
Abstract
The larval stages of Carmenta theobromae Busck (1910) and Simplicivalva ampliophilobia Davis, Gentili-Poole and Mitter (2008) attack the subcortical zone and pith in guava trees, respectively, in the first productive nucleus of fruit trees in Colombia: Hoya del Río Suárez (HRS). The presence of pest insects has been reported in 98% of the farms sampled in HRS (n = 124), with up to 96 and 11 simultaneous larvae per tree, respectively. Although the aspects of the basic biology and life cycle of both pests have been resolved, there are no strategies for managing populations in the field. Therefore, the aim of this study was to evaluate different management alternatives under laboratory and field conditions in HRS. In laboratory conditions, a completely randomized design was used in two separate experiments, each with six treatments: T1: Spinosad (a mixture of Spinosad A and D); T2: S-1,2-di(ethoxycarbonyl) ethyl 0,0-dimethylphosphorodithioate (chemical control); T3: Lecanicillium lecanii; T4: Beauveria bassiana; T5: Mix of B. bassiana and B. brongniartii, and T6: distilled water (control). The number of dead larvae per replicate per treatment was evaluated (DL), with experimental units of five and three larvae, respectively. In the field, to the two best alternatives found for each pest in the laboratory, pruning and keeping the area around the plants free of weeds were added as cultural management, in two separate additional experiments, each with three larvae as experimental unit per treatment. For C. theobromae, the best laboratory alternatives were chemical control (DL: 3.78) and L. lecanii (DL: 2.33), followed without statistical differences by B. bassiana (DL: 1.67). In the field, the virulence of B. bassiana improved (DL: 3), and together with pruning and keeping the area around the plants clear of weeds (DL: 3), they stood out as the best alternatives. For S. ampliophilobia under laboratory conditions, the best alternatives were Spinosad (2.74) and chemical control (DL: 2.66), without significant difference. In the field, there were no statistical differences between the alternatives, except for the control. This statistical parity of cultural practices, and biological and chemical management is an argument in favor of the use of the former to the detriment of the third, especially when the harmful effects of the molecule S-1,2 di (ethoxycarbonyl) ethyl 0, 0-dimethyl phosphorodithioate have been proven in air, water and agricultural soils, in addition to its association with thyroid cancer in humans. This is a strong argument to favor the use of synergies of cultural and biological management methods framed in IPM, as opposed to the use of chemical agents whose harmful effects are strongly documented, and whose use is becoming increasingly prohibited.
Similar content being viewed by others
Introduction
The Sesiidae family (Boisduval, 1828) (Lepidoptera) is comprised of 151 genera and 1,370 species, including 50 subspecies1. Their larvae are of agricultural interest because they are xylophagous insects that bore live wood (stems, branches, and roots) from shrubby, arboreal, and herbaceous hosts2. Most research on this family focuses on the biological aspects of the << dogwood borer >>, larvae of the Synanthedon genus found in multiple plant hosts, especially timber species from the northern and southern temperate zones3,4. However, towards the tropical zone in recent decades, the presence of the genus Carmenta, with the species C. foraseminis and C. theobromae5, stands out. The larval stages of C. theobromae have been reported attacking mainly cacao fruits5,6, as well as the subcortical zone of guava trees6,7, and myrtle6 in Venezuela and Colombia.
The attack of C. theobromae is considered a limitation in the two largest guava production centers in Colombia: Hoya del Río Suárez (HRS) and the northern region of Valle del Cauca (NVC)6. The presence of the pest insect has been reported in 98% of the farms sampled in HRS (n = 124), with approximately 10 infested trees over 40 observed, and up to 96 simultaneous larvae per tree. In the NVC its presence has been registered in the north and center of the department, in technified systems of pear guava Palmira ICA-1 and wild plots of common guava, as well as in cacao plantations6.
Likewise, in the same hosts, a recently described species has been found, identified as Simplicivalva ampliophilobia Davis, Gentili-Poole and Mitter (2008)8. The larval stage of this species can reach 5 cm in length and has been observed drilling the stem pith and primary and secondary branches of guava trees in HRS. Reports indicate that it can induce significant damage to the attacked trees causing their death and has been considered to be more aggressive than C. theobromae and other common guava pests, with an average incidence of 96%, and a severe attack degree with up to 11 larvae per tree.
