
Abstract
Caloric restriction (CR) can attenuate the general loss of health observed during aging, being one of the mechanisms involved the reduction of hormonal alteration, such as insulin and leptin. This change could also prevent age-specific fluctuations in brain monoamines, although few studies have addressed the effects of CR on peripheral hormones and central neurotransmitters exhaustively. Therefore, the variations in brain monoamine levels and some peripheral hormones were assessed here in adult 4-month old and 24-month old male Wistar rats fed ad libitum (AL) or maintained on a 30% CR diet from four months of age. Noradrenaline (NA), dopamine (DA), serotonin (5-HT) and its metabolites were measured by high-performance liquid chromatography with electrochemical detection (HPLC-ED) in nine brain regions: cerebellum, pons, midbrain, hypothalamus, thalamus, hippocampus, striatum, frontal cortex, and occipital cortex. In addition, the blood plasma levels of hormones like corticosterone, insulin and leptin were also evaluated, as were insulin-like growth factor 1 and other basal metabolic parameters using enzyme-linked immunosorbent assays (ELISAs): cholesterol, glucose, triglycerides, albumin, low-density lipoprotein, calcium and high-density lipoprotein (HDLc). CR was seen to increase the NA levels that are altered by aging in specific brain regions like the striatum, thalamus, cerebellum and hypothalamus, and the DA levels in the striatum, as well as modifying the 5-HT levels in the striatum, hypothalamus, pons and hippocampus. Moreover, the insulin, leptin, calcium and HDLc levels in the blood were restored in old animals maintained on a CR diet. These results suggest that a dietary intervention like CR may have beneficial health effects, recovering some negative effects on peripheral hormones, metabolic parameters and brain monoamine concentrations.
Similar content being viewed by others

Introduction
The brain undergoes a series of physiological and functional alterations as it ages, which can produce greater susceptibility to neurodegenerative disorders. The changes in the aging brain are very heterogeneous, the most noticeable alterations evident in the hippocampus, the frontal cortex and the striatum1. Other regions affected by ageing are the thalamus, hypothalamus, cerebellum and brainstem2, whereas one of the least affected regions is the occipital cortex3. Age-related biochemical changes include altered levels of neurotransmitters and peripheral hormones, as well as their receptor interactions. Monoaminergic neurotransmitters like DA, NA and 5-HT have been shown to decline with age in some of the aforementioned regions. In rats, DA levels diminish in the hippocampus4,5,6,7, striatum5,7,8,9,10,11 and brainstem11, while significant age-induced declines in NA have been detected in the striatum and hippocampus6,7,9, the midbrain10,12 and the pons-medulla5. 5-HT is also reduced in the frontal cortex, hippocampus and striatum5,6,7,9,10,13. Nevertheless, ageing appears to produce increases in the 5-hydroxyindoleacetic acid/serotonin ratio (5-HIAA/5-HT) in the frontal cortex, amygdala and striatum, suggesting an age-related decrease in 5-HT synthesis is coupled to its enhanced metabolism5.
Age-related impairment of brain function is also associated with other physiological changes, including alterations of peripheral hormones like insulin14, leptin15 and insulin-likegrowth factor 1 (IGF-1)16, as well as increased levels of stress hormones like corticosterone17. Dysregulation of insulin and leptin affects neuronal inflammation and oxidative stress, as reflected by the changes in proinflammatory cytokine release18,19. In this context, it was proposed that immune system pathways may influence the biosynthesis of monoamines20 and indeed, age-related brain inflammation has been associated with abnormal tryptophan and tyrosine metabolism21, which may in turn be associated with decreases in the synthesis of monoamines during aging.
The rate and severity of the deleterious effects of normal aging can be attenuated by a wide variety of factors, such as appropriate diets. CR, defined as a reduction in the caloric intake without causing malnutrition22, ameliorates brain aging by reducing oxidative stress, improving mitochondrial function, activating anti-inflammatory responses, promoting neurogenesis and enhancing synaptic plasticity23,24. In addition, CR seems to modify leptin and insulin concentrations and metabolic parameters, which in turn may attenuate the age-related decreases in monoamines. Despite the relationship between CR, monoamines and peripheral hormones, no studies have yet combined and correlated these elements, analyzing their relationships in different brain areas.
In this context, the present study set out to evaluate the effect of life-long CR on monoamines in 24-month old Wistar rats, evaluating NA, DA and 5HT, as well as their metabolites L-DOPA, 3,4-dihydroxyphenylacetic acid (DOPAC), tyrosine hydroxylase (TH), homovanilic acid (HVA), 5-HIAA, tryptophan hydroxylase (TrpH), and the peripheral hormones insulin, leptin, corticosterone, IGF-1. Monoamines were analyzed in the brain regions most affected by aging, such as the prefrontal cortex, hippocampus, striatum and thalamus, as well as in the cerebellum, midbrain and pons, using the occipital cortex as a control region. To control the effects of CR on the animals’ general health, other biochemical parameters were also analyzed, such as their cholesterol, glucose, total protein, triglyceride, albumin, calcium, low-density lipoprotein (LDL-c), high-density lipoprotein (HDL-c) and alkaline phosphatase (ALP).
