
Abstract
Diverse taxa have undergone phenological shifts in response to anthropogenic climate change. While such shifts generally follow predicted patterns, they are not uniform, and interspecific variation may have important ecological consequences. We evaluated relationships among species’ phenological shifts (mean flight date, duration of flight period), ecological traits (larval trophic specialization, larval diet composition, voltinism), and population trends in a butterfly community in Pennsylvania, USA, where the summer growing season has become warmer, wetter, and longer. Data were collected over 7–19 years from 18 species or species groups, including the extremely rare eastern regal fritillary Speyeria idalia idalia. Both the direction and magnitude of phenological change over time was linked to species traits. Polyphagous species advanced and prolonged the duration of their flight period while oligophagous species delayed and shortened theirs. Herb feeders advanced their flight periods while woody feeders delayed theirs. Multivoltine species consistently prolonged flight periods in response to warmer temperatures, while univoltine species were less consistent. Butterflies that shifted to longer flight durations, and those that had polyphagous diets and multivoltine reproductive strategies tended to decline in population. Our results suggest species’ traits shape butterfly phenological responses to climate change, and are linked to important community impacts.
Similar content being viewed by others


Introduction
Phenological changes are among the most noticeable responses by plants and animals to anthropogenic climate change1,2,3. Although some taxa may fail to respond, or respond in ways that are maladaptive4, others may undergo evolutionary change or respond via phenotypic plasticity5. Among animals, more pronounced changes and faster responses often arise in ectotherms than in endotherms6, likely because the increased ambient temperatures and changes in precipitation associated with climate change have more direct effects on the metabolic rates, activity patterns, and developmental rates of ectotherms. Among butterflies, which have become prominent ectotherm models, several general patterns are now clear: many species have advanced their date of first seasonal appearance7,8,9, prolonged their duration of seasonal activity10 and increased their number of generations per season8. Nonetheless, phenological changes in response to climate change are far from uniform6, and can vary in magnitude and direction even among species experiencing similar environmental conditions11.
Understanding variation in phenological response is an important scientific and conservation challenge, because phenological changes may determine which species’ populations are harmed—such as via trophic mismatches (e.g., via desynchronization with a host plant12), lack of sufficient time to breed one last generation (the “lost generation” hypothesis;13,14), or limitations on species’ capacities to mitigate extreme temperatures or drought (such as via aestivation/diapause4)—and which are able to respond rapidly and effectively to environmental change15,16 through modification of phenology to correspond to changing climatic conditions17,18. Further, a lack of understanding of inter-specific variation hinders scientists’ and managers’ ability to predict shifts in species’ phenology and complicates the identification and management of species at high risk from climate change.
Several studies have suggested that species-specific ecological traits may predict the extent and direction of phenological shift2,19. Specifically, the intensity of phenological responses to climate change may be influenced by species-specific ecological traits such as larval diets (e.g., more constrained activity window in woody plant feeders8), number of generations (e.g., increased numbers of generations in multivoltine species8 or the facilitation of another generation in univoltine species20), habitat use (e.g., earlier spring emergence in more open habitats21,22), adult thermoregulation behavior (e.g., high-temperature dwellers are expanding at the expense of low-temperature dwellers6,15), or seasonal occurrence (e.g., spring broods show more pronounced advances in flight times than late-season fliers23, but see24 ). Thus, analysis of phenological responses to climate change in the context of species‐specific ecological traits provides a potentially powerful framework for identifying vulnerable species25,26,27,28 and making more robust generalizations about species’ phenological responses to climate change29.
In this study, we sought to improve understanding of how butterfly phenology (timing and duration of the flight period) has shifted over time, and to evaluate how species’ traits (larval trophic specialization, larval diet composition, voltinism) mediate these phenological shifts. We also examined changes in local climatic variables (temperature, aridity, and growing degree days) during the study period and over recent decades, and we sought to evaluate the sensitivity of phenological variation to seasonal fluctuations in temperature. We further related species’ traits and phenological patterns with the butterflies’ long-term population trends. We focused on late-flying butterflies in a temperate grassland community in Pennsylvania, USA. Phenology was derived from standardized repeated surveys of butterflies during late spring, summer, and early autumn over 19 years (1998–2016) for the extremely rare eastern regal fritillary Speyeria idalia idalia, and over 7–10 years (most recent years ending in 2016) for 17 other studied organisms.
We expected butterflies overall to be sensitive to temperature variation and, in accordance with predictions for climate change, to gradually shift seasonal flight periods to begin earlier and last for longer durations over the years since the beginning of the study period30. We also expected, however, that phenological changes would be associated with species-specific diets and reproduction strategies, as follows. First, we expected less pronounced phenological changes (both seasonal timing and duration) over the years in butterfly species with more restricted larval diets (oligophagous feeders) than those with more generalized larval diets (polyphagous feeders). This is because the direct response of oligophagous species to abiotic conditions altered by climate change may be tempered by their high dependence on their host plants1. Second, we expected larval woody feeders to show more pronounced changes in the seasonal timing and duration of the adult flight period compared to larval herb feeders since herbs can produce fresh shoots throughout the season while leaves of woody plants are available only for a short time period8,31 and the newly flushing leaves of woody plants are increasingly appearing earlier in response to climate change32. Thus, woody feeders may track the earlier availability of fresh leaves of woody plants, and transition to the adult flighted stage earlier. We further expected that, because flowering plants used by adults may undergo more limited temporal shifts than the host plants used by larvae, these same woody-feeding butterflies would also increase the overall duration of their flight period. Third, we expected univoltine butterfly species (those with only one generation per season) to show less pronounced phenological shifts than multivoltine species, which generally exhibit more plastic responses to variable environmental conditions33,34, and which have the ability to respond to longer growing seasons with an increased number of generations in a season35. Fourth, we assumed that each organism would be under continuous selective pressure to match its phenology to environmental conditions, but that more specialized or less variable ecological traits would constrain genetic or plastic shifts in behavior. Thus, we expected that species exhibiting oligophagy and univoltinism would exhibit lower sensitivity to annual temperature fluctuations than other species36. Finally, butterflies may face divergent pressures while responding both to altered abiotic conditions and host plants that separately shift phenology with climate change37. Such separate pressures could trigger trophic mismatches in phenology1,16, which could reduce survival38 most acutely among specialized butterflies. Similarly, limitations on individual longevity, increased mortality, or an inability to add additional generations in a season could constrain univoltine species’ response to longer growing seasons. Thus, in the context of ongoing climate change, we expected populations of oligophagous and univoltine species would fare worse over time than polyphagous or multivoltine species.
