
Abstract
Epistasis plays an important role in manipulating rice tiller number, but epistatic mechanism still remains a challenge. Here we showed the process of epistatic analysis between tillering QTLs. A half diallel mating scheme was conducted based on 6 single segment substitution lines and 9 dual segment pyramiding lines to allow the analysis of 4 epistatic components. Additive-additive, additive-dominance, dominance-additive, and dominance-dominance epistatic effects were estimated at 9 stages of development via unconditional QTL analysis simultaneously. Unconditional QTL effect (QTL cumulative effect before a certain stage) was then divided into several conditional QTL components (QTL net effect in a certain time interval). The results indicated that epistatic interaction was prevalent, all QTL pairs harboring epistasis and one QTL always interacting with other QTLs in various component ways. Epistatic effects were dynamic, occurring mostly within 14d and 21–35d after transplant and exhibited mainly negative effects. The genetic and developmental mechanism on several tillering QTLs was further realized and perhaps was useful for molecular pyramiding breeding and heterosis utilization for improving plant architecture.
Similar content being viewed by others
Introduction
Breeding for ideotypes implies to take qualified morphological and physiological characteristics to concentrate on one plant so as to obtain the highest utilization rate and transformation rate of light energy1,2,3. Tiller number in cereal is not only a plant type trait but also one of the important agronomic traits for grain production, and moreover a model trait for the study of developmental behaviors4,5. Therefore, the understanding tillering genetic mechanism will address the fundamental issues of plant science and facilitate the breeding of high-yielding crop varieties.
Using molecular marker maps and QTL mapping technology, mapping of QTLs for tiller number has been extensively conducted6,7,8,9. Two common methods were used for developmental traits, one being by analyzing the performance of a trait observed at a fixed stage of ontogenesis to estimate QTL accumulated effects from the beginning to the investigation stage, and the other being analyzing successively the observations at various developmental stages to reveal QTL expression dynamics8. However net expression of QTLs at a certain stage kept mysterious.
Understanding gene expression is one of the major goals in developmental genetics. A separate analysis of data derived from times t-1 and t can provide inferences for the cumulative gene effects from the initial time to t-1 or t but not for the net effect of gene expression in the period t-1 to t10. A conditional analysis method was proposed to estimate the extra genetic effect from t-1 to t10,11, and QTL analysis on the extra genetic effect (named conditional QTL analysis) had widely been applied12,13,14,15. Conditional QTL reflected the net expression of QTL at a certain time. However traditional QTL mapping can not obtain an unbiased estimation of QTLs under the conventional mapping populations such as F2, RILs, or DHLs.
It has been long to recognize the advantages of near isogenic lines or single segment substitution lines for QTL identification16,17,18. Based on single segment substitution lines numerous QTLs were identified19,20,21,22,23,24,25,26,27,28,29,30,31,32. However, the important genetic component of epistasis was often ignored or incomprehensive. Japanese rice genome projects (JRGP) and Guangdong Key Lab of Plant Molecular Breeding provided typical cases to analyze epistatic interactions between QTLs, in which four types of epistatic components were effectively estimated on heading date in rice via secondary mapping populations33,34,35,36,37. In this paper, we applied previous cultured materials to estimate epistasis between QTLs on tiller number via adopting both the unconditional and conditional analysis methods. The aims were to further confirm the prevalence of epistatic interactions and deeply understand genetic and developmental mechanisms on several tillering QTLs so as to excavate useful alleles for molecular pyramiding breeding and heterosis utilization for improving plant architecture.
Results
Identification of tillering QTLs
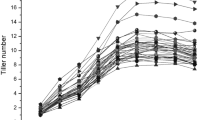
Tiller number appeared “S” type of growth curve, after slow, rapid growth it reaching the peak and then decreasing slightly (Supplementary Figure S1). Analysis of variance at various stages revealed the significant difference of tiller numbers existed among genotypes (Supplementary Table S1). This information supported the existence of tillering QTLs. The contrast test indicated that the 6 SSSLs really harbored tillering QTLs (Table 1). All carried with heterozygote effects, and S2 with additional homozygote effects. That QTLs appeared repeatedly at several tested stages guaranteed the truth of putative QTLs, but inconsistent estimations throughout stages exhibited dynamics of QTL expressions. The effects of QTLs were mainly to enhance tillering and appeared mostly before t3. It was suggested that utilization of these QTLs on heterosis and in developmental early stages.
