
Abstract
While the introduction of herbicide tolerant crops provided growers new options to manage weeds, the widespread adoption of these herbicides increased the risk for herbicide spray drift to surrounding vegetation. The impact of herbicide drift in sensitive crops is extensively investigated, whereas scarce information is available on the consequences of herbicide drift in non-target plants. Weeds are often abundant in field margins and ditches surrounding agricultural landscapes. Repeated herbicide drift exposure to weeds could be detrimental to long-term management as numerous weeds evolved herbicide resistance following recurrent-selection with low herbicide rates. The objective of this study was to evaluate if glyphosate, 2,4-D, and dicamba spray drift could select Amaranthus spp. biotypes with reduced herbicide sensitivity. Palmer amaranth and waterhemp populations were recurrently exposed to herbicide drift in a wind tunnel study over two generations. Seeds from survival plants were used for the subsequent rounds of herbicide drift exposure. Progenies were subjected to herbicide dose-response studies following drift selection. Herbicide drift exposure rapidly selected for Amaranthus spp. biotypes with reduced herbicide sensitivity over two generations. Weed management programs should consider strategies to mitigate near-field spray drift and suppress the establishment of resistance-prone weeds on field borders and ditches in agricultural landscapes.
Similar content being viewed by others
Introduction
Current weed management practices such as herbicide applications tend to focus on property-level management decisions, where actions usually neglect landscape-scale outcomes1. The recurrent herbicide drift complaints in agricultural landscapes evidences this situation. After the introduction of glyphosate tolerant crops in the late nineties, and the recent introduction of 2,4-D and dicamba tolerant crops, growers gained new herbicide options and flexibility to manage weeds2,3,4. However, the widespread adoption of these herbicides in weed management programs increased the risk of off-target movement associated with herbicide applications. Spray drift is the part of the pesticide application deflected away from the target area during or following applications5. Glyphosate, 2,4-D, and dicamba spray drift have been reported to cause severe injury on sensitive vegetation and crops, especially when best practices are not adopted during applications6,7,8,9,10,11,12,13.
While the consequences of herbicide drift towards sensitive crops are extensively investigated in the literature, scarce information is available on the consequences of herbicide drift towards other plant communities surrounding agricultural landscapes14. Weed species such as waterhemp [Amaranthus tuberculatus (Moq.) J. D. Sauer] and Palmer amaranth (Amaranthus palmeri S. Wats.) are often abundant in field margins and ditches throughout the US15,16,17. Herbicide drift exposure could be detrimental to long-term weed management as numerous weed species have evolved herbicide resistance following recurrent applications of low herbicide rates18,19,20,21,22,23,24,25,26,27,28, and we reported in a previous study that spray drift can expose weeds to herbicide doses associated with herbicide resistance selection14. In addition, herbicide resistance has been widely reported in weed populations inhabiting field margins and ditches surrounding agricultural landscapes15,17,29.
Recurrent selection to low herbicide doses can gradually select for metabolism alleles present within the standing genetic variation of the population, which can progressively lead to herbicide resistance on weeds20,30,31,32. Some researchers also suggest that low rates of herbicides could induce new stress-related mutations and epigenetic alterations on weeds, ultimately leading to reduced herbicide sensitivity33,34,35.
Recombination and accumulation of minor resistance alleles can occur at a faster rate in cross-pollinated species, such as Palmer amaranth and waterhemp, during recurrent selection with low rates of herbicides21,22,36. These C4 summer annual Amaranthus spp. are among the most troublesome weed species in the US row crop production systems37. Both are obligate outcrossing dioecious weed species with a fast growth habitat, extended emergence window, and prolific seed production with high genetic plasticity which pose a challenge to their management37,38,39,40,41,42,43,44. Numerous Palmer amaranth and waterhemp populations have evolved resistance to herbicides that target 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), 4-hydroxyphenylpyruvate dioxygenase (HPPD), photosystem II, protoporphyrinogen oxidase (PPO), auxin receptors, microtubule assembly, and acetolacte synthase (ALS) in the US15,17,45,46,47,48,49,50,51,52,53,54. Moreover, pollen mediated gene flow has been reported as a major contributor to herbicide resistance dissemination in Palmer amaranth and waterhemp in the US Midwest55,56.
Although controlling weed populations on field margins and ditches is considered a best management practice to delay herbicide resistance evolution, these weed populations are often neglected in agricultural landscapes15,16,17,29. The hypothesis of this study is that repeated herbicide drift in field borders and ditches can select weed biotypes with reduced herbicide sensitivity. Therefore, the objective of this study was to evaluate whether drift from glyphosate, 2,4-D, and dicamba applications could select for Amaranthus spp. with reduced herbicide sensitivity over two generations in a wind tunnel study.
