,28,41, but disclose notable similarities between Middle and Late Pleistocene populations in the area24,25,26,27. In order to track the evolution of Northern African populations and potential influences from adjacent areas during the Middle and Late Pleistocene, we quantify mandibular variation in Kébibat, Contrebandiers 1, Dar-es-Soltane II 5 and El Harhoura along with late Early/Middle Pleistocene hominins, Neanderthals, early and later H. sapiens (Supplementary S1, S2, Figs. S1 and S2, Table S1).
We use 3D geometric morphometrics to perform principal component analyses in Procrustes shape space and to visualize group mean shapes as well as differences in absolute mandibular dimensions. The strength of this approach is the perspective on mandibular shape separately from size, allowing us to track shape continuity across different time periods. The complementary record of discrete mandibular traits provides insights into differences between individuals.
Results and discussion
In the complete mandible data set Principal Component (PC) 1 accounts for 29.7% of the total variation and reveals substantial overlap between groups (Supplementary Fig. S3). Along PC 2, accounting for 18.9% of total variation, archaic groups (late Early/Middle Pleistocene hominins and Neanderthals) separate well from most H. sapiens, however, early H. sapiens and ancient sub-Saharans overlap with both clusters (Fig. 3a). The Tighenif mandibles are distinct from Neanderthals. European Middle Pleistocene individuals and Neanderthals fall towards the negative end of PC 2 while Holocene humans and Iberomaurusians plot towards the positive extreme. Natufian and Upper Paleolithic specimens attain intermediate scores, but intersect considerably the former two groups. In the corpus data set archaic groups separate best from H. sapiens along PC1 (Fig. 3b), accounting for 34.1% of the total variation. Again, early H. sapiens and ancient sub-Saharans fall in the middle. A three-dimensional visualization of group mean shapes (Fig. 4) supports the intermediate position of early H. sapiens disclosing a morphological succession in time from African Middle Pleistocene hominins to later H. sapiens. Except for Jebel Irhoud 113 (Table 1; Supplementary Tables S3–S7), early H. sapiens display a reduction in dental arcade length, gonial area, ramus breadth and coronoid size, pushing back the onset of masticatory gracilization11 to the Middle/Late Pleistocene transition. This process persists into the Holocene, explaining why Holocene people feature on average the smallest mandibles followed by Natufian, ancient sub-Saharan and Upper Paleolithic groups (Fig. 5; Supplementary Tables S3–S7). Iberomaurusians are clearly larger than their contemporaries, covering a similar size range as Aterians and early H. sapiens. Out of all samples, African Middle Pleistocene mandibles show on average the largest dimensions.

Principal component (PC) analyses of the pooled samples of the mandible (n = 166) and corpus (n = 192) data sets in shape space. Warps of PC extremes are displayed in grey (PC1), in bone color (PC2) and red (PC3) for ± 3 SD. They illustrate shape changes that drive mandibular variation along each PC.

Mean shapes of African late Early/Middle Pleistocene hominins, early Homo sapiens, and later H. sapiens (Iberomaurusians/Natufians/ancient sub-Saharans/Upper Paleolithic/Holocene) warped onto a 3D surface model of an individual from the respective group (a) and as wireframes (b).

Box plots depicting centroid size for each group in each data set. Horizontal lines represent the median of each group. Boxes show the interquartile range (IQR, 25th to 75th percentile). Whiskers extend to 1.5 times IQR, but minimum to the lower/upper 25% of the data.
Procrustes nearest neighbors of Maghrebian late Early/Middle Pleistocene mandibles (Tighenif, Thomas Quarry) disclose shape affinities to Iberomaurusians and recent humans from the region (Supplementary Table S2). They also reveal a close relatedness between Tighenif 2, Irhoud 11 and Dar-es-Soltane II 5 as well as between Irhoud 11 and Thomas Gh10717 – visualized by three-dimensional superimpositions (Figs. 6 and 7). These results indicate morphological continuity in the Maghreb, substantiated by numerous archaic traits that late Early/Middle Pleistocene individuals (Tighenif, Thomas Quarry, Kébibat) share with Jebel Irhoud 11 and Aterians (Table 1). One third of the closest Procrustes neighbors to Dar-es-Soltane II 5 are Neanderthals (Table S2), reflecting PCA results (Fig. 3a). According to these lines of evidence, Dar-es-Soltane II 5 is more similar in mandibular shape to Neanderthals than to Iberomaurusians. Procrustes nearest neighbors (Supplementary Table S2) and a 3D superimposition of El Harhoura versus Taforalt XVIII (Fig. 7) partially support a morphologic link between Aterian and Iberomaurusian crania24,26. Iberomaurusian mandibles were distinct from penecontemporaneous sub-Saharan specimens, namely Ishango, Jebel Sahaba, Mumbwa 3, Olduvai 1, Gobero, Asselar, and Shum Laka (Fig. 3a).