Even though the basic biology and life cycle aspects of both the guava bander C. theobromae and the guava borer S. ampliophilobia have already been studied7,9, as well as the natural enemies present in the guava nuclei of HRS and NVC, and some monitoring methods including a pheromone for the bander5,10,11, so far there are no reports on the evaluation of integrated management strategies for these two pests.
Integrated pest management (IPM) has been considered as a promising solution to problems caused by insects within a perspective of sustainable agriculture12. In Colombia, there are reports of successful cases of applying IPM for crop limiting insects in cassava13 and Cape gooseberry14. However, the adoption of IPM in Latin America is, in general, marginal12. One reason is the few positive results in field validations, especially when live biological management agents are involved. In this sense, it is necessary to obtain laboratory results that allow predicting the behavior of the management alternatives in their application under field conditions. With this, it is possible to focus efforts on the most promising alternatives, saving resources in their validation15. From this perspective, the aim of this study is to evaluate different management alternatives for C. theobromae and S. ampliaphilobia, both in the laboratory and in commercial guava fields in HRS, to identify the best alternatives. This seeks to provide a mediated response to a problem with relevance for the development of guava cultivation in Colombia.
Materials and methods
Specimens were collected in silvo-pastoral guava plots located in HRS in the municipalities of Vélez, Jesús María, Guavatá, Puente Nacional, San Benito, Chipatá, Güepsa and Barbosa, in the department of Santander, and in the municipalities of Moniquirá, Provincia de Ricaurte, and Briceño and Tununguá in the western region of the department of Boyacá, Colombia. Silvo-pastoral and non-technified plots were chosen because they have high percentages of infestation of pest insects.
Laboratory studies were carried out at the agricultural entomology laboratory of the research center C.I. Tibaitatá of Agrosavia, Cimpa campus, located between 5°56′51″ N and 73°36′24″ W. The experimental work in the field was carried out in Vélez, department of Santander, in the sector Los Guayabos, district of Aco and Peña Blanca, in the farm El Paraíso located between 5°58′27″ N and 73°39′45″ W, and above 1905 m above the sea level.
Larvae were collected between March 27, 2015, and May 04, 2016, to obtain individuals from both pests for laboratory and field tests. Larva collections from I to VI instars were carried out through direct captures at sites showing recent activity (Fig. 1a,c,d). Pupae were also collected to obtain newborn larvae in the laboratory. All the captures imply the destruction of the guava tree to achieve the extraction of the individuals of S. ampliophilobia (Fig. 1 b).

Source: Pulido Blanco (2020).
a Presence of S. ampliophilobia inside a guava tree due to observing a typical capped hole made by this insect. b S. amphoraphilobia larva inside a sectioned guava tree. c Presence of C. theobromae. d C. theobromae larva devouring the vascular cambium.
The biological material (larvae and pupae) was transported in chambers conditioned at 22 °C, and 70% relative humidity (RH), and entered the laboratory emulating field conditions: 25 ± 3 °C and 60 ± 10% RH, with a photoperiod of 0:24. All the larvae were disinfected in a laminar flow cabinet, washing with 0.5% v/v NaClO for 5 s, followed by two washes with distilled and deionized water for 30 s.
The disinfected larvae were measured with an entomological ruler under a stereomicroscope and weighed with a Sartorius CPA3235 balance (d = 0.001 g) (supplementary Tables 1, 2, 3 and 4), for their subsequent distribution among treatments. Some larvae had to be immobilized with cold following the protocol of16. The total collections were recorded in a laboratory log and reported to the Alexander Von Humboldt institute through the Collection Permit of the Colombian Ministry of Environment and Sustainable Development. The larvae were deposited as follows: C. theobromae in a meridic diet of cacao prepared based17,18, varying the main ingredient, the antibiotic, the fat source, and the water concentration (Table 1). On the other hand, S. ampliophilobia was given a guava oligidic diet.
For each pest, a completely randomized block design was used, in which the block factor was the size of the larvae, and therefore, the age19 (supplementary tables 5, 6, 7, and 8). The response unit was the death event for each larva per repetition per treatment (DL); and the experimental unit, as well as the number of repetitions, varied for each pest in the laboratory and field according to larvae availability. In total, four separate experiments were carried out: two for each pest, one in the laboratory and one in the field. The experimental units differed in size, as noted in the Table 2.