Estimations of epistatic effects
The epistatic effect was generally estimated as the deviation between the dual-SSSL pyramiding effect and the sum of two single-SSSL effects. Estimations of four epistatic components, additive-additive (aa), additive-dominance (ad), dominance-additive (da), and dominance-dominance (dd) for nine pairs of QTLs confirmed epistatic prevalence (Table 2). All pairs of Si/Sj held interaction effects, involving in one or more epistatic components and exhibiting at least two stages. Interactions in dd component, with negative effects and at early stages were in the majority, which closely related to d component, positive effects and early development of single QTL. Consistent function directions appeared among epistatic components and across stages. However, epistasis changed with developmental stages, reflecting dynamics of it. Additionally, one QTL always interacted with multiple other QTLs in various component ways.
Conditional QTL analysis
Unconditional QTL analysis mentioned above indicated that tillering QTLs changed dynamically (Tables 1, 2). Conditional QTLs revealed expression quantities of tillering QTLs in various time intervals (Table 3). For instance, the cumulative heterozygote effect of S1 was 1.50* at t3 (Table 1), which was divided into three components, 0.78*, − 0.26 and 0.50, representing the net expressions in t0–t1, t1–t2 and t2–t3, respectively (Table 3). Similarly, the cumulative ad component of S1/S2 was − 3.91** at t3 (Table 2), which expressed − 0.58, − 2.52** and 0.31 in t0–t1, t1–t2 and t2–t3, respectively (Table 3). It was indicated that QTL effects estimated at t(n) were in fact the accumulation of expression quantities in various time intervals before t(n). Expression quantities provided information about both action periods and dynamic patterns of QTLs. QTLs expressed homozygote effects mainly concentrating on t5–t6, heterozygote effects in t0–t1 and t3–t6, while epistasis in t0–t2 and t3–t5. Great differences took place among epistatic components and developmental periods. Negative expressions were main, occupied up 66.7% of all significant effects, where negative heterozygote effects and negative epistatic effects hold 47.1% and 75.7%, respectively. Inverse correlation between heterozygote effects and epistatic effects was detected in t0–t2 and t3–t5, where positive (negative) d resulting in negative (positive) e in general. Lots of expressions were feeble so that fail to be detected, and large expressions became invisible for the reason of error ascending.
Discussion
Tiller number related to plant architecture and yield components. Generally, tiller number was investigated at the maximum tillering stage and was then analyzed. This however didn’t provide ample information about QTLs. QTLs detected at t6 (the maximum tillering stage, see Supplementary Figure S1) were significantly less than those at t2 or t3 (Tables 1, 2). It is suggested that an alternative strategy to explore tillering QTLs concentrating in 14–21d after transplant. QTL analysis of time-specific measures possessed great advantages, providing comprehensive and convincing QTL information (Tables 1, 2). Especially time-specific measures allowed to excavate QTL dynamics. Conditional QTL analysis estimated net expression quantities of tillering QTLs in various developmental time intervals (Table 3), solving the puzzle of gene dynamic behavior. Since SSSLs harbored tillering QTLs mostly with positive homozygote effects and heterozygote effects (Table 1), negative epistatic effects were detected between many SSSL pairs (Table 2). QTLs expressed selectively in certain time intervals, homozygote effects appearing mostly in 35–42d, heterozygote effects in 0–7d and 21–42d, and epistasis in 0–7d and 21–35d after transplant (Table 3).