Results
Changes in local climate
Over recent decades (1981–2016), the growing season at the field site became progressively warmer, wetter, and longer (Fig. 1). Mean annual temperature revealed a significant upward trend with time (Mann–Kendall Test; t = 149, P = 0.02, Slope estimate = 0.02), with warming of about 0.7 °C over the 35-year period. The mean of the annual aridity index showed a significant upward trend (Mann–Kendall Test; t = 203, P = 0.002, Slope estimate = 0.33), signifying progressively wetter conditions with an increase of 1.05 mm/ °C over the period. There was also a significant positive trend in growing degree days (GDD) (Mann–Kendall Test; t = 215, P = 0.001, Slope estimate = 0.025), signifying a progressively longer growing season with an increase of 5.25 °C GDD over the period.

Positive trends in (a) mean annual temperature, (b) aridity, and (c) growing degree days across a 35-year period at Fort Indiantown Gap National Guard Training Center (FIG-NGTC). Note that for the Aridity index, the smaller the value the higher the level of aridity. Solid lines = annual change, dotted lines = linear regression line over time.
Temporal trends in butterfly phenology and abundance
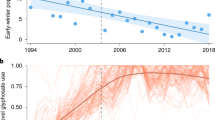
Applying individual median-based linear models we found that the majority (72%) of our organisms shifted either the mean date or the duration of their flight period or both (Table 1). Specifically, six of the 18 taxa (Boloria bellona, Euptoieta claudia, Papilio glaucus, Papilio polyxenes, both sexes of S. idalia idalia, and Colias eurytheme/philodice) advanced their mean flight date over time, while three others (Phyciodes tharos, Polygonia interrogationis/comma, Satyrodes eurydice/Enodia anthedon) showed some delay. Shifts in mean date were marginal in two of these taxa, B. bellona (P = 0.07) and P. interrogationis/comma (P = 0.06) (Table 1, Fig. S1). In addition, seven taxa prolonged the duration of the flight period (Epargyreus clarus, Euptoieta claudia, Limenitis archippus, S. idalia idalia (F), P. interrogationis/comma, Satyrodes eurydice/Enodia anthedon, Speyeria cybele/aphrodite) over time, while Papilio troilus shortened its duration (Table 1, Fig. S1). Shifts in duration were marginal in two of these taxa, L. archippus (P = 0.07) and P. interrogationis/comma (P = 0.06).
Population trends based on repeated counts at various sites were estimated on a model-based imputation model. Overall, populations were found to have an upward trend for most of the studied organisms (69%) (Table 1). However, when comparing assemblages by ecological traits, populations increased over time in a greater portion of oligophagous (80%) than polyphagous (50%) species, and a much greater portion of univoltine (100%) than multivoltine species (31%). (Table 1, Fig. 2). Further, all but one of the species found to have declined were both polyphagous and multivoltine (Table 1).

Population shifts grouped per ecological trait. Species for which model did not converge or population trend was not important are not shown. The two sexes of S. idalia idalia are treated as separate entities.
Surprisingly, phylogenetic regressions showed that there was a negative relationship between shifts in flight period and population trend (F-statistic: 5.8, slope = -0.06, P = 0.02). That is, species that shortened flight duration over time tended to increase in population while species that lengthened their flight duration tended to decrease in population (Table 1). No relationship between shifts in mean date and population trend was detected (P > 0.05).
Sensitivity of species traits to phenological and temperature change
Phylogenetic regressions showed that species differing in larval trophic specialization increasingly differed in mean date over the years (F-statistic = 4.5, P = 0.05), with oligophagous species increasingly flying later in the season and polyphagous species increasingly flying earlier (Fig. 3a, Table S1). Similarly, species differed by larval diet composition (F-statistic = 8.1, P = 0.01), with woody feeders increasingly flying later and herb feeders increasingly flying earlier (Fig. 3b, Table S1). No relationship was detected between voltinism and mean date (P > 0.05). Only a marginal difference in duration by larval trophic specialization was detected (F-statistic = 3.5, P = 0.07), with a tendency toward shorter flight periods in oligophagous and longer flight periods in polyphagous species (Fig. 4, Table S1). No relationship was detected between larval diet composition and duration, or voltinism and duration (P > 0.05 in all cases).

Sensitivity of larval trophic specialization (LTS) and larval diet composition (LDC) on shifts in mean date of appearance over the years. Error bars depict predicted mean values (circle) and standard error (upper and lower horizontal lines). LTS is shown on panel (a) (oligophagous in light red; polyphagous in deep red) and LDC is shown on panel (b) (herb feeders in light green; woody feeders in deep green).

Sensitivity of larval trophic specialization (LTS) on shifts in duration of the flight period over the years. Error bars depict predicted mean values (circle) and standard error (upper and lower horizontal lines).
Lastly, species differed by voltinism in the extent to which the flight period was sensitive to the spring–summer (F-statistic = 5.8, P = 0.03) (Fig. 5a, Table S1) and previous autumn–winter (F-statistic = 7, P = 0.02) temperatures (Fig. 5b, Table S1). Specifically, multivoltine species substantially prolonged the duration of the flight period in response to warmer temperatures during both seasonal periods, and especially the spring–summer temperatures, while univoltine species slightly prolonged the flight period in response to warmer spring–summer temperatures but modestly decreased them in response to warmer temperatures in the previous autumn and winter (Fig. 5). No relationship was detected by larval trophic specialization or larval diet composition in the slope of duration versus either seasonal temperature (P > 0.05 in all cases). Likewise, no relationship was detected by any of the three ecological traits in the slope of mean date versus either seasonal temperature (P > 0.05 in all cases). Finally, no relationship was detected by any of the three ecological traits in either the slope of the mean date or the slope of duration versus annual (12-month) temperature (P > 0.05 in all cases).