Superimpositions in Procrustes space of the reconstructed Tighenif 2 (dark gray), Kébibat (orange), the reconstructed Dar-es-Soltane II 5 (green), and Taforalt XX (blue).

Superimpositions in Procrustes space of the original Irhoud 11 (pink), Thomas I Gh10717 (white), Dar-es-Soltane II 5 (green), El Harhoura (violet), Taforalt XVIII (turquoise), and a reconstructed version of Irhoud 11 (pink transparent).
Kébibat has been recently classified as a phylogenetic link between North African archaic and modern populations, exhibiting diagnostic H. sapiens traits on the cranium, but lacking them in the mandible8. We also identified the proposed high robusticity of Kébibat, its anteroposteriorly uniform corpus height, vertical symphysis and lack of chin. Saban27 disclosed a considerable number of archaic characteristics that Kébibat shares with the much older mandibular fragments from Sidi Abderrahman (400 ka BP60) and the younger dated Contrebandiers 1 mandible, supported by our data (Table 1, Supplementary Table S2). Our PCA (Fig. 3b) substantiates the view that the “Rabat-Kébibat Man” joins a complex transition from archaic to modern populations in Africa, previously indicated by its cranial morphology27,28. As many diagnostic parts of the mandible are missing (retromolar area, gonion, mandibular notch) and others are damaged (symphysis, dental arcade), we could not conclusively assign Kébibat to either early H. sapiens or an archaic Middle Pleistocene form. The mix between archaic and modern traits persists throughout the Aterian sample and is consistent with a mosaic evolution of the H. sapiens lineage24,26,27. At the same time there is a discernable shift in the frequency of modern traits, visible as differences between individuals (Table 1, Fig. 3) and likely due to an accretion of modern traits through time. This gradual accretion develops non-linearly through time with modern characters carried by individuals dated earlier, archaic characters carried by more recent specimens3,28, and a variable mix of features among penecontemporaneous specimens. For example, despite a large temporal distance between them and the presence of archaic traits, Irhoud 11 and Dar-es-Soltane II 5 exhibit a strong anteroposterior height decrease in their mandibular corpus (Figs. 1 and 7). In contrast, the younger dated fossil from El Harhoura 1 completely lacks this distinct modern feature and even displays an inferior transverse torus; at the same time it has a full human chin. Such mixed morphology is corroborated by patterns of dental morphology that Aterians share with early H. sapiens from the Levant23, North3 and East Africa61. In particular, megadontia expressed in the development of mass-additive traits on the teeth7,23 and of large dental root dimensions3,62 are reminiscent of archaic hominins whereas dental tissue proportions and root shape fall already into the range of modern human variation.
From a regional perspective, resemblances in mandibular shape (Supplementary Table S2, Figs. 6 and 7) and discrete features (Table 1) indicate that the Tighenif, Thomas Quarry and Kébibat hominins were part of the same evolving lineage as the Jebel Irhoud humans, Aterians, Iberomaurusians and recent North Africans. Absolute sizes of Aterian mandibles are in the range of early H. sapiens and Iberomaurusians (Fig. 5). Even though we have no proof of an in-situ population succession, Aterian morphology fits the human fossil gap between Jebel Irhoud 11 and Iberomaurusians, suggesting a greater time depth for regional continuity in Northern Africa than previously established57,58. The archaeological hiatus at the Middle/Later Stone Age transition29,30 might result from a demographic bottleneck, but not from a population replacement of Aterians by Iberomaurusians. Yet, a morphological link of Dar-es-Soltane II 5 to the Iberomaurusians, as suggested by Ferembach24, remains vague as this group is only distantly related in mandibular shape (Table S2, Fig. 3). This concurs with a facial analysis of Dar-es-Soltane II 541, yet we agree with the authors that it might also be obscured by the large size of the specimen.