Results
The effects of CR on monoaminergic systems
Noradrenergic system
The concentration of neurotransmitters and their metabolites was assessed in each brain region (mean ± SEM, ng/g tissue: Table 1) and in general, the concentration of monoamines differed between old CR and AL animals. ANOVA revealed significant between-group differences in the NA levels in the cerebellum [F(2,27) = 4.341; p < 0.05], midbrain [F(2,27) = 5.844; p < 0.01], hypothalamus [F(2,27) = 4.695; p < 0.05], thalamus [F(2,27) = 3.60; p < 0.05], hippocampus [F(2,27) = 4.92; p < 0.05] and striatum [F(2,27) = 5.612; p < 0.01]. The contrast analysis identified a decrease in NA in aged animals relative to the adult group in the cerebellum, midbrain, hypothalamus and striatum, and an increase in the hippocampus. Nevertheless, the CR intervention compensated such deficits as old CR animals had higher levels of NA than old AL rats in the cerebellum, hypothalamus, thalamus and striatum, with levels similar to those in the adult animals.
Dopaminergic system
Some significant between-group differences in the dopaminergic system were detected in specific brain areas (Table 1). There was a significant decrease in DA in the striatum of old AL animals relative to adult animals [F(2,27) = 10.966, p < 0.001], as well as in DOPAC [F(2,27) = 7.659, p < 0.01], HVA [F(2,27) = 4.987, p < 0.05] and TH [F(2,17) = 4.248, p < 0.05] (Fig. 1A,B and Fig. 1 supplementary information), such a decline was attenuated in the old CR group. Moreover, old AL rats had less DOPAC in the midbrain [F(2,27) = 4.252, p < 0.05] and prefrontal cortex [F(2,27) = 6.514, p < 0.01], and an increase was seen in the hippocampus [F(2,27) = 5.183, p < 0.05]. The old CR group had higher levels of DOPAC in the midbrain and more DA in the hypothalamus [F(2,27) = 3.733, p < 0.05] than the old AL rats, with similar levels to the adult group. There were no significant differences in L-DOPA in any of the brain areas studied.

Histograms of the integrated densitometry of proteins represented as the percentage change, taking the intensity in adult’ rats as 100% for each region: (A) Striatum TH and (B) Hippocampal TrpH. The values represent the mean ± SEM of the three groups: #p < 0.05 Old Al vs Adult; *p < 0.05 Old AL vs Old CR.
Serotoninergic system
There were significant between-group differences in the levels of 5-HT in the midbrain [F(2,27) = 7.461, p < 0.01], hypothalamus [F(2,27) = 4.695, p < 0.05] and Striatum [F(2,27) = 10.096, p < 0.001], and in the levels of 5-HT and 5-HIAA in the Pons [F(2,27) = 9.650, p < 0.001; F(2,27) = 4.516, p < 0.05]. In the hippocampus, there were also significant between-group differences in 5-HT [F(2,27) = 6.077, p < 0.01], 5-HIAA [F(2,27) = 10.677, p < 0.001] and TrpH [F(2,18) = 5.170, p < 0.05] (Fig. 1C,D). The contrast analysis showed that the old AL animals expressed less 5-HT in the pons, midbrain and striatum than the adult rats. Moreover, old CR animals had more 5-HT in the pons, hypothalamus and striatum than those in the old AL group. Additionally, in old AL animals there was more 5-HIAA in the pons and hippocampus than in adult rats, while an increase in 5-HIAA in the thalamus of old CR animals was accompanied by a decrease in the hippocampus relative to old AL.
The effects of CR on peripheral hormones
In terms of the peripheral hormones evaluated, there was a significant between-group difference in insulin [F(2,28) = 5.843, p = 0.008] and leptin [F(2,28) = 21.821, p < 0.001] (Table 2), while no such differences were detected in corticosterone [F(2,28) = 0.688, p = 0.511] and IGF-1 [F(2,28) = 0.481, p = 0.624]. Contrast analysis indicated that the old AL animals exhibited significantly more insulin and leptin than adult rats, which was partially ameliorated in the old CR group. However, no significant differences were detected between old CR and adult rats.
