
Abstract
Recessive variants of the SLC26A4 gene are an important cause of hereditary hearing impairment. Several transgenic mice with different Slc26a4 variants have been generated. However, none have recapitulated the auditory phenotypes in humans. Of the SLC26A4 variants identified thus far, the p.T721M variant is of interest, as it appears to confer a more severe pathogenicity than most of the other missense variants, but milder pathogenicity than non-sense and frameshift variants. Using a genotype-driven approach, we established a knock-in mouse model homozygous for p.T721M. To verify the pathogenicity of p.T721M, we generated mice with compound heterozygous variants by intercrossing Slc26a4+/T721M mice with Slc26a4919-2A>G/919-2A>G mice, which segregated the c.919-2A > G variant with abolished Slc26a4 function. We then performed serial audiological assessments, vestibular evaluations, and inner ear morphological studies. Surprisingly, both Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M showed normal audiovestibular functions and inner ear morphology, indicating that p.T721M is non-pathogenic in mice and a single p.T721M allele is sufficient to maintain normal inner ear physiology. The evidence together with previous reports on mouse models with Slc26a4 p.C565Y and p.H723R variants, support our speculation that the absence of audiovestibular phenotypes in these mouse models could be attributed to different protein structures at the C-terminus of human and mouse pendrin.
Similar content being viewed by others

Introduction
Recessive variants in the SLC26A4 (PDS, GeneID: 5172) gene are a common cause of hereditary hearing impairment (HHI)1. In certain populations, SLC26A4 variants can be identified in approximately 15% to 20% of patients with HHI2. SLC26A4 encodes pendrin, a chloride/iodide/bicarbonate transporter expressed in the thyroid, inner ears, kidneys, lungs, liver, and heart3,4. Recessive SLC26A4 variants lead to Pendred syndrome (PS; MIM #274,600) and non-syndromic DFNB4 (MIM# 600,791). DFNB4 is characterized by isolated sensorineural hearing impairment (SNHI), which is associated with a common inner ear malformation called enlarged vestibular aqueduct (EVA; MIM 603,545), whereas patients with PS have goiter in addition to EVA5. To date, approximately 700 SLC26A4 variants have been identified (https://databases.lovd.nl/shared/genes/SLC26A4). Clinically, patients with SLC26A4 variants, either with DFNB4 or PS, usually suffer from progressive or fluctuating SNHI6.
The pathogenesis of SNHI in patients with DFNB4 and PS has been partially elucidated in various mouse models. Several mouse models have been generated, including knock-out Slc26a4-/- mice7, Slc26a4loop/loop mice with the p.S408F variant8, Slc26a4919-2A>G9/919-2A>G mice with the c.919–2 A > G variant9, Slc26a4H723R/H723R mice with the p.H723R variant10, Slc26a4L236P/L236P mice with the p.L236P variant11, Slc26a4C565Y/C565Y mice with p.C565Y variant12, conditional knock-out Tg[E]; Tg[R]; Slc26a4Δ/Δ mice13, and humanized hH723R Tg mice with the p.H723R variant in the human SLC26A4 sequence14. None of these models recapitulated SNHI phenotypes in humans. Knock-out Slc26a4-/-, Slc26a4loop/loop, and Slc26a4919-2A>G9/919-2A>G mice showed congenital profound SNHI that was too severe compared to their human counterparts7,8,9. Slc26a4H723R/H723R and Slc26a4C565Y/C565Y mice showed normal hearing without any hearing loss phenotypes10,12. Although Tg[E]; Tg[R]; Slc26a4Δ/Δ mice demonstrated hearing loss of various severities, doxycycline was required to induce the phenotype13. The auditory phenotypes of Slc26a4L236P/L236P and hH723R Tg mice were milder than those of Slc26a4-/-, Slc26a4loop/loop, and Slc26a4919-2A>G9/919-2A>G mice11,14, yet the absence of progressive hearing loss could not reflect the clinical symptoms in patients with SLC26A4 mutations.