Unconditional QTL and conditional QTL analysis produced approximate consistent results (Tables 1–3). In theory, the unconditional QTL effect is the accumulation of several conditional QTL components10. Generally, QTL effects at time tn are the sum of QTL effect at time tn-1 and the extra effect Gd. Gd was ever estimated by the deviation of QTL effects between tn and tn-1, but it couldn’t provide the net effects of gene expression in the period tn-1 to tn since Gd correlated with QTL effect at tn-110,11. The conditional QTL effect was as the unbiased estimation of Gd, namely \(QTL_{{t_{n} }} = QTL_{{t_{n - 1} }} + QTL_{{t_{n} |t_{n - 1} }}\) since \(QTL_{{t_{n} |t_{n - 1} }}\) was independent of \(QTL_{{t_{n - 1} }}\)12,13,14,15. The formula could be extended easily as \(QTL_{{t_{n} }} = QTL_{{t_{1} }} + QTL_{{t_{2} |t_{1} }} + QTL_{{t_{3} |t_{2} }} + \cdots + QTL_{{t_{n} |t_{n - 1} }}\). The relationship was well validated by estimated results (Supplementary Table S2), in which the correlation coefficient between \(QTL_{{t_{9} }}\) effects and the sums of all conditional QTL effects before t9 reached 0.9379**. Furthermore, only did conditional QTL variations reflected dynamic expressions of a QTL throughout the whole developmental stage.
In multiple gene system, gene interaction is inevitable except for gene accumulative effects11. A half diallel mating design allowed to estimate various components simultaneously, time-specific measures provided repeated testing, and conditional analysis revealed net expressions of epistasis (Table 2). All SSSL pairs involved in dd and another epistatic component at least, with 90.6% negative estimations and 65.6% of them detected in 21d after transplant (Table 2). QTLs expressed epistasis mostly with negative effects (occupied 75.7%) and mainly in 14d and 21–35d after transplant (Table 3). These epistatic features attributed to the inverse correlation between heterozygote effect and epistasis. Additionally, expression quantity differences exhibited abundantly the dynamics of epistasis with the development of tillers.
Tillering QTLs were widely located via traditional mapping populations such as F2, DHLs and RILs etc6,7,8,9. However, since both the materials and markers applied in these studies differed, it was difficult to compare the locations of existing QTLs on tiller number. Our lab was engaged in the study of QTLs using the population of single segment substitution lines for a long time, it was possible to detect the same chromosome regions for different QTLs. In 2008, we identified 18 QTLs on panicle number in rice using SSSLs38, and then nine of them were analyzed for their dynamic expression in 200930. Similarly, 14 tillering QTLs were detected with a single segment substitution population of 26 lines by unconditional QTL mapping method, and their net expressions were estimated within time intervals by conditional QTL mapping simultaneously in 201031, and then twelve of them were analyzed for their dynamic expression in different planting seasons and under different cropping densities in 201232. These studies excavated tillering QTLs and showed dynamic expressions for some of them on additive, dominance and dominance-dominance epistatic component, but lacking the components of additive-additive, additive-dominance and dominance-additive epistatic components since the limitation of pyramiding line materials. In 2018 based on eight single segment substitution lines, each of which was tested with heading QTLs, a half diallel crossing population was constructed to allow estimating simultaneously the four epistatic components of QTLs37. Using this population, six single segment substitution lines were tested to be with tillering QTLs, and then their dynamic expressions of the four epistatic components at various developmental stages were explored in this paper. Comparing with the previous studies, the chromosome interval of S1 in this paper might be consistent with SSSL 23 in 2008, S2 with SSSL 14 in 2008, Tn6-2, Tn6-3 and Tn6-4 in 2010, and S3 with SSSLs 3, 14, 22 in 2008, Tn6-2, Tn6-3, Tn6-4 in 2010 (Supplementary Figure S2) .
Methods
Plant materials
The similar plant materials as the previous experiment were applied in this trial37 (Table 4). Huajingxian 74 (HJX74) is an elite indica variety with many excellent properties cultured by our laboratory from South China. SSSLs possessed only a single substituted segment from donors under HJX74 genetic background, and harbored putative QTL/gene on heading date37. Double segment substitution lines (DSSLs) were cultivated based on the F2 populations from the crossing of two SSSLs. Together with of 6 SSSLs, a total of nine DSSLs were selected as parental materials (Table 2), Some of DSSLs were lacking since their seeds of F1 weren’t achieved. And then the experimental population was compounded via a half diallel mating scheme37. The population in total included 49 materials, HJX74, 6 SSSLs, 9 DSSLs and 33 crossing combinations among them.