Management treatments were: T1: Spinosad (a mixture of Spinosad A and D); T2: S-1,2-di(ethoxycarbonyl) ethyl 0,0-dimethyl phosphorodithioate (chemical handling); T3: Lecanicillium lecanii (commercial strain (cp)); T4: Beauveria bassiana (cp); T5: Mix of B. bassiana and B. brongniartii (cp), and T6: distilled water (control). All treatments were sprayed to the drop formation point following the manufacturer's instructions20,21,22,23, using conventional atomizers, to a final volume of 250 cm3 (Table 3). Treatments T1 to T5 are commercial products readily available in the study areas. Every three days, observation and recording of dead individuals were made as a response variable.
Based on the laboratory results, the two best alternatives were prioritized in the field, added to the chemical management and cultural management practices, including formation pruning and keeping the area around the plants clear of weeds, as well as the control. The age of the collected larvae, the position of the trees where the treatments were established, and the edge effect were blocked.
The data were analyzed through ANOVAs with Tukey's mean comparison test at a significance of 5%, verifying the assumptions of the model with the IBM SPSS Statics software, version 19.
Results and discussion
Collections
In total, 2313 biological states of C. theobromae were collected, corresponding to 2152 larvae between I and VI instars (93%) and 161 pupae (7%), and 441 biological states of the guava borer (S. ampliophilobia), corresponding to 239 larvae between the I and VI instars (54.2%), 200 pupae (45.35%), and two adults (sporadic capture) (0.45%). However, for both pests, high mortality was evidenced in larvae from I to III instars, as expected for Lepidoptera with a type II survival curve (r strategists), where the first instars are the most vulnerable24 (supplementary Tables 1, 2, 3, and 4). There was a higher decrease in biological states of S. ampliophilobia compared to those observed for C. theobromae, due to the lack of overlap of biological states and a very long univoltine life cycle with only one generation per year.
High concentrations of individuals per tree delimited the focal nature of the pests, which could be due to the reduction of habitats suitable for their survival. Individuals of C. theobromae and S. ampliophilobia were not collected in western Boyacá, but unidentified Sesiidae and Buprestidae individuals were collected. This demonstrates that trophic niches, whenever available, will be occupied by species with trophic analogies25.
Laboratory management alternatives
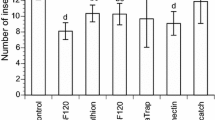
All model assumptions were achieved in the laboratory experiments. There are statistically significant differences between the management alternatives of C. theobromae and S. ampliophilobia under laboratory conditions after 37 and 42 days of evaluation of post-application dead larvae, respectively (n = 160; p value = 0.000; < α = 0.05, and n = 60; p value = 0.0001; α = 0.05, respectively) (supplementary Fig. 1 ).
For C. theobromae, the chemical treatment S-1,2-di(ethoxycarbonyl) ethyl 0,0-dimethyl phosphorodithioate registered the highest average dead larvae per replica (3.78), followed, with statistically significant differences, by the treatment with the L. lecanii fungus (2.33). The treatment of B. bassiana in a liquid vehicle (1.67) was statistically different from the chemical control, but not to the L. lecanii fungus (Fig. 2a). The Spinosad and mix of B. bassiana and B. brongniartii powder treatments did not have statistically significant differences with the control and were discarded for field evaluation. The evaluation of management alternatives for S. ampliophilobia showed an early response in terms of the number of dead larvae per experimental unit: between the second and sixth days, with a new larval death event between the tenth and sixteenth days. The death events after these periods were verified under laboratory conditions due to starvation because of inappetence. Therefore, the recommendation is to avoid an evaluation that extends beyond 20 days of follow-up. The best treatment was Spinosad (2.74), followed, without statistical differences by the chemical treatment (2.66). The mixture treatment of B. bassiana and B. brongniartii powder and the one with L. lecanii did not show statistically significant differences between them (Fig. 2b). The treatment of B. bassiana in a liquid vehicle yielded the least number of dead S. amphiophilobia larvae among the alternatives, regardless of the control, which behaved as expected, without causing death events.

Source: Elaborated by the authors.
Average number of dead larvae per replicate under laboratory conditions. Different letters indicate statistically significant differences. a C. theobromae and b S. ampliophilobia.
