
Abstract
Acinetobacter baumannii is a common pathogen of nosocomial infection, and its ability to form biofilms further contributes to its virulence and multidrug resistance, posing a great threat to global public health. In this study, we investigated the inhibitory effects of five biofilm inhibitors (BFIs) (zinc lactate, stannous fluoride, furanone, azithromycin, and rifampicin) on biofilm formation of nine extensively drug-resistant A. baumannii (XDRAB), and assessed the synergistic antibacterial effects of these BFIs when combined with one of four conventional anti-A. baumannii antibiotics (imipenem, meropenem, tigecycline, and polymyxin B). Each of the five BFIs tested was found to be able to significantly inhibit biofilm formation of all the clinical isolates tested under sub-minimal inhibitory concentrations. Then, we observed synergistic effects (in 22%, 56% and 11% of the isolates) and additive effects (56%, 44% and 44%) when zinc lactate, stannous fluoride and furanone were combined with tigecycline, respectively. When zinc lactate and stannous fluoride were each used with a carbapenem (imipenem or meropenem), in 33% and 56–67% of the isolates, they showed synergistic and additive effects, respectively. Additivity in > 50% of the isolates was detected when rifampicin was combined with imipenem, meropenem, tigecycline, or polymyxin B; and a 100% additivity was noted with azithromycin-polymyxin B combination. However, antagonism and indifference were noted for polymyxin B in its combination with zinc lactate and stannous fluoride, respectively. In conclusion, five BFIs in combination with four antibacterial drugs showed different degrees of in vitro synergistic and additive antibacterial effects against XDRAB.
Similar content being viewed by others


Introduction
Acinetobacter baumannii is a major gram-negative opportunistic pathogen causing hospital infections1. Clinical isolates of A. baumannii not only possess strong intrinsic resistance to a variety of structurally unrelated antibiotics but also show increasing acquired multidrug or extensive drug resistance1,2. In particular, carbapenem-resistant A. baumannii has been categorized by the World Health Organization as one of the 12 top priority resistant bacteria that pose the greatest threat to human health with the urgent need for developing new antibiotics3. A. baumannii also possesses strong biofilm formation ability, which further enhances its resistance to antibacterial drugs and circumvents the host immune-mediated clearance, and thus greatly contributes to recurrent infections or chronic infections4,5,6,7.
Biofilms are surface-attached population of bacterial cells, which are encased in self-produced extracellular polymers. The latter mainly includes a range of substances such as polysaccharides, proteins, nucleic acids, and lipids, forming a dense physical barrier that renders, along with other mechanisms, biofilm cells much more resistant to antibacterial drugs than their non-adherent planktonic cells8,9. In recent years, much attention has been directed to uncovering new approaches including combination therapy against biofilm infections10,11.
Control of bacterial biofilm formation is considered as one major strategy against bacterial infections, and thus efforts have been made to identify effective inhibitors that go beyond conventional antibiotics for their ability to disrupt biofilm formation10. For example, it has been reported that three chemical substances, zinc lactate, stannous fluoride and furanone, and two antibiotics, azithromycin and rifampicin, can inhibit bacterial biofilm formation, but cannot effectively control the infection caused by biofilm-forming bacteria12,13,14,15,16,17. Yet, information available is largely not specific to A. baumannii. In this study, we aimed to assess inhibitory effects of five aforementioned biofilm inhibitors (BFIs) on the biofilm formation of extensively drug-resistant A. baumannii (XDRAB) and to explore if these inhibitors could synergistically interplay with one of four conventional anti-A. baumannii antibiotics (imipenem, meropenem, tigecycline or polymyxin B) against XDRAB. The results obtained warrant further investigations towards the combination strategy for their potential in the prevention and treatment of the infections caused by biofilm-forming XDRAB.
