
Abstract
Forest restoration have had limited success due to intense and prolonged droughts in Mediterranean-type ecosystems. In this context, knowledge of growth and physiology in seedlings of different provenances can be useful in the selection of appropriate seed sources for restoration. In this study we investigated variations in survival, growth, and leaf-level physiology of five provenances of Quillaja saponaria Mol. and five provenances of Cryptocarya alba Mol. originated from coastal and Pre Andean sites exhibiting latitudinal-related climate differences in central Chile. Seedlings were grown in a nursery on 600 mL pots for 18 months and then planted in a dryland site severely damaged by fire. One year after establishment, we measured survival, growth, and leaf-level physiology. We also analyzed the relationship between outplanting survival with seedling characteristics prior to planting, and the relationship between growth and survival with physiological traits and with climate variables. Growth and survival were similar among provenances of Q. saponaria and C. alba, with the exception of differing heights observed within the provenance of Q. saponaria. Initial root collar diameter of Q. saponaria was observed to be positively correlated to outplanting survival. With the exception of photosynthesis in Q. saponaria, all provenances of both species differed in the leaf-level physiological traits. Those provenances originating from interior dryland sites exhibited lower stomatal conductance and used water more efficiently. The opposite was true for provenances coming from coastal sites. In outplanting sites with Mediterranean-type climates that have been damage by severe fire, selections based on larger diameter seedlings, especially for Q. saponaria and from interior and pre-Andean provenances, will likely improve outplanting success.
Similar content being viewed by others
Introduction
Mediterranean-type forest ecosystems are very fragile and susceptible to degradation1. In South America, the Chilean Mediterranean-type forest ecosystem has been under permanent threat since the European colonization and 83% of its original coverage is already lost mainly because of threats such as land conversion for agricultural purposes2. The restoration of this degraded ecosystem have had limited success and have created concern about the factors influencing the success for its restoration, which may greatly depend on factors such as the appropriate seed sources3 and seedling morphological attributes determining the quality of the planting stock, i.e., desirable phenotypic traits; such as shoot height, stem diameter and root system, that promotes successful seedling establishment4,5.
Provenance origin and seed source plays an important role in the successful restoration of degraded areas6. Local seed sources are better adapted to local conditions, which improves survival and growth7 as has been found in Quercus ilex L. and Quillaja saponaria Mol.8,9. Adaptations of sclerophyllus species growing across a range of Mediterranean-type climates has shown that provenances from mesic environments have elongated and large leaves10, while provenances from dry climates have a reduced leaf area as an adaptation to face water restriction11. Stomatal regulation in response to drought stress is an important mechanism of Mediterranean sclerophyllus species such as Q. ilex to prevent water loss12. The species exhibits early stomatal closure before to postpone the onset of water stress13.
Seedling quality is another important factor that constrains the success of the restoration because it influences plant survival and growth14,15,16,17. Seedling quality is assessed by several morphological and physiological attributes, but height, diameter, and root:shoot ratio are typically considered as good predictors of the outplanting survival5. In general, large seedlings increases their survival and growth in degraded drylands because they develop larger roots that can reach moisture in deep soil horizons16,18,19,20,21,22,23.
Quillaja saponaria Mol. and Cryptocarya alba Mol. are two endemic sclerophyllus tree species widely distributed in the Mediterranean drylands of central Chile. Populations of both species were severely degraded by the fires occurred in central Chile during summer 2017, being the worst catastrophic wildfires of the last 50 years that burned nearly 520,000 ha of land24. This situation aggravated the conditions for forest recovery, and exacerbated the early mortality of planted seedlings9,25. The distribution range of both species spans from the coastal areas in the arid northern part of Chile (30° S) to the humid climates in the southern part of the country (40° S)26. Both species have specific adaptations to survive in Mediterranean-type climates. Q. saponaria is a shade-intolerant and deep-rooted species that exhibits water potentials of c.a. − 2.0 MPa during summer, while C. alba is a shade-tolerant and shallow-rooted species that maintain water potentials of c.a. − 4.0 MPa in the summer period9,27,28. Q. saponaria has a higher photosynthetic rate and stomatal conductance than C. alba29,30. Although both species are routinely planted in restoration projects, its outplanting performance is highly unpredictable and variable25. There is still a gap in the information on the outplanting performance after a severe fire in relation to seed-source provenance for both species, causing concerns of the appropriate seed sources for restoration under climatic change context. Severe fires are known to cause losses of organic matter and nutrients, increases soil erosion31 and spatial variation of the soil properties, decreases in site productivity32, and plant growth, and survival33, and decreases in post-fire recruitment34. As seedlings experience higher rates of mortality in the first year after planting, monitoring the early performance can give guidelines for achieving restoration goals. Thus, the aim of this study was to assess variation in growth, survival, and leaf-level physiology across a latitudinal gradient of seed sources for Q. saponaria and C. alba that were established on a dryland site severely damaged by fire. We hypothesized that provenance origin will have a great effect on variation in outplanting success of Q. saponaria and C. alba in harsh sites.