The chemical treatment S-1,2-di(ethoxycarbonyl) ethyl 0,0-dimethyl phosphorodithioate is widely used in Colombia, especially in HRS, for the effective and cheap control of the guava weevil Conotrachelus psidii Marshall (Coleoptera: Curculionidae), that is the main pest of the guava fruit tree, and a real limiting factor in bocadillo production (a popular snack in Colombia)26. Although the results on C. theobromae reinforce the thesis that the use of chemical synthesis insecticides persists due to their effectiveness27,28,29, toxic effects of S-1,2-di(ethoxycarbonyl) ethyl 0,0-dimethyl phosphorodithioate has been demonstrated by bioaccumulation in animal and plant species such as the common river carp Cyprinus carpio carpio30, cucumber31, and in native microorganism species in Colombian soils32. This molecule has been shown to pollute surface water, groundwater, soil, and even air persistently with few applications33. Its effects are not restricted to the organisms of the cropping field; its association with the increased risk of thyroid cancer in humans has been demonstrated34. Therefore, its use in fruit pest management programs has been restricted35,36. The results on S. ampliophilobia contribute to this trend, showing that an alternative such as Spinosad, a molecule of organic origin, has better experimental results than the chemical synthesis molecule most widely used in HRS.
The product based on the Spinosad molecule is registered by37 for the control of populations of Tephritidae in guava26, mango, and coffee in the HRS; furthermore, the molecule is globally recognized as a serious alternative to the control of this same family in a spectrum of tropical fruit trees with commercial interest38,39. Therefore, its use would not only contribute to the control of S. ampliophilobia populations but simultaneously to the management of fruit flies, especially the Anastrepha spp. complex and the Mediterranean fly Ceratitis capitata26.
Treatment with B. bassiana did not show the degree of effectiveness expected in the laboratory, which for Sesiidae related to C. theobromae has reached up to 76% of control, as in the case of Synanthedon myopaeformis in apple40,41. This may be due to the high specificity shown by some pathosystems, where the host exerts selection pressure on the genotypes of the pathogen, leading to the formation of host-specific forms42.
Field management alternatives
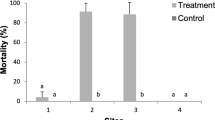
All model assumptions were achieved in the field experiments. There are statistically significant differences between the management alternatives of C. theobromae (Fig. 3a) and S. ampliophilobia (Fig. 3b) under field conditions after 19 and 22 days of evaluation of dead larvae after spray inoculation, respectively (n = 90; p value = 0.013; < α = 0.05, and n = 60; p value = 0.000; α = 0.05, respectively) (supplementary Fig. 2). However, the differences in S. ampliophilobia occur between the alternatives and the control.

Source: Elaborated by the authors.
Average number of dead larvae per replicate under field conditions. Different letters indicate statistically significant differences. a C. theobromae. b S. ampliophilobia.
For C. theobromae, B. bassiana in a liquid vehicle and the cultural treatment that included pruning and keeping the area around the plants clear of weeds, registered the highest average dead larvae per replica after aspersion (3), without having statistically significant differences with the chemical treatment (2.67) (Fig. 3a). This situation reflects that the use of entomopathogenic fungi, in combination with adverse conditions for the establishment of pests, is an alternative that competes with the effects of chemical synthesis pesticides43. Treatment with the fungus L. lecanii did not have statistically significant differences with water.
Modifying the habitat of pests through pruning is positioned as a successful practice included in good agricultural practices44,45 point out that in Venezuela, the combination of pruning with the release of biological controllers is a frequent practice in approximately 6% of the guava producers in the country, while46 demonstrated that the combination with entomopathogenic fungi had a synergistic effect that potentiated the lethality of those formulated against Premnotrypes vorax larvae in potato. However, the study of cultural methods associated with entomopathogenic fungi has focused more on the virulence of the strains used than on the discernment of the mechanisms that allow synergies. This treatment bases its actions on the affectation of the two main abiotic variables that insects require to live, especially in the egg and larva stages: temperature and humidity24. Pruning increases the temperature and reduces humidity in the branches and stems that are exposed to sunlight47, while pruning and keeping the area around the plants clear of weeds eliminates sheltered areas near the stem. This causes unfavorable conditions for the oviposition of insects, which depends on the choice of a host that meets the necessary conditions to ensure the larvae cycle48,49,50,51,52. Thus, this treatment continuously affects the establishment of a pest near its substrate.