Material and Methods
Plant material
Palmer amaranth and waterhemp seeds were collected from 10–20 putative herbicide susceptible plants in wheat (Triticum aestivum L.) and corn (Zea mays L.) fields in Nebraska (Table 1). Seeds from within a single field were identified as a population (Chase and Perkins for Palmer amaranth, and Thayer and Stanton for waterhemp) and stored at −20 °C for a minimum of three months to overcome dormancy. Following sowing, seedlings were transplanted into plastic tubes (1 L) containing commercial potting mix (Berger BM7 Bark Mix, Saint Modeste, QC, Canada) and maintained under greenhouse conditions (30/20 °C [day/night] with a 16 h photoperiod) at the Pesticide Application Technology Laboratory (University of Nebraska-Lincoln, West Central Research and Extension Center, North Platte, NE)17. Supplemental light (LED growth lights 520 μmol s−1, Philips Lighting, Somerset, NJ, USA) was provided to ensure a 16-h photoperiod. Plants were supplied with water including fertilizer solution (0.2% v/v) as needed (UNL 5-1-4, Wilbur-Ellis Agribusiness, Aurora, CO, USA)17.
Herbicide drift recurrent selection
Herbicide drift simulations were conducted in the low speed wind tunnel at the Pesticide Application Technology Laboratory. Glyphosate, 2,4-D, and dicamba solutions were prepared at 140 L ha−1 carrier volume (Table 2). The glyphosate solution had the addition of ammonium sulfate at 5% v/v to overcome antagonistic effects of cationic salts in hard water (Bronc, Wilbur-Ellis Agribusiness, Aurora, CO, USA). Herbicide applications were performed at 140 L ha−1 with two even nozzles, a conventional flat-fan nozzle (TP95015EVS) and an air-inclusion (AI) nozzle (AI95015EVS) (TeeJet Technologies Spraying Systems Co., Glendale Heights, IL, USA) at 230 kPa with constant wind speed of 4.47 m s−1 as we described in a previous study14. Nozzles were selected to provide high (Fine spray classification) and low (Ultra Coarse spray classification) drift potentials. The average air temperature and relative humidity during applications were 25 °C and 45%, respectively. Palmer amaranth and waterhemp plants (15–20 cm-tall) were positioned at four downwind distances: 1.0, 1.5, 2.0, 2.5 m from the nozzle simulating plants inhabiting field margins. Eighty plants of each population were exposed to herbicide*nozzle drift treatments, with 20 plants per distance. During applications, nozzles were positioned at 70 cm from the ground. Following herbicide drift exposure, plants were returned and kept under greenhouse conditions as previously described. Plant mortality was evaluated at 35 days after treatment (DAT).
Survivors of each herbicide*nozzle*population treatment were enclosed within tents (plants from all distances were pooled) constructed with 213-cm by 152-cm pollination bags (Vilutis & Co., Frankfort, IL, USA) to ensure cross-pollination exclusively within specific treatments. Tents were periodically shaken to facilitate pollination. Seeds from all plants within each treatment were collected at maturity, pooled, and termed P1 seeds. Seeds were dried at greenhouse room temperature and stored at −20 °C for 15 days. P1 seeds of each herbicide*nozzle*population treatment were used for the subsequent round of herbicide drift selection. Plant material, herbicide drift treatments, and isolation on pollination tents were conducted as previously described, and survivors from the second herbicide drift selection were grown to seed to establish the P2 progeny for each treatment. During each herbicide drift selection (P1 and P2 selection), a group of 40 untreated plants per population (Chase, Perkins, Thayer, and Stanton) was maintained and isolated on pollination tents using the same procedure previously described to establish P1 and P2 unselected controls. The study had a factorial arrangement with weed species, nozzle, and herbicide as factors in a completely randomized design. Plant mortality of treatment combinations were analyzed with a Beta-binomial distribution using a logit link function with a generalized mixed model in SAS software (SAS v9.4, SAS Institute Inc., Cary, NC, USA) and comparisons among treatments were performed using Fisher’s least significant difference procedure at significance level α = 0.05.
Herbicide dose response
Palmer amaranth and waterhemp P2 progenies (herbicide*population*nozzle treatments and non-selected controls) were subjected to glyphosate, 2,4-D, and dicamba dose-response study (respective to herbicide drift selection treatment) in the Pesticide Application Technology Laboratory. Following sowing, seedlings from P2 progenies were transplanted into plastic tubes containing commercial potting mix and maintained under greenhouse conditions as previously described. Plants (10- to 12-cm tall) were sprayed with different glyphosate, 2,4-D, and dicamba rates (Table 3) using a research spray chamber (DeVries, Hollandale, MN, USA) calibrated to deliver 93.5 L ha−1 using an AI95015EVS nozzle at 414 kPa. The experiment was conducted in a randomized complete design with four replications per treatment in which a single plant was considered as an experimental unit. Plant above ground biomass was harvested at 30 DAT and oven dried at 65 °C to constant weight. Biomass data were converted into percentage of biomass reduction as compared to the untreated control. A non-linear regression model was fitted to dry weight data in response to herbicide dose using the drc package in R software (R Foundation for Statistical Computing, Wien, Austria)17,57. The effective-dose to reduce 90% of plant biomass (GR90) was estimated for each P2 progeny using a four-parameter log-logistic model: y = c + {d − c/1 + exp[b(log x − log e)]} in which y corresponds to the biomass reduction (%), b is the slope at the inflection point, c is the lower limit of the model (fixed to 0%), d is the upper limit (fixed to 100%), and e is the inflection point (effective dose to reduce plant biomass in 50%). Resistance ratios were calculated as the ratio of the GR90 for each selected P2 population to the respective P2 unselected population17. The experiment was conducted twice and data were combined.