Correlations between hormone and monoamine levels
When the relationship between the hormones and brain monoamine levels were assessed, significant differences were evident (Table 3). The levels of both insulin and leptin were negatively correlated with the levels of NA in the striatum and prefrontal cortex (p < 0.05), and with the DOPAC, DA and 5-HT levels in the striatum (p < 0.05). Moreover, leptin was negatively correlated to the levels of DOPAC in the prefrontal cortex (p < 0.05).
The effects of CR on biochemical parameters
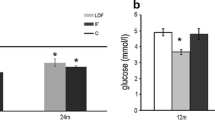
Regarding the other biochemical parameters assessed, significant between-group differences were only detected for the levels of triglyceride [F(2.28) = 8.479. p = 0.001], albumin [F(2.28) = 7.836. p = 0.002], calcium [F(2.28) = 4.833. p = 0.016] and HDL-c [F(2.28) = 3.841. p = 0.035], but not for the other parameters examined: glucose F(2.28) = 1.961, p = 0.161; total protein F(2.28) = 2.458, p = 0.105; ALP F(2.28) = 3.008, p = 0.067; cholesterol F(2.28) = 1.516, p = 0.241; and LDL-c F(2.28) = 3.012, p = 0.067 (Table 4). A post hoc analysis demonstrated that the old AL group accumulated more triglycerides (p < 0.01) and less albumin (p < 0.05), calcium (p < 0.05) and HDL-c (p < 0.05) than the adult rats. Moreover, the old CR rats had more albumin (p < 0.01), calcium (p < 0.05) and HDL-c (p < 0.05) than the old AL rats. In terms of the other parameters measured, no significant differences were detected between the old CR and adult rats.
Discussion
In this study, maintaining a CR diet from 4-months of age ameliorated the decline in monoamines in specific brain areas, as well as the increase in plasma leptin and insulin associated with aging. The correlation results also indicate that the beneficial effects of CR on some peripheral hormones parallel the positive changes in monoamine neurotransmission in old animals. Moreover, the analysis of several biochemical parameters, such as glucose, total protein and ALP, did not indicate health alterations associated with the CR diet.
Our findings suggest that the changes in monoamines and the effects of the CR diet might be specific for particular brain regions and specific neurotransmitters. Thus, there is a decrease in the NA in the striatum, cerebellum and midbrain of old AL rats, and an increase in the hippocampus relative to the adult rats. These results are consistent with previous data in rats whereby age-related deficits in noradrenergic transmission were described in the midbrain5,10,12 and striatum in male7 and in female6 animals. Moreover, an increase of NA in the hippocampus has also been reported previously10,12. Although aging has also been seen to produce a decrease in hippocampal NA in old male Wistar rats9 and female Long-Evans rats6 or even no changes7,8, as in the hypothalamus7,25,26 and frontal cortex in male8 and in females rats6. Interestingly, CR increased NA in these brain regions relative to rats fed AL, as well as in the thalamus and cerebellum. Moreover, previous studies indicated increases in NA levels in the hippocampus27 and decreases in the caudate nucleus and hypothalamus28. In the latter study, in which CR diet was more severe than the one used in the present study, the NA reduction in the striatum was detected in male but not in female rats, contrary to what was observed in the hypothalamus.
The results presented here confirm that the levels of DA and its metabolites tend to decline with age in the striatum of male Wistar rats5,8,9,10,13,29. However, results in female rats seem to be different, as no changes of DA in this brain area have been reported6,30. Indeed, a previous study reported that neurons of the substantia nigra degenerate with age in male rats, especially those projecting to the dorsolateral region of the striatum31. In turn, a lower rate of DA replacement25, and in the synthesis and expression of the DA transporter, has been seen in the striatum of male aged rats32. Regarding the hippocampus, no changes in the levels of DA and metabolites have been detected, similarly to previous reports in male rats4,7,8,13,30,33, but in contrast to the decreases observed in Long-Evans female rats6 and Sprague-Dawley male rats4,5. In terms of the effects of diet and when compared to old AL animals, old rats subjected to CR expressed more TH, DOPAC and DA in the striatum, more DOPAC in the frontal cortex and midbrain, and less DOPAC in the hippocampus. Previous studies in male rats have found that CR increased DA in the Substantia nigra34,35, and that a reduction in body weight improved DA receptor signaling36 and enhanced striatum DA release37. However, a more severe CR diet reduced DA in the striatum of male rats28.
Concerning the serotonergic system, the old AL rats exhibited more 5-HT and 5-HIAA in the hippocampus than adult animals, a modification that was reverted in the CR rats. Increases in HT and/or 5-HIAA in the hippocampus have been previously reported in rats4,5,7,8,29 but such a results are in disagreement with others who found no decreases9,10,30 or even no effect of age6,11,13,33. An age-dependent decline in the serotoninergic pathways was also found here in the striatum5,6,7,9,13 and hypothalamus26. In addition, CR recovered the age-dependent 5-HT decrease in the pons29, midbrain, hypothalamus and striatum. In general, CR seems to combat the effects of aging on monoamine levels, although the action of CR in specific brain areas remain unclear as the type of diet, age, sex and strain/species may affect, suggesting that neurotransmitter brain levels are highly sensitive to such variables that are important to take into consideration in future studies. In fact, it has been described that dietary interventions can have different effects on monoamines depending on the sex of the animals6,7,30.