Field experiments
The same phenotypic trial as the previous study was applied in this trial37. The trial site located at the teaching and experiment station of South China Agricultural University, in Guangzhou, China (23°79ʹ N, 113°159ʹ E). In the early season (duration from March to July) of 2018, 49 materials were grown in a completely randomized block design with three replications. Germinated seeds were sowed in a seedling bed, and then seedlings were transplanted to a rice field 20 days later with one plant per hill and the density of 16.7 cm \(\times\) 16.7 cm. A plot consisted of four rows, ten plants per row. Local standard practices were used for the trails. Tiller numbers per hill on 10 central plants were investigated in each plot from seven days after transplanting onwards, data every 7 days once was continuously recorded nine weeks (denoted by t1 to t9). Averages over ten plants in each plot were as inputting data for statistical analysis.
Statistical analysis and estimation of QTL effects
The model \(y_{ij} = \mu + G_{i} + B_{j} + e_{ij}\) was adopted to variance analysis on data at each stage, where \(y,\mu ,G,B\) and \(e\) were the observations each plot, the population mean, genotype, block and error effect, respectively. i, j represented serial numbers of genotypes and blocks, respectively. Homozygote effect (a) or heterozygote effect (d) of QTL was estimated by \((S_{i} - HJX74)\), where \(S_{i}\) represented homozygote or heterozygote of SSSLi. Epistatic effect between QTLs (e) was estimated by \((D_{kl} + HJX74 - S_{k} - S_{l} )\), where \(D_{kl} ,S_{k} ,S_{l}\) indicated dual segment and its two single segment materials, respectively, which might be homozygotes or heterozygotes. Four epistatic components could be estimated by,
where \(aa,ad,da,dd\) indicated additive-additive, additive-dominance, dominance-additive and dominance-dominance epistatic components, respectively. \(k = i, \, i^{\prime}, \, l = j, \, j^{\prime}\), i and i′, j and j′ represented homozygotes and heterozygotes of substitution segments. Statistics \(t_{\alpha } \sqrt {\frac{{2S_{e}^{2} }}{n}}\) and \(t_{\alpha } \sqrt {\frac{{4S_{e}^{2} }}{n}}\) were adopted to test of significance for a/d and e, respectively. Where \(t_{\alpha }\), \(S_{e}^{2}\), \(n\) indicated as critical t-value under \(\alpha\) probability level and error freedom degree, error mean square and replication numbers, respectively. Formula \(y_{t|t - 1} = y_{t} - b_{t/t - 1} (y_{t - 1} - \overline{y}_{t - 1} )\) was used to estimate conditional variable \(y_{t|t - 1}\), indicating phenotypic values at time t conditional on the phenotypic value at given time t-1, where \(y_{t - 1} {\text{ and }} \overline{y}_{t - 1} , \, y_{t} {\text{ and }} \overline{y}_{t}\) were the phenotypic values and means at time t-1 and time t, respectively. \(b_{t/t - 1}\) was the regression coefficient for phenotypic values at time t versus time t-1. Statistical analysis was imposed on the conditional variable \(y_{t|t - 1}\) to generate conditional QTLs10.
Statistical analysis and estimation of QTL effects were carried out with aov() and lm() functions in R language (https://www.r-project.org/).
References
Wang, Y. H. & Li, J. Y. Genes controlling plant architecture. Curr. Opin. Biotechnol. 17, 1–7 (2006).
Peng, S. B., Khush, G. S., Virk, P. D., Tang, Q. Y. & Zou, Y. B. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 108, 32–38 (2008).
Markel, K., Belcher, M. S. & Shih, P. M. Defining and engineering bioenergy plant feedstock ideotypes. Curr. Opin. Biotechnol. 62, 196–201 (2020).