Results
Antimicrobial susceptibility testing
Antibacterial activity of 15 antibiotics against nine clinical isolates of A. baumannii is shown in Table 1. These tested agents include a variety of structurally unrelated antibiotics belonging to nine classes or subclasses of β-lactam-β-lactamase inhibitors, aminoglycosides, carbapenems, cephalosporins, fluoroquinolones, glycylcyclines, phenicols, polymyxins, and tetracyclines. Most of these antibiotics tested are clinically relevant as drugs of choice in the treatment of A. baumannii infections1,18. According to the interpretive categories established by the Clinical and Laboratory Standards Institute (CLSI)18, these isolates were either susceptible or non-susceptible to tigecycline and/or polymyxin B, but were resistant to other 13 antibacterial drugs (Table 1), and such drug resistance profiles placed these isolates as XDRAB19. In addition, the five BFIs selected for this study, zinc lactate, stannous fluoride, furanone, azithromycin, and rifampicin, displayed different degrees of antibacterial effects on these XDRAB, with identical MIC values of 512, 512, 256, 1024, and 1 µg/ml, respectively, for all nine isolates tested. Based on these MIC values, except for rifampicin which antibacterial activity is demonstrated, activity of zinc lactate, stannous fluoride, furanone and azithromycin against A. baumannii is consider limited.
Sub-inhibitory concentrations of the biofilm inhibitors
Based on literature information12,13,14,15,16,17, we have identified a range of metal salts, biocides and conventionally used antibiotics with ability to affect bacterial biofilm formation. However, no or little data were specifically available to A. baumannii. We found that zinc lactate, stannous fluoride, furanone, azithromycin, and rifampicin did not affect the growth of planktonic bacterial cells at the sub-inhibitory concentrations (i.e., 1/8–1/2 MICs) of 256, 256, 32, 256, and 0.25 µg/ml, respectively (Fig. 1). Therefore, these concentrations were established as the working concentrations of the BFIs for assessing their effects on biofilm formation and combination use with anti-A. baumannii drugs.

Bacterial growth curves of 9 extensively drug-resistant A. baumannii in the presence of sub-inhibitory concentrations of 5 biofilm inhibitors tested. Control, no biofilm inhibitor; AMZ azithromycin (1/4 MIC = 256 µg/ml), FUR furanone (1/8 MIC = 32 µg/ml), RIF rifampicin (1/4 MIC = 0.25 µg/ml), SnF stannous fluoride (1/2 MIC = 256 µg/ml), and ZnL zinc lactate (1/2 MIC = 256 µg/ml). Data are shown as mean ± SD (n = 9).
Effects of sub-inhibitory concentrations of BFIs on XDRAB biofilm formation
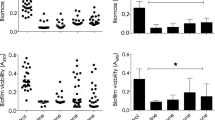
With the sub-inhibitory concentrations of 5 BFIs described above, we tested the effects of these BFIs on biofilm formation. Compared with the no BFI control group, at the sub-inhibitory concentration of each agent, the five BFIs were found to significantly inhibit the biofilm formation of all 9 XDRAB isolates (P < 0.05 or < 0.01) (Fig. 2). The strongest effect was observed with zinc lactate (mean OD570 decreased of 0.78, from 1.52 to 0.74), followed by stannous fluoride, furanone, and rifampicin. The weakest effect was observed when azithromycin was used, with an average value of OD570 decrease of 0.27 (from 1.52 to 1.25) (Fig. 2). At the assay conditions of this study, the inhibitory effects of the five tested BFIs on the biofilm formation of XDRAB are ranked as: zinc lactate > stannous fluoride > furanone > rifampicin > azithromycin.

Effects of the sub-inhibitory concentration of 5 biofilm inhibitors on the biofilm formation of 9 isolates of extensively drug-resistant A. baumannii as measured via the decolorization solution of crystal violet stained biofilm cells. Control, cells with no biofilm inhibitor; AMZ azithromycin (1/4 MIC = 256 µg/ml), FUR furanone (1/8 MIC = 32 µg/ml), RIF rifampicin (1/4 MIC = 0.25 µg/ml), SnF stannous fluoride (1/2 MIC = 256 µg/ml); ZnL zinc lactate (1/2 MIC = 256 µg/ml). Data are shown as mean ± SD (n = 9; significance **P < 0.01 and *P < 0.05 calculated by t-test).