The effects of CR on peripheral hormones like leptin and insulin are consistent with earlier studies confirming that CR attenuates the age-related increase in insulin and leptin38. Indeed, the reduced activity of these metabolic pathways promotes health and enhances the animal’s lifespan39. Moreover, both insulin and leptin are negatively correlated with the levels of NA in the striatum and prefrontal cortex, and the levels of DOPAC, DA and 5-HT in the striatum, while the leptin concentration is also negatively correlated to DOPAC levels in the prefrontal cortex, which suggests that regulating these hormones through CR may enhance monoaminergic neurotransmission in specific brain regions. As for other hormones, no differences in IGF-1 and corticosterone were evident between adult and old animals, which contrasts with earlier studies showing that dietary interventions might diminish40 or even augment16 serum IGF-1 levels, and upregulate circulating blood levels of corticosterone41. However, similar levels of corticosterone were previously found in aged AL and CR animals42, in accordance with our results and confirming that CR does not affect stress hormones.
To control the effects of CR on the animals’ general health, biochemical parameters were analyzed, such as cholesterol, glucose, total proteins, triglycerides, albumin, calcium, LDL-c, HDL-c and ALP. Old AL rats had more triglycerides and less HDL-c, albumin and calcium in the blood than the adult rats, while CR reverted some of these changes (e.g. triglycerides, albumin, HDL-c and calcium in the blood). The LDL-c levels were higher in both the aged groups relative to the adult animals, in agreement with a previous study on the effects of dietary restriction in rats43. Moreover, our results confirm the capacity of CR to attenuate triglycerides, thereby improving cardiac function and health during aging44. Finally, no changes in total protein and glucose have been detected due to aging, although CR could ameliorate the age related decline in plasma albumin and calcium.
The present study is one of the few experimental studies to investigate the effects of CR on monoamines and peripheral hormones, although some limitations should be noted. First, the experiment was performed on males and as discussed before, sex may be an important variable to consider. Another important factor is the experimental design, as we used a long-life dietary intervention evaluating the animals when they were 24-months-old, while other studies have used shorter periods of CR or intermittent interventions. Moreover, we did not record the biometric parameters of animals, such as body fat, lean body mass, bone mass or length. Considering that the CR lasted for almost two years, these parameters could well be of particular interest to assess the effects of CR on the animals’ health. Furthermore, the lack of cognitive data makes it difficult to interpret the functional effects associated with the neurochemical changes found. However, other studies in our laboratory showed positive effects of CR on monoaminergic transmission that correlate with better spatial memory27. Thus, further research is needed to identify the potential role of sex-dependent differences in caloric requirements and how a CR diet applied at different time points in life affects monoamine levels5,13.
Currently, dietary interventions and healthier eating habits remain the most applicable and cost-efficient means of preventing a wide variety of age-related diseases. Although a low-caloric diet throughout life is unlikely to be feasible for most people, a growing body of evidence demonstrates that CR exerts beneficial effects on brain aging at multiple levels. In a recent study on humans45, it was demonstrated that reduced caloric intake is geroprotective, which indicates that this metabolic intervention influences the immune system and modifies the aging process. However, further studies will be needed to fully elucidate whether CR may represent a novel therapeutic intervention for healthy aging.
In summary, it has been long known that CR has some protective effects on aging brain, but studies investigating the biochemical effects of CR on brain aging are scarce. In this study, we show that in general, a lifelong CR diet can attenuate the age-related decline in monoaminergic neurotransmission observed in some brain regions. The beneficial effects of CR on brain monoamines parallels the improved metabolic regulation. Hence, it cannot be ruled out that attenuation of the age-related increase in insulin and leptin may help to restore brain monoamine levels in brain areas like the hippocampus and frontal cortex.