Parallel to the transgenic mouse models, studies on cell lines have provided crucial insights into the pathogenicity of SLC26A4 variants. It has been reported that different SLC26A4 variants may result in different degrees of protein misexpression and/or dysfunction15,16,17. Among the SLC26A4 variants, whose pathogenicity has been investigated in cell lines, the p.T721M variant is especially interesting. The expression of pendrin in cell lines with p.T721M could not be rescued after salicylate treatment, but that in cell lines with p.H723R could, indicating that p.T721M might confer a stronger pathogenicity than p.H723R15. Based on these results, we hypothesized that mice with the Slc26a4 p.T721M variant might exhibit auditory phenotypes milder than those of c.919-2A > G mice but more severe than those of p.H723R mice. In this study, we generated a knock-in mouse model with the p.T721M variant, as well as compound heterozygous (Slc26a4919-2A>G/T721M) mice in which we tried to manipulate the severity of phenotypes by abolishing the other functional Slc26a4 allele9. We then characterized the audiovestibular phenotypes and inner ear pathology in these mouse models (Fig. 1).

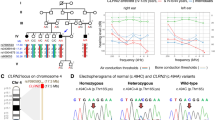
Generation of mice with Slc26a4 p.H721M variant using CRISPR/Cas9. (A) Design diagram. SgRNA for CRISPR/Cas9 and silent mutations for enzyme cutting sites as a check were designed to incorporate the c.2162C > T (p.H721M) variant into the genome of C57BL/6 mice. To generate the Slc26a4 p.T721M variant, the codon “ACG” was mutated to “ATG”. (B) Sanger sequencing was performed to confirm the nucleotide change in transgenic mice. The sequence was read in reverse.
Results
Auditory phenotypes
Wild-type mice (Slc26a4+/+), heterozygous mice (Slc26a4+/T721M), and homozygous mice (Slc26a4T721M/T721M) (n = 10 each) were subjected to audiological evaluations at 12, 28, and 44 weeks (Fig. 2). Both Slc26a4+/T721M and Slc26a4T721M/T721M mice had normal hearing up to 44 weeks, indicating that the p.T721M allele did not cause deafness in mice.

Auditory phenotypes. (A) The ABR waveforms in clicks did not differ significantly among heterozygous Slc26a4+/T721M, homozygous Slc26a4T721M/T721M, compound heterozygous Slc26a4919-2A>G/T721M, and wild-type Slc26a4+/+ mice. The ABR waveforms of Slc26a4919-2A>G/919-2A>G mice with profound hearing loss were also determined for comparison. (B) Heterozygous, homozygous, and compound heterozygous mice showed normal hearing thresholds across different frequencies, similar to those in wild-type Slc26a4+/+ mice. There were no significant differences among these four groups of mice at 12, 28, and 44 weeks.
To confirm the pathogenicity of the p.T721M allele in mice, we invalidated the Slc26a4 allele by intercrossing Slc26a4+/T721M mice with Slc26a4919-2A>G/919-2A>G mice to generate compound heterozygous mice (i.e., Slc26a4919-2A>G/T721M)9. Similar to heterozygous mice with the c.919-2A > G variant (i.e., Slc26a4+/919–2-2A>G), Slc26a4919-2A>G/T721M mice (n = 10) had normal hearing up to 44 weeks. Furthermore, physical collisions such as falling from rotarods did not induce hearing loss in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice (data not shown). These findings indicate that the p.T721M allele is not pathogenic and a single allele with p.T721M is sufficient to maintain the auditory function in mice with the p.T721M variant.
Inner ear morphology and pendrin expression
Cochlear morphology was investigated in homozygous (Slc26a4T721M/T721M) and compound heterozygous mice (Slc26a4919-2A>G/T721M). Cochlear morphologies of wild-type mice and profoundly deaf Slc26a4919-2A>G/919-2A>G mice were also obtained for comparison. The endolymphatic sac was severely enlarged only in Slc26a4919-2A>G/919-2A>G mice, and remained normal in size in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice (Fig. 3A). Similarly, abnormal morphological findings in Slc26a4919-2A>G/919-2A>G mice, including dilatation of the scala media, atrophy of the stria vascularis, and degeneration of the cochlear hair cells, were not observed in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice (Fig. 3B–D). Quantitative analyses of the endolymphatic space revealed that the cross-sectional area of the scala media in Slc26a4919-2A>G/919-2A>G mice was significantly larger than those in wild-type, Slc26a4T721M/T721M, and Slc26a4919-2A>G/T721M mice (271.6 ± 2.2; 59.6 ± 2.0; 56.5 ± 3.1; 58.9 ± 2.2 *1000 μm2, respectively, n = 3 each) (Fig. 3E).