Combination antimicrobial drug susceptibility
Given the observed effects from sub-inhibitory BFIs on biofilm formation, we further tested how a combination use of a BFI with a clinically-relevant anti-A. baumannii antibiotic could interplay. Based on the fractional inhibitory concentration index (FICI) values generated from the combination drug susceptibility testing (Table 2), we observed that when zinc lactate was used in combination with imipenem, meropenem, and tigecycline respectively, 33%, 33%, and 22% of the isolates showed synergistic effects, and 67%, 67%, and 56% showed additive effects. However, when combined with polymyxin B, we detected an antagonistic effect. When stannous fluoride was used with imipenem, meropenem, and tigecycline, synergism was detected in 33%, 44%, and 56% of the isolates, respectively, while additivity was noted in 67%, 56%, and 44% of the isolates. Its combination with polymyxin B resulted in indifferent effects. Additivity was detected when furanone was combined with imipenem, meropenem or tigecycline (100%, 100% or 44% respectively). The combination of furanone and polymyxin B yielded either indifferent or antagonistic effect. Combination of rifampicin with imipenem, meropenem, tigecycline or polymyxin B led to additive effects (78%, 56%, 67% and 100%, respectively). When azithromycin was combined with polymyxin B, we observed an additive effect. Yet, azithromycin showed indifferent effects when combined with imipenem, meropenem or tigecycline.
Discussion
Our antimicrobial susceptibility testing results show that all nine isolates tested were XDRAB (i.e., non-susceptible to ≥ 1 agent in all tested drug categories but ≤ 2 categories)19, with resistance to commonly used antibacterial drugs, including β-lactam plus β-lactamase inhibitor, carbapenems, third/fourth-generation cephalosporins, aminoglycosides, and fluoroquinolones18. Some of the isolates were even non-susceptible to tigecycline and/or polymyxin B, which rendered these isolates close to be pandrug-resistant (i.e., non-susceptible to all tested drug categories)19. These findings are consistent with the fact that XDRAB has been increasingly isolated in clinical settings globally, which calls for combination therapy11. Indeed, the clinical choice of antibacterial drugs for XDRAB infections is limited, and the new therapeutic regime with combination drug use has been pursued16,17,20,21.
Given that biofilms contribute to bacterial virulence and resistance22, we targeted agents with anti-biofilm property for their potential in combination drug use against A. baumannii. At their sub-MIC levels, our results revealed that the five BFIs tested in this investigation showed different degrees of inhibitory effects on the biofilm formation of XDRAB strains, especially the zinc lactate had the strongest effect, followed by stannous fluoride, furanone, rifampicin and azithromycin at our assay conditions. The inhibition of biofilm formation by these BFIs likely occurs through different mechanisms. Studies have shown that zinc compounds can inhibit the synthesis of extracellular polysaccharides or the formation of matrix networks, and stannous fluoride can destroy the biofilm structure by loosening the structure of the biofilm matrix13,23. Furanone, a quorum-sensing system inhibitor, inhibits the biofilms formation of bacterial by replacing the binding sites of quorum sensing signal molecules24. Azithromycin can inhibit the synthesis of alginate in the biofilm of Pseudomonas aeruginosa, thereby destroying the biofilm structure, leading to the formation of channels on the biofilm that may allow the synergistic drug to pass through the biofilm and thus to reach and kill the bacteria inside25. Our findings of the effects of sub-inhibitory zinc lactate, stannous fluoride and furanone further expand the understanding of sub-inhibitory conventional antibiotics including azithromycin and rifampicin in the reduction of A. baumannii biofilm formation26.
Although BFIs of zinc lactate, stannous fluoride, furanone, and azithromycin have effects on XDRAB, their MIC values are high. Only rifampicin has a low MIC value. However, for the latter, the rapid development of RNA polymerase subunit-encoding rpoB gene mutation-mediated resistance to rifamycins limits the use of rifampicin alone against bacterial infections including A. baumannii27. According to the drug treatment principles for XDRAB, carbapenems (imipenem or meropenem), polymyxins (colistin or polymyxin B) and tigecycline are used as basic drugs in combination with other types of antibacterial drugs. Therefore, there is a clinical value in exploring how these BFIs interplay when an anti-A. baumannii antibacterial drug is used in combination with a BFI. The combination drug susceptibility testing results, as shown in Table 2, mainly reveals different levels of synergistic and additive antibacterial effects on XDRAB isolates, which suggests that BFIs may exert their actions via the reduction of biofilm formation and/or direct effect on bacterial growth to interplay with anti-A. baumannii antibiotics against XDRAB. With respect to rifampicin or azithromycin combination use with one of 4 anti-A. baumannii antibiotics, our results are largely in agreement with published in vitro studies28,29,30. The synergistic or additive effect from the combination use of zinc lactate, stannous fluoride, furanone or rifampicin with imipenem or meropenem against carbapenem-resistant XDRAB is not totally unexpected because the BFIs exerts different modes of action from that of carbapenems. An in vivo study has demonstrated efficacy of imipenem-rifampicin against carbapenem-resistant A. baumannii31. However, as our studies have limitations that focus on the measurement of in vitro activity, the clinical significance of these observations remains to be determined.