Materials and methods
Subjects
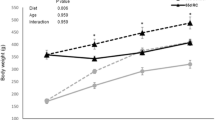
This study was carried out on 30 male Wistar rats from our laboratory’s breeding stock (Prolabor, Charles River Laboratories, Abresle, France). The CR group of old rats (n = 12; age = 23–24 months; weight = 483.1 g ± 91 g) were kept under conditions of CR from four months of age, with a 30% reduction of total food intake (18–20 g/day) coupled to free access to water. Another group of old rats (n = 8; age = 23–24 months; weight = 789.1 g ± 68.2 g) were given Ad libitum (AL) access to food and water, as was a control group of adult animals (n = 10; age = 3–4 months; weight = 369.6 ± 22.9 g). The animals were fed with dry pellets (Harlan Laboratories Inc., Madison, USA) produced and packed by Mucedola Sri. (MI, Italy). Animals were pair-housed from the beginning of the experiment and maintained at a constant temperature (21 °C) on a controlled 12 h light/dark cycle. The animal housing procedures are explained in more detail elsewhere46. All procedures were performed in compliance with protocols approved by the Animal Care and Use Committee of Autonomous University of Barcelona (CEEAH number 3866), with authorization from Department of Environment of the Generalitat de Catalunya, and with guidelines approved by the EU Council Directive for the care and use of laboratory animals (2010/63/EU).
Blood samples and plasma analysis
Food was removed the day before sacrifice to minimize the biochemical and hormonal differences between the groups due to the amount of food consumed immediately before47 or due to lipidemia48. After sacrificing the animals by decapitation, blood samples were collected in Heparin tubes (Sodium Heparin, 5000 USP/mL; Chiesi Spain, SA, Spain) and placed on ice for slow coagulation. Cell-free plasma was obtained by centrifugation at 6000 rpm for 15 min at 4 °C (RCF 2361 × g) and it was stored at − 80 °C until biochemical analysis. These frozen plasma samples were allowed to reach room temperature before processing and analyzing the levels of: corticosterone, insulin, leptin, IGF-1, cholesterol, glucose, total protein, triglycerides, albumin, LDL-c, calcium, HDL-c and ALP. Corticosterone levels were measured at the Servei de Bioquímica, Clinica Veternitaria (Universitat Autònoma de Barcelona) with a competitive Corticosterone EIA (Immunodiagnostic Systems Ltd, Bolton, UK), while the insulin, leptin and IGF-1 levels were assessed using Sandwich ELISAs, all quantified on a EMS Reader MF V.2.9-0: Mercodia Rat Insulin ELISA (Mercodia AB, Sweden); Quantikine ELISA Mouse/Rat leptin (R&D Systems, Inc. USA) Quantikine Mouse/Rat IGF-I ELISA (R&D Systems, Inc. USA). Cholesterol was analyzed by the CHE/POD1 enzyme method, glucose by the Hexokinase method, and the total protein was calculated by the Biuret method. Triglycerides were quantified by the Glycerol-3-phosphate oxidase method, albumin levels by the Bromocresol green method, calcium levels by the Arzenazo III method, LDL-c was quantified by the selective protection method and HDL by the immunoinhibition method, and the ALP levels were assessed by the Substrate 4-nitrophenyl phosphate method using AMP buffer. All these analyses were carried out on an AU400 Olympus analyzer (Germany) with OSR reagents (Olympus System Reagent, Beckman Coulter, Ireland).
Tissue collection and processing
After decapitation, the rat’s brain was rapidly removed and dissected into different regions that were weighed, frozen and stored at − 80 °C. The following bilateral brain regions were collected and processed for HPLC-ED analysis: frontal cortex, hippocampus, striatum, thalamus, cerebellum, pons, midbrain, hypothalamus, and occipital cortex. The striatum and hippocampus were also analyzed in Western Blots.
Quantification of monoamines and their metabolites by HPLC-ED
Brain samples were homogenized in buffer (perchloric acid 60% w/w 0.25 M, sodium metabisulphite 100 µM, EDTA Na2·2H2O 250 µM) in a 9/1 ratio (p/v; ml/mg). A polytron homogenizer was used to rapidly disrupt the animal tissue and the homogenate was centrifuged for 10 min at 15,000 rpm and 4 °C (5417 R centrifuge). The supernatant recovered was filtered and 50 µL aliquots were analyzed on a reverse phase column (Cromolith Performance, 4.6 mm internal diameter × 10 cm length) coupled to a pre-column (4.6 mm × 5 cm). The mobile phase consisted of 0.1 M citric acid, 0.05 M EDTA, 1.2 mM SOS, 10% acetonitrile (v/v) adjusted to pH 2.75 with tetraethylammonium. Elution was performed at a flow rate of 0.8 mL/min and the HPLC apparatus (LaChrom Elite) was coupled to an electrochemical detector (ED: ESA Coulochem 5100A), with an ESA analytical dual electrode cell 5011A (the detection potential for electrodes 1 and 2 was set at 70.05 and + 0.4 V, respectively). Before and after each round of processing the samples, one calibration mix was analyzed at seven dilutions: 12.5, 25, 50, 62.5, 125, 250 and 500 μg/mL. Depending on the retention time of each component, calibration lines (r2 = 0.999) allowed of the concentration of the monoamines (NA, DA, 5-HT) and their metabolites (L-DOPA, DOPAC, HVA, 5-HIAA) to be calculated. EZChrom Elite Software was used to determine the appropriate concentrations, expressed as ng/g tissue.