Results and Discussion
Herbicide drift exposure
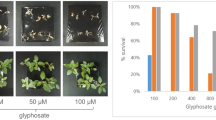
Glyphosate, 2,4-D, and dicamba drift exposure resulted in Palmer amaranth and waterhemp mortality (Tables 4 and 5). Amaranthus spp. mortality was influenced by nozzle type (p < 0.0001) and herbicide by weed species interaction (p < 0.0001). Herbicide drift from the flat fan nozzle resulted in 54–69% (CI 95%) overall mortality when the other variables were pooled, whereas the air inclusion nozzle resulted in 19 to 32% (CI 95%). These results corroborate previous field and wind tunnel results where applications with air inclusion nozzles resulted in less particle drift compared to flat fan nozzles58,59,60,61,62. The preorifice component of air inclusion nozzles is designed to reduce the solution pressure as it exits the nozzle, thereby increasing the droplet size of the spray and consequently reducing the drift potential63,64. We reported in a previous study that herbicide applications in a wind tunnel (4.47 m s−1 wind speed) with the flat fan nozzle resulted in 32, 23, 17, and 14% of herbicide drift (in relation to volume sprayed) at 1, 1.5, 2.0, and 2.5 m from the nozzle, respectively, whereas applications with the air inclusion nozzle resulted in 11, 7, 5, and 3% herbicide drift in the same downwind distances14.
Glyphosate drift resulted in increased plant mortality on Palmer amaranth (81–95% CI 95%) compared to 2,4-D (16–36% CI 95%) and dicamba (23–45% CI 95%). 2,4-D drift resulted in higher plant mortality on waterhemp (37–61% CI 95%) compared to glyphosate (18–41% CI 95%) and dicamba (16–36% CI 95%). Both species had similar mortality when exposed to dicamba drift. Inherent differences in herbicide response between Palmer amaranth and waterhemp were reported in previous studies, with Palmer amaranth being more tolerant to PPO-inhibitors50, but more susceptible to glyphosate17. Palmer amaranth was very susceptible to glyphosate drift, especially with applications using the flat fan nozzle. As a result, P1 and P2 progenies were not established for both Palmer amaranth populations (Perkins and Chase) exposed to glyphosate drift with the flat fan nozzle. Although a P1 progeny was established for the Chase population exposed to glyphosate drift using the air inclusion nozzle, a P2 progeny was not established as plants did not survive the second round of herbicide drift exposure.
A previously susceptible Palmer amaranth population evolved levels of glyphosate resistance following four selection rounds with low rates of glyphosate, with 58, 43, 51, and 79% plant mortality during selection rounds26. Similar resistance shift results were reported in an annual ryegrass (Lolium rigidum Gaudin) population recurrently selected with low rates of glyphosate in field conditions, although higher plant mortality ranging from 71 to 90% during four selection rounds was observed22. A wild radish (Raphanus raphanistrum L.) population evolved levels of 2,4-D resistance after four rounds of selection with sublethal rates of 2,4-D, with 71, 88, 77, and 76% mortality during each selection round18. Similarly, a Palmer amaranth population evolved higher levels of dicamba resistance after recurrent selection with low dicamba rates, with 47, 68, 29, and 79% plant mortality during each of the selection rounds27.
Herbicide drift recurrent selection
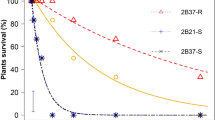
The Palmer amaranth population from Perkins County evolved glyphosate resistance (54.7-fold in the GR90) after being recurrently exposed to glyphosate drift with the air inclusion nozzle (Fig. 1). The Perkins population exposed to 2,4-D drift with the air inclusion nozzle had 2.5-fold shift in the GR90 after two selection rounds, whereas the progeny exposed to 2,4-D drift with the flat fan nozzle had a 1.8-fold shift (Fig. 2). On the other hand, the Palmer amaranth population from Chase County had no resistance shift after being recurrently selected with 2,4-D drift with both air inclusion and flat fan nozzles (Table 6). Moreover, both Palmer amaranth populations had no sensitivity shift following dicamba drift selection with both air inclusion and flat fan and nozzles (Fig. 3).

Biomass reduction for the Palmer amaranth population (P2) from Perkins County (NE) following recurrent selection to glyphosate spray drift at 30 days after treatment in the glyphosate dose response study.

Biomass reduction for the Palmer amaranth population (P2) from Perkins County (NE) following recurrent selection to 2,4-D spray drift at 30 days after treatment in the 2,4-D dose response study.

Biomass reduction for the Palmer amaranth population (P2) from Chase County (NE) following recurrent selection to dicamba spray drift at 30 days after treatment in the dicamba dose response study.