Certain combinations also showed partly indifferent or antagonistic effects. For example, there were antagonistic and indifferent effects occurring in 100% of the isolates when zinc lactate and stannous fluoride were each combined with polymyxin B, respectively. This could partly be explained by that the tested XDRAB isolates were not resistant to polymyxins and anti-A. baumannii activities of polymyxin B may mask the role of zinc and stannous compounds or these cationic metal compounds may potentially affect polymyxin’s mode of action in disrupting bacterial membrane integrity32. In other word, the positively charged group of polymyxin B can bind to the negatively charged phosphate in the phospholipids of the bacterial cell membrane, leading to the death of the bacteria32, while zinc lactate and stannous fluoride are metal cation biofilms inhibitors therefore they may compete with polymyxin B for drug targets, showing antagonistic or indifferent effect33. In contrast, the observation of indifferent effects from azithromycin combination with imipenem, meropenem, or tigecycline could be attributable to the weak anti-A. baumannii activity of azithromycin. In this regard, one study shows that another macrolide, clarithromycin, exerts antagonistic the effect with a carbapenem on P. aeruginosa34. However, the additivity between azithromycin and polymyxin is likely due to the membrane disruption of XDRAB by polymyxin B that resulted in improved accumulation of azithromycin into the cells and thus antibacterial activity32,35.
Furthermore, it is of importance to consider our in vitro drug combination synergistic results from the pharmacokinetic point of view for their potential clinical implications. Azithromycin and rifampicin are systematically administered antibiotics. While an azithromycin concentration comparable to its high MICs for A. baumannii is unlikely achievable in an in vivo situation, a pharmacokinetic-pharmacodynamic analysis of rifampicin has indicated to readily obtain pharmacokinetic parameters (such as the maximum serum concentration [Cmax] value) corresponding to relatively low rifampicin MICs for A. baumannii36. Thus, our data are in support for the use of rifampicin as one of the combination agents for treatment of XDRAB infections37,38. On other hand, the three chemical BFIs, zinc lactate, stannous fluoride and furanone, are not expected to be administered systematically, partly due to toxicity concerns. However, their inhibitory concentrations can be readily reached in topical or local use such that as zinc lactate and stannous fluoride have been used in oral care formulations (e.g., mouthwash with 1.4 mg/ml for zinc lactate or up to 16 mg/ml for stannous fluoride)39,40, which warrants further studies in their potential topical use to combat wound infections associated with A. baumannii1,41.
In conclusion, we have presented data demonstrating the inhibitory effects of five BFIs on the biofilm formation of XDRAB at the sub-inhibitory concentrations and interplay between BFIs and anti-A. baumannii antibiotics against XDRAB. Further studies are warranted for their potential clinical implications in combating biofilm-associated bacterial infections.
Methods
Bacterial strains
Nine isolates of XDRAB were derived from clinical specimen (septum, endotracheal aspirate or respiratory lavage fluid) of patients from the critical care units, geriatrics, internal medicine and emergency department at the First Affiliated Hospital of Chengdu Medical College in 2018 to 2019 (Chengdu, Sichuan, China). These isolates were identified by standard laboratory methods and ATB New (bioMérieux, France) and also were further verified by PCR of two genes, 16S rRNA (with primers 5′-CATTATCACGGTAATTAGTG-3′ and 5′-AGAGCACTGTGCACTTAAG-3′) and blaOXA-51 (with primers 5′-TAATGCTTTGATCGGCCTTG-3′ and 5′-TGGATTGCACTTCATCTTGG-3′)42,43. Staphylococcus aureus ATCC29213 and Escherichia coli ATCC25922 used as quality control strains in antimicrobial susceptibility testing were obtained from the American Type Culture Collection (USA).