Liang, W. H., Shang, F., Lin, Q. T., Lou, C. & Zhang, J. Tillering and panicle branching genes in rice. Gene 537, 1–5 (2014).
Shao, G. N. et al. Tiller bud formation regulators MOC1 and MOC3 cooperatively promote tiller bud outgrowth by activating FON1 expression in rice. Mol. Plant 12, 1090–1102 (2019).
Lei, L. et al. Genetic dissection of rice (Oryza sativa L) tiller, plant height, and grain yield based on QTL mapping and meta analysis. Euphytica 214, 109–113 (2018).
Li, Z. B. et al. Quantitative trait locus mapping for yield-associated agronomic traits in a BC2F6 population of Japonica hybrid rice Liaoyou 5218. J. Plant Growth Regul. 39, 60–71 (2020).
Wu, W. R., Li, W. M., Tang, D. Z., Lu, H. R. & Worland, A. J. Time-related mapping of quantitative trait loci underlying tiller number in rice. Genetics 151, 297–303 (1999).
Liu, G. F., Li, M., Wen, J., Du, Y. & Zhang, Y. M. Functional mapping of quantitative trait loci associated with rice tillering. Mol. Genet. Genom. 284, 263–271 (2010).
Zhu, J. Analysis of conditional genetic effects and variance components in developmental genetics. Genetics 141, 1633–1639 (1995).
Atchley, W. R. & Zhu, J. Developmental quantitative genetics, conditional epigenetic variability and growth in mice. Genetics 147, 765–776 (1997).
Yan, J. Q., Zhu, J., He, C. X., Benmoussa, M. & Wu, P. Quantitative trait loci analysis for the developmental behavior of tiller number in rice (Oryza sativa L.). Theor. Appl. Genet. 97, 267–274 (1998).
Yan, J. Q., Zhu, J., He, C. X., Benmoussa, M. & Wu, P. Molecular dissection of developmental behavior of plant height in rice (Oryza sativa L.). Genetics 150, 1257–1265 (1998).
Cao, G. C. et al. Impact of epistasis and QTL×environment interaction on the developmental behavior of plant height in rice (Oryza sativa L.). Theor. Appl. Genet. 103, 153–160 (2001).
Liu, G. F., Yang, J., Xu, H. M., Hayat, Y. A. & Zhu, J. Genetic analysis of grain yield conditioned on its component traits in rice (Oryza sativa L.). Aust. J. Agric. Res. 59, 189–195 (2008).
Tanksley, S. Mapping polygenes. Annu. Rev. Genet. 27, 205–233 (1993).
Eshed, Y. & Zamir, D. An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 141, 1147–1162 (1995).
Eshed, Y. & Zamir, D. Less-than-additive epistatic interactions of quantitative trait loci in tomato. Genetics 143, 1807–1817 (1996).
Wang, S. K. et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 44(8), 950–954 (2012).
Wang, S. K. et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat. Genet. 47(8), 949–954 (2015).
Zhu, H. T. et al. Detection and characterization of epistasis between QTLs on plant height in rice using single segment substitution lines. Breed. Sci. 65(3), 192–200 (2015).
Zhu, H. T. et al. Analysis of QTLs on heading date based on single segment substitution lines in rice (Oryza Sativa L.). Sci. Rep. 8, 13232 (2018).
Dai, Z. J. et al. Development of a platform for breeding by design of CMS lines based on an SSSL library in rice (Oryza sativa L.). Euphytica 205(1), 63–72 (2015).
Dai, Z. J. et al. Development of a platform for breeding by design of CMS restorer lines based on an SSSL library in rice (Oryza sativa L.). Breed. Sci. 66(5), 768–775 (2016).
Guo, J. et al. Overcoming inter-subspecific hybrid sterility in rice by developing indica-compatible japonica lines. Sci. Rep. 6(1), 26878 (2016).
He, N. et al. Development and trait evaluation of chromosome single segment substitution lines of O. meridionalis in the background of O. sativa. Euphytica 213, 281–296 (2017).