The waterhemp population from Stanton County showed no evidence of resistance shift when recurrently selected with glyphosate drift with the air inclusion nozzle, whereas plants exposed to glyphosate drift with the flat fan nozzle had a 2-fold glyphosate resistance shift (Table 7). The Thayer population had a 2.4 and 3.3-fold glyphosate resistance shift after being recurrently exposed to glyphosate drift with the air inclusion and the flat fan nozzles, respectively (Fig. 4). The Thayer population also had its 2,4-D sensitivity reduced after recurrent exposure to 2,4-D drift using the air inclusion (2.2-fold) and the flat fan nozzle (1.7-fold), whereas no shifts were observed for the Stanton population (Fig. 5). Recurrent exposure to dicamba drift with the air inclusion and the flat fan nozzles resulted in dicamba sensitivity shifts in the Thayer population (1.5 and 2.2-fold shift, respectively). The Stanton population also had its sensitivity to dicamba increased, but only for progenies exposed to dicamba drift with the flat fan nozzle (2.4-fold) (Fig. 6).

Biomass reduction for the waterhemp population (P2) from Thayer County (NE) following recurrent selection to glyphosate spray drift at 30 days after treatment in the glyphosate dose response study.

Biomass reduction for the waterhemp population (P2) from Thayer County (NE) following recurrent selection to 2,4-D spray drift at 30 days after treatment in the 2,4-D dose response study.

Biomass reduction for the waterhemp population (P2) from Stanton County (NE) following recurrent selection to dicamba spray drift at 30 days after treatment in the dicamba dose response study.
The reduced herbicide sensitivity shifts reported herein are consistent with resistance shifts reported in previous recurrent selection studies with intentional applications of low herbicide rates. Glyphosate sensitivity shift of 2.15-fold in the lethal dose required to 95% control (LD95) was reported in a Palmer amaranth population recurrently selected for four generations with low rates of glyphosate26. Similar results were reported in an annual ryegrass population, where resistance ratios in the GR50 ranged from 1.68 to 1.87 in progenies recurrently selected with low rates of glyphosate22. A wild radish population had its 2,4-D sensitivity reduced 3.4-fold (LD50) after recurrent selection with low rates of 2,4-D following two selection rounds18. Moreover, authors reported a resistance shift of 8.6-fold as recurrent selection continued during two additional selection rounds. A similar trend was reported for a Palmer amaranth population recurrently selected with low rates of dicamba, where a 2.6-fold dicamba sensitivity shift (LD90) was reported following two rounds of selection27. Additionally, the authors reported a 3.9-fold dicamba resistance shift in the third selection round.
The 54.7-fold glyphosate sensitivity shift in the Palmer amaranth progeny from Perkins County is unprecedented in the literature. This large resistance shift indicates that although the initial Palmer amaranth progeny was classified as glyphosate-susceptible, individuals with a major glyphosate resistance mechanism were already present within the population prior to glyphosate drift selection. In a further screening, four out of 195 plants of the initial unselected Perkins population (P0) survived a diagnostic glyphosate rate (197 g ae ha−1). Investigations conducted at the Molecular Weed Science Laboratory at Colorado State University revealed that P0 plants from Perkins County that survived the 197 g ae ha−1 glyphosate rate had increased EPSPS copy number with 11 to 38 copies relative to ALS gene (low copy control gene) (supplementary information). The glyphosate resistance trait rapidly became predominant in this Palmer amaranth population following recurrent exposure to glyphosate drift.
Herbicide sensitivity reduction in this study varied across weed species, weed population, spray drift potential (nozzle), and herbicide active ingredient. In this study, waterhemp was more prone to herbicide sensitivity shifts following herbicide drift selection compared to Palmer amaranth. Moreover, the waterhemp population from Thayer County had more herbicide sensitivity shifts following herbicide drift selection compared to the Stanton County population. A similar trend was observed for Palmer amaranth, where the population from Perkins County was more prone to herbicide sensitivity reduction following herbicide drift selection compared to the population from Chase County. Across Amaranthus spp. populations tested herein, glyphosate sensitivity reduction was predominant over 2,4-D and dicamba following drift selection with the respective herbicides. Nozzle type influenced resistance shifts following herbicide drift exposure with glyphosate and dicamba, where progenies selected with the flat fan nozzle had greater selection intensity (higher mortality), and consequently larger resistance shifts. Interestingly, this trend was not observed for 2,4-D drift, where recurrent selection with the air inclusion nozzle resulted in slightly larger resistance shifts compared to the flat fan nozzle despite differences in selection intensity between nozzles.
Recurrent selection with low doses of herbicides can progressively favor for metabolism alleles present within the standing genetic variation of the population, which additively leads to non-target-site herbicide resistance20,32. A study reported that a previous susceptible annual ryegrass population evolved diclofop resistance following recurrent selection with low rates of diclofop24. Further investigations revealed that the recurrent selection with low rates of diclofop selected for non-target-site resistance with enhanced diclofop metabolism, likely mediated by cytochrome P450 monooxygenases (P450)65. A RNA-Seq transcriptome study with this population confirmed that not only P450 genes, but nitronate monooxygenase (NMO), glutathione transferase (GST), and glucosyltransferase (GT) genes were upregulated in diclofop-resistant plants31. Further studies reported upregulation of metabolic genes (GST) in a pyroxasulfone-resistant annual ryegrass population recurrently selected with low rates of the herbicide19,30. While Palmer amaranth and waterhemp populations with target-site glyphosate resistance (EPSPS gene amplification) were reported in Nebraska66,67, metabolic resistance to other herbicides is more frequent in waterhemp. A 2,4-D-resistant waterhemp population reported in Nebraska had rapid 2,4-D metabolism mediated by P450 enzymes68. Enhanced herbicide metabolism via P450 enzymes was also reported in a waterhemp population resistant to HPPD-inhibitor herbicides in Nebraska69,70. Atrazine resistance with rapid herbicide metabolism via enhanced GST conjugation was widespread in waterhemp populations in Nebraska54. Although non-target-site glyphosate resistance with metabolism in plants is relatively rare71, non-target-site resistance with reduced glyphosate translocation was identified in waterhemp biotypes in Mississippi72. Waterhemp biotypes with non-target-site resistance to glyphosate were also reported in Missouri52. Glyphosate metabolism with increased aldo-keto reductase (AKR) activity was reported in Echinochloa colona in Australia73.