Drugs and reagents
A wide range of clinically-used antibacterial agents including ampicillin, ampicillin-sulbactam (2:1), cefepime, ceftazidime, doxycycline, minocycline, amikacin, gentamicin, ciprofloxacin, levofloxacin, chloramphenicol, imipenem, meropenem, tigecycline, polymyxin B, azithromycin and rifampicin were purchased from Meilun Biological (Dalian, Liaoning, China). Zinc lactate, stannous fluoride and furanone were obtained, respectively, from Yuanye Bio-Technology (Shanghai, China), Macklin Biochemical (Shanghai), and Sigma-Aldrich (St. Louis, Missouri, USA). Bacterial culture media, Muller-Hinton broth (MHB), cation-adjusted MH broth (CAMHB), and tryptic soy broth (TSB) medium were purchased from Haibo Biotechnology (Qingdao, Shandong, China).
Antimicrobial susceptibility testing
The minimal inhibitory concentrations (MICs) of antibacterial agents and BFIs for A. baumannii were determined by the broth microdilution method according to the guidelines from the CLSI44. The testing was independently carried out at least twice. Briefly, bacterial cells were inoculated in Luria–Bertani agar medium at 37 °C for 16–20 h, and then resuspended in saline (0.9% sodium chloride) to produce 0.5 McFarland turbidity standard, followed by a 20-fold dilution. Antimicrobial solutions were prepared according to the CLSI44. Zinc lactate and stannous fluoride were prepared in phosphate buffered solution (PBS). Furanone was initially dissolved in dimethyl sulfoxide (50%) and then diluted in PBS (the presence of dimethyl sulfoxide at the assay conditions [≤ 1.6%] was not found to affect A. baumannii growth or their biofilm formation). Together, 180 μl CAMHB, 10 μl of a serially diluted antimicrobial agent, and 10 μl of new prepared bacterial suspension solution were add into 96-well plate. The value of OD600 was measured after incubation at 37 °C for 16–20 h. Less than 0.10 is considered as sterile growth or the colored drugs are observed with the naked eye for the determination of MICs44,45.
Sub-inhibitory concentrations of the biofilm inhibitors
Using the 96-well cell culture plate, 170 μl of TSB medium was added to each well, followed by the addition of 10 μl of PBS (control group) or BFI solution (treatment group) and 20 μl of bacterial suspension solution (at OD600 value of 0.12). Six duplicates were set for each strain. The value of OD600 was measured after incubation at 37 °C every 6 h. When the concentrations of BFIs were used at 1 × MIC, 1/2 MIC, 1/4 MIC, and 1/8 MIC, the growth curves of nine isolates of A. baumannii within 24 h were plotted respectively, and the sub-inhibitory concentrations that did not affect the bacterial growth was finally selected as the working concentration for subsequent experiments46.
Bacterial biofilm formation assay under sub-inhibitory concentrations of the biofilm inhibitors
The biofilm formation assay was performed (with six duplicates for each isolate) using 96-well cell culture plate model with crystal violet staining47. The assay was repeated independently twice. Briefly, 170 μl of TSB medium and 10 μl of PBS (control group) or BFI solution (treatment group) were added into each well, followed by the inoculation of 20 μl of bacterial suspension (OD600 of 0.12). After incubating at 37 °C for 24 h, non-adherent bacterial cells were removed by washing three times with 200 μl of PBS and dried in air. The remaining adherent bacterial cells were stained by adding 200 μl of 0.1% crystal violet. Following incubation of the plates at room temperature for 15 min, the plates were washed three times with PBS and then dried in air. Subsequently, 200 μl of 95% ethanol was added to each well for 5 min and the absorbance of the decolorization solution from each well was measured at the wavelength of 570 nm.
Combination antimicrobial susceptibility testing
The synergistic antibacterial effects of each of the five BFIs with one of the four anti-A. baumannii antibiotics on nine XDRAB isolates were evaluated by microdilution checkerboard method for determining FICI values. The highest concentration of each agent was 2 times its MIC with twofold sequential dilutions, and 8 concentration gradients were tested. Briefly, 170 μl of MHB, 10 μl of each of the series diluents of agents A and B, and 10 μl of freshly prepared bacterial suspension solution were added into the 96-well plate. The checkerboard consisted of columns in which each well contained the same amount of the drug being diluted along the x-axis, and rows in which each well contained the same amount of the drug being diluted on the y-axis, which resulted in each well of a checkerboard containing a unique combination of the two agents being tested. The results were visualized by incubation at 37 °C for 16–20 h. The FICI values were determined for two agents (A and B) by the equation: FICI = A (MICcombined /MICalone) + B (MICcombined/MICalone), with the interpretive criteria as: FICI ≤ 0.5 for synergy, 0.5 < FICI ≤ 1 for additivity, 1 < FICI ≤ 2 for indifference, and FICI > 2 for antagonism48,49.