Cochlea histology. (A) Gross morphology of the vestibular aqueduct and endolymphatic sac. The vestibular aqueduct and endolymphatic sac were enlarged in Slc26a4919-2A>G/919-2A>G mice. The length of the green lines represents the width of vestibular aqueduct. The red dash lines denote the contour of the sac. (B) Histology of the cochlea harvested from 3-month-old mice. On histological examination, the scala media was dilated in Slc26a4919-2A>G/919-2A>G mice, but not in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice (RM, Reissner's membrane; SV, stria vascularis; bar = 150 μm). (C) In the view of the organ of Corti panel, degeneration of hair cells was observed in Slc26a4919-2A>G/919-2A>G mice, but not in the other three groups of mice (IHC, inner hair cells; OHC 1–3, three rows of outer hair cells; bar = 150 μm). (D) Histology of the cochlear hair cells harvested from 3-month-old mice. Myosin VIIA expression was normal in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice, but was diminished in Slc26a4919-2A>G/919-2A>G mice (red: Myosin VIIA, blue: DAPI; bar = 50 μm). (E) Quantitative analysis of endolymphatic space. The cross-sectional area of the scala media (middle turn) in Slc26a4919-2A>G/919-2A>G mice was significantly larger than that in the other three groups. (F) The histology and expression of pendrin in the stria vascularis. Significant atrophy of stria vascularis and poor protein expression of pendrin were observed in Slc26a4919-2A>G/919-2A>G mice. In contrast, pendrin was normally distributed in the spiral prominence (SP) and root cells (RC) in Slc26a4CT721M/T721M and Slc26a4919-2A>G/T721M mice, similar to that in wild-type mice. Tissues were harvested from 3-month-old mice (green: pendrin, white dash line: the contour of stria vascularis; bar = 50 μm).
We then examined the expression of pendrin in the cochlea of Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice by immunolocalization (Fig. 3F). In both strains, pendrin was normally distributed in the spiral prominence and root cells, indicating that the expression of pendrin was normal in the p.T721M mice. In addition, the contour of the stria vascularis (indicated by white dashed lines in the figure) showed atrophic changes in Slc26a4919-2A>G/919-2A>G mice, but not in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice.
Vestibular function evaluation
Vestibular morphology was also investigated in homozygous mice (Slc26a4T721M/T721M) and compound heterozygous mice (Slc26a4919-2A>G/T721M) (Fig. 4A). Both strains of mice showed normal vestibule morphologies. In contrast, giant otoliths were observed in Slc26a4919-2A>G/919-2A>G mice. Fluorescence confocal microscopy demonstrated that vestibular hair cells in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice did not degenerate (Fig. 4B).

Morphology, histology, and phenotypes of the vestibule system. (A) Histology of the vestibular organs. Giant otoconia was observed in Slc26a4919-2A>G/919-2A>G mice, but the otoconia was normal in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice (bar = 150 μm). (B) Fluorescence confocal microscopy observations. In contrast to the Slc26a4919-2A>G/919-2A>G mice, vestibular hair cells in Slc26a4T721M/T721M and Slc26a4919-2A>G/T721M mice were not degenerated (green: Myosin VIIA; bar = 50 μm). (C–D) In contrast to the Slc26a4919-2A>G/919-2A>G mice, the homozygous Slc26a4T721M/T721M and compound heterozygous Slc26a4919-2A>G/T721M mice performed well in rotarod test (C) and swimming test (D), similar to wild-type Slc26a4+/+ mice.
Fifteen mice of each group including Slc26a4+/+, Slc26a4+/T721M, Slc26a4T721M/T721M, and Slc26a4919-2A>G/T721M mice, were subjected to vestibular evaluations. Similar to the normal audiological phenotypes, neither heterozygous mice (i.e., Slc26a4+/T721M and Slc26a4919-2A>G/T721M) nor homozygous mice (i.e., Slc26a4T721M/T721M) showed vestibular deficits such as head tilting and circling behavior, and both groups performed normally on the rotarod (Fig. 4C) and in swimming tests (Fig. 4D). These findings indicate that a single p.T721M allele is sufficient to maintain normal vestibular function in mice.