Herbicide resistance alleles may be originally present within the standing genetic variation of the population or may immigrate via pollen or seeds from other populations74. As populations were collected in commercial cropping fields, and considering the rampant pollen-mediated gene flow and seeds transferring herbicide resistant alleles across waterhemp and Palmer amaranth populations in Nebraska, it can be inferred that herbicide resistance alleles could already be present within the standing genetic variation of the Amaranthus spp. populations tested herein55,56. This could explain the differences in herbicide sensitivity shift between waterhemp and Palmer amaranth, and even the differences among populations (different genetic background) following recurrent selection with herbicide drift. The influence of selection intensity (nozzle type), weed species, and weed population on glyphosate and dicamba sensitivity shifts following drift selection suggest that resistance alleles present within the standing genetic variability of populations were progressively selected during selection rounds. Some researchers suggest that low rates of herbicides could also act as stress agents inducing new stress-related mutations and epigenetic alterations that could ultimately lead to reduced herbicide sensitivity33,34,35. On the other hand, a study where over 70 million Amaranthus hypochondriacus L. seedlings were screened with imazethapyr showed no evidence suggesting that herbicide stress increased mutation rates conferring ALS resistance, although authors mentioned that stress-mediated increase of mutation rates leading to herbicide resistance remains biologically possible74. Both Perkins and Chase Palmer amaranth plants were physiologically stressed following dicamba drift and did not evolve levels of dicamba resistance following drift exposure for two generations, although we recognize that additional selection rounds would be necessary to expand the discussion. Interestingly, the 2,4-D sensitivity shifts in Palmer amaranth (Perkins) and waterhemp (Thayer) following drift selection were independent of selection intensity (nozzle type). Further studies are necessary to investigate the molecular basis of the sensitivity shifts found in the Amaranthus spp. following recurrent herbicide drift selection in this study.
The results of this study confirm that herbicide drift towards field margins can rapidly select for biotypes with reduced herbicide sensitivity with minor and major herbicide resistance mechanisms. Preventing the establishment of resistance prone weeds on field margins and ditches in agricultural landscapes is an important management strategy to delay herbicide resistance, especially for cross-pollinated weed species such as Palmer amaranth and waterhemp16,75. Weed management programs should consider strategies to mitigate near-field spray drift, and suppress weed populations on field borders and ditches in agricultural landscapes16,62,75,76.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Bagavathiannan, M. V. et al. Considering weed management as a social dilemma bridges individual and collective interests. Nat. Plants 5, 343–351 (2019).
Behrens, M. R. et al. Dicamba Resistance: Enlarging and Preserving Biotechnology-Based Weed Management Strategies. Science 316, 1185–1188 (2007).
Padgette, S. R. et al. Development, Identification, and Characterization of a Glyphosate-Tolerant Soybean Line. Crop Sci. 35, 1451–1461 (1995).
Wright, T. R. et al. Robust crop resistance to broadleaf and grass herbicides provided by aryloxyalkanoate dioxygenase transgenes. Proc. Natl. Acad. Sci. USA 107, 20240–20245 (2010).
Matthews, G., Bateman, R. & Miller, P. Pesticide Application Methods 4th edition. (Wiley-Blackwell, 2014).
Buehring, N. W., Massey, J. H. & Reynolds, D. B. Shikimic Acid Accumulation in Field-Grown Corn (Zea mays) Following Simulated Glyphosate Drift. J. Agric. Food Chem. 55, 819–824 (2007).
Ding, W. et al. Biological Response of Soybean and Cotton to Aerial Glyphosate Drift. J. Crop Improv. 25, 291–302 (2011).
Egan, J. F., Barlow, K. M. & Mortensen, D. A. A Meta-Analysis on the Effects of 2,4-D and Dicamba Drift on Soybean and Cotton. Weed Sci. 62, 193–206 (2014).
Egan, J. F. & Mortensen, D. A. Quantifying vapor drift of dicamba herbicides applied to soybean. Environ. Toxicol. Chem. 31, 1023–1031 (2012).
Hewitt, A. J. Spray drift: impact of requirements to protect the environment. Crop Prot. 19, 623–627 (2000).
Jones, G. T., Norsworthy, J. K., Barber, T., Gbur, E. & Kruger, G. R. Off-target Movement of DGA and BAPMA Dicamba to Sensitive Soybean. Weed Technol. 33, 51–65 (2019).