Entomopathogenic fungi showed a dual behavior in their passage from laboratory to field: B. bassiana improved its effectiveness, while L. lecanii decreased it. The improvement of B. bassiana agrees with46,53 who observed efficiency of B. bassiana isolates in the control of Premnotrypes vorax, both in vitro and in field conditions. However, contrasts with the results reported by54. These authors recorded a decrease in the virulence of their own B. bassiana strains on Hypothenemus hampei selected in the laboratory and evaluated in the field. Although54 recognized that the interactions of the entomopathogen and the host with the environment are unknown, in terms of control, these can occur in a two-way process: both negative and positive55. Just as environmental factors can inhibit the pathogenicity of entomopathogenic fungi, as happened with the mix of B. bassiana and B. brongniartii on S. ampliophilobia, mechanisms not established in this study improved the pathogenicity of the fungal strain of the T4 treatment. In this regard56,57,58 warn that the effect of entomopathogenic fungi depends on the contact of the fungus with the host, the latter being one of the main determinants in the success of IPM methods.
In that sense, the loss of effectiveness L. lecanii reaffirm the observations of56,57,58, who warn that fungi are susceptible to solar radiation and depend on the available water to activate, maintain and end their biological cycle. We believe that entomopathogens were differentially affected by the conditions of high radiation and low rainfall during the period of strong summer caused by the phenomenon “El Niño” during June 2015 to April 201659. This duality in the field results of entomopathogenic fungi has been an argument against their use: there is less certainty of their effectiveness than, for example, chemical alternatives58.
The results for S. ampliophilobia, are revealing: suggest that between commercial biological strategies (spinosad), chemical (S-1,2 di (ethoxycarbonyl) ethyl 0,0-dimethyl phosphorodithioate) and cultural (pruning and silvering), there are no statistical differences that allow one method to be privileged over another, corroborating the growing trend towards a decrease in the gap between pesticides, biopesticides and cultural practices (Bettiol et al.29), which, in statistical terms of the effectiveness of S. ampliophilobia population management under HRS conditions, would have no contrast (Fig. 3b). Therefore, the choice of the management method would not depend on its effectiveness but on its cost, ease of acquisition, implementation and little harm to human and animal health and the environment. Under all the previous premises, cultural treatment is positioned as the best alternative.
Conclusions
Although the chemical treatment showed the highest efficiency in laboratory conditions against pests, especially against C. theobromae, it was found that the field conditions represented especially in the pruning and keeping the area around the plants clear of weeds as treatment, offer adverse conditions for the survival of pests, as much or even more efficient than biopesticides or chemical control. The foregoing shows that the constant application of pruning, sanitation, hygiene, and sanitary waste management practices in orchards is essential to control these two pests.
This statistical parity of cultural practices, and biological and chemical management is an argument in favor of the use of the former to the detriment of the third, especially when the harmful effects of the molecule S-1,2 di (ethoxycarbonyl) ethyl 0, 0-dimethyl phosphorodithioate have been proven in air, water and agricultural soils, in addition to its association with thyroid cancer in humans. This is a strong argument to favor the use of synergies of cultural and biological management methods framed in IPM, as opposed to the use of chemical agents whose harmful effects are strongly documented, and whose use is becoming increasingly prohibited.
References
Jin, Q., Wang, S. X. & Li, H. Catalogue of the family Sesiidae in China (Lepidoptera: Sesiidae). Shil. Rev. Lepidopterol. 36, 507–526 (2008).
Lacey, L. A. et al. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 132, 1–41 (2015).
Harms, K. E. & Aiello, A. Seed-boring by tropical clearwing moths (Sesiidae): Aberrant behavior or widespread habit?. J. Lepid. Soc. 49, 43–48 (1995).
Lee, C. M., Bae, Y. S. & Arita, Y. Morphological description of Synanthedon bicingulata (Staudinger, 1887) in life stages (Lepidoptera, Sesiidae). J. Asia. Pac. Entomol. 7, 177–185 (2004).
Delgado-Puchi, N. Caracterización morfológica de los Sesiidae (Insecta: Lepidoptera) perforadores del fruto del cacao (Theobroma cacao L.), presentes en la región costera del Estado Aragua. Venezuela. Entomotropica 20, 97–111 (2005).
Carabalí, A., Insuasty, O., Pulido, V. & Canacuán, D. Insectos plagas de importancia económica en el cultivo de la guayaba y sus estrategias de control. (2015).