A previous allometric study on human mandibles had revealed that some aspects of shape variation in adults correlate with mandibular size of the individual11. In this context, most Neanderthal-like shape features in early H. sapiens could be attributed to their large size. Interestingly, two large Aterian specimens in the current study carry similar features, namely a prognathic U-shaped dental arcade (El Harhoura), and a wide ramus with a large gonial angle as well as a wide bicondylar breadth (Dar-es-Soltane II 5). As Aterians chronologically align more with early H. sapiens than with later groups of humans, we assume similar allometric constraints . The amounts of shape variance explained by mandibular size in early H. sapiens (10.2%) and the pooled sample (4.6%) are alike11 (Table 2). This suggests that the Neanderthal-like appearance of Dar-es-Soltane II 5 (Fig. 3a; Table S2) might be linked to its large mandibular size (Supplementary Table S5). Bicondylar distance in Dar-es-Soltane II 5 even exceeds Neanderthal/European Middle Pleistocene means (Supplementary Tables S6, S8). Cranial and facial morphology are close to the Irhoud humans24,41, a sample that also shows size-driven Neanderthal-like morphology11. Likewise, the prognathic and U-shaped El Harhoura corpus (Table 1, Fig. 1) correlates with an exceptional size (Fig. 5).
In principle, the presence of diagnostic H. sapiens features (anteroposteriorly decreasing corpus height, incipient chin) throughout the Aterian sample allows us to group it within a single evolving lineage. Aterians fill a temporal gap in the global human fossil record between early and later H. sapiens, their heterogenous mandibular shape demonstrates an accretional pattern for the emergence of modern morphology. Apart from regional continuity, Aterians and other ancient North Africans (Tighenif, Thomas Quarry I) resemble Natufians, sub-Saharans and Upper Paleolithic people (Supplementary Table S2), shedding some light on the nature of potential exchanges between North Africa and adjacent areas. Genetically, this finding parallels a close relatedness of Iberomaurusians to Natufians, southern Europeans57, and—on a smaller scale—to sub-Saharan Africans59. To that end, late glacial back-to-Africa migrations via the Mediterranean63,64,65,66,67 and via the Near-East59,68 offer explanatory scenarios. Such population movements depended either on low sea-levels during glacial periods54,69 or on the periodic emergence of green corridors throughout the Sahara, Sinai, Negev and Nafud deserts70,71,72,73. The latter also allowed for human exchanges between North Africa and the Sahelian zone, accounting for the exceptional skeletal variation and/or signs of gene flow74,75 in most finds from the African humid period (Later Stone Age until mid-Holocene). Among our samples, especially the El Harhoura mandible matches substantially to the Jebel Sahaba series (Supplementary Table S2).
Conclusion
African population dynamics were determined by shifting ecological boundaries between the Mediterranean and the tropical zone71. During green windows of reduced aridity migration corridors through Northern Africa, the Levant and the Arabian Peninsula caused pulses of hominin/human dispersals that ultimately were followed by population contractions when humid phases came to an end71,72,73. The Sahara formed a major part of this ecotone, periodically entailing bidirectional encounters not only with the Near-East, but also with sub-Saharan Africa. In this context, Northern Africa occupies a unique position for understanding supra-regional activities of hominin groups in the Middle and Late Pleistocene. Our data substantiate sporadic human exchanges between Northern Africa, the Near-East57,59, Europe57,63,64 and sub-Saharan Africa59 that would have been limited to periods of favorable climatic conditions70,71,72,73. Nevertheless, fossil mandibles dating to the Maghrebian MSA (Kébibat, Irhoud 11, Aterian) exhibit notable similarities in shape and discrete traits to preceding and succeeding populations in the area (Table 1, Supplementary Table S2; Figs. 6 and 7), implying a long-term succession of hominins in Northern Africa. In this debate, Kébibat and the Aterians fill not only a chronological but also a morphological gap of considerable interest between the earliest currently known H. sapiens from Jebel Irhoud and later H. sapiens. At the same time, their mosaic mandibular morphology sheds some light onto Late Pleistocene diversity and is in line with an accretion of modern traits through time, consistent with an ongoing masticatory gracilization11.