Kalsing, A. et al. Effect of Formulations and Spray Nozzles on 2,4-D Spray Drift under Field Conditions. Weed Technol. 32, 379–384 (2018).
Reddy, K. N. et al. Biological responses to glyphosate drift from aerial application in non-glyphosate-resistant corn. Pest Manag. Sci. 66, 1148–1154 (2010).
Vieira, B. C. et al. Response of Amaranthus spp. following exposure to sublethal herbicide rates via spray particle drift. Plos One 14, e0220014 (2019).
Bagavathiannan, M. V. & Norsworthy, J. K. Multiple-Herbicide Resistance Is Widespread in Roadside Palmer Amaranth Populations. Plos One 11, e0148748 (2016).
Norsworthy, J. K. et al. Reducing the Risks of Herbicide Resistance: Best Management Practices and Recommendations. Weed Sci. 60, 31–62 (2012).
Vieira, B. C. et al. Distribution of glyphosate-resistant Amaranthus spp. in Nebraska. Pest Manag. Sci. 74, 2316–2324 (2018).
Ashworth, M. B., Walsh, M. J., Flower, K. C. & Powles, S. B. Recurrent selection with reduced 2,4-D amine doses results in the rapid evolution of 2,4-D herbicide resistance in wild radish (Raphanus raphanistrum L.). Pest Manag. Sci. 72, 2091–2098 (2016).
Busi, R., Gaines, T. A., Walsh, M. J. & Powles, S. B. Understanding the potential for resistance evolution to the new herbicide pyroxasulfone: field selection at high doses versus recurrent selection at low doses. Weed Res. 52, 489–499 (2012).
Busi, R., Neve, P. & Powles, S. Evolved polygenic herbicide resistance in Lolium rigidum by low-dose herbicide selection within standing genetic variation. Evol. Appl. 6, 231–242 (2013).
Busi, R., Girotto, M. & Powles, S. B. Response to low-dose herbicide selection in self-pollinated Avena fatua. Pest Manag. Sci. 72, 603–608 (2016).
Busi, R. & Powles, S. B. Evolution of glyphosate resistance in a Lolium rigidum population by glyphosate selection at sublethal doses. Heredity 103, 318–325 (2009).
Manalil, S., Busi, R., Renton, M. & Powles, S. B. Rapid Evolution of Herbicide Resistance by Low Herbicide Dosages. Weed Sci. 59, 210–217 (2011).
Neve, P. & Powles, S. Recurrent selection with reduced herbicide rates results in the rapid evolution of herbicide resistance in Lolium rigidum. Theor. Appl. Genet. 110, 1154–1166 (2005).
Neve, P. & Powles, S. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 95, 485–492 (2005).
Norsworthy, J. K. Repeated Sublethal Rates of Glyphosate Lead to Decreased Sensitivity in Palmer Amaranth. Crop Manag. https://doi.org/10.1094/CM-2012-0403-01-RS, (2012).
Tehranchian, P. et al. Recurrent Sublethal-Dose Selection for Reduced Susceptibility of Palmer Amaranth (Amaranthus palmeri) to Dicamba. Weed Sci. 65, 206–212 (2017).
Vila-Aiub, M. M. & Ghersa, C. M. Building up resistance by recurrently exposing target plants to sublethal doses of herbicide. Eur. J. Agron. 22, 195–207 (2005).
Regnier, E. E. et al. Certified Crop Advisors’ Perceptions of Giant Ragweed (Ambrosia trifida) Distribution, Herbicide Resistance, and Management in the Corn Belt. Weed Sci. 64, 361–377 (2016).
Busi, R., Porri, A., Gaines, T. A. & Powles, S. B. Pyroxasulfone resistance in Lolium rigidum is metabolism-based. Pestic. Biochem. Physiol. 148, 74–80 (2018).
Gaines, T. A. et al. RNA-Seq transcriptome analysis to identify genes involved in metabolism-based diclofop resistance in Lolium rigidum. Plant J. 78, 865–876 (2014).
Neve, P., Busi, R., Renton, M. & Vila-Aiub, M. M. Expanding the eco-evolutionary context of herbicide resistance research. Pest Manag. Sci. 70, 1385–1393 (2014).
Dyer, W. E. Stress-induced evolution of herbicide resistance and related pleiotropic effects. Pest Manag. Sci. 74, 1759–1768 (2018).
Gressel, J. Low pesticide rates may hasten the evolution of resistance by increasing mutation frequencies. Pest Manag. Sci. 67, 253–257 (2010).
Markus, C., Pecinka, A., Karan, R., Barney, J. N. & Merotto, A. Epigenetic regulation – contribution to herbicide resistance in weeds? Pest Manag. Sci. 74, 275–281 (2018).
Yu, Q. & Powles, S. Metabolism-Based Herbicide Resistance and Cross-Resistance in Crop Weeds: A Threat to Herbicide Sustainability and Global Crop Production. Plant Physiol. 166, 1106–1118 (2014).
Steckel, L. E. The Dioecious Amaranthus spp.: Here to Stay. Weed Technol. 21, 567–570 (2007).