Semi-quantitative western blots
Striatal and hippocampal tissue was collected at different time-points and washed in lysis buffer (25 mM Tris–HCl, 150 mM NaCl, 0.5% Sodium deoxycholate, 0.1% SDS, 1% NP-40 [pH 7.6]). The lysates were homogenized with a pestle (Sigma-Aldrich Corp., Madrid, Spain), sonicated and quantified using a BCA assay (Pierce Chemical Co.). Equal amounts of protein (30 µg/well) were resolved on SDS-PAGE gels and transferred at 100 V to a nitrocellulose membrane for 1 h (Whatman, Dassel, Germany) in a Mini TransBlot Cell (Bio-Rad; Hercules, CA, USA). The membranes were blocked for 1 h at 20–25 °C with 5% non-fat dry milk in Tris-buffered saline (TBS: 75 mM NaCl, 1.5 mM KCl, 12.4 mM Tris [pH 7.4]) and they were then probed overnight at 4 °C with the corresponding primary antibody diluted in 5% (w/v) bovine serum albumin (BSA), with shaking. The following primary antibodies were used: monoclonal mouse anti-tyrosine hydroxylase (T2928, Sigma, USA, 1:1000), monoclonal mouse anti-tryptophan hydroxylase (T06981, Sigma, USA, 1:500), and mouse anti-β-tubulin (Becton–Dickinson, Franklin Lakes, NJ, USA, 1:1000). After several washes with TBS 0.1% + Tween 20, the membranes were incubated for 1 h with the corresponding horseradish peroxidase conjugated secondary antibody, anti-mouse-HRP (Dako Denmark, Glostrup, Denmark, 1:3000). Blots were developed using a chemoluminiscent mix 1:1 (0.5 M luminol, 79.2 mM p-coumaric acid, 1 M Tris–HCl [pH 8.5], and 8.8 M hydrogen peroxide, 1 M Tris–HCl [pH 8.5]). All the samples to be compared were processed at the same time, transferred simultaneously to a membrane and probed with the same antibody dilution. The apparent molecular weight of the proteins was determined by calibrating the blots with pre-stained molecular weight markers (All Blue: Pierce Chemical Co., USA) and densitometry was carried out using ImageJ software (National Institute of Health, Bethesda, MD, USA). The total content of each specific protein (TH and TrpH) was assayed after the membranes were stripped for 1 h at 20–25 °C with Glycine (0.1 mM, pH 2.3), blocked again and incubated with the corresponding primary antibody. Chemiluminescence signals of the bands obtained were all within the linear range of the imaging system and were not saturated (ChemiDoc XRS + System, Bio-Rad Laboratories). Densitometry and quantitation was carried out using ChemiDoc MP Imaging System, Image Lab program (Bio-Rad) and Microsoft Excel was used to determine the levels of proteins. The adult group of subjects was used as a control group, and the total level of proteins is expressed as a percentage.
Statistical analysis
The data were analyzed using the SPSS v20 package and plotted as the mean ± SEM. Statistical analysis was performed using one-way ANOVA (groups: adult, old AL and old CR) and a Bonferroni correction was applied for multiple comparisons. In addition, Spearman Rank correlations were established to examine the relationship between the hormonal and biochemical variables. A P-value of ≤ 0.05 was considered to be statistically significant.
References
Abbott, A. Cognition: The brain’s decline. Nature 492, S4–S5 (2012).
Anderton, B. H. Ageing of the brain. Mech. Ageing Dev. 123, 811–817 (2002).
Fjell, A. M. & Walhovd, K. B. Structural brain changes in aging: Courses, causes and cognitive consequences. Rev. Neurosci. 21, 187–221 (2010).
Godefroy, F., Bassant, M. H., Weil-Fugazza, J. & Lamour, Y. Age-related changes in dopaminergic and serotonergic indices in the rat forebrain. Neurobiol. Aging 10, 187–190 (1989).
Míguez, J. M., Aldegunde, M., Paz-Valiñas, L., Recio, J. & Sánchez-Barceló, E. Selective changes in the contents of noradrenaline, dopamine and serotonin in rat brain areas during aging. J. Neural Transm. 106, 1089–1098 (1999).
Stemmelin, J., Lazarus, C., Cassel, S., Kelche, C. & Cassel, J. C. Immunohistochemical and neurochemical correlates of learning deficits in aged rats. Neuroscience 96, 275–289 (2000).