References
Schlebusch, C. M. et al. Genomic variation in seven Khoe-San groups reveals adaptation and complex african history. Science 338, 374–379 (2012).
Mallick, S. et al. The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature 538, 201–206 (2016).
Hublin, J.-J. et al. New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature 546, 289–292 (2017).
Grün, R. et al. Dating the skull from Broken Hill, Zambia, and its position in human evolution. Nature 580, 372–375 (2020).
Richter, D. et al. The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature 546, 293–296 (2017).
Woodward, A. S. A new cave man from Rhodesia, South Africa. Nature 108, 371–372 (1921).
Hublin, J.-J. Northwestern African Middle Pleistocene hominids and their bearing on the emergence of Homo sapiens. In Human Roots. Africa and Asia in the Middle Pleistocene (eds. Barham, L. & Robson-Brown, K.) 99–121 (Bristol Western Academic & Specialist Press Ltd, 2001).
Oujaa, A., Arnaud, J., Bardey-Vaillant, M. & Grimaud-Hervé, D. The fossil human from Rabat-Kébibat (Morocco): Comparative study of the cranial and mandibular fragments. Afr. Archaeol. Rev. 34, 511–523 (2017).
Scerri, E. M. L. et al. Did our species evolve in subdivided populations across Africa, and why does it matter?. Trends Ecol. Evol. 33, 582–594 (2018).
Klein, R. G. Population structure and the evolution of Homo sapiens in Africa. Evol. Anthropol. Issues News Rev. 28, 179–188 (2019).
Bergmann, I., Hublin, J.-J., Gunz, P. & Freidline, S. E. How did modern morphology evolve in the human mandible? The relationship between static adult allometry and mandibular variability in Homo sapiens. J. Hum. Evol. 157, 103026 (2021).
Fu, Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr. Biol. 23, 553–559 (2013).
Posth, C. et al. Pleistocene mitochondrial genomes suggest a single major dispersal of non-Africans and a Late Glacial population turnover in Europe. Curr. Biol. 26, 557–561 (2016).
Dibble, H. L. et al. On the industrial attributions of the Aterian and Mousterian of the Maghreb. J. Hum. Evol. 64, 194–210 (2013).
Richter, D., Moser, J., Nami, M., Eiwanger, J. & Mikdad, A. New chronometric data from Ifri n’Ammar (Morocco) and the chronostratigraphy of the Middle Palaeolithic in the Western Maghreb. J. Hum. Evol. 59, 672–679 (2010).
Bouzouggar, A. & Barton, R. N. E. The identity and timing of the aterian in Morocco. In Modern Origins: A North African Perspective (eds Hublin, J.-J. & McPherron, S. P.) 93–105 (Springer Netherlands, 2012). https://doi.org/10.1007/978-94-007-2929-2_7.
Sehasseh, E. M. et al. Early Middle Stone Age personal ornaments from Bizmoune Cave, Essaouira, Morocco. Sci. Adv. 7, eabi8620 (2021).
Nespoulet, R. et al. Le contexte archéologique des restes humains atériens de la région de Rabat-Témara (Maroc): Apport des fouilles des grottes d’El Mnasra et d’El Harhoura 2. Actes Quatr. Rencontres Quat. Maroc. RQM4 Quat. Maroc. Dans Son Contexte Méditerranéen 356–375 (2008).
El Hajraoui, M. A. L’industrie osseuse atérienne de la grotte d’El Mnasra (Région de Témara, Maroc). Préhistoire Anthropol. Méditerranéennes 3, 91–94 (1994).
Bouzouggar, A. et al. 90,000 year-old specialised bone technology in the Aterian Middle Stone Age of North Africa. PLoS ONE 13, e0202021 (2018).
Hallett, E. Y. et al. A worked bone assemblage from 120,000–90,000 year old deposits at Contrebandiers Cave, Atlantic Coast, Morocco. iScience 24, 102988 (2021).
Rightmire, G. P. & Deacon, H. J. Comparative studies of Late Pleistocene human remains from Klasies River Mouth, South Africa. J. Hum. Evol. 20, 131–156 (1991).
Hublin, J.-J. et al. Dental evidence from the Aterian human populations of Morocco. In Modern Origins 189–204 (Springer, 2012).