Bensch, C. N., Horak, M. J. & Peterson, D. Interference of Redroot Pigweed (Amaranthus retroflexus), Palmer Amaranth (A. palmeri), and Common Waterhemp (A. rudis) in Soybean. Weed Sci. 51, 37–43 (2003).
Horak, M. J. & Loughin, T. M. Growth Analysis of Four Amaranthus Species. Weed Sci. 48, 347–355 (2000).
Keeley, P. E., Carter, C. H. & Thullen, R. J. Influence of Planting Date on Growth of Palmer Amaranth (Amaranthus palmeri). Weed Sci. 35, 199–204 (1987).
Sauer, J. Recent Migration and Evolution of the Dioecious Amaranths. Evolution 11, 11–31 (1957).
Sauer, J. D. The Grain Amaranths: A Survey of Their History and Classification. Ann. Mo. Bot. Gard. 37, 561–632 (1950).
Sauer, J. D. The Grain Amaranths and Their Relatives: A Revised Taxonomic and Geographic Survey. Ann. Mo. Bot. Gard. 54, 103–137 (1967).
Werle, R., Sandell, L. D., Buhler, D. D., Hartzler, R. G. & Lindquist, J. L. Predicting Emergence of 23 Summer Annual Weed Species. Weed Sci. 62, 267–279 (2014).
Bernards, M. L., Crespo, R. J., Kruger, G. R., Gaussoin, R. & Tranel, P. J. A Waterhemp (Amaranthus tuberculatus) Population Resistant to 2,4-D. Weed Sci. 60, 379–384 (2012).
Crespo, R. J. et al. Multiple-Herbicide Resistance in a 2,4-D—Resistant Waterhemp (Amaranthus tuberculatus) Population from Nebraska. Weed Sci. 65, 743–754 (2017).
Evans, C. M. et al. Characterization of a waterhemp (Amaranthus tuberculatus) population from Illinois resistant to herbicides from five site-of-action groups. Weed Technol. 33, 400–410 (2019).
Heap, I. International survey of herbicide resistant weeds. weedscience.org (2019).
Jhala, A. J., Sandell, L. D., Rana, N., Kruger, G. R. & Knezevic, S. Z. Confirmation and Control of Triazine and 4-Hydroxyphenylpyruvate Dioxygenase-Inhibiting Herbicide-Resistant Palmer Amaranth (Amaranthus palmeri) in Nebraska. Weed Technol. 28, 28–38 (2014).
Lillie, K. J., Giacomini, D. A., Green, J. D. & Tranel, P. J. Coevolution of resistance to PPO inhibitors in waterhemp (Amaranthus tuberculatus) and Palmer amaranth (Amaranthus palmeri). Weed Sci. 67, 521–526 (2019).
Murphy, B. P., Larran, A. S., Ackley, B., Loux, M. M. & Tranel, P. J. Survey of Glyphosate-, Atrazine- and Lactofenresistance Mechanisms in Ohio Waterhemp (Amaranthus tuberculatus) Populations. Weed Sci. 67, 296–302 (2019).
Schultz, J. L., Chatham, L. A., Riggins, C. W., Tranel, P. J. & Bradley, K. W. Distribution of Herbicide Resistances and Molecular Mechanisms Conferring Resistance in Missouri Waterhemp (Amaranthus rudis Sauer) Populations. Weed Sci. 63, 336–345 (2015).
Shergill, L. S., Barlow, B. R., Bish, M. D. & Bradley, K. W. Investigations of 2,4-D and Multiple Herbicide Resistance in a Missouri Waterhemp (Amaranthus tuberculatus) Population. Weed Sci. 66, 386–394 (2018).
Vennapusa, A. R. et al. Prevalence and Mechanism of Atrazine Resistance in Waterhemp (Amaranthus tuberculatus) from Nebraska. Weed Sci. 66, 595–602 (2018).
Oliveira, M. C. et al. Interspecific and intraspecific transference of metabolism-based mesotrione resistance in dioecious weedy Amaranthus. Plant J. 96, 1051–1063 (2018).
Sarangi, D. et al. Pollen-mediated gene flow from glyphosate-resistant common waterhemp (Amaranthus rudis Sauer): consequences for the dispersal of resistance genes. Sci. Rep. 7, srep44913 (2017).
Knezevic, S. Z., Streibig, J. C. & Ritz, C. Utilizing R Software Package for Dose-Response Studies: The Concept and Data Analysis. Weed Technol. 21, 840–848 (2007).
Alves, G. S. et al. Spray Drift from Dicamba and Glyphosate Applications in a Wind Tunnel. Weed Technol. 31, 387–395 (2017).
Alves, G. S. et al. Dicamba Spray Drift as Influenced by Wind Speed and Nozzle Type. Weed Technol. 31, 724–731 (2017).
Ferguson, J. C. et al. Determining the drift potential of Venturi nozzles compared with standard nozzles across three insecticide spray solutions in a wind tunnel. Pest Manag. Sci. 72, 1460–1466 (2016).
Johnson, A. K., Roeth, F. W., Martin, A. R. & Klein, R. N. Glyphosate Spray Drift Management with Drift-Reducing Nozzles and Adjuvants. Weed Technol. 20, 893–897 (2006).