Tanila, H., Taira, T., Piepponen, T. P. & Honkanen, A. Effect of sex and age on brain monoamines and spatial learning in rats. Neurobiol. Aging 15, 733–741 (1994).
Lee, J. M. et al. Spatial learning deficits in the aged rat: Neuroanatomical and neurochemical correlates. Brain Res. Bull. 33, 489–500 (1994).
Esteban, S. et al. Improving effects of long-term growth hormone treatment on monoaminergic neurotransmission and related behavioral tests in aged rats. Rejuvenation Res. 13, 707–716 (2010).
Koprowska, M., Krotewicz, M., Romaniuk, A. & Strzelczuk, M. Age-related changes in fear behavior and regional brain monoamines distribution in rats. Acta Neurobiol. Exp. (Wars) 64, 131–142 (2004).
Ponzio, F. et al. Changes in monoamines and their metabolite levels in some brain regions of aged rats. Neurobiol. Aging 3, 23–29 (1982).
Ida, Y. et al. Effects of age and stress on regional noradrenaline metabolism in the rat brain. Neurobiol. Aging 3, 233–236 (1982).
Yurek, D. M., Hipkens, S. B., Hebert, M. A., Gash, D. M. & Gerhardt, G. A. Age-related decline in striatal dopamine release and motoric function in Brown Norway/Fischer 344 hybrid rats. Brain Res. 791, 246–256 (1998).
Baranowska-Bik, A. & Bik, W. Insulin and brain aging. Przeglad Menopauzalny 16, 44–46 (2017).
Arai, Y., Kamide, K. & Hirose, N. Adipokines and aging: Findings from centenarians and the very old. Front. Endocrinol. 10, 142 (2019).
Sonntag, W. E., Ramsey, M. & Carter, C. S. Growth hormone and insulin-like growth factor-1 (IGF-1) and their influence on cognitive aging. Ageing Res. Rev. 4, 195–212 (2005).
Lupien, S. B., Bluhm, E. J. & Ishii, D. N. Systemic insulin-like growth factor-I administration prevents cognitive impairment in diabetic rats, and brain IGF regulates learning/memory in normal adult rats. J. Neurosci. Res. 74, 512–523 (2003).
van den Beld, A. W. et al. The physiology of endocrine systems with ageing. Lancet Diabetes Endocrinol. 6, 647–658 (2018).
Santos, C. L., Bobermin, L. D., Souza, D. O. & Quincozes-Santos, A. Leptin stimulates the release of pro-inflammatory cytokines in hypothalamic astrocyte cultures from adult and aged rats. Metab. Brain Dis. 33, 2059–2063 (2018).
van den Ameele, S. et al. Markers of inflammation and monoamine metabolism indicate accelerated aging in bipolar disorder. Front. Psychiatry 9, 250 (2018).
Capuron, L. et al. Chronic low-grade inflammation in elderly persons is associated with altered tryptophan and tyrosine metabolism: Role in neuropsychiatric symptoms. Biol. Psychiatry 70, 175–182 (2011).
Ribarič, S. Diet and aging. Oxid. Med. Cell. Longev. 2012, 1–20 (2012).
Hadem, I. K. H., Majaw, T., Kharbuli, B. & Sharma, R. Beneficial effects of dietary restriction in aging brain. J. Chem. Neuroanat. 95, 123–133 (2019).
Van Cauwenberghe, C., Vandendriessche, C., Libert, C. & Vandenbroucke, R. E. Caloric restriction: Beneficial effects on brain aging and Alzheimer’s disease. Mamm. Genome 27, 300–319 (2016).
Carfagna, N., Trunzo, F. & Moretti, A. Brain catecholamine content and turnover in aging rats. Exp. Gerontol. 20, 265–269 (1985).
Rodríguez-Gómez, J. A., de la Roza, C., Machado, A. & Cano, J. The effect of age on the monoamines of the hypothalamus. Mech. Ageing Dev. 77, 185–195 (1995).
Rojic-Becker, D., Portero-Tresserra, M., Martí-Nicolovius, M., Vale-Martínez, A. & Guillazo-Blanch, G. Caloric restriction modulates the monoaminergic and glutamatergic systems in the hippocampus, and attenuates age-dependent spatial memory decline. Neurobiol. Learn. Mem. 166, 107107 (2019).
Kolta, M. G., Holson, R., Duffy, P. & Hart, R. W. Effect of long-term caloric restriction on brain monoamines in aging male and female fischer 344 rats. Mech. Ageing Dev. 48, 191–198 (1989).
Goicoechea, C., Ormazábal, M. J., Alfaro, M. J. & Martín, M. I. Age-related changes in nociception, behavior, and monoamine levels in rats. Gen. Pharmacol. 28, 331–336 (1997).