Ferembach, D. Les restes humains de la Grotte de Dar-es-Soltane II (Maroc). Campagne 1975. Bull. Mém. Société Anthropol. Paris 3, 183–193 (1976).
Debénath, A. Nouveaux restes humains atériens du Maroc. CR Acad. Sci. Paris 290, 851–852 (1980).
Ferembach, D. Le crâne atérien de Témara (Maroc atlantique). Bull. Archéologie Marocaine 18, 19–66 (1998).
Saban, R. Les restes humains de Rabat (Kébibat). Annls PaléontolVert 61, 151–207 (1975).
Jaeger, J. J. Les hommes fossiles du Pléistocène moyen du Maghreb dans leur cadre géologique, chronologique et paléoécologique. In Homo erectus. Papers in Honor of Davidson Black (eds Sigmon, B. A. & Cybulski, J. S.) 159–187 (The University of Toronto Press, 1981).
Camps, G. Les civilisations préhistoriques de l’Afrique du Nord et du Sahara Vol. 348 (Doin Paris, 1974).
Barton, R. N. E. et al. Origins of the Iberomaurusian in NW Africa: New AMS radiocarbon dating of the Middle and Later Stone Age deposits at Taforalt Cave, Morocco. J. Hum. Evol. 65, 266–281 (2013).
Marçais, J. Découverte de restes humains fossiles dans les grès quaternaires de Rabat, Maroc (Maison et Cie, 1934).
Chahid, D. et al. New lithostratigraphic and chronostratigraphic data for the fossil human skull-bearing eolianite of Rabat-Kebibat (Morocco). Quat. Rev. Assoc. Fr. Pour Létude Quat. https://doi.org/10.4000/quaternaire.14287 (2020).
Stearns, C. E. & Thurber, D. L. Th230/U234 dates of late Pleistocene marine fossils from the Mediterranean and Moroccan littorals. Prog. Oceanogr. 4, 293–305 (1965).
Jacobs, Z. et al. Single-grain OSL dating at La Grotte des Contrebandiers (‘Smugglers’ Cave’), Morocco: Improved age constraints for the Middle Paleolithic levels. J. Archaeol. Sci. 38, 3631–3643 (2011).
Dibble, H. L. et al. New excavations at the site of Contrebandiers Cave, Morocco. PaleoAnthropology 2012, 145–201 (2012).
Roche, J. Chronostratigraphie des restes atériens de la grotte des Contrebandiers à Témara (Province de Rabat). Bull. Mém. Société Anthropol. Paris 3, 165–173 (1976).
Vallois, H. V. & Roche, J. L. La mandibule acheuléenne de Témara. Comptes Rendus Hebd. Séances Académie Sci. 246, 3113–3116 (1958).
Debénath, A. Le site de Dar-es-Soltane II à Rabat (Maroc). Géologie et préhistoire. Bull. Mém. Société Anthropol. Paris 3, 181–182 (1976).
Schwenninger, J.-L. et al. A new luminescence chronology for Aterian cave sites on the Atlantic coast of Morocco. South-East. Mediterr. Peoples Between 130, 18–36 (2010).
Raynal, J.-P. & Occhietti, S. Amino chronology and an earlier age for the Moroccan Aterian. In Modern Origins: A North African Perspective (eds Hublin, J.-J. & McPherron, S. P.) 79–90 (Springer Netherlands, 2012). https://doi.org/10.1007/978-94-007-2929-2_6.
Harvati, K. & Hublin, J.-J. Morphological continuity of the face in the late Middle and Late Pleistocene hominins from northwestern Africa: A 3D geometric morphometric analysis. In Modern Origins 179–188 (Springer, 2012).
Debénath, A. & Sbihi-Alaoui, F. Découverte de deux nouveaux gisements préhistoriques près de Rabat (Maroc). Bull. Société Préhistorique Fr. Comptes Rendus Séances Mens Paris 76, 11–14 (1979).
Debénath, A., Raynal, J.-P. & Roche, J. Stratigraphie, Habitat, Typologie et devenir de l’Atérien. Anthropol. Paris 90, 233–246 (1986).
Debénath, A. L. peuplement préhistorique du Maroc: données récentes et problèmes. L’Anthropologie 104, 131–145 (2000).