Methods
Study site
The experiment was established at Las Brisas Experimental Station (35° 34′ S, 72° 06′ W, 254 m a.s.l) which belongs to the Universidad de Chile and it is located 50 km west of the San Javier city, Maule Region, central Chile. In 2017, the total area of the experimental station (93 ha) was completely burned by fire (de la Barrera et al. 2018). We estimated the burn severity at the study site, based on the relativized delta normalized burn ratio (RdNBR) from pre- and post- fire Sentinel-2 (20 m) images. The RdNBR is an index developed from satellite imagery to estimate vegetation burn severity maps using pre and post-fire images. The index values usually can be grouped into three categories as low, medium, and high severity, depending on alterations in the soil, canopy cover, and vegetation mortality35,36. The results indicated that a high percentage of the study area experienced a high burn severity. Prior to the fire, the area supported different vegetation types such as: (1) isolated trees of native forests, dominated by species such as Q. saponaria, Acacia caven (Mol). Mol., Lithraea caustica (Mol.) Hook. & Arn., Escallonia pulverulenta (Ruiz et Pav.) Pers., and Peumus boldus Mol., (2) plantations of Pinus radiata D. Don, and (3) introduced pasture grasses such as Aira cariophyla, Briza minor, and Bromus hordeaceus among others. The climate at the study site is considered Mediterranean, with annual rainfall of 734 mm occurring mostly in the winter months (675 mm from June to August). The dry period is around 7 months, and summer is typically hot and dry (maximum daily temperature of 36 °C during January). The soil is neutral (pH 6.1), sandy clay (47% sand, 17% lime, 36% clay), low electrical conductivity (0.03 dS m−1), and 1.5% organic matter content. The available soil nutrient content is 4 mg kg−1 N, 8 mg kg−1 P, and 168 mg kg−1 K. Based on SPAW Hydrology (a free online water budgeting tool from the USDA Natural Resource Conservation Service) we obtained volumetric water content and bulk density at a depth of 30 cm. Values were 8.42 cm at field capacity, 5.65 cm at wilting point, and 1.49 g cm−3 bulk density.
Plant material
Five provenances of Q. saponaria and C. alba exhibiting latitudinal-related climate differences (Table 1, Fig. 1) were grown in a nursery, property of the Forestal Arauco Company (35° 18′ S, 72° 23′ W, 10 m a.s.l), located in the city of Constitución, central Chile. A mix of seeds from different mother trees represented each provenance. Seedlings were grown in 600 mL pots filled with composted bark of P. radiata which was combined with the slow release fertilizer BASACOTE 9 M, and cultured under ambient conditions of temperature and light. In both species, seeds were sown on March 2017 and maintained for 18 months under daily irrigation.

Location of the provenances origin and the experimental site. The approximate range of distribution of both species in central Chile is indicated by a square in the inset graph.
Experimental design
The field experiment was a randomized complete block design with seven blocks and included five provenances per species and a row-plot of 7-seedlings as the experimental unit (5 provenances × 7 blocks × 7 seedlings per block = 245 seedlings per species). No sun protection was provided for seedlings at the field test site. The site was fenced to avoid herbivore damage. Seedlings were hand planted during August 2018, in planting holes (25 × 25 × 30 cm) at a spacing of 1 × 1 m. All seedlings were watered once a month (4 L plant−1 month−1) during four months (December 2018 to March 2019) and no fertilizer was added to the planting holes.
Survival and growth traits
Prior to planting the field experiments, all seedling were measured for height (Hi) and root collar diameter (Di) using a meter stick and a digital caliper, respectively. The slenderness index was estimated as the height to diameter ratio (Hi/Di). Then, one growing season after field establishment, seedlings were measured for H (Hp), D (Dp), and survival (SUR). SUR was measured as a categorical trait (i.e., alive seedling = 1, dead seedling = 0), and then expressed as percentage at plot level. Increments for D (INCd) and H (INCh) were calculated as the differences in those traits prior to planting with those one year after establishment.