Luine, V., Bowling, D. & Hearns, M. Spatial memory deficits in aged rats: contributions of monoaminergic systems. Brain Res. 537, 271–278 (1990).
Diaz-Ruiz, C., Rodriguez-Perez, A. I., Beiroa, D., Rodriguez-Pallares, J. & Labandeira-Garcia, J. L. Reciprocal regulation between sirtuin-1 and angiotensin-II in the Substantia nigra: Implications for aging and neurodegeneration. Oncotarget 6, 26675–26689 (2015).
Cruz-Muros, I. et al. Aging effects on the dopamine transporter expression and compensatory mechanisms. Neurobiol. Aging 30, 973–986 (2009).
Nakamura, S. & Ohno, T. Spatial learning impairment in aged rats: Comparing between aged basal forebrain lesioned and normal aged rats. Behav. Brain Res. 70, 69–76 (1995).
Salvatore, M. F. et al. Dissociation of striatal dopamine and tyrosine hydroxylase expression from aging-related motor decline: Evidence from calorie restriction intervention. J. Gerontol. A. Biol. Sci. Med. Sci. 73, 11–20 (2017).
Salvatore, M. F. et al. Initiation of calorie restriction in middle-aged male rats attenuates aging-related motoric decline and bradykinesia without increased striatal dopamine. Neurobiol. Aging 37, 192–207 (2016).
Carr, K., Tsimberg, Y., Berman, Y. & Yamamoto, N. Evidence of increased dopamine receptor signaling in food-restricted rats. Neuroscience 119, 1157–1167 (2003).
Diao, L. H., Bickford, P. C., Stevens, J. O., Cline, E. J. & Gerhardt, G. A. Caloric restriction enhances evoked DA overflow in striatum and nucleus accumbens of aged Fischer 344 rats. Brain Res. 763, 276–280 (1997).
Most, J. & Redman, L. M. Impact of calorie restriction on energy metabolism in humans. Exp. Gerontol. 133, 110875 (2020).
Santos, J., Leitão-Correia, F., Sousa, M. J. & Leão, C. Dietary restriction and nutrient balance in aging. Oxid. Med. Cell. Longev. 2016, 1–10 (2016).
Fontana, L., Weiss, E. P., Villareal, D. T., Klein, S. & Holloszy, J. O. Long-term effects of calorie or protein restriction on serum IGF-1 and IGFBP-3 concentration in humans. Aging Cell 7, 681–687 (2008).
Montaron, M. F. et al. Lifelong corticosterone level determines age-related decline in neurogenesis and memory. Neurobiol. Aging 27, 645–654 (2006).
Moneo, M., Zúñiga, J. M. & Morón, I. Caloric restriction in grouped rats: Aggregate influence on behavioural and hormonal data. Lab. Anim. https://doi.org/10.1177/0023677216686805 (2017).
Chen, J. H. et al. A moderate low-carbohydrate low-calorie diet improves lipid profile, insulin sensitivity and adiponectin expression in rats. Nutrients 7, 4724–4738 (2015).
Mager, D. E. et al. Caloric restriction and intermittent fasting alter spectral measures of heart rate and blood pressure variability in rats. FASEB J. 20, 631–637 (2006).
Ma, S. et al. Caloric restriction reprograms the single-cell transcriptional landscape of Rattus Norvegicus aging. Cell 180, 984-1001.e22 (2020).
Portero-Tresserra, M., Martí-Nicolovius, M., Guillazo-Blanch, G., Boadas-Vaello, P. & Vale-Martínez, A. d-Cycloserine in the basolateral amygdala prevents extinction and enhances reconsolidation of odor-reward associative learning in rats. Neurobiol. Learn. Mem. 100, 1–11 (2013).
Kmiec, Z. Central regulation of food intake in ageing. J. Physiol. Pharmacol. 57(Suppl 6), 7–16 (2006).
Nikolac, N. Lipemia: Causes, interference mechanisms, detection and management. Biochem. Med. 24, 57–67 (2014).
Acknowledgements
This work was supported by funds from the Ministerio de Economía y Competitividad (PSI2017-84290-R). The authors thank Dr Mark Sefton (BiomedRed SL) for his support with author editing.
Author information
Authors and Affiliations
Contributions
M.P. and M.M. wrote the main manuscript text. M.P., D.R. and C.V performed all the experimental work. M.P., M.M and G.G. designed the experiment. M.P. and C.V. prepared tables and figures. M.P. and A.V. performed and review thoroughly the statistical analysis. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Portero-Tresserra, M., Rojic-Becker, D., Vega-Carbajal, C. et al. Caloric restriction modulates the monoaminergic system and metabolic hormones in aged rats. Sci Rep 10, 19299 (2020). https://doi.org/10.1038/s41598-020-76219-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-76219-7
- Springer Nature Limited