Discussion
The knock-in mouse generated in this study, the Slc26a4T721M/T721M with the Slc26a4 p.T721M variant, demonstrated normal audiovestibular phenotypes and inner ear morphologies, similar to wild-type mice. To investigate whether the p.T721M variant could contribute to SNHI in mice through the haplo-insufficiency mode, we further generated mice with compound heterozygous variants (Slc26a4919-2A>G/T721M) by intercrossing Slc26a4+/T721M mice with Slc26a4919-2A>G/919-2A>G mice, which segregated the c.919-2A > G variant with abolished function. Compound heterozygous mice for p.T721M and c.919-2A > G (i.e., Slc26a4919-2A>G/T721M mice) also had normal audiovestibular phenotypes, indicating that a single p.T721M allele was sufficient to maintain normal inner ear physiology in the mice.
The SLC26A4 p.T721M variant has been documented sporadically in hearing-impaired families worldwide, including two Mediterranean families18, two Iranian families19, two Japanese family20,21, three Chinese families22,23, and one Taiwanese family24. Although this variant is widely distributed across different populations, its prevalence is relatively low compared to other SLC26A4 variants, such as c.919-2A > G or p.H723R. According to the American College of Medical Genetics and Genomics (ACMG) guidelines in the Varsome platform25, SLC26A4 p.T721M is classified as “pathogenic” by fulfilling the criteria of PM1, PM2, PP2, PP3, and PP5. It is located in the hotspot region of the SLC26A4 gene, where the majority of pathogenic variants occur (PM1). Its allele frequency is < 0.0001 across various ethnic groups in gnomAD (PM2). The majority of non-VUS missense variants in SLC26A4 have been reported as “pathogenic” in UniProt (PP2). In well-established databases (e.g., ClinVar, DVD26, and UniProt) and in several prediction algorithms (e.g., SIFT, Polyphen2, LRT, FATHMM, Mutation Taster, etc.), p.T721M was categorized as “pathogenic” (PP3 and PP5). Clinically, both homozygosity for p.T721M and compound heterozygosity for p.T721M with another SLC26A4 mutation have been linked to non-syndromic DFNB4 or PS, as characterized by EVA, progressive or fluctuating severe-to-profound SNHI, and/or goiter18,19,20,21. Specifically, the hearing features have been detailed in two compound heterozygotes, one with symmetric SNHI (right/left: 103.75/110 dBHL)21, and the other with asymmetric SNHI (right/left: 112.5/68.75 dBHL)20. These typical clinical manifestations in p.T721M homozygotes and compound heterozygotes suggest that p.T721M is a pathogenic SLC26A4 variant in humans.
In previous cell line studies, a number of SLC26A4 variants, including p.P123S, p.M147V, p.L236P, p.S657N, p.T721M, and p.H723R, have been demonstrated to confer pathogenicity by affecting the trafficking process, rather than the expression level of pendrin15,17. However, the affected trafficking process can be rescued by certain treatments15,17. Low-temperature incubation has been reported to rescue the trafficking of pendrin with p.H723R, which originally accumulated in the endoplasmic reticulum, but not the trafficking of pendrin with p.L236P, which originally accumulated in centrosomal regions17. Similarly, salicylate treatment could rescue the trafficking of pendrin with p.P123S, p.M147V, p.S657N, and p.H723R, and restore the function of pendrin as an anion exchanger, but not that of pendrin with p.T721M15. These lines of evidence also support the view that the SLC26A4 p.T721M variant is pathogenic and implies that p.T721M is more pathogenic compared to other missense SLC26A4 variants, such as p.H723R.
To our surprise, the pathogenicity of p.T721M as predicted by the ACMG guidelines and evidenced by the cell line studies was not observed in our mouse model with the Slc26a4 p.T721M variant. These findings are consistent with our previous studies in mouse models with Slc26a4 p.H723R10 and p.C565Y12 variants. In the cochlea of mice, pendrin is expressed at the spiral prominence and outer sulcus cells27, which is almost the same as its expression in the cochlea of primates28. Therefore, the position of pendrin expression does not appear to be a major factor contributing to the phenotypic discrepancy between species.