Sarmiento-Naizaque, Z., Insuasty, O., Martínes, J. & Barreto-Triana, N. Aspectos biológicos del anillador de la guayaba Carmenta theobromae (Lepidoptera: Sesiidae) en Santander Colombia. Rev. Colomb. Entomol. 42, 176–183 (2016).
Davis, S. R., Gentili-Poole, P. & Mitter, C. A revision of the Cossulinae of Costa Rica and cladistic analysis of the world species (Lepidoptera: Cossidae). Zool. J. Linn. Soc. 154, 222–277 (2008).
Pulido Blanco, V. C., Insuasty Burbano, O. I., Sarmiento-Naizaque, Z. X. & Durán, J. R. Guava borer worm (Lepidoptera: Cossidae), a limiting pest in guava: biology, lifecycle and management alternatives. Heliyon 5, 1–29 (2019).
del Sánchez, M., Navarro, R., Marín, C., Casares, R. & Fuentes, V. Duración de la fase adulta y emergencia de machos y hembras del perforador del fruto de cacao en Choroní y Maracay, estado Aragua. Agron. Trop. 61, 241–251 (2011).
González, A., Altesor, P., Sellanes, C. & Rossini, C. Aplicación de Feromonas Sexuales en el Manjejo de Plaga de Cultivos Agrícolas. in Temas Selectos en Ecología Química de Insectos. (eds. Rojas., J. C. & Malo, E. A.) 343–360 (2012).
Alcázar, J. & Palacios, M. L. Enseñanza del Manejo Integrado de Plagas en el Cultivo de la Papa: La Experiencia del CIP en la Zona Andina del Perú. Rev. Latinoam. la Papa 9, 1–23 (2016).
Cuartas, P. E. et al. Novel biopesticide based on Erinnyis ello betabaculovirus: Characterization and preliminary field evaluation to control Erinnyis ello in rubber plantations. Pest Manag. Sci. 75, 1391–1399 (2019).
Chaves-Gómez, J. L., Chavez-Arias, C. C., Cotes Prado, A. M., Gómez-Caro, S. & Restrepo-Díaz, H. Physiological response of cape gooseberry seedlings to three biological control agents under fusarium oxysporum f. Sp. Physali Infection. Plant Dis. 104, 388–397 (2020).
Postali, J. & Coelho, A. Applied Biological Control in Brazil: From Laboratory Assays to Field Application. J. Insect Sci. 19, 5 (2019).
Lorea, L. Guía para la Captura y Conservación de Insectos. (2004).
Santos, A. M. et al. Nucleopoliedrovirus de Spodoptera frugiperda SfNPV003: Compatibilidad con agroquímicos y estabilidad en condiciones de almacenamiento. Corpoica Cienc. y Tecnol. Agropecu. 15, 219 (2014).
Greene, G. L., Leppla, N. C. & Dickerson, W. A. Velvetbean caterpillar: A rearing procedure and artificial medium123. J. Econ. Entomol. 69, 487–488 (1976).
Dyar, H. G. The number of molts of lepidopterous larvae. Psyche (Stuttg). 5, 420–421 (1890).
DowAgroScience. Success GF-120 0.02 CB. Etiqueta Web—Colombia (2017).
ADAMA. Malathion 57% (2018). https://www.adama.com/colombia/es/crop-protection/insecticide/malathion.html.
BIO-CROP. ADRAL W.P. Insecticida Biológico (2010). http://bio-crop.com/wp-content/uploads/2016/09/ADRAL-W.P-Ficha_Tecnica-BIO-CROP2.pdf.
SANOPLANT. Beauveriplant. Ficha técnica (2009). https://issuu.com/sanoplant/docs/ficha_tecnica_beauveriplant.
Zalucki, M. P., Clarke, A. R. & Malcolm, S. B. Ecology and behavior of first instar larval lepidoptera. Annu. Rev. Entomol. 47, 361–393 (2002).
Duarte Cueva, F. El control biológico como estrategia para apoyar las exportaciones agrícolas no tradicionales en Perú: Un análisis empírico. Contab. y Negocios 7, 81–100 (2012).
Insuasty, O., Monroy, R., Díaz, A. & & Bautista, J. Manejo Fitosanitario del cultivo de la Guayaba. Publicación del ICA (Imprenta nacional de Colombia, 2012).