Raynal, J.-P. et al. Hominid Cave at Thomas Quarry I (Casablanca, Morocco): Recent findings and their context. Quat. Int. 223–224, 369–382 (2010).
Ennouchi, É. Découverte d’un Pithécanthropien au Maroc. (1969).
Sausse, F. L. mandibule atlanthropienne de la carrière Thomas I (Casablanca). Anthropologie. 79, 81–112 (1975).
Geraads, D. Plio-Pleistocene Mammalian biostratigraphy of Atlantic Morocco. Quaternaire 13, 43–53 (2002).
Raynal, J. P., SbihiAlaoui, F. Z., Geraads, D., Magoga, L. & Mohi, A. The earliest occupation of North-Africa: The Moroccan perspective. Quat. Int. 75, 65–75 (2001).
Rhodes, E. J., Singarayer, J. S., Raynal, J.-P., Westaway, K. E. & Sbihi-Alaoui, F. Z. New age estimates for the Palaeolithic assemblages and Pleistocene succession of Casablanca, Morocco. Quat. Sci. Rev. 25, 2569–2585 (2006).
Arambourg, C. & Hoffstetter, R. Le gisement de Ternifine Vol. 1 (Masson, 1963).
Geraads, D. et al. The Pleistocene hominid site of Ternifine, Algeria: New results on the environment, age, and human industries. Quat. Res. 25, 380–386 (1986).
Sahnouni, M. & van der Made, J. The Oldowan in North Africa within a biochronological framework. In The Cutting Edge: New Approaches to the Archaeology of Human Origins 179–210 (Stone Age Institute Press, 2009).
Geraads, D. Pleistocene Carnivora (Mammalia) from Tighennif (Ternifine), Algeria. Geobios 49, 445–458 (2016).
Geraads, D. Tighennif (Ternifine), Algeria: environments of the earliest human fossils of North Africa in the late Early/early Middle Pleistocene. In African Paleoecology and Human Evolution (Cambridge University Press, In press).
Martini, P. & Geraads, D. Camelus thomasi Pomel, 1893 from the Pleistocene type-locality Tighennif (Algeria). Comparisons with modern Camelus. Geodiversitas 40, 115–134 (2018).
Kéfi, R. et al. On the origin of Iberomaurusians: New data based on ancient mitochondrial DNA and phylogenetic analysis of Afalou and Taforalt populations. Mitochondrial DNA Part A 29, 147–157 (2018).
Fregel, R. et al. Ancient genomes from North Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe. Proc. Natl. Acad. Sci. 115, 6774–6779 (2018).
van de Loosdrecht, M. et al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations. Science 360, 548–552 (2018).
Arambourg, C. & Biberson, P. The fossil human remains from the Paleolithic site of Sidi Abderrahman (Morocco). Am. J. Phys. Anthropol. 14, 467–489 (1956).
White, T. D. et al. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423, 742–747 (2003).
Kupczik, K. & Hublin, J.-J. Mandibular molar root morphology in Neanderthals and Late Pleistocene and recent Homo sapiens. J. Hum. Evol. 59, 525–541 (2010).
Hernández, C. L. et al. Early holocenic and historic mtDNA African signatures in the Iberian Peninsula: The Andalusian Region as a Paradigm. PLoS ONE 10, e0139784 (2015).
Hernández, C. L. et al. The distribution of mitochondrial DNA haplogroup H in southern Iberia indicates ancient human genetic exchanges along the western edge of the Mediterranean. BMC Genet. 18, 46 (2017).
Zampetti, D. La question des rapports entre la Sicile et l’Afrique du Nord pendant le Paléolithique Supérieur Final: la contribution de l’archéologie. BAR. Int. Series 508, 459–476 (1989).
Straus, L. G. Africa and Iberia in the Pleistocene. Quat. Int. 75, 91–102 (2001).
Bianchi, F., BorgogniniTarli, S. M., Marchi, M. & Paoli, G. An attempt of application of multivariate statistics to the problems of the Italian mesolithic samples. HOMO J. Comp. Hum. Biol. 31, 153–166 (1980).
Secher, B. et al. The history of the North African mitochondrial DNA haplogroup U6 gene flow into the African, Eurasian and American continents. BMC Evol. Biol. 14, 109 (2014).