Sensitivity of voltinism on shifts in duration of the flight period over the seasonal fluctuations of temperature. Error bars depict predicted mean values (circle) and standard error (upper and lower horizontal lines). Slope of the relationship between flight period and spring–summer temperature is shown on panel (a) and slope of the relationship between flight period and autumn–winter temperature is shown on panel (b) (multivoltine in yellow; polyphagous in orange).
Discussion
The phenology of life history events represents a central mechanism by which species respond to environmental change, and recent shifts in phenology provide a crucial window into how species will respond to accelerating climate change. In this study, we sought to understand how butterfly phenology has shifted over time, how species’ ecological traits influence these shifts, and the consequences for butterfly populations. We found phenological shifts over time in 13 of the 18 butterfly taxa studied. Local increases in temperature, humidity, and the length of the growing season may suggest potential associations of the observed phenological shifts with climate change. We observed that shifts in mean flight date varied among assemblages differing in larval trophic specialization and larval diet composition. Shifts in flight duration also differed by larval trophic specialization. Further, species assemblages differed by voltinism in how flight duration was influenced by seasonal temperature. In addition, species with different ecological traits (larval trophic composition and voltinism) exhibited divergent population trends and shifts in flight duration were negatively associated with population trend. Together, these results suggest species’ ecological traits mediate the direction and intensity of climate-driven phenological shifts, and that these shifts have important consequences for population dynamics in the context of climate change.
Phenological change by taxon
Phenological studies have mostly focused on early and spring flyers9,39, but here we demonstrate that flyers of late summer or summer broods of multivoltine species can also be responsive to climate change. First, 1/3 of the studied taxa advanced their mean flight date over time (by 0.59–5.04 days/year). Such phenological advances in the timing of butterfly flight are expected under warmer climate scenarios9,10,40. No changes were detected over time in half of the species. This may be because of limited time period (7–10 years) most species were observed. It may also be because we did not include early flyers in our analysis, which may have limited our ability to detect species responses during springtime, a period in which species responses are most pronounced2,24,41. A contrasting pattern, with a substantial delay in mean flight date over time, was observed in three taxa (1.16–3.48 days/year) and was most clearly evident in P. tharos. The distribution and flight behavior of this butterfly species is highly related to host plant availability (asters, milkweeds, thistles and sunflowers)42. However, no similar delays for species that use the same host plants (e.g. P. glaucus, P. polyxenes) were observed37. Another plausible explanation could be an extended aestival diapause because of the long photoperiod and high and increasing temperatures in the local summer climate.
Second, in accordance with expectations for warmer conditions21, seven of the butterflies prolonged their flight period (by 0.32–4.76 days per year). Of particular interest for conservation was that S. i. idalia, a globally threatened species that has begun to recover after a long population decline43, exhibited both a longer flight duration and earlier mean flight date. The lack of change we observed in the flight duration of ten taxa could again be attributed to the limited 7–10 year period that some of the studied organisms had been observed, but it is also consistent with the idea that local habitats can buffer ecological communities against coarse-scale trends and patterns of ongoing environmental change22,44. For instance, fine-scale habitat heterogeneity can create microrefugia, which may mediate species’ responses to climate changes45. In contrast to expectations, one species, P. troilus, shortened the duration of its flight period (0.4 days/year). Differences among taxa in observed phenological change may reflect not only direct effects of climate on butterflies, but also indirect effects of climate on the phenologies of host-plants46.
The role of ecological traits in mediating phenological shifts over time
One key factor shaping changes in species’ phenology across years was larval nutrition. Oligophagous species delayed and shortened their flight periods while polyphagous species advanced and lengthened theirs. The expression of opposite phenological patterns suggests a different dynamic on species groups’ resilience to environmental change. For instance, species with generalized feeding habits have a wider niche and an ecological advantage to survive in areas with unpredictable and changing environment47. The large species pool used as food by polyphagous feeders may be a beneficial life history trait that allows organisms to maximize fitness as they track environmental change and respond via seasonally plastic strategies.
Another feeding trait influencing the magnitude of phenological change was larval diet composition. Herb feeders advanced their mean flight date whereas woody feeders showed moderate delays. Previous plant studies, including those ranging from the level of single individuals to entire biomes, have documented a clear advancement of leaf flushing in woody plants in the temperate zone in response to climate change48 but the response of leaf flushing is highly variable because woody plants also depend on cold temperatures to break bud dormancy49. If winter warming leads to insufficient bud dormancy49, woody plants will postpone the onset of newly flushing leaves50, also affecting the phenology of woody feeders that depend on their resources51. Moreover, at temperate latitudes, winter warming may increase intraspecific variation in leafing and flowering dates52. In addition, woody feeders often target the newly flushed leaves because of new leaves’ nutritional profile and/or less well-developed defenses53, but the association between plant palatability and shifts in phenology represents a largely overlooked pattern in the ecology of temperate butterflies. Together, these disparate plant responses to climate change could in turn alter butterfly community composition, by increasingly shifting the timing of herb and woody feeders’ life history events in the coming years.
The influence of ecological traits in mediating phenological responses to seasonal temperature
Organisms often exhibit heterogeneity in their sensitivity to environmental factors54. It had previously been unknown, however, whether voltinism influenced the sensitivity of butterfly phenologies to seasonal temperature. We found multivoltine butterflies had high sensitivity to seasonal fluctuations in temperature, with consistent increases in flight duration in response to warmer seasonal temperatures, especially spring–summer temperatures. In contrast, univoltine species exhibited more modest shifts that increased flight duration with warmer spring–summer temperatures but decreased flight duration with warmer autumn–winter temperatures. These results accord with previous evidence for the higher responsiveness to climate of the multivoltine reproductive strategy. For instance, multivoltine species have greater synchrony in mean flight date across temperature gradients45, and they exhibit plastic phenological variation tied to annual climatic conditions and habitat use22. Further, warmer ambient conditions have increased the number of butterfly generations in dozens of temperate butterflies55, and can even facilitate a second generation in principally univoltine species20, where such species are constrained solely by the length of the growing season from having a second generation.