Alternatively, the inter-species phenotypic discrepancy may be attributed to the structural differences between mouse and human pendrin.
To date, five mouse models with missense Slc26a4 variants, including p.L236P11, p.S408F8, p.C565Y12, p.T721M (this study), and p.H723R10, have been documented in the literature (Fig. 5A). Abnormal audiovestibular phenotypes were observed in mice with p.L236P and p.S408F, but not in mice with p.C565Y, p.T721M, and p.H723R. Notably, p.L236P and p.S408F are located in the transmembrane domains of pendrin27,28. In contrast, p.C565Y, p.T721M, and p.H723R are located in the C-terminus of pendrin comprised of amino acid residues 508–78027,28 (Fig. 5B). From an evolutionary perspective, the amino acid sequence of the pendrin C-terminus is less conserved, sharing only 86% identity between mice and humans. In contrast, the amino acid sequence of the transmembrane domains shared 92% identity between the two species (https://www.expasy.org/&https://www.uniprot.org/). Accordingly, we speculate that the absence of phenotypes in mice with Slc26a4 p.C565Y, p.T721M, and p.H723R variants could be attributed to different protein structures at the C-terminus of human and mouse pendrin.

Five documented murine missense Slc26a4 variants. Five mouse models with missense Slc26a4 variants, including p.L236P, p.S408F, p.C565Y, p.T721M, and p.H723R, have been documented in the literature. (A) Scheme of the genomic DNA. Squares represent exons and lines represent introns. (B) Scheme of the 12-transmembrane domain model of pendrin. The five missense variants are indicated with stars, with p.L236P indicated in green, p.S408F in purple, p.C565Y in orange, p.T721M in red, and p.H723R in blue. L236P and p.S408F are located in the transmembrane domains, whereas p.C565Y, p.T721M, and p.H723R are located at the C-terminus of pendrin.
Another line of evidence that supports our speculation is the hH723R Tg mouse model14. Instead of creating the variant on murine genomic DNA, the authors delivered a sequence of human cDNA harboring the p.H723R variant into pronuclear-stage mouse embryos. This humanized transgenic mouse model revealed profound SNHI10, suggesting that C-terminus SLC26A4 variants identified in humans might require a human peptide backbone to exhibit their pathogenicity.
Rapp et al. analyzed the locations of SLC26A4 mutations and identified a high density of SLC26A4 mutations in the anion-binding transmembrane domains (TMs), including TM1, 3, 8, and 1029. The findings indicate that when present in these transmembrane domains, SLC26A4 variants are more likely to be pathogenic. This could possibly explain the higher evolutionary conservation of amino acid sequences between human and mouse pendrin at the transmembrane domains, as well as the clear phenotypic manifestation of profound SNHI in mice with Slc26a4 variants at the transmembrane domains, such as Slc26a4L236P/L236P and Slc26a4loop/loop mice (i.e., mice with p.S408F).
In summary, using a genotype-driven approach, we generated a knock-in mouse model segregating the deafness-associated SLC26A4 p.T721M variant in humans. Surprisingly, mice with the Slc26a4 p.T721M variant exhibited a normal audiovestibular phenotype and inner ear morphology. Because there might be differences in the pathogenicity of specific SLC26A4 variants between humans and mice, caution is needed while extrapolating the results of animal studies to humans.
Methods
Ethics statement
All animal experiments were carried out in accordance with animal welfare guidelines and approved by the Institutional Animal Care and Use Committee of the National Taiwan University College of Medicine (approval no. 20160337). Also, all animal experiments followed the guidelines of ARRIVE 2.0.
Construction of Slc26a4 T721M/T721M knock-in mice
Transgenic mice were generated by the Transgenic Mouse Models Core (TMMC, Taiwan) using the CRISPR technology-associated RNA-guided endonuclease Cas9 to mutate the Slc26a4 gene and generate the Slc26a4+/T721M mouse line. Specific guiding RNAs (sgRNAs) were developed to target exon 15 of the Slc26a4 gene in C57BL/6 mice. The sgRNA and CRISPR/Cas9 RNA were delivered into the mouse zygote to generate founders. The two male founder mice that were obtained from each, harbored the p.T721M (c.2162C > T) variant in the Slc26a4 gene. After germline transmission of the targeted variant allele, we produced the congenic Slc26a4+/T721M mouse line used in this study by repeated backcrossing into the C57BL/6 inbred strain for 6–10 generations. Mice homozygous for the variant (Slc26a4T721M/T721M) were obtained by intercrossing heterozygous mice (i.e., Slc26a4+/T721M) (Fig. 1). Reverse transcription-PCR (RT-PCR) of mRNA of inner ear extract and direct sequencing confirmed a pure non-chimeric genetic background without unintentional wild-type Slc26a4 expression in Slc26a4T721M/T721M mice.