Gas-exchange measurements at the field experiment
During January 2019 light-saturated photosynthetic rate (Asat, µmol CO2 m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1), and the derived intrinsic water use efficiency (WUEi = Asat/gs) were measured in fully-developed leaves located on the upper third of the seedlings. These measurements were taken between 09:00 and 12:00 (local time) using an LI-6800 photosynthesis system (LI-COR Inc., Lincoln, NE, USA). The conditions of temperature, air CO2 concentration, and light source inside the chamber were set to 25 °C, 400 ppm, and 1800 mmol m2 s1, respectively.
Data analyses
We conducted a first analysis to assess variations in survival, growth, and leaf-level physiology of species and provenances under study. Growth traits at the time of planting were analyzed with a lineal model considering provenance as a fixed factor, whereas survival, growth, and leaf-level physiology traits 1 year after outplanting were analyzed using a lineal mixed model including blocks as a random factor. Blocks were included to diminish the influence of environmental variation due to the heterogeneity of site conditions after the fire and thereby improve statistical power. The analyses of variance for each trait was carried out on a plot-mean basis (i.e., average of row-plot of 7-seedlings) for each species separately. To meet the assumptions of normality and constant variances, growth, and leaf-physiological traits were transformed according to the Box-Cox transformation when necessary37. In the case of survival, categorical values (i.e., 1, 0), were expressed in percentages at the plot level and transformed by the arcsine square root transformation. To correct those values that included 0 (zero), we added 1 (one) to each survival value. Mean comparisons were made using the Tukey test at P ≤ 0.05.
Because our interest was also to analyze the effect of seedling morphological attributes on outplanting performance, we used a simple regression model separately by species to assess (1) the relationship between seedling height and diameter at the time of planting with the survival one year after outplanting, and (2) the relationship between diameter, height, and survival with leaf-level physiological traits 1 year after outplanting. In both cases, we checked assumptions of normality and homogeneity of variances and the goodness of fit for the regression models was assessed by computing the coefficient of determination and the Pearson’s coefficient of correlation. In addition, we conducted a Path analysis with the aim to assess how the independent variables (Di, Hi, Dp, Hp, Asat and gs), influenced the seedling outplanting survival. In this analysis we omitted the traits derived as ratios i.e., Hi/Di and WUEi (Asat/gs), to avoid multicollinearity.
Finally, a Principal Component Analysis (PCA) was carried out on all database with the aim to explore the relationship between seedling performance at the time of planting and after outplanting (Di, Hi, Dp, Hp, Asat, gs, and WUEi) with the climate at provenance locations where seed was collected using mean annual temperature and precipitation, and the De Martonne aridity index. Mean annual temperature and precipitation were taken from the Worldclim (www.worldclim.org) high-resolution dataset. All the statistical analyses were performed with SPSS version 18.0 software (SPSS Inc, Chicago, Illinois, USA) and INFOSTAT version 2018 (Group Infostat, Universidad Nacional de Córdoba, Argentina).
Results
Differences in height and diameter at time of planting
Both species exhibited differences in growth traits at the provenance level. In the case of C. alba, Di was higher in the provenances of Los Queñes (Pre Andean site), while Hi was higher in the provenance of El Carmen (interior site). The Linares provenance (Pre Andean site) had the lowest Di and exhibited the highest Hi/Di ratio (Fig. 2C). As Di was more variable among provenances (Fig. 2A) than Hi (Fig. 2B), the differences in Hi/Di are better explained by the differences in Di than in Hi. In the case of Q. saponaria, seedlings of the Cholguán and Pocillas provenances (both from interior sites) were the tallest and the shortest, respectively (Fig. 2E), while seedlings from Vichuquén and Curepto; from coastal sites, exhibited the thickest Di (Fig. 2D). As with Hi, the provenance Cholguán (from interior sites) had the highest Hi/Di ratio (Fig. 2F). The differences in Hi/Di for Q. saponaria were more associated to differences in Hi.

Seedling characteristics for C. alba and Q. saponaria at time of planting. Di root collar diameter, Hi height, Hi/Di height to diameter ratio. Different letters indicate significant differences among provenances (Tukey test; p ≤ 0.05). The error bars indicate the standard error. Codes for provenances are according to Table 1. Provenances are ordered from left to right by latitude of origin.