Morales, M., Carlos, A. & Ibis, A. Efectividad De insecticidas químicos en el control de thrips palmi karny en el cultivo de la papa. Fitosanidad 6, 41–45 (2002).
Ruiz Sánchez, E. et al. Comparación de la efectividad de un insecticida botánico y dos químicos convencionales en el control del picudo (Anthonomus eugenii cano) (Coleoptera: Curculionidae) en chile habanero (Capsicum chinense Jacq.). Fitosanidad 13, 117–120 (2009).
Bettiol, Wagner, Rivera, M. C., Mondino, Pedro, ed. IV. Montealegre A., Jaime R., C. & C, Y. Control biológico de enfermedades de plantas en America latina y el caribe. Control biológico de enfermedades de plantas en América Latina y el Caribe (2014).
Yonar, S. M. Toxic effects of malathion in carp, Cyprinus carpio carpio: Protective role of lycopene. Ecotoxicol. Environ. Saf. 97, 223–229 (2013).
Lofty, H. M., Abd El-Aleem, A. E.-A. A. & Monir, H. H. Determination of insecticides malathion and lambda-cyhalothrin residues in zucchini by gas chromatography. Bull. Fac. Pharmacy, Cairo Univ. 51, 255–260 (2013).
Chaves-Bedoya, G., Ortíz-Moreno, M. L. & Ortiz-Rojas, L. Y. Efecto de la aplicación de agroquímicos en un cultivo de arroz sobre los microorganismos del suelo. Acta Agron. 62, 66–72 (2013).
Montenegro, R. Informe sobre los riesgos sanitarios y ambientales del malatión. (2001).
Lerro, C. C. et al. Organophosphate insecticide use and cancer incidence among spouses of pesticide applicators in the Agricultural Health Study. Occup. Environ. Med. 72, 736–744 (2015).
Martínez-Haro, M. et al. Relationship of the toxicity of pesticide formulations and their commercial restrictions with the frequency of animal poisonings. Ecotoxicol. Environ. Saf. 69, 396–402 (2008).
Osteen, C. D. & Fernandez-Cornejo, J. Economic and policy issues of U.S. agricultural pesticide use trends. Pest Manag. Sci. 69, 1001–1025 (2013).
Instituto Colombiano Agropecuario (ICA). Registros nacionales de plaguicidas químicos de uso agrícola. https://www.ica.gov.co/getdoc/d3612ebf-a5a6-4702-8d4b-8427c1cdaeb1/registros-nacionales-pqua-15-04-09.aspx (2018).
Pelz, K. S., Isaacs, R., Wise, J. C. & Gut, L. J. Protection of fruit against infestation by apple maggot and blueberry maggot (Diptera: Tephritidae) using compounds containing spinosad. J. Econ. Entomol. 98, 432–437 (2005).
Yee, W. L. & Alston, D. G. Effects of spinosad, spinosad bait, and chloronicotinyl insecticides on mortality and control of adult and larval western cherry fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 99, 1722–1732 (2006).
Cossentine, J. E., Judd, G. J. R., Bissett, J. D. & Lacey, L. A. Susceptibility of apple clearwing moth larvae, synanthedon myopaeformis (lepidoptera: Sesiidae) to beauveria bassiana and metarhizium brunneum. Biocontrol Sci. Technol. 20, 703–707 (2010).
Cossentine, J., Aurelian, V. M. & Judd, G. J. Biological Control Programmes in Canada 2001–2012. in Journal of Chemical Information and Modeling (eds. Mason, P. G. & GillespieD.R.) 285 (2013).
Motta-Delgado, P. A. & Murcia-Ordoñez, B. Hongos entomopatógenos como alternativa para el control biológico de plagas. Ambient. e Agua - An Interdiscip. J. Appl. Sci. 6, 77–90 (2011).
Faria, M. R. d. & Wraight, S. P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 43, 237–256 (2007).
Guzman, M. Control biológico y cultural de la sigatoka-negra. Trop. Plant Pathol. 37 (suplem), (2012).
Chirinos, D. T. & Geraud Pouey, F. El manejo de plagas agrícolas en Venezuela. Análisis y reflexiones sobre algunos casos. Interciencia 36, 192–199 (2011).