In our study, ecological traits were not otherwise associated with the relationship between phenology and seasonal temperatures. It should be noted, however, that phenology could be associated with variations in temperature that occur at a finer spatial or temporal scale than those measured in the aggregate with our site-level and season-long climate variables.
Population consequences of climate and phenological changes
Earlier emergence or mean flight dates have been widely documented1,9,15,31,56,57 and have often been assumed to enable species to adjust to a changing climate. Shifts in mean flight dates were not related to population change across all species in our study, however. We did observe that, while not universal across all species, changes to flight duration were generally negatively related to changes in population over time. For instance, the P. interrogationis/ comma species group, which had the greatest increase in flight duration of any species (at 4.8 days per year), also had the largest population decline of all species (0.2 individuals observed per transect walk per year). The reasons for this pattern are unclear, but one potential mechanism could derive from the changing availability of floral resources at a community scale. In temperate zones, climate warming is expected to advance the phenology of early-flowering plants and delay it in late-flowering ones, and this divergent pattern of plant phenological shift could create a mid-summer gap in floral resources that might disproportionately affect species with longer summer activity periods37.
Overall, we observed an increasing population trend for most (69%) of our species pool, though percentages differed by assemblage: high portions of oligophagous and univoltine species exhibited population increases, while declining species were almost always both polyphagous and multivoltine. This contrasted with previous expectations that the greater plasticity exhibited by generalist feeders and via a reproductive strategy with multiple broods per season would confer an ecological advantage in a variable environment, and that this advantage would extend to directional environmental change caused by climate change47. For multivoltine species, one possible explanation is the lost generation hypothesis: if an extended growing period leads juvenile stages to complete development in late summer instead of entering the over-wintering stage, the chance of losing this generation is high and population declines may occur14,58. The declines we observed among multivoltine species further highlight that species exhibiting greater responsiveness to climate do not necessarily fare better than less responsive ones, at least in the short term. In contrast, evolved strategies in univoltine species, such as the use of summer aestivation, may have allowed them to effectively respond to longer periods of thermal stress and drought, and to climate-driven shifts in plant phenology.
Even so, it is unclear whether strategies that have thus far benefitted univoltine butterflies will continue to prove beneficial as climate change accelerates. For instance, in univoltine species like Maniola or Speyeria butterflies, in which important life history events like summer dormancy or oviposition are induced by photoperiod59, shifts in plant phenology and climate might lead to an increasing temporal mismatch with optimal conditions for growth, survival, and reproduction58. Alternatively, the use of extended aestivation to adjust to a warming climate could incur several costs, including shortened activity periods needed for mating or to meet nutritional needs prior to entering aestivation, increased susceptibility to predation or disease during the longer aestivation period, or heightened survival risk if the increasingly warmer or drier conditions during aestivation exceed the butterflies’ environmental tolerances4.
Methods
Study site
Data were collected at Fort Indiantown Gap National Guard Training Center (FIG-NGTC), a military training area in southeastern Pennsylvania, USA (40°26′13.15′'N, 76°34′33.8′'W). The landscape consists of a mosaic of forests, semi-natural grasslands, and rangelands heavily used for military training. FIG-NGTC extends over 6920 ha, with altitudes from 110 to 437 m above mean sea level60. We focused on grassland habitat (88.22 ha) within this mosaic.
The climate is humid continental with February the driest and May the most humid month. The grassland habitat of FIG-NGTC is of particular interest for conservation both because it hosts a diverse native butterfly community and because it harbors the only remaining viable population of an extremely rare butterfly, S. i. idalia61,62, which is Critically Imperiled63.
Butterfly species
For 19 years (1998–2016), counts of S. i. idalia were made weekly along five fixed routes in grassland habitats by trained biologists and volunteers, following the Pollard walk transect method64. Since S. i. idalia is a large, sexually-dichromatic butterfly that is detectable and can be identified to species and sex from a distance, surveys focused on an 18.3 m (20 yards) band on either side of the transect. Sampling included the months of June to September in all years, but sampling sometimes began earlier (in May) or extended later (until October), when weather conditions were suitable for butterfly activity. Beginning in 2007 and again in 2008, additional species and species groups were recorded during transect walks, with the same methods. From 2010 onwards, sampling was further extended to all identifiable species or species groups in the butterfly community. While this yields observations of just 7–10 years in most species, butterflies have been found to respond to climate change over similar time frames in other studies15,65.
These butterfly surveys resulted in a monitoring dataset that included 41 species or species groups (sets of species with similar appearance lumped together in field data because they could not be properly distinguished visually at a distance) that were consistently identifiable during surveys. From this dataset, we removed observations that could contribute to bias, as follows. First, we removed species groups if different species within the same group had different traits of interest (6 species groups excluded). The species groups that were retained were always pairs of species and these pairs were more closely related to each other than to other taxa in this analysis. Second, to avoid trying to draw conclusions from small sample sizes, we removed species that were observed infrequently (≤ 4 counts per year or ≤ 4 years of records) from analyses (16 species). Third, we excluded an early univoltine flyer for which the emergence day was before sampling began. For multivoltine species included in the analysis we also ignored the first flight period of each year (which sometimes was also before sampling), and focused on the second flight period, which always fell during the sampling period. For Epargyreus clarus the mean date and duration are calculated from the total time period encompassing the two flight periods. The resulting dataset included 14 species and 4 species groups (hereafter, “taxa”; Table 1). The dataset also included separate data on both sexes of S. i. idalia because, unlike all other species in this analysis, a different phenology characterizes each sex43. Males emerge in June and die after mating in July, while females emerge in July and after a summer dormancy, they oviposit their eggs from late August to mid-September and then die.
Species traits
For each of these 18 taxa, we then determined three life-history traits that are likely correlated with changes in flight period in response to climate change40,56. These included (i) larval trophic specialization (oligophagous versus polyphagous)8 ; (ii) the larval diet composition (woody versus herb feeders)40 ; (iii) voltinism (univoltine versus multivoltine species)25,40 (Table 1). Data on these life history traits were primarily from field observations by the authors and secondarily from field guides66,67.