Auditory evaluations
Mice were anesthetized and placed in a head-holder within an acoustically and electrically insulated and grounded test room10. We used an evoked potential detection system (Smart EP 3.90; Intelligent Hearing Systems, Miami, FL, USA) to measure auditory brainstem response (ABR) thresholds in mice. Click sounds, as well as 8, 16, and 32 kHz tone bursts at various intensities were generated to evoke ABRs. Response signals were detected using subcutaneous needle electrodes. The active electrodes were inserted into the vertex and the ipsilateral retroauricular region with a ground electrode on the back of each mouse. The ABRs were recorded from postnatal 12–44 weeks, to trace changes in auditory function.
ABRs were measured bilaterally. As there were no significant differences between the two ears, only the data of the left ears were presented and analyzed.
Inner ear morphology studies
For light microscopy studies, tissues were stained with hematoxylin and eosin (H&E). The morphology of each sample was examined using a Leica optical microscope10. First, inner ear tissues from adult mice (P28-P30) were fixed by perilymphatic perfusion with 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) through round and oval windows, and a small fenestra in the apex of the cochlear bony capsule. The specimens were decalcified for one week. The samples were then dehydrated and embedded in paraffin. Subsequently, serial Sects. (7 mm) were stained with H&E. ImageJ software (NIH, http://imagej.nih.gov/ij/download.html) was used to quantify the cross-sectional area of the middle turn scala media.
Whole-mount studies of mouse inner ear specimens were performed as previously described10,30. The tissues were stained with rabbit anti-Myosin-VIIA primary antibody (1:200; Proteus Bioscience Inc., Ramona, CA, USA). Later, the tissue sections were incubated with 4',6-diamidino-2-phenylindole (DAPI; 1:5000; Thermo Fisher Scientific, Waltham, MA, USA) and Alexa Fluor 568-conjugated goat anti-rabbit IgG (H + L) secondary antibodies (1:200; Thermo Fisher Scientific) at 4 °C overnight. After washing in PBS, the tissues were mounted using the ProLong Antifade kit (Molecular Probes, Eugene, OR, USA) for 20 min at room temperature. Images of the tissues were obtained using a model LSM 800 laser scanning confocal microscope (Carl Zeiss, Oberkochen, Germany).
Expression of pendrin
Tissue sections were prepared from the inner ears of wild-type, Slc26a4919-2A>G/919-2A>G, and Slc26a4T721M/T721M mice. The sections were mounted on silane-coated glass slides, deparaffinized in xylene, and rehydrated in ethanol. Tissues were stained with a 1:1000 dilution of rabbit anti-pendrin antibody kindly provided by Dr. Jinsei Jung, Yonsei University College of Medicine, Seoul, Republic of Korea. This was followed by exposure to DAPI (1:5000) and Alexa Fluor 488-conjugated goat anti-rabbit IgG (H + L) secondary antibody (1:200; Thermo Fisher Scientific). After incubation, the slides were washed with PBS and mounted with ProLong Antifade kit at 25 °C. Images were obtained using the aforementioned LSM 880 laser scanning confocal microscope10.
Vestibular function evaluation
Mice were subjected to a series of tests at 8 weeks, including the swimming and rotarod tests. For the swimming test, the swimming performance of the mice was scored from 0 to 3, with 0 representing normal swimming and 3 representing underwater tumbling31. For the rotarod tests, the mice were placed on the rotating rod for a maximum of 180 s. The speed of the rods was accelerated from 5 rpm to a maximum speed of 20 rpm in one min. The length of time each mouse remained on the rotating rod was recorded32.