References
Peleg, A. Y., Seifert, H. & Paterson, D. L. Acinetobacter baumannii: emergence of a successful pathogen. Clin. Microbiol. Rev. 21, 538–582. https://doi.org/10.1128/CMR.00058-07 (2008).
Ling, B.-D., Zhang, L. & Li, X.-Z. Antimicrobial resistance and drug efflux pumps. In Acinetobacter in Efflux-Mediated Antimicrobial Resistance in Bacteria: Mechanisms, Regulation and Clinical Implications (eds Li, X.-Z. et al.) 329–358 (Springer, Berlin, 2016).
Tacconelli, E. et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. https://doi.org/10.1016/S1473-3099(17)30753-3 (2018).
Blackledge, M. S., Worthington, R. J. & Melander, C. Biologically inspired strategies for combating bacterial biofilms. Curr. .Opin Pharmacol. 13, 699–706. https://doi.org/10.1016/j.coph.2013.07.004 (2013).
Badave, G. K. & Kulkarni, D. Biofilm producing multidrug resistant Acinetobacter baumannii: an emerging challenge. J. Clin. Diagn. Res. 9, DC08-10. https://doi.org/10.7860/JCDR/2015/11014.5398 (2015).
Sanchez, C. J. Jr. et al. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 13, 47. https://doi.org/10.1186/1471-2334-13-47 (2013).
Henig, O. et al. The impact of multidrug-resistant organisms on outcomes in patients with diabetic foot infections. Open Forum Infect. Dis. 7, ofaa161. https://doi.org/10.1093/ofid/ofaa161 (2020).
Donlan, R. M. & Costerton, J. W. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 15, 167–193. https://doi.org/10.1128/cmr.15.2.167-193.2002 (2002).
Hall, C. W. & Mah, T. F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 41, 276–301. https://doi.org/10.1093/femsre/fux010 (2017).
Algburi, A., Comito, N., Kashtanov, D., Dicks, L. M. T. & Chikindas, M. L. Control of biofilm formation: antibiotics and beyond. Appl. Environ. Microbiol. https://doi.org/10.1128/AEM.02508-16 (2017).
Spellberg, B. & Bonomo, R. A. Combination therapy for extreme drug-resistant Acinetobacter baumannii: ready for prime time?. Crit .Care Med. 43, 1332–1334. https://doi.org/10.1097/CCM.0000000000001029 (2015).
Ledder, R. G., Sreenivasan, P. K., DeVizio, W. & McBain, A. J. Evaluation of the specificity and effectiveness of selected oral hygiene actives in salivary biofilm microcosms. J. Med. Microbiol. 59, 1462–1468. https://doi.org/10.1099/jmm.0.024372-0 (2010).
Yu, S., Zhang, M. & Ma, L. Anti-biofilm effects of Zn lactate·3H2O and SnF2 on Pseudomonas aeruginosa, Acinetobacter baumannii and Streptococcus mutans. Chin. J. Biotechnol. 33, 1478–1488. https://doi.org/10.13345/j.cjb.170141 (2017).
Rabin, N. et al. Agents that inhibit bacterial biofilm formation. Future Med. Chem. 7, 647–671. https://doi.org/10.4155/fmc.15.7 (2015).
Song, J. Y., Cheong, H. J., Noh, J. Y. & Kim, W. J. In vitro comparison of anti-biofilm effects against carbapenem-resistant Acinetobacter baumannii: imipenem, colistin, tigecycline, rifampicin and combinations. Infect. Chemother. 47, 27–32. https://doi.org/10.3947/ic.2015.47.1.27 (2015).
Brochado, A. R. et al. Species-specific activity of antibacterial drug combinations. Nature 559, 259–263. https://doi.org/10.1038/s41586-018-0278-9 (2018).
Seleem, N. M., Abd El Latif, H. K., Shaldam, M. A. & El-Ganiny, A. Drugs with new lease of life as quorum sensing inhibitors: for combating MDR Acinetobacter baumannii infections. Eur. J. Clin. Microbiol. Infect. Dis. https://doi.org/10.1007/s10096-020-03882-z (2020).
CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed. CLSI supplement M100. (Wayne, Pennsylvania: Clinical and Laboratory Standards Institute, 2020).
Magiorakos, A. P. et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. https://doi.org/10.1111/j.1469-0691.2011.03570.x (2012).
Bae, S. et al. In vitro synergistic activity of antimicrobial agents in combination against clinical isolates of colistin-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 60, 6774–6779. https://doi.org/10.1128/AAC.00839-16 (2016).
Pimchan, T., Maensiri, D. & Eumkeb, G. Synergy and mechanism of action of alpha-mangostin and ceftazidime against ceftazidime-resistant Acinetobacter baumannii. Lett. Appl. Microbiol. 65, 285–291. https://doi.org/10.1111/lam.12789 (2017).
Eze, E. C., Chenia, H. Y. & El Zowalaty, M. E. Acinetobacter baumannii biofilms: effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 11, 2277–2299. https://doi.org/10.2147/IDR.S169894 (2018).
Mahamuni-Badiger, P. P. et al. Biofilm formation to inhibition: role of zinc oxide-based nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 108, 110319. https://doi.org/10.1016/j.msec.2019.110319 (2020).
Mayer, C. et al. Multiple quorum quenching enzymes are active in the nosocomial pathogen Acinetobacter baumannii ATCC17978. Front. Cell Infect. Microbiol. 8, 310. https://doi.org/10.3389/fcimb.2018.00310 (2018).
Imperi, F., Leoni, L. & Visca, P. Antivirulence activity of azithromycin in Pseudomonas aeruginosa. Front. Microbiol. 5, 178. https://doi.org/10.3389/fmicb.2014.00178 (2014).
Bogdan, M. et al. In vitro effect of subminimal inhibitory concentrations of antibiotics on the biofilm formation ability of Acinetobacter baumannii clinical isolates. J. Chemother. 30, 16–24. https://doi.org/10.1080/1120009X.2017.1378835 (2018).
Nordqvist, H., Nilsson, L. E. & Claesson, C. Mutant prevention concentration of colistin alone and in combination with rifampicin for multidrug-resistant Acinetobacter baumannii. Eur. J. Clin. Microbiol. Infect. Dis. 35, 1845–1850. https://doi.org/10.1007/s10096-016-2736-3 (2016).
Majewski, P. et al. In vitro activity of rifampicin alone and in combination with imipenem against multidrug-resistant Acinetobacter baumannii harboring the blaOXA-72 resistance gene. Scand. J. Infect. Dis. 46, 260–264. https://doi.org/10.3109/00365548.2013.865141 (2014).
Bai, Y. et al. In vitro activities of combinations of rifampin with other antimicrobials against multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 59, 1466–1471. https://doi.org/10.1128/AAC.04089-14 (2015).
Wareham, D. W. & Bean, D. C. In-vitro activity of polymyxin B in combination with imipenem, rifampicin and azithromycin versus multidrug resistant strains of Acinetobacter baumannii producing OXA-23 carbapenemases. Ann. Clin. Microbiol. Antimicrob. 5, 10. https://doi.org/10.1186/1476-0711-5-10 (2006).
Pachon-Ibanez, M. E. et al. Efficacy of rifampin and its combinations with imipenem, sulbactam, and colistin in experimental models of infection caused by imipenem-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 54, 1165–1172. https://doi.org/10.1128/AAC.00367-09 (2010).
Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 56, 395–411 (1992).
Baron, S., Hadjadj, L., Rolain, J. M. & Olaitan, A. O. Molecular mechanisms of polymyxin resistance: knowns and unknowns. Int. J. Antimicrob. Agents 48, 583–591. https://doi.org/10.1016/j.ijantimicag.2016.06.023 (2016).
Kadar, B. et al. In vitro activity of clarithromycin in combination with other antimicrobial agents against biofilm-forming Pseudomonas aeruginosa strains. Acta Microbiol. Immunol. Hung. 57, 235–245. https://doi.org/10.1556/AMicr.57.2010.3.8 (2010).
Li, X.-Z., Plesiat, P. & Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 28, 337–418. https://doi.org/10.1128/CMR.00117-14 (2015).