Growth and survival in the field plantation and their relationship with seedling attributes at time of planting
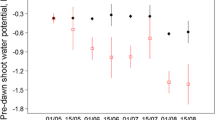
Provenances of Q. saponaria did not differ in survival and this trait ranged from 41 to 52% for provenances Cabrero and Curepto, respectively (Fig. 3E). Our results showed a significant provenance effects only for Hp and INCh (Table 2). Provenances Pocillas and Curepto exhibited the tallest and shortest seedlings, respectively (Fig. 3C), but INCh was higher and positive only in the Pocillas provenance. Most planting stocks of the other provenances exhibited negative height growth (Fig. 3D). Similar to survival, no provenance differentiation was found for INCd (Fig. 3B) and Dp ranged from 3.7 to 4.5 mm for provenances Curepto and Pocillas, respectively (Fig. 3A). In the case of C. alba, no differences in Dp, Hp, INCd, INCh and SUR were observed at the provenance level (Fig. 3F–I). SUR was very low in this species and ranged from 0 to 6% (Fig. 3J), while Hp and Dp of the surviving seedlings ranged from 8.1 to 10.5 cm, and from 5.2 to 5.9 mm, respectively. Most seedlings in all provenances exhibited negative INCh. The relationship between seedlings characteristics measured at the time of planting and SUR in the field showed low and poor correlations. Di and Hi/Di were positively and negatively correlated with SUR in Q. saponaria, but Hi exhibited no relationship with this trait (Fig. 4A–C). No significant correlations were found in C. alba (Fig. 4D–F).

Survival (SUR), root collar diameter (Dp), height (Hp) and their respective increments in D (INCd) and H (INCh) for Q. saponaria and C. alba seedlings according to the provenance origin. Different letters indicate significant differences among provenances (Tukey test; p ≤ 0.05). The error bars indicate the standard error. Codes for provenances are according to Table 1. Provenances are ordered from left to right by latitude of origin.

Relationship among the seedling attributes measured at the time of planting and survival measured 1 year after outplanting. Root collar diameter (Di), height (Hi), height to diameter ratio (Hi/Di), survival (SUR).
Gas-exchange responses and its relationship with growth and survival
Provenances of Q. saponaria differed in gs and WUEi but not in Asat (Fig. 5D, Table 3). The provenances of Vichuquén and Pocillas exhibited the highest gs but the lowest WUEi. On the contrary, the provenance of Cabrero had the highest WUEi while the provenance of Curepto exhibited low rates of gs (Fig. 5E,F). In this species, SUR was positively correlated with Asat (r = 0.54, p = 0.017) and with gs (r = 0.58, p = 0.009). The relationship between other seedlings characteristics and physiological traits was non-significant. In the case of C. alba, the provenance Linares still had alive seedlings when gas exchange analysis was done (i.e., January 2019), but they died after the summer months. This is why this provenance appears with 0% survival, but with gas exchange measurements. Additionally, because no surviving seedlings were observed in the trial for the provenance of Hualañé, it was not possible to obtain physiological measurements. Therefore, the provenances of Los Queñes (pre-Andean provenance) and Coelemu (coastal provenance) consistently exhibited the highest and lowest values for all gas exchange traits under study (Fig. 5A–C). The relationship between seedlings characteristics and physiological traits of C. alba showed non-significant correlations (data not shown).

Gas exchange traits of C. alba and Q. saponaria seedlings according to the provenance. Different letters indicate significant differences among provenances (Tukey test; p ≤ 0.05). The error bars indicate the standard error. Codes for provenances are according to Table 1. Provenances are ordered from left to right by latitude of origin. In the case of C. alba, the provenance Hualañe (HU) had no alive seedlings after the post transplanting shock and this is why this provenance is missing from (A–C).
Path analysis of relationships between outplanting survival and independent variables
We used Path analysis to quantify simultaneously the direct and indirect contributions of seedling attributes and physiological responses on seedling outplanting survival (Table 4). The variable with the greatest direct effect on seedling SUR was gs, followed by Dp, Asat, Di, Hp, and Hi; but only Dp had directly significant negative effects on seedling survival (p < 0.05). In addition, this trait, via negatively affecting gs, indirectly affected seedling survival. Hp, Asat, and gs had significant indirect effects on seedling survival through their interrelated effects. There was a significant indirect correlation between SUR with Hp and Asat (correlation of 0.44 and 0.51) which is mainly determined by gs. Similarly, the significant indirect correlation between SUR and gs was mainly determined by Asat.