Statistical analyses
Data are presented as the mean ± standard deviation. Statistical analyses were conducted using an unpaired Student’s t-test with Bonferroni correction for continuous variables. Statistical significance was set at p < 0.05. All analyses were performed using SPSS software (version 15.0; SPSS Inc., Chicago, IL, USA).
Data availability
The authors confirm that the data supporting the findings of this study are available within the article.
References
Wolf, A. et al. A novel mutation in SLC26A4 causes nonsyndromic autosomal recessive hearing impairment. Otol. Neurotol. 38(2), 173–179 (2017).
Han, M. et al. A quantitative cSMART assay for noninvasive prenatal screening of autosomal recessive nonsyndromic hearing loss caused by GJB2 and SLC26A4 mutations. Genet. Med. 19(12), 1309 (2017).
Everett, L. A., Morsli, H., Wu, D. K. & Green, E. D. Expression pattern of the mouse ortholog of the Pendred’s syndrome gene (Pds) suggests a key role for pendrin in the inner ear. Proc. Natl. Acad. Sci. 96(17), 9727–9732 (1999).
Pelzl, L. et al. DOCA sensitive pendrin expression in kidney, heart, lung and thyroid tissues. Cell. Physiol. Biochem. 30(6), 1491–1501 (2012).
Li, X. C. et al. A mutation in PDS causes non-syndromic recessive deafness. Nat. Genet. 18(3), 215 (1998).
Mori, T., Westerberg, B. D., Atashband, S. & Kozak, F. K. Natural history of hearing loss in children with enlarged vestibular aqueduct syndrome. J. Otolaryngol. Head Neck Surgery. 37(1), 112–118 (2008).
Everett, L. A. et al. Targeted disruption of mouse Pds provides insight about the inner-ear defects encountered in Pendred syndrome. Hum. Mol. Genet. 10(2), 153–161 (2001).
Dror, A. A. et al. Calcium oxalate stone formation in the inner ear as a result of an Slc26a4 mutation. J. Biol. Chem. 285(28), 21724–21735 (2010).
Lu, Y.-C. et al. Establishment of a knock-in mouse model with the SLC26A4c. 919–2A> G mutation and characterization of its pathology. PLoS ONE 6(7), e22150 (2011).
Lu, Y.-C. et al. Differences in the pathogenicity of the p. H723R mutation of the common deafness-associated SLC26A4 gene in humans and mice. PLoS ONE 8(6), 64906 (2013).
Wen, Z. et al. A knock-in mouse model of Pendred syndrome with Slc26a4 L236P mutation. Biochem. Biophys. Res. Commun. 515(2), 359–365. https://doi.org/10.1016/j.bbrc.2019.05.157 (2019).
Hu, C.-J. et al. Toward the pathogenicity of the SLC26A4 p C565Y variant using a genetically driven mouse model. Int. J. Mol. Sci. 22(6), 2789 (2021).
Choi, B. Y. et al. Mouse model of enlarged vestibular aqueducts defines temporal requirement of Slc26a4 expression for hearing acquisition. J. Clin. Investig. 121(11), 4516–4525 (2011).
Choi, H. J. et al. DNAJC14 ameliorates inner ear degeneration in the DFNB4 mouse model. Mol. Therapy-Methods Clin. Develop. 17, 188–197 (2020).
Ishihara, K. et al. Salicylate restores transport function and anion exchanger activity of missense pendrin mutations. Hear. Res. 270(1–2), 110–118 (2010).
Choi, B. Y. et al. Hypo-functional SLC26A4 variants associated with nonsyndromic hearing loss and enlargement of the vestibular aqueduct: Genotype-phenotype correlation or coincidental polymorphisms?. Hum. Mutat. 30(4), 599–608 (2009).
Yoon, J. S. et al. Heterogeneity in the processing defect of SLC26A4 mutants. J. Med. Genet. 45(7), 411–419. https://doi.org/10.1136/jmg.2007.054635 (2008).
López-Bigas, N. et al. Identification of five new mutations of PDS/SLC26A4 in Mediterranean families with hearing impairment. Hum. Mutat. 18(6), 548 (2001).
Kahrizi, K. et al. Identification of SLC26A4 gene mutations in Iranian families with hereditary hearing impairment. Eur. J. Pediatr. 168(6), 651 (2009).