Phenology variables
Two phenological variables were calculated for each butterfly species each year. The first variable, the weighted mean date of adult flight (“mean date” hereafter) was calculated as:
where pk is the relative abundance of species (nk is the number of individuals of species k per visit, Nk is the total number of individuals of species k per year), t is the date in Julian days (1 = January 1st), and J is the last observation date56. The second variable, the duration of the flight period, was calculated as the SD about the mean date9,68,69. These variables are less sensitive to sampling effort, extreme events, and population trends than variables like first observation or time between first and last observation9,21,70.
Butterfly phylogeny
Traits of closely related taxa such as food source preferences, number of broods and temporal sensitivities to temperature fluctuations may be similar due to common ancestry and hence statistically dependent in comparative analyses71. In order to account for phylogenetic relatedness we used the ultrametric phylogenetic tree published by72 to construct a phylogenetic tree of our species pool (Fig. S2, Table S2).
Climate variables
First, to investigate whether there has been a significant directional change in local climate during the last decades, a 35-year period (1981–2016) was considered. Meteorological data were obtained for the precise location of the study area from the PRISM climate model73, which estimates temperature and precipitation variables for the continental United States, as interpolated from multiple neighboring weather stations while controlling for physiographic variables74. Climate variables examined were annual temperature (Temperature), aridity index (Aridity) and growing degree-days (GDD). Aridity index is a measure of drought, calculated as P/2T, where P is mean annual precipitation (mm) and T is mean annual temperature (°C)75. Note that the higher the aridity index value, the less arid the conditions. Cumulative seasonal heat units, expressed as growing-degree days (GDD), were used as an indirect measure of the effect of temperature on butterfly populations. The GDD value represents the accumulated number of degrees over a season where the average daily temperature is greater than a threshold temperature [(Temperaturemax + Temperaturemin)/2 − Temperaturethreshold]; this measure is known to contribute to organismal development76. Here the threshold was set as 10 °C, the minimal thermal condition conducive to butterfly development and survival40,57.
Then, to document the sensitivity of butterfly phenology to abiotic factors, we considered the primary sampling period in this study (2007–2016) and we estimated the three following climatic cues (Donoso et al. 2016): (1) a 12-month period starting with the month after the last butterfly survey (i.e., beginning in October), (2) the spring–summer (March–August) period that overlapped with the annual surveys, and (3) the autumn–winter (September—February) period prior to each annual survey.
Statistical analysis
To detect possible trends in each one of the climate time series we ran trend analyses. The significance of the trend was assessed by a nonparametric rank-based test (Mann–Kendall test) and evidence for an increasing trend versus the null hypothesis (no trend) was tested at a 0.95 level of confidence.
Next, to evaluate changes in species’ phenology over the study period, we assessed each taxon separately with median-based linear models, as this method is robust to outliers77. Time was the explanatory variable, and phenological descriptors were the responses. Positive slopes indicated mean flight date was later or duration of the flight period was longer in more recent years for the species. We used mblm() function78,79,80 in R package NSM381. Diagnostic graphs for residuals independency and homogeneity and the Shapiro–Wilk test for normality were used to check if models’ assumptions were met. In addition, we estimated population trends for each taxon. Annual counts were modeled as a function of the fixed effects of “time” (years) and “site” (a suite of five sampling grasslands) using the trim() function in the R package rtrim82. Sampling effort was similar across months, sites, and years83.
Furthermore, we evaluated whether butterflies responding appropriately to climate change (via a shift to an earlier date or a shift to longer duration) would benefit with increased population numbers (or at least remain stable) while butterflies that showed minimal or no shifts would suffer more from the effects of climate change. We therefore tested whether population trends and phenological shifts tended to co‐vary, while accounting for phylogenetic relatedness as described below.
Attributing sensitivity of species traits to shifts in phenology and seasonal temperature
We then used a phylogenetic analysis to evaluate how butterfly phenology has changed over time and in response to seasonal temperature fluctuations, and to determine whether the direction and magnitude of those changes is associated to specific ecological traits of butterflies. First, we regressed the mean date and duration by year for each of the 18 taxa, and then we included the slopes of those regressions (one per taxon) as a response variable in a comparative analysis. Then, we grouped our species pool into subsets of species (hereafter, species assemblages) on the basis of their ecological traits: (1) larval trophic specialization (oligophagous versus polyphagous species), (2) larval diet composition (woody versus herb feeders) and (3) voltinism (univoltine versus multivoltine species).
We used phylogenetic generalized least squares (PGLS) to test for an association between mean date of appearance/duration and species traits using the gls() function in R package nlme84. The slope between the mean date or duration and years was treated as response variable and species traits as categorical variables. The PGLS approach allowed us to examine relationships using the most common models for evolutionary change (Pagel's lambda, Brownian motion, Ornstein–Uhlenbeck)85. We compared model fit using Akaike’s Information Criterion (AIC) and used the model with the best fit to estimate linear regression associations. Furthermore, to test the sensitivity of butterfly phenology to climatic cues, we first calculated the relationship (the slope between mean date or duration and seasonal temperature), and then we included the relationship in the comparative analysis described earlier. To account for phylogenetic structure, we also fit PGLS trait evolution models and compared model fit using Akaike’s Information Criterion.
References
Parmesan, C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Glob. Change Biol. 13, 1860–1872. https://doi.org/10.1111/j.1365-2486.2007.01404.x (2007).
Peñuelas, J. et al. Evidence of current impact of climate change on life: A walk from genes to the biosphere. Glob. Change Biol. 19, 2303–2338. https://doi.org/10.1111/gcb.12143 (2013).
Thackeray, S. J. et al. Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments. Glob. Change Biol. 16, 3304–3313. https://doi.org/10.1111/j.1365-2486.2010.02165.x (2010).
Dapporto, L. et al. Rise and fall of island butterfly diversity: Understanding genetic differentiation and extinction in a highly diverse archipelago. Divers. Distrib. 23, 1169–1181. https://doi.org/10.1111/ddi.12610 (2017).
Hendry, A. P., Farrugia, T. J. & Kinnison, M. T. Human influences on rates of phenotypic change in wild animal populations. Mol. Ecol. 17, 20–29. https://doi.org/10.1111/j.1365-294X.2007.03428.x (2008).
Devictor, V. et al. Differences in the climatic debts of birds and butterflies at a continental scale. Nat. Clim. Change 2, 121–124. https://doi.org/10.1038/nclimate1347 (2012).