Stöck, M. et al. Post-Messinian evolutionary relationships across the Sicilian channel: Mitochondrial and nuclear markers link a new green toad from Sicily to African relatives. BMC Evol. Biol. 8, 56 (2008).
Drake, N. A., Blench, R. M., Armitage, S. J., Bristow, C. S. & White, K. H. Ancient watercourses and biogeography of the Sahara explain the peopling of the desert. Proc. Natl. Acad. Sci. 108, 458–462 (2011).
Larrasoaña, J. C., Roberts, A. P. & Rohling, E. J. Dynamics of green Sahara periods and their role in hominin evolution. PLoS ONE 8, e76514 (2013).
Rosenberg, T. M. et al. Middle and Late Pleistocene humid periods recorded in palaeolake deposits of the Nafud desert, Saudi Arabia. Quat. Sci. Rev. 70, 109–123 (2013).
Breeze, P. S. et al. Palaeohydrological corridors for hominin dispersals in the Middle East ∼250–70,000 years ago. Quat. Sci. Rev. 144, 155–185 (2016).
Crevecoeur, I. Étude anthropologique du squelette du Paléolithique supérieur de Nazlet Khater 2 (Égypte): Apport à la compréhension de la variabilité passée des hommes modernes Vol. 8 (Leuven University Press, 2008).
Harvati, K. et al. The later stone age calvaria from Iwo Eleru, Nigeria: Morphology and chronology. PLoS ONE 6, e24024 (2011).
Acknowledgements
We thank the following scientists, institutions, curators and museums for providing access to mandibular remains of fossil and recent humans: Patrick Semal, Institut royal des Sciences naturelles de Belgique, Brussels (La Naulette, Spy, Ishango, Shum Laka) Stéphane Louryan, Laboratoire d’anatomie, biomécanique et organogenèse, Université Libre de Bruxelles (Ishango); Marie-Antoinette de Lumley, Centre Européen de Préhistoire Tautavel (Arago); Aurelie Fort, Antoine Balzeau, Muséum national d'histoire naturelle, Paris (Abri Pataud 1, Cro Magnon 1+3, La Ferrassie 1, Montmaurin 1); Henry de Lumley, Amelie Vialet, Institut de Paléontologie humaine, Paris (Afalou Bou Rhummel, Asselar, Grotte de Gramat, Isturitz, Rabat 1, Taforalt); Patrick Simon (Directeur), Jerome Magail, Musée d'Anthropologie préhistorique de Monaco (Grotte des Enfants 4); Andrea Pascale, Museo Archeologico del Finale (Arene Candide 2); Elisabetta Starnini, Museo Nazionale Preistorico ed area archeologica dei Balzi Rossi (Barma Grande 2, 4, 5); Roberto Macchiarelli, Didier Berthet, Arnaud Mazurier, Musée des Confluences, Lyon (La Quina H9); Roberto Macchiarelli, Christine Argot, Laboratoire de Paleontologie, MNHN Paris (Tighenif 1+3); Clément Zanolli and Arnaud Mazurier, who scanned the Tighenif mandibles at the University of Poitiers; José Miguel Carretero, Lawrence Guy Straus, University of Burgos/Laboratory of Human Evolution, Burgos (El Mirón); Montserrat Sanz, Joan Daura, Antonio Rosas, Museo Nacional de Ciencias Naturales, Madrid (El Sidrón 1 + 2); Juan Luis Arsuaga & Ana Pantoja Pérez, Centro mixto UCM-ISCIII de Evolución y Comportamiento Humanos, Madrid (Sima de los Huesos/Atapuerca); Michael Walker, Universidad de Murcia (Sima de las Palomas 6+23); Martin Oliva, Zdeněk Tvrdý, Anthropos Institute, Moravian Museum, Zclny Brno (Dolní Věstonice 3); Sergey V. Vasilyev & Natalia Kharlamova, Paleoanthropological collections of the Institute of Ethnology and Anthropology, Russian Academy of Sciences (Sunghir 1); Mauro Rubini, Anthropological Service of S.A.B.A.P.