Miyagawa, M., Nishio, S. Y. & Usami, S. I. Mutation spectrum and genotype–phenotype correlation of hearing loss patients caused by SLC26A4 mutations in the Japanese: A large cohort study. J. Human Genet. 59(5), 262 (2014).
Okamoto, Y. et al. Subgroups of enlarged vestibular aqueduct in relation to SLC26A4 mutations and hearing loss. Laryngoscope 124(4), E134–E140 (2014).
Yuan, Y. et al. Molecular epidemiology and functional assessment of novel allelic variants of SLC26A4 in non-syndromic hearing loss patients with enlarged vestibular aqueduct in China. PLoS ONE 7(11), e49984 (2012).
Zhang, M., Han, Y., Zhang, F., Bai, X. & Wang, H. Mutation spectrum and hotspots of the common deafness genes in 314 patients with nonsyndromic hearing loss in Heze area China. Acta Oto-laryngol 139(7), 612–617 (2019).
Wu, C.-C. et al. Genetic epidemiology and clinical features of hereditary hearing impairment in the Taiwanese population. Genes 10(10), 772 (2019).
Kopanos, C. et al. VarSome: The human genomic variant search engine. Bioinformatics 35(11), 1978–1980 (2019).
Azaiez, H. et al. Genomic landscape and mutational signatures of deafness-associated genes. Am. J. Hum. Genet. 103(4), 484–497 (2018).
Bassot, C., Minervini, G., Leonardi, E. & Tosatto, S. C. Mapping pathogenic mutations suggests an innovative structural model for the pendrin (SLC26A4) transmembrane domain. Biochimie 132, 109–120 (2017).
Royaux, I. E. et al. Pendrin, the protein encoded by the Pendred syndrome gene (PDS), is an apical porter of iodide in the thyroid and is regulated by thyroglobulin in FRTL-5 cells. Endocrinol. 141(2), 839–845 (2000).
Rapp, C., Bai, X. & Reithmeier, R. A. Molecular analysis of human solute carrier SLC26 anion transporter disease-causing mutations using 3-dimensional homology modeling. Biochim. Biophys. Acta 1859(12), 2420–2434 (2017).
Cheng, Y. F. et al. Generation and pathological characterization of a transgenic mouse model carrying a missense PJVK mutation. Biochem. Biophys. Res. Commun. 532(4), 675–681. https://doi.org/10.1016/j.bbrc.2020.07.101 (2020).
Hardisty-Hughes, R. E., Parker, A. & Brown, S. D. A hearing and vestibular phenotyping pipeline to identify mouse mutants with hearing impairment. Nat. Protoc. 5(1), 177 (2010).
Isgrig, K. et al. Gene therapy restores balance and auditory functions in a mouse model of Usher syndrome. Mol. Ther. 25(3), 780–791 (2017).
Acknowledgements
We thank the imaging core at the First Core Labs, National Taiwan University College of Medicine, for the technical support in image acquisition and analysis. We also thank Ministry of Science and Technology (MOST109-2628-B-075-024), Taipei Veteran General Hospital-National Taiwan University Hospital Joint Research Program (VN110-10), National Health Research Institute (NHRI-EX110-10914PI), and National Taiwan University Hospital (110-S5093) for the grant support.
Author information
Authors and Affiliations
Contributions
C.J.Hsu., T.C.L., C.C.W. conceived and designed the study. I.S.Y. and S.W.L. generated the transgenic mouse. C.J.Hu., Y.C.L., Y.H.C. and Y.S.L. performed histological experiments and animal behavior analysis. C.Y.T. provided clinical data analysis. Y.H.C., Y.C.L., and C.J.Hu. performed the explants experiments analyzed the data. C.J.Hu., Y.C.L., Y.F.C., and C.C.W. drafted the manuscript. T.H.Y., C.C.W., C.J.Hsu., T.C.L., P.H.L., and Y.F.C. supervised the whole study and provided critical revision. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hu, CJ., Lu, YC., Tsai, CY. et al. Insights into phenotypic differences between humans and mice with p.T721M and other C-terminal variants of the SLC26A4 gene. Sci Rep 11, 20983 (2021). https://doi.org/10.1038/s41598-021-00448-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-00448-7
- Springer Nature Limited