Lepe, J. A., Garcia-Cabrera, E., Gil-Navarro, M. V. & Aznar, J. Rifampin breakpoint for Acinetobacter baumannii based on pharmacokinetic-pharmacodynamic models with Monte Carlo simulation. Rev. Esp. Quimioter. 25, 134–138 (2012).
Viehman, J. A., Nguyen, M. H. & Doi, Y. Treatment options for carbapenem-resistant and extensively drug-resistant Acinetobacter baumannii infections. Drugs 74, 1315–1333. https://doi.org/10.1007/s40265-014-0267-8 (2014).
Aydemir, H. et al. Colistin vs. the combination of colistin and rifampicin for the treatment of carbapenem-resistant Acinetobacter baumannii ventilator-associated pneumonia. Epidemiol. Infect. 141, 1214–1222. https://doi.org/10.1017/S095026881200194X (2013).
Srisilapanan, P., Roseman, J. & Likitsatian, T. Clinical effect of toothpaste and mouth rinse containing zinc lactate on oral malodor reduction. J. Clin. Exp. Dent. 11, e346–e352. https://doi.org/10.4317/jced.55418 (2019).
Marinho, V. C., Chong, L. Y., Worthington, H. V. & Walsh, T. Fluoride mouthrinses for preventing dental caries in children and adolescents. Cochrane Database Syst. Rev. 7, CD00284. https://doi.org/10.1002/14651858.CD002284.pub2 (2016).
Davis, S. C., Pissani, F. & Montero, R. B. Effects of commonly used topical antimicrobial agents on Acinetobacter baumannii: an in vitro study. Mil. Med. 173, 74–78. https://doi.org/10.7205/milmed.173.1.74 (2008).
Chang, Y. et al. Characterization of carbapenem-resistant Acinetobacter baumannii isolates in a Chinese teaching hospital. Front. Microbiol. 6, 910. https://doi.org/10.3389/fmicb.2015.00910 (2015).
Lin, F. et al. Molecular characterization of reduced susceptibility to biocides in clinical Isolates of Acinetobacter baumannii. Front. Microbiol. 8, 1836. https://doi.org/10.3389/fmicb.2017.01836 (2017).
CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed. CLSI supplement M100. (Wayne, Pennsylvania: Clinical and Laboratory Standards Institute, 2018).
Zhang, C. et al. A study on the antibacterial effect of Chinese medicine plumbagin on tigecycline-resistant Acinetobacter baumannii. J. Chengdu Med. Coll. 12, 6–10. https://doi.org/10.3969/j.issn.1674-2257.2017.02.001 (2017).
Aranda, J. et al. Acinetobacter baumannii RecA protein in repair of DNA damage, antimicrobial resistance, general stress response, and virulence. J. Bacteriol. 193, 3740–3747. https://doi.org/10.1128/JB.00389-11 (2011).
Selasi, G. N. et al. Differences in biofilm mass, expression of biofilm-associated genes, and resistance to desiccation between epidemic and sporadic clones of carbapenem-resistant Acinetobacter baumannii sequence type 191. PLoS ONE 11, e0162576. https://doi.org/10.1371/journal.pone.0162576 (2016).
Hsieh, M. H., Yu, C. M., Yu, V. L. & Chow, J. W. Synergy assessed by checkerboard. A critical analysis. Diagn. Microbiol. Infect. Dis. 16, 343–349. https://doi.org/10.1016/0732-8893(93)90087-n (1993).
Zuo, G. Y. et al. Synergistic antibacterial and antibiotic effects of bisbenzylisoquinoline alkaloids on clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). Molecules 16, 9819–9826. https://doi.org/10.3390/molecules16129819 (2011).
Marchaim, D. et al. Major variation in MICs of tigecycline in Gram-negative bacilli as a function of testing method. J. Clin. Microbiol. 52, 1617–1621. https://doi.org/10.1128/JCM.00001-14 (2014).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 81373454).
Author information
Authors and Affiliations
Contributions
Q.P. designed and performed the experiments, and drafted the manuscript; F.L. contributed to study design and revising the manuscript; B.L. initiated and supervised the study, and revised the manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peng, Q., Lin, F. & Ling, B. In vitro activity of biofilm inhibitors in combination with antibacterial drugs against extensively drug-resistant Acinetobacter baumannii. Sci Rep 10, 18097 (2020). https://doi.org/10.1038/s41598-020-75218-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-75218-y
- Springer Nature Limited