Principal component analysis of relationship among seedling performance and climate at provenance locations
The first (PC 1) and second (PC 2) principal components explained a significant proportion of the variability (78.2%) in the species and provenances under study (Fig. 6). The PC1 captures seedling performance, whereas the PC2 captures climate variables. SUR appeared to be positively correlated to the leaf-level physiological traits Asat and gs, but negatively correlated to seedling diameter at the time of planting and after outplanting. Some provenances of Q. saponaria are associated to a higher SUR, Asat and gs, whereas the provenances of C. alba are associated to larger diameters. Climatic variables did not show any correlation with the other variables, especially MAT.

Principal Component Analysis among seedling performance and climate at provenance locations. Each squares and triangle represents a provenance of C. alba and Q. saponaria, respectively. In the case of C. alba, provenances HU and LI were absent due to they had no alive seedlings after the post transplanting shock. SUR survival after outplanting, Di diameter at planting, Hi height at planting, Dp diameter after outplanting, Hp height after outplanting, Asat photosynthesis after outplanting, gs stomatal conductance after outplanting, WUEi water use efficiency after outplanting, MAP mean annual precipitation, MAT mean annual temperature, Martonne The De Martonne aridity index.
Discussion
The growth and survival were low and highly variable for both species. In the case of Q. saponaria, prior to planting seedlings differed in Di, Hi, and Hi/Di by provenance; however, with the exception of Hp no clear evidence of provenance differentiation was found for Dp and SUR 1 year after outplanting. The larger seedlings with the highest height increments were found in the provenance Pocillas, whose seed was collected at 24 km from the field test site and seedlings were the shortest prior to planting. As local genotypes performs better than distant genotypes38 it may be possible that this provenance have adaptations to the climatic conditions of the planting site that favored growth and positive height increments. Studies on sclerophyllous Mediterranean species have shown that survival improved using larger seedlings19,21,23, but in our study site characterized by low soil water availability and high temperatures during the growing season, seedling Hi was not related to field survival; however, the principal component analysis suggests that seedling Hp is associated to field survival. The Path analysis showed significant and negative direct effects of Dp on survival which; indirectly affecting gs, influenced survival. The initial stem diameter is a reliable indicator of field performance5 and our results suggests that, in sites with severe summer droughts, seedlings with ticker diameters could be associated to plants with higher transpiratory demands and stomatal conductances, with detrimental effects on seedling survival. It has been observed that small seedlings are better prepared to survive in dry environments39 because the low transpiring surfaces they develop and the consequent low risk of desiccation4. SUR at field was not correlated with Hi; which was corroborated by the Path analysis, but it was negatively correlated with Hi/Di, which was near the value of 6. Although the optimum value for Hi/Di that signifies good quality seedlings is unknown in Q. saponaria, it is recommendable to avoid the establishment of seedlings with excessive low or high Hi/Di values (i.e., stocky or spindly seedlings, respectively), as they have a low chance to survive in dry and windy sites5.
Both species experienced a high mortality, but this was particularly true for all provenances of C. alba that additionally experienced negative height growth. Negative height growth of C. alba was associated to a considerable decrease in leaf-level gas-exchange traits of seedlings submitted to a water restriction40. In addition, negative increments in height in Quercus pagoda Raf. and Quercus phellos L. were associated to the container type in which seedlings were cultured. Seedlings cultured in large containers were the largest in the nursery and experienced positive height growth at field41. Thus, the poor performance of provenances of C. alba in our study site might be explained by the shock experienced by small seedlings15,42,43. Planting small seedlings, as those of C. alba in our experiment (i.e., Hi < 20 cm), implies a reduced volume exploration by roots and consequently an insufficient water uptake and survival22. C. alba seedlings were also stocky prior to planting (Hi/Di in the 3–4 range), which might have contributed to low field survival5. However, in our study, there was not enough evidence supporting the effect of the seedling size prior to planting on the outplanting growth and survival of the different provenances. In this respect, in the sclerophyllous species Q. ilex and Quercus coccifera L., no significant correlations were found between field survival and seedling traits at nursery14,44,45. It seems that in harsh environments; as our study site that was additionally severely damaged by fire, it is not clear which seedling attributes determines establishment success46. The estimated burn severity index (i.e., RdNBR) indicated a high severity damage by fire in the study area. This condition probably changed the spatial patterns of the soil properties, exacerbating microsite variation and negatively affecting outplanting performance of both species, but this needs to be further investigated.