Vieira, B. C. et al. Spray particle drift mitigation using field corn (Zea mays L.) as a drift barrier. Pest Manag. Sci. 74, 2038–2046 (2018).
Creech, C. F., Henry, R. S., Fritz, B. K. & Kruger, G. R. Influence of Herbicide Active Ingredient, Nozzle Type, Orifice Size, Spray Pressure, and Carrier Volume Rate on Spray Droplet Size Characteristics. Weed Technol. 29, 298–310 (2015).
Dorr, G. J. et al. A comparison of initial spray characteristics produced by agricultural nozzles. Crop Prot. 53, 109–117 (2013).
Yu, Q., Han, H., Cawthray, G. R., Wang, S. F. & Powles, S. B. Enhanced rates of herbicide metabolism in low herbicide-dose selected resistant Lolium rigidum. Plant Cell Environ. 36, 818–827 (2013).
Chahal, P. S., Varanasi, V. K., Jugulam, M. & Jhala, A. J. Glyphosate-Resistant Palmer Amaranth (Amaranthus palmeri) in Nebraska: Confirmation, EPSPS Gene Amplification, and Response to POST Corn and Soybean Herbicides. Weed Technol. 31, 80–93 (2017).
Chatham, L. A. et al. A Multistate Study of the Association Between Glyphosate Resistance and EPSPS Gene Amplification in Waterhemp (Amaranthus tuberculatus). Weed Sci. 63, 569–577 (2015).
Figueiredo, M. R. et al. Metabolism of 2,4-dichlorophenoxyacetic acid contributes to resistance in a common waterhemp (Amaranthus tuberculatus) population. Pest Manag. Sci. 74, 2356–2362 (2018).
Kaundun, S. S. et al. Mechanism of resistance to mesotrione in an Amaranthus tuberculatus population from Nebraska, USA. Plos One 12, e0180095 (2017).
Oliveira, M. C. et al. Reversing resistance to tembotrione in an Amaranthus tuberculatus (var. rudis) population from Nebraska, USA with cytochrome P450 inhibitors. Pest Manag. Sci. 74, 2296–2305 (2018).
Sammons, R. D. & Gaines, T. A. Glyphosate resistance: state of knowledge. Pest Manag. Sci. 70, 1367–1377 (2014).
Nandula, V. K., Ray, J. D., Ribeiro, D. N., Pan, Z. & Reddy, K. N. Glyphosate Resistance in Tall Waterhemp (Amaranthus tuberculatus) from Mississippi is due to both Altered Target-Site and Nontarget-Site Mechanisms. Weed Sci. 61, 374–383 (2013).
Pan, L. et al. Aldo-keto reductase metabolizes glyphosate and confers glyphosate resistance in Echinochloa colona. Plant Physiol. 00979.2019, https://doi.org/10.1104/pp.19.00979 (2019).
Casale, F. A., Giacomini, D. A. & Tranel, P. J. Empirical investigation of mutation rate for herbicide resistance. Weed Sci. 1–8, https://doi.org/10.1017/wsc.2019.19 (2019).
Riar, D. S. et al. Adoption of Best Management Practices for Herbicide-Resistant Weeds in Midsouthern United States Cotton, Rice, and Soybean. Weed Technol. 27, 788–797 (2013).
Foster, H. C., Sperry, B. P., Reynolds, D. B., Kruger, G. R. & Claussen, S. Reducing Herbicide Particle Drift: Effect of Hooded Sprayer and Spray Quality. Weed Technol. 32, 714–721 (2018).
Acknowledgements
The authors would like to thank CAPES (Brazilian Government Foundation) for the financial support to the graduate student Bruno Canella Vieira (proc 013041/2013-04). This project was partially supported by the Nebraska Agricultural Experiment Station with funding from the Hatch Multistate Research capacity funding program from the USDA National Institute of Food and Agriculture. The authors also would like to thank all undergraduate students, graduate students, and professional staff at the University of Nebraska-Lincoln who assisted with data collection and analysis.
Author information
Authors and Affiliations
Contributions
B.V., J.L., K.A., T.G., R.W. and G.K. designed the study. B.V. performed the experiment, analyzed the data, and wrote the manuscript. All authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vieira, B.C., Luck, J.D., Amundsen, K.L. et al. Herbicide drift exposure leads to reduced herbicide sensitivity in Amaranthus spp.. Sci Rep 10, 2146 (2020). https://doi.org/10.1038/s41598-020-59126-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-59126-9
- Springer Nature Limited
This article is cited by
-
A post-emergence herbicide program for weedy sunflower (Helianthus annuus L.) control in maize
Phytoparasitica (2024)
-
Structural-based analysis of sulfonamide derivatives: from solid states to acetolactate synthase enzyme interactions
Rendiconti Lincei. Scienze Fisiche e Naturali (2024)
-
Towards reducing chemical usage for weed control in agriculture using UAS imagery analysis and computer vision techniques
Scientific Reports (2023)
-
Particle drift simulation from mesotrione and rimsulfuron plus thifensulfuron-methyl mixture through two nozzle types to field and vegetable crops
Environmental Science and Pollution Research (2022)