-RM-MET, Rome (Guattari 2+3); Isabelle Crevecoeur, Bordeaux 1 University (Ishango), George McGlynn, Staatssammlung für Anthropologie und Paläoanatomie, München (Olduvai 1); Robert Kruszynski, Natural History Museum, London (Tabun C1); Daniel E. Lieberman, Michèle Morgan, Katherine Meyers (Associate Archivist), Peabody Museum of Archeology and Ethnology, Harvard (Skhul V); David Katz, University of California Davis; Jay T. Stock & Tom Davies & Marielle Brown, University of Western Ontario (Jebel Sahaba); Lauren Conroy, University of Chicago (Gobero); Isabelle Ribot & Yassmine Ghalem (access to and 3D scanning of Shum Laka); Israel Hershkovitz, Dani Nadel, Julia Abramov, Dan David Center for Human Evolution and Biohistory, Tel Aviv University (Ain Mallaha, Hayonim Cave 25+ 27+ 29+ 29a, Nahal Oren 6+18+20,); Israel Hershkovitz, Baruch Arensburg, Sackler School of Medicine, Tel Aviv University (Amud 1, Hayonim Cave 8+ 17+ 19+ 20, Kebara 2, Nahal Oren 8+14, Qafzeh 9, Tabun 2 C2); Rockefeller Museum Jerusalem (Skhul IV); Abdelouahed Ben-Ncer & Mohammed Abdeljalil El Hajraoui & Samir Raoui, Institut National des Sciences du Patrimoine et de l’Archeólogie and “Direction du Patrimoine Culturel”, Rabat (Djebel Irhoud 11; El Harhoura, Dar-es-Soltane II, Grotte des Contrebandiers); Yves Coppens, Collège de France Paris & Kornelius Kupczik, Max-Planck-Institut für evolutionäre Anthropologie & Alexandra Franz, Universität Leipzig (Thomas Quarry); Emma Mbua, National Museums of Kenya Nairobi (Baringo-Kapthurin), Wendy Black, Miss Erica Bartnick, Iziko Museums of South Africa, Kapstadt (Klasies River Mouth); Bernhard Zipfel, Medical School of the University of the Witwatersrand, Johannesburg (Border Cave, Mumbwa); Emma Mbua, Department of Earth Science, National Museums of Kenya (Lothagam); Priscilla Bayle, PACEA/Université Bordeaux 1 (Sima de las Palomas 6) Ekaterina Stansfield/University of York, A. Soficaru/Francisc I. Rainer Institute of Anthropology (Schela Cladovei, Vărăști Gulmenița); Erik Trinkaus, Washington University St. Louis (Oase 1), Ana Luísa Santos, CIAS Research Center for Anthropology and Health/Department of Life Sciences, Universidade de Coimbra (recent Coimbra sample); Ulrich A. Glasmacher, Institut für Geowissenschaften, Universität Heidelberg (Mauer); Christine Feja & Katharina Spanel-Borowski, Anatomische Sammlung Universität Leipzig and Adeline Le Cabec & Stefanie Stelzer & Nadia Scott & Simon Neubauer, Max Planck Institute for Evolutionary Anthropology Leipzig (recent ULAC series). This research was made possible by INSAP (Institut National de l’Archéologie et du Patrimoine), Rabat, Morocco, and funded by the Max Planck Society. We thank David Plotzki, Heiko Temming, Andreas Wintzer (technicians) for assistance with CT scanning and reconstructing computed tomography data. We are grateful to Shannon McPherron and Will Archer for assistance in photogrammetry. We want to thank Zeljko Rezek for discussions and comments on the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
I.B. analyzed the data and wrote the main manuscript as well as supplemental files. J-J.H., P.G., S.E.F., A.B-N. and F.Z.S-A. reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bergmann, I., Hublin, JJ., Ben-Ncer, A. et al. The relevance of late MSA mandibles on the emergence of modern morphology in Northern Africa. Sci Rep 12, 8841 (2022). https://doi.org/10.1038/s41598-022-12607-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-12607-5
- Springer Nature Limited
This article is cited by
-
The Undescribed Juvenile Maxilla from Contrebandiers Cave, Morocco—A Study on Middle Stone Age Facial Growth
Journal of Paleolithic Archaeology (2024)
-
Symbolic innovation at the onset of the Upper Paleolithic in Eurasia shown by the personal ornaments from Tolbor-21 (Mongolia)
Scientific Reports (2023)
-
Identifying signatures of positive selection in human populations from North Africa
Scientific Reports (2023)