Unlike growth and survival, our study showed that Q. saponaria and C. alba exhibited differences among provenances in gas exchange parameters. The Principal component analysis showed no association between leaf-level physiological traits with climate variables but it reveals that most of the provenances Q. saponaria are associated to a higher SUR, Asat and gs. The analysis of variance indicated that, with the exception of Asat, provenances of this species differed in gs, and WUEi. We found low values for gs and higher WUEi in the provenance Cabrero, a provenance originating from sites with mean annual precipitation of 1206 mm year−1, sandy soils with low available water capacity, and an extended dry season from 5 to 7 months47. The superior WUEi in the Cabrero provenance may be attributed to a reduction in gs because Asat was not different among provenances. The likely natural adaptation of that provenance to the drier conditions from which it was originated might have allowed it a higher stomatal control and water conservancy. On the contrary, the Vichuquén provenance maintained superior levels of gs but had low WUEi. As this provenance is originated from sites close to the coast (20 km from the coast) with higher humidity and metamorphic soils, when established in the dry site of the study area, it became less water use efficient as an apparent adaptation to its local growing conditions. In a similar experiment it was found a higher WUEi and low gs in a provenance of Q. saponaria originated in sites with a Mediterranean-type climate and whose seed was collected at a similar latitude than the Cabrero provenance of our experiment48. Similarly, the authors found superior levels of gs and low WUEi in a provenance whose seed was collected in a site located at 10 km from the coast. In Q. saponaria, the relationship between SUR with Asat and gs was positive but needs to be interpreted with caution. Asat was not different among provenances and the provenances that exhibited higher gs did not exhibit a clear superiority in survival.. Our path analysis showed complex relationships among Asat and gs in indirectly affecting seedling survival. As both traits are closely related, our interpretation is that the control of stomatal closure in the summer months after the transplanting shock will promote a high survival in the harsh conditions of the planting site, corroborating the importance of stomatal regulation as a mechanism to prevent water loss and assure survival in Mediterranean sclerophyllus species12,13. In the case of C. alba, the contrasting differences in gas exchange between the provenances Coelemu and Los Queñes might be also related to its adaptation to specific site conditions. The provenance of Los Queñes comes from a pre-Andean site located at an altitude of 850 m.a.s.l. (Table 1); characterized by low mean annual temperature and negative temperatures in winter, and seedlings were able to sustain the highest Asat and WUEi of the provenances under study. This provenance might have evolved to optimize water use under cold condition as surviving under these conditions requires great stress tolerance, phenotypic plasticity or both. On the contrary, the provenance of Coelemu is a coastal provenance from altitudes close to the sea level and sites with warmer temperatures, and seedlings of this provenance were among the lowest performers in leaf-level physiology. The behavior of this provenance suggests that it follows the specialization theory49 in which genotypes adapted to favorable conditions; a coastal environment in this case, may have a decreased performance in restrictive environments, represented by the harsh conditions of our planting site. This could imply a low adaptability of coastal provenances when transferred to more dry sites, which needs to be further investigated.
References
Peñuelas, J. et al. Assessment of the impacts of climate change on Mediterranean terrestrial ecosystems based on data from field experiments and long-term monitored field gradients in Catalonia. Environ. Exp. Bot. 152, 49–59 (2017).
Salazar, A. et al. Land use and land cover change impacts on the regional climate of non-Amazonian South America: A review. Glob. Planet Change 128, 103–119 (2015).
Broadhurst, L. M. et al. Seed supply for broadscale restoration: Maximizing evolutionary potential. Evol. Appl. 1, 587–597 (2008).
Grossnickle, S. C. Why seedlings survive: Influence of plant attributes. New For. 43, 711–738 (2012).
Grossnickle, S. C. & MacDonald, J. E. Why seedlings grow: Influence of plant attributes. New For. 49, 1–34 (2018).
Chambers, J. C. Seed movements and seedling fates in disturbed sagebrush steppe ecosystems: Implications for restoration. Ecol. Appl. 10, 1400–1413 (2000).
Broadhurst, L. & Boshier, D. H. Seed provenance for restoration and management: Conserving evolutionary potential and utility. In Genetic Considerartions in Ecosystem Restoration Using Native Tree Species (eds Bozzano, M. et al.) 27–37 (Elsevier, 2014).
Gratani, L., Meneghini, M., Pesoli, P. & Crescente, M. F. Structural and functional plasticity of Quercus ilex seedlings of different provenances in Italy. Trees 17, 515–521 (2003).