Forister, M. L. & Shapiro, A. M. Climatic trends and advancing spring flight of butterflies in lowland California. Glob. Change Biol. 9, 1130–1135. https://doi.org/10.1046/j.1365-2486.2003.00643.x (2003).
Altermatt, F. Tell me what you eat and I’ll tell you when you fly: Diet can predict phenological changes in response to climate change. Ecol. Lett. 13, 1475–1484. https://doi.org/10.1111/j.1461-0248.2010.01534.x (2010).
Stefanescu, C., Penuelas, J. & Filella, I. Effects of climatic change on the phenology of butterflies in the northwest Mediterranean Basin. Glob. Change Biol. 9, 1494–1506. https://doi.org/10.1046/j.1365-2486.2003.00682.x (2003).
Roy, D. B. & Sparks, T. H. Phenology of British butterflies and climate change. Glob. Change Biol. 6, 407–416. https://doi.org/10.1046/j.1365-2486.2000.00322.x (2000).
Diez, J. M. et al. Forecasting phenology: from species variability to community patterns. Ecol. Lett. 15, 545–553. https://doi.org/10.1111/j.1461-0248.2012.01765.x (2012).
Schweiger, O., Settele, J., Kudrna, O., Klotz, S. & Kühn, I. Climate change can cause spatial mismatch of trophically interacting species. Ecology 89, 3472–3479. https://doi.org/10.1890/07-1748.1 (2008).
Glazaczow, A., Orwin, D. & Bogdziewicz, M. Increased temperature delays the late-season phenology of multivoltine insect. Sci. Rep. https://doi.org/10.1038/srep38022 (2016).
van der Kolk, H.-J., WallisDeVries, M. F. & van Vliet, A. J. H. Using a phenological network to assess weather influences on first appearance of butterflies in the Netherlands. Ecol. Indicators 69, 205–212, https://doi.org/10.1016/j.ecolind.2016.04.028 (2016).
Zografou, K. et al. Signals of climate change in butterfly communities in a mediterranean protected area. PLoS ONE 9, e87245. https://doi.org/10.1371/journal.pone.0087245 (2014).
Visser, M. Keeping up with a warming world; assessing the rate of adaptation to climate change. Proc. Biol. Sci. R. Soc. 275, 649–659, https://doi.org/10.1098/rspb.2007.0997 (2008).
Kharouba, H. M., Paquette, S. R., Kerr, J. T. & Vellend, M. Predicting the sensitivity of butterfly phenology to temperature over the past century. Glob. Change Biol. 20, 504–514. https://doi.org/10.1111/gcb.12429 (2014).
Roy, D. B. et al. Similarities in butterfly emergence dates among populations suggest local adaptation to climate. Glob. Change Biol. 21, 3313–3322. https://doi.org/10.1111/gcb.12920 (2015).
Rapacciuolo, G. et al. Beyond a warming fingerprint: Individualistic biogeographic responses to heterogeneous climate change in California. Glob. Change Biol. 20, 2841–2855. https://doi.org/10.1111/gcb.12638 (2014).
Fischer, K. & Fiedler, K. Life-history plasticity in the butterfly Lycaena hippothoe: Local adaptations and trade-offs. Biol. J. Lin. Soc. 75, 173–185. https://doi.org/10.1046/j.1095-8312.2002.00014.x (2002).
Zografou, K. Who flies first?—Habitat-specific phenological shifts of butterflies and orthopterans in the light of climate change: A case study from the south-east Mediterranean Lepidoptera and Orthoptera phenology change. Ecol. Entomol. 40, 562–574. https://doi.org/10.1111/een.12220 (2015).
Suggitt Andrew, J. et al. Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos 120, 1–8, https://doi.org/10.1111/j.1600-0706.2010.18270.x (2010).
Dell, D., Sparks, T. & Dennis, R. Climate change and the effect of increasing spring temperatures on emergence dates of the butterfly Apatura iris (Lepidoptera: Nymphalidae). Eur. J. Entomol. 102, 161–167. https://doi.org/10.14411/eje.2005.026 (2005).
Zipf, L., Williams, E. H., Primack, R. B. & Stichter, S. Climate effects on late-season flight times of Massachusetts butterflies. Int. J. Biometeorol. 61, 1667–1673. https://doi.org/10.1007/s00484-017-1347-8 (2017).
Diamond, S. E., Frame, A. M., Martin, R. A. & Buckley, L. B. Species’ traits predict phenological responses to climate change in butterflies. Ecology 92, 1005–1012. https://doi.org/10.1890/i0012-9658-92-5-1005 (2011).
Melero, Y., Stefanescu, C. & Pino, J. General declines in Mediterranean butterflies over the last two decades are modulated by species traits. Biol. Cons. 201, 336–342. https://doi.org/10.1016/j.biocon.2016.07.029 (2016).
Stefanescu, C., Peñuelas, J. & Filella, I. Butterflies highlight the conservation value of hay meadows highly threatened by land-use changes in a protected Mediterranean area. Biol. Cons. 126, 234–246. https://doi.org/10.1016/j.biocon.2005.05.010 (2005).
Sparks, T. H., Huber, K. & Dennis, R. L. H. Complex phenological responses to climate warming trends? Lessons from history. Eur. J. Entomol. 103, 379–386 (2006).
Wong, M. K. L., Guénard, B. & Lewis, O. T. Trait-based ecology of terrestrial arthropods. Biol. Rev. 94, 999–1022. https://doi.org/10.1111/brv.12488 (2019).
Gutiérrez, D. & Wilson, R. J. Intra- and interspecific variation in the responses of insect phenology to climate. J. Anim. Ecol. https://doi.org/10.1111/1365-2656.13348 (2020).
Zografou, K. et al. Butterfly phenology in Mediterranean mountains using space-for-time substitution. Ecol. Evolut. 10, 928–939. https://doi.org/10.1002/ece3.5951 (2020).
Steltzer, H. & Post, E. Seasons and life cycles. Science 324, 886–887. https://doi.org/10.1126/science.1171542 (2009).
Hale, R., Morrongiello, J. R. & Swearer, S. E. Evolutionary traps and range shifts in a rapidly changing world. Biol. Let. 12, 20160003. https://doi.org/10.1098/rsbl.2016.0003 (2016).