Zhou, Y. L. et al. Substitution mapping of QTLs controlling seed dormancy using single segment substitution lines derived from multiple cultivated rice donors in seven cropping seasons. Theor. Appl. Genet. 130(6), 1191–1205 (2017).
Wang, X. L. et al. QTL epistatic analysis for yield components with single-segment substitution lines in rice. Plant Breed. 137(3), 346–354 (2018).
Zhao, H. et al. Genetic characterization of the chromosome single-segment substitution lines of O. glumaepatula and O. barthii and identification of QTLs for yield-related traits. Mol. Breed. 39, 51–70 (2019).
Liu, G. F. et al. Dynamic expression of nine QTLs for tiller number detected with single segment substitution lines in rice. Theor. Appl. Genet. 118(3), 443–453 (2009).
Liu, G. F. et al. Unconditional and conditional QTL mapping for the developmental behavior of tiller number in rice (Oryza sativa L.). Genetica 138(8), 885–893 (2010).
Liu, G. F., Zhu, H. T., Zhang, G. G., Li, L. H. & Ye, G. Y. Dynamic analysis of QTLs on tiller number in rice (Oryza sativa L.) with single segment substitution lines. Theor. Appl. Genet. 125, 143–153 (2012).
Lin, H. X., Yamamoto, T., Sasaki, T. & Yano, M. Characterization and detection of epistatic interactions of 3 QTLs, Hd1, Hd2 and Hd3, controlling heading date in rice using nearly isogenic lines. Theor. Appl. Genet. 101, 1021–1028 (2000).
Yamamoto, T., Lin, H. X., Sasaki, T. & Yano, M. Identifcation of heading date quantitative trait locus Hd6 and characterization of its epistatic interactions with Hd2 in rice using advanced backcross progeny. Genetics 154, 885–891 (2000).
Chen, J. B. et al. Characterization of epistatic interaction of QTLs LH8 and EH3 controlling heading date in rice. Sci. Rep. 4(1), 4263 (2014).
Qin, M. et al. Bigenic epistasis between QTLs for heading date in rice analyzed using single segment substitution lines. Field Crops Res. 178, 16–25 (2015).
Yang, Z. F. et al. Analysis of epistasis among QTLs on heading date based on single segment substitution lines in rice. Sci. Rep. 8, 3059 (2018).
Liu, G. F. et al. Detection of QTLs with additive effects and additive-by-environment interaction effects on panicle number in rice (Oryza sativa L.) with single-segment substitution lines. Theor. Appl. Genet. 116, 923–931 (2008).
Acknowledgements
This research was supported by Guangzhou Science and Technology Planning Project Foundation, China (201707010340), The National Key Research and Development Program of China (2016YFD0100406), Special Project for Leading Talents in Innovation of Science and Technology of Guangdong Province, China (2016TX03N224) and Research and Development Program in Key Areas of Guangdong Province, China (2018B020202004).
Author information
Authors and Affiliations
Contributions
H.Q.Z., W.F.Y. and S.P.M. were the executives of trials and the writers, X.L., H.T.Z., A.M.W., C.L.H., B.R. and S.Z.D. were the participators, L.J.M. was the cooperator, S.K.W. and G.Q.Z. were the leaders of our research team, and G.F.L. was the constitutor of this study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhou, H., Yang, W., Ma, S. et al. Unconditional and conditional analysis of epistasis between tillering QTLs based on single segment substitution lines in rice. Sci Rep 10, 15912 (2020). https://doi.org/10.1038/s41598-020-73047-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-73047-7
- Springer Nature Limited
This article is cited by
-
Pyramiding of Low Chalkiness QTLs Is an Effective Way to Reduce Rice Chalkiness
Rice (2024)
-
QTL epistasis plays a role of homeostasis on heading date in rice
Scientific Reports (2024)
-
Epistatic interaction has the reverse effects with its constitutive quantitative trait loci
Scientific Reports (2024)
-
Dynamic analysis of QTLs on plant height with single segment substitution lines in rice
Scientific Reports (2022)
-
Substitution Mapping of Two Closely Linked QTLs on Chromosome 8 Controlling Grain Chalkiness in Rice
Rice (2021)