Ovalle, J. F., Arellano, E. C. & Ginocchio, R. Trade-offs between drought survival and rooting strategy of two south American mediterranean tree species: Implications for dryland forests restoration. Forests 6, 3733–3747 (2015).
Lumaret, R., Mir, C., Michaud, H. & Raynal, V. Phylogeographical variation of chloroplast DNA in holm oak (Quercus ilex L.). Mol. Ecol. 11, 2327–2336 (2002).
Baldocchi, D. D. & Xu, L. What limits evaporation from Mediterranean oak woodlands—The supply of moisture in the soil, physiological control by plants or the demand by the atmosphere?. Adv. Water. Resour. 30, 2113–2122 (2007).
Mediavilla, S. & Escudero, A. Stomatal response to drought at a Mediterranean site: A comparative study of co-occurring woody species differing in leaf longevity. Tree Physiol. 23, 78–996 (2003).
Fotelli, M. N., Radoglou, K. M. & Constantinidou, H. I. A. Water stress responses of seedlings of four Mediterranean oak species. Tree Physiol. 20, 1065–1075 (2000).
Tsakaldimi, M., Zagas, T., Tsitsoni, T. & Ganastas, P. Root morphology, stem growth and field performance of seedlings of two Mediterranean evergreen oak species raised in different container types. Plant Soil 278, 85–93 (2005).
Jacobs, D. F. et al. Nursery stock quality as an indicator of bottomland hardwood forest restoration success in the Lower Mississippi River Alluvial Valley. Scand. J. For. Res. 27, 255–269 (2012).
Villar-Salvador, P. et al. Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New For. 43, 755–770 (2012).
Banister, J. R. et al. Major bottlenecks for the restoration of natural forests in Chile. Restor. Ecol. 26(6), 1039–1104 (2018).
Aphalo, P. & Rikala, R. Field performance of silver-birch planting-stock grown at different spacing and in containers of different volume. New For. 25, 93–108 (2003).
Puértolas, J., Gil, L. & Pardos, J. A. Effects of nutritional status and seedling size on field performance of Pinus halepensis planted on former arable land in the Mediterranean basin. Forestry 76, 159–168 (2003).
Chirino, E., Vilagrosa, A., Hernández, E. I., Matos, A. & Vallejo, V. R. Effects of a deep container on morpho-functional characteristics and root colonization in Quercus suber L. seedlings for reforestation in Mediterranean climate. For. Ecol. Manage. 256, 779–785 (2008).
Villar-Salvador, P., Planelles, R., Enriquez, E. & Peñuelas-Rubira, J. Nursery cultivation regimes, plant functional attributes and field performance relationships in the Mediterranean oak Quercus ilex L.. For. Ecol. Manage. 196, 257–266 (2004).
Oliet, J. A. et al. Field performance of Pinus halepensis planted in Mediterranean arid conditions: Relative influence of seedling morphology and mineral nutrition. New For. 37, 313–331 (2009).
Cuesta, B., Villar-Salvador, P., Puértolas, J., Jacobs, D. F. & Rey Benayas, J. M. Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For. Ecol. Manage. 260, 71–78 (2010).
de la Barrera, F., Barraza, P., Favier, Ph., Ruiz, V. & Quense, J. Megafires in Chile 2017: Monitoring multiscale environmental impacts of burned ecosystems. Sci. Total. Environ. 637–638, 1526–1536 (2018).
Becerra, P. I., Cruz, G., Ríos, S. & Castelli, G. Importance of irrigation and plant size in the establishment success of different native species in a degraded ecosystem of central Chile. Bosque 34, 23–24 (2013).
Donoso, C. Reseña ecológica de los bosques Mediterráneos de Chile. Bosque 4, 117–146 (1982).
Giliberto, J. & Estay, H. Seasonal water stress in some Chilean Matorral shrubs. Bot. Gaz. 139, 236–240 (1978).
Hoffmann, A. & Kummerow, J. Root studies in the Chilean Matorral. Oecologia 69, 57–69 (1978).
Brito, C. E., Bown, H. E., Fuentes, J. P., Franckm, N. & Perez-Quezada, J. F. Mesophyll conductance constrains photosynthesis in three common sclerophyllous species in Central Chile. Rev. Chil. Hist. Nat. 87, 8 (2014).
Bown, H. E., Fuentes, J. P. & Martínez, A. M. Assessing water use and soil water balance of planted native tree species under strong water limitations in Northern Chile. New For. 49, 871–892 (2018).
Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 143, 1–10 (2005).
Rab, M. A. Soil physical and hydrological properties following logging and slash burning in the Eucalyptus regnans forest of southeastern Australia. For. Ecol. Manage. 84, 159–175 (1996).
Boyd, C. S. & Davies, K. W. Shrub microsite influences post-fire perennial grass establishment. Rangel. Ecol. Manage. 63, 248–252 (2010).
Boucher, D. et al. How climate change might affect tree regeneration following fire at northern latitudes: A review. New For. 51, 543–571 (2020).
Miller, J. D. & Thode, A. E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens. Environ. 109, 66–80 (2007).
Parks, S. A., Dillon, G. K. & Miller, C. A new metric for quantifying burn severity: The relativized burn ratio. Remote Sens. 6, 1827–1844 (2014).
Box, G. E. P. & Cox, D. R. An analysis of transformations. J. R. Stat. Soc. Ser. B 26, 211–252 (1964).
Vander Mijnsbrugge, K., Bischoff, A. & Smith, B. A question of origin: Where and how to collect seed for ecological restoration. Basic Appl. Ecol. 11, 300–311 (2010).
Trubat, R., Cortina, J. & Vilagrosa, A. Nutrient deprivation improves field performance of woody seedlings in a degraded semi-arid shrubland. Ecol. Eng. 37(8), 1164–1173 (2011).
Donoso, S., Peña, K., Pacheco, C., Luna, G. & Aguirre, A. Respuesta fisiológica y de crecimiento en plantas de Quillaja saponaria y Cryptocarya alba sometidas a restricción hídrica. Bosque 32, 187–195 (2011).
Renninger, H. J., Miles, C. H. & Ezzel, A. W. Seasonal physiology and growth of planted oaks with implications for bottomland hardwood restoration. New For. 51, 191–212 (2020).
Vallejo, V. R. et al. Restoration strategies and actions in Mediterranean degraded lands. In Desertification in Europe: Mitigation Strategies, Land Use Planning (eds Enne, G. et al.) 221–233 (Elsevier, 2000).
Ward, J. S., Gent, M. P. N. & Stephens, G. R. Effects of planting stock quality and browse protection-type on height growth of northern red oak and eastern white pine. For. Ecol. Manage. 127, 205–216 (2000).
Wilson, B. C. & Jacobs, D. F. Quality assessment of temperate zone deciduous hardwood seedlings. New For. 31, 417–433 (2006).
Del Campo, A. D., Navarro, R. M. & Ceacero, C. J. Seedling quality and field performance of commercial stocklots of containerized holm oak (Quercus ilex) in Mediterranean Spain: An approach for establishing a quality standard. New For. 39, 19–37 (2010).
Trubat, R., Cortina, J. & Vilagrosa, A. Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J. Arid Environ. 74, 491–497 (2010).
Huber, A., Iroumé, A. & Bathurst, J. Effect of Pinus radiata plantations on water balance in Chile. Hydrol. Process. 22, 142–148 (2008).
Espinoza, S., Yáñez, M., Magni, C., Santelices, R. & Cabrera, A. Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size. iForest 13, 33–40 (2020).
Lortie, C. J. & Aarssen, L. W. The specialization hypothesis for phenotypic plasticity in plants. Int. J. Plant. Sci. 157, 484–487 (1996).
Acknowledgements
Seedlings of the experiments were kindly provided by the Vivero Quivolgo from Forestal Arauco S.A. Special thanks to Carolina Rojas from Forestal Arauco S.A., for helping us to obtain seedlings. We are indebted with those who provided support in the fieldwork, especially Nicole Toro, Cristian Hernández, Gustavo Hernández, and Francisco Méndez, from the Universidad de Chile.
Author information
Authors and Affiliations
Contributions
Formulated the idea of the manuscript: S.E.E. Designed and performed the experiment: S.E.E. and C.R.M. Analysed the data: S.E.E. and M.A.Y. Wrote the manuscript: S.E.E. Edited the manuscript: M.A.Y., M.R.C.B., S.J.V., J.G., E.E.M. and C.R.M. All authors gave final approval for publication and agree to be held accountable for the work performed therein.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Espinoza, S.E., Yañez, M.A., Martínez, E.E. et al. Influence of provenance origin on the early performance of two sclerophyllous Mediterranean species established in burned drylands. Sci Rep 11, 6212 (2021). https://doi.org/10.1038/s41598-021-85599-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-85599-3
- Springer Nature Limited