Ghalambor, C. K., McKay, J. K., Carroll, S. P. & Reznick, D. N. Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct. Ecol. 21, 394–407. https://doi.org/10.1111/j.1365-2435.2007.01283.x (2007).
Macgregor, C. J. et al. Climate-induced phenology shifts linked to range expansions in species with multiple reproductive cycles per year. Nat. Commun. 10, 4455. https://doi.org/10.1038/s41467-019-12479-w (2019).
Pau, S. et al. Predicting phenology by integrating ecology, evolution and climate science. Glob. Change Biol. 17, 3633–3643. https://doi.org/10.1111/j.1365-2486.2011.02515.x (2011).
Sherry, R. A. et al. Divergence of reproductive phenology under climate warming. Proc. Natl. Acad. Sci. 104, 198. https://doi.org/10.1073/pnas.0605642104 (2007).
Wilson, R. J. & Fox, R. Insect responses to global change offer signposts for biodiversity and conservation. Ecol. Entomol. https://doi.org/10.1111/een.12970 (2020).
Brooks, S. J. et al. The influence of life history traits on the phenological response of British butterflies to climate variability since the late-19th century. Ecography 40, 1152–1165. https://doi.org/10.1111/ecog.02658 (2017).
Cayton, H. L., Haddad, N. M., Gross, K., Diamond, S. E. & Ries, L. Do growing degree days predict phenology across butterfly species?. Ecology 96, 1473–1479. https://doi.org/10.1890/15-0131.1 (2015).
Stocker, T. F. et al. Summary for policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, 3–29 (2013).
Swengel, A. B. Effects of fire and hay management on abundance of prairie butterflies. Biol. Cons. 76, 73–85 (1996).
Zografou, K. et al. Severe decline and partial recovery of a rare butterfly on an active military training area. Biol. Cons. 216, 43–50. https://doi.org/10.1016/j.biocon.2017.09.026 (2017).
Gillingham, P. K., Huntley, B., Kunin, W. E. & Thomas, C. D. The effect of spatial resolution on projected responses to climate warming. Divers. Distrib. 18, 990–1000. https://doi.org/10.1111/j.1472-4642.2012.00933.x (2012).
Roy David, B. et al. Similarities in butterfly emergence dates among populations suggest local adaptation to climate. Global Change Biol. 21, 3313–3322, https://doi.org/10.1111/gcb.12920 (2015).
Lemoine, N. P. Climate change may alter breeding ground distributions of eastern migratory monarchs (Danaus plexippus) via range expansion of asclepias host plants. PLoS ONE 10, e0118614. https://doi.org/10.1371/journal.pone.0118614 (2015).
Slansky, F. Phagism relationships among butterflies. J. N. Y. Entomol. Soc. 84, 91–105 (1976).
Morin, X., Roy, J., Sonié, L. & Chuine, I. Changes in leaf phenology of three European oak species in response to experimental climate change. New Phytol. 186, 900–910. https://doi.org/10.1111/j.1469-8137.2010.03252.x (2010).
Chuine, I., Morin, X. & Bugmann, H. Warming. Photoperiods Tree Phenol. 329, 277–278. https://doi.org/10.1126/science.329.5989.277-e%JScience (2010).
Luedeling, E., Girvetz, E. H., Semenov, M. A. & Brown, P. H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 6, e20155. https://doi.org/10.1371/journal.pone.0020155 (2011).
Fu, Y. S. H. et al. Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species. Proc. Natl. Acad. USA 111, 7355–7360. https://doi.org/10.1073/pnas.1321727111%JProceedingsoftheNationalAcademyofSciences (2014).
Renner, S. S. & Zohner, C. M. Climate change and phenological mismatch in trophic interactions among plants, insects, and vertebrates. Annu. Rev. Ecol. Evol. Syst. 49, 165–182. https://doi.org/10.1146/annurev-ecolsys-110617-062535 (2018).
Barton, K. E., Edwards, K. F. & Koricheva, J. Shifts in woody plant defence syndromes during leaf development. Funct. Ecol. 33, 2095–2104. https://doi.org/10.1111/1365-2435.13435 (2019).
Cohen, J. M., Lajeunesse, M. J. & Rohr, J. R. A global synthesis of animal phenological responses to climate change. Nat. Clim. Change 8, 224–228. https://doi.org/10.1038/s41558-018-0067-3 (2018).
Altermatt, F. Climatic warming increases voltinism in European butterflies and moths. Proc. R. Soc. B Biol. Sci. 277, 1281–1287. https://doi.org/10.1098/rspb.2009.1910 (2010).
Illán, J. G., Gutiérrez, D., Díez, S. B. & Wilson, R. J. Elevational trends in butterfly phenology: Implications for species responses to climate change. Ecol. Entomol. 37, 134–144. https://doi.org/10.1111/j.1365-2311.2012.01345.x (2012).
Nufio, C. R., McGuire, C. R., Bowers, M. D. & Guralnick, R. P. Grasshopper community response to climatic change: Variation along an elevational gradient. PLoS ONE 5, e12977. https://doi.org/10.1371/journal.pone.0012977 (2010).
Van Dyck, H., Bonte, D., Puls, R., Gotthard, K. & Maes, D. The lost generation hypothesis: Could climate change drive ectotherms into a developmental trap?. Oikos 124, 54–61. https://doi.org/10.1111/oik.02066 (2015).
Scott, J. A. The Butterflies of North America: A Natural History and Field Guide. (Stanford University Press, 1992).
Division, E. Final Integrated Natural Resources Management Plan 17003–25002 (The Pennsylvania Department of Military and Veterans Affairs, Annville, 2002).
Shuey, J. et al. Landscape-scale response to local habitat restoration in the regal fritillary butterfly (Speyeria idalia) (Lepidoptera: Nymphalidae). J. Insect Cons. 20, 773–780. https://doi.org/10.1007/s10841-016-9908-4 (2016).
Metzler, E., Shuey, J., Ferge, L., Henderson, R. & Goldstein, P. Contributions to the understanding of tallgrass prairie-dependent butterflies and moths (Lepidoptera) and their biogeography in the United States. Ohio Biol. Surv. Bull. New Ser. 15, 1–143 (2005).