Villamil C., J. E., Martínez O., J. W. & Pinzón, E. H. Actividad biológica de hongos entomopatógenos sobre Premnotrypes vorax Hustache (Coleoptera: Curculionidae). Rev. Ciencias Agrícolas 33, 34 (2016).
López Juárez, S. A., Sol-Sánchez, Á., Córdova Ávalos, V. & Gallardo López, F. Efecto de la poda en plantaciones de cacao en el estado de Tabasco, México. Rev. Mex. Ciencias Agrícolas 2807 (2017). https://doi.org/10.29312/remexca.v0i14.452.
Lawton, J. H. Plant architecture and the diversity of phytophagous insects. Annu. Rev. Entomol. 28, 23–39. https://doi.org/10.1146/annurev.en.28.010183.000323 (1983).
Thompson, J. N. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects. Entomol. Exp. Appl. 47, 3–14 (1988).
Hanks, L. M., Paine, T. D., Millar, J. G. & Paine, D. International association for ecology host species preference and larval performance in the wood-boring beetle phoracantha. Oecologia 95, 22–29 (1993).
Hanks, L. M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 44, 483–505 (1999).
Suárez, L. H., Aguilar, A. V. & Gonzáles, W. L. Patrones de infestación por insectos xilófagos en renovales de Nothofagus obliqua Mirb. y Nothofagus dombeyi (Mirb.) Oerst. (Fagales: Nothofagaceae). Rev. Chil. Hist. Nat. 78, 23–32 (2005).
Villamil, J., Martínez, J. & Pinzón, E. Actividad entomopatogénica de aislamientos autóctonos de beauveria spp. sobre premnotrypes vorax (hustache) in vitro. Temas Agrar. 20, 19 (2015).
Cárdenas-Ramírez, Á. B., Villalba-Guott, D. A., Bustillo-Pardey, Á. E., Montoya-Restrep, E. C. & Góngora-Botero, C. Eficacia de mezclas de cepas del hongo beauveria bassiana en el control de la broca del café. Cenicafe 55, 136–149 (2004).
Tobar-h, S. P., Eugenia, P. & Montoya-restrepo, E. Evaluación en campo de un aislamiento de beauveria bassiana seleccionado por resistencia a la luz ultravioleta. Cenicafé 50, 195–204 (1999).
Asaff, A., Reyes, Y., López, E. & De la Torre, M. Guerra entre insectos y microorganismos: Una estrategia natural para el control de plagas. Av. y Perspect. 21, 291–295 (2002).
Lozano, M., Rodríguez, M., Vásquez, N. & Gutiérrez, G. Efecto de Metarhizium anisopliae sobre plagas rizófagas de arracacha (Arracacia xanthorriza) en Colombia. Manejo Integr. plagas 58–64 (2000).
García, M. A. G., García, S. C., Gordillo, J. M. L. & Martínez, R. F. M. Hongos entomopatógenos como una alternativa en el control biológico. Kuxulkab’ 15, (2015).
IDEAM. Impacto del fenómeno “El Niño” 2015–2016 en los nevados y alta montaña en Colombia. Subdirección de Ecosistemas e Información Ambiental Grupo de suelos y tierras. 2016. http://www.ideam.gov.co/documents/11769/132669/Impacto+de+El+Niño+en+la+alta+montaña+colombiana.pdf/dd41d158-0944-41d5-917e-44fdb524e8ea (2016).
Acknowledgements
This article is derived from the results and information obtained in the development of the project “Alternatives for the management of the bander (Lepidoptera: Sesiidae) and borer worms (Lepidoptera: Cossidae) identified in Hoya del Río Suarez, and the coiler (Lepidoptera: Sesiidae) identified in Valle del Cauca in guava (Psidium guajava L.).” Corporación Colombiana de Investigación Agropecuaria – AGROSAVIA executed this project within the framework of the Corporate Dynamic Agenda, financed by Ministerio de Agricultura y Desarrollo Rural (MADR).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pulido-Blanco, V.C., Pinzón-Sandoval, E.H., González-Chavarro, C.F. et al. Management alternatives for Carmenta theobromae (Busck, 1910) (Lepidoptera: Sesiidae) and Simplicivalva ampliophilobia (Lepidoptera: Cossidae), limiting pests of guava in Colombia. Sci Rep 11, 3076 (2021). https://doi.org/10.1038/s41598-021-81830-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-81830-3
- Springer Nature Limited