PNHP. PNHP Species Lists. Pennsylvania Natural Heritage Program. http://www.naturalheritage.state.pa.us/Species.aspx (2019).
Pollard, E. & Yates, T. J. Monitoring Butterflies for Ecology and Conservation (1993).
Nufio, C. R., McGuire, C. R., Bowers, M. D. & Guralnick, R. P. Grasshopper community response to climatic change: Variation along an elevational gradient. PLoS ONE https://doi.org/10.1371/journal.pone.0012977 (2010).
Glassberg, J. Butterflies through binoculars, the East. A field guide to the butterflies of Eastern North America, 242. (Oxford University Press, Inc., 1999).
Brock, J. P. & Kaufman, K. Field Guide to Butterflies of North America., 391 (Hillstar Editions L.C, 2003).
Brakefield, P. M. Geographical variability in, and temperature effects on, the phenology of Maniola jurtina and Pyronia tithonus (Lepidoptera, Satyrinae) in England and Wales. Ecol. Entomol. 12, 139–148. https://doi.org/10.1111/j.1365-2311.1987.tb00993.x (1987).
de Arce Crespo, J. I. & Gutiérrez, D. Altitudinal trends in the phenology of butterflies in a mountainous area in central Spain. Eur. J. Entomol. 108, 651–658 (2011).
Moussus, J.-P., Julliard, R. & Jiguet, F. Featuring 10 phenological estimators using simulated data. Methods Ecol. Evol. 1, 140–150. https://doi.org/10.1111/j.2041-210X.2010.00020.x (2010).
Penny, D. The comparative method in evolutionary biology. J. Classif. 9, 169–172. https://doi.org/10.1007/BF02618482 (1992).
Earl, C., Belitz, M. et al. Spatial phylogenetics of butterflies in relation to environmental drivers and angiosperm diversity across North America. BioRxiv:2020.2007.2022.216119, https://doi.org/10.1101/2020.07.22.216119 (2020).
PRISM. Climate Group, Parameter-elevation Regressions on Independent Slopes Model. Oregon State University, http://prism.oregonstate.edu. Accessed 24 July 2018.
Daly, C. et al. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 28, 2031–2064. https://doi.org/10.1002/joc.1688 (2008).
Peñuelas, J. et al. Response of plant species richness and primary productivity in shrublands along a north-south gradient in Europe to seven years of experimental warming and drought: Reductions in primary productivity in the heat and drought year of 2003. Glob. Change Biol. 13, 2563–2581. https://doi.org/10.1111/j.1365-2486.2007.01464.x (2007).
McMaster, G. S. & Wilhelm, W. W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 87, 291–300. https://doi.org/10.1016/S0168-1923(97)00027-0 (1997).
Walters, E. J., Morrell, C. H. & Auer, R. E. An investigation of the median-median method of linear regression. J. Stat. Educ. https://doi.org/10.1080/10691898.2006.11910582 (2006).
Theil, H. in Henri Theil’s Contributions to Economics and Econometrics: Econometric Theory and Methodology (eds Baldev Raj & Johan Koerts) 345–381 (Springer Netherlands, 1992).
Sen, P. K. Estimates of the regression coefficient based on Kendall’s Tau. J. Am. Stat. Assoc. 63, 1379–1389. https://doi.org/10.2307/2285891 (1968).
Siegel, A. F. Robust regression using repeated medians. Biometrika 69, 242–244. https://doi.org/10.2307/2335877 (1982).
Schneider, G., Chicken, E. & Becvarik, R. NSM3: Functions and Datasets to Accompany Hollander, Wolfe, and Chicken - Nonparametric Statistical Methods, Third Edition. R Package Version 1.15. https://CRAN.R-project.org/package=NSM3. (2020).
Patrick Bogaart, Loo, M. v. d. & Pannekoek, J. rtrim: Trends and Indices for Monitoring Data. R Package Version 2.1.1. https://CRAN.R-project.org/package=rtrim. (2020).
Zografou, K. et al. Stable generalist species anchor a dynamic pollination network. Ecosphere 11, e03225. https://doi.org/10.1002/ecs2.3225 (2020).
Pinheiro J, Bates D, DebRoy S & D, S. nlme: Linear and Nonlinear Mixed Effects Models. R Package v. 3.1‐117. (www document). https://CRAN.R-project.org/package=nlme. (2015).
Felsenstein, J. Phylogenies and quantitative characters. Annu. Rev. Ecol. Syst. 19, 445–471. https://doi.org/10.1146/annurev.es.19.110188.002305 (1988).
Acknowledgements
We thank the many biologists and volunteers who contributed to data collection and to FTIG-NGTC for managing S. i. idalia and its habitat and for supporting the long-term monitoring program for this species. We would also like to thank two anonymous reviewers for their many insightful comments and Chandra Earl for sharing phylogeny data. This research was supported by funding from the Pennsylvania Department of Military and Veterans Affairs (4300420055 and 4300599299) to Brent J. Sewall.
Author information
Authors and Affiliations
Contributions
K.Z., G.C.A., B.J.S. analyzed the data and wrote the paper; M.T.S. supervised and performed the field work; M.T.S., V.P.T., E.N.M. performed the field work; B.J.S.: designed the project and gained financial support. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zografou, K., Swartz, M.T., Adamidis, G.C. et al. Species traits affect phenological responses to climate change in a butterfly community. Sci Rep 11, 3283 (2021). https://doi.org/10.1038/s41598-021-82723-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-82723-1
- Springer Nature Limited
This article is cited by
-
The composition and phenology of butterflies are determined by their functional trait in Indian tropical dry forests
Biodiversity and Conservation (2024)
-
Weather anomalies more important than climate means in driving insect phenology
Communications Biology (2023)
-
Negative density-dependence buffers against mismatch-induced population decline in the Sinai baton blue butterfly
Oecologia (2023)
-
Differences in phenology, daily timing of activity, and associations of temperature utilization with survival in three threatened butterflies
Scientific Reports (2022)
-
Consistent trait-temperature interactions drive butterfly phenology in both incidental and survey data
Scientific Reports (2022)