
Abstract
Meiotic recombination is essential for producing healthy gametes, and also generates genetic diversity. DNA double-strand break (DSB) formation is the initiating step of meiotic recombination, producing, among other outcomes, crossovers between homologous chromosomes (homologs), which provide physical links to guide accurate chromosome segregation. The parameters influencing DSB position and repair are thus crucial determinants of reproductive success and genetic diversity. Using Schizosaccharomyces pombe, we show that the distance between sequence polymorphisms across homologs has a strong impact on meiotic recombination rate. The closer the sequence polymorphisms are to each other across the homologs the fewer recombination events were observed. In the immediate vicinity of DSBs, sequence polymorphisms affect the frequency of intragenic recombination events (gene conversions). Additionally, and unexpectedly, the crossover rate of flanking markers tens of kilobases away from the sequence polymorphisms was affected by their relative position to each other amongst the progeny having undergone intragenic recombination. A major regulator of this distance-dependent effect is the MutSα-MutLα complex consisting of Msh2, Msh6, Mlh1, and Pms1. Additionally, the DNA helicases Rqh1 and Fml1 shape recombination frequency, although the effects seen here are largely independent of the relative position of the sequence polymorphisms.
Similar content being viewed by others

Introduction
Correct chromosome segregation during meiosis depends on pairing and physical connection of homologous chromosomes (homologs). Physical connections are established by the repair of programmed DNA double-strand breaks (DSBs) using the homolog rather than the sister chromatid as a template (i.e. interhomolog recombination) and by ensuring that interhomolog recombination intermediates are processed into crossovers (COs). The formation of DSBs by the transesterase Spo11 is thus a key step in initiating recombination during meiosis1. Regions of high-frequency Spo11 recruitment, and thus DSB formation, are called hotspots2. One of the best characterized category of hotspots are cAMP-responsive elements in Schizosaccharomyces pombe, created by point mutations in the ade6 gene that represent binding sites for the Atf1-Pcr1 transcription factor2,3. These include the ade6-M26 hotspot and its derivatives, which are defined by the DNA sequence heptamer 5′-ATGACGT-3′3. Although binding of Atf1-Pcr1 and the associated transcription already creates open chromatin at M26-like hotspots3,4, a very high frequency of meiotic recombination requires a conducive chromatin environment in a wider genomic context5,6. This network of parameters determines the overall level of DSB formation at a given genomic locus.
Following break formation, DSB ends are resected to initiate homologous recombination, which during meiosis follows either a Holliday junction/D-loop resolution or a synthesis-dependent strand annealing (SDSA) pathway1,7. As a repair template, either the sister chromatid or the homolog can be used8. Based on this, it has been suggested that the governance of meiotic recombination could be viewed as a two-tiered decision system9. The first decision being template choice (interhomolog vs. intersister recombination), and the second being how the recombination intermediate is resolved - i.e. the CO/non-crossover (NCO) decision. The template choice decision is mainly driven by meiosis-specific factors of the chromosome axis and by the meiotic recombinase Dmc1 supported by its mediators8. In budding yeast there is a basic understanding of how the interhomolog bias is established, although some mechanistic details still remain to be elucidated10. Since homologs are not necessarily identical on a DNA sequence level, a DSB end invading the homolog for repair can generate mismatch-containing heteroduplex DNA. Mismatches can be corrected by the mismatch repair system, consisting of the highly conserved MutS and MutL proteins11. Additionally, the MutS-MutL complex can also block strand invasion to avoid recombination between non-homologous sequences11. The CO/NCO-decision happens as the next step; here the decision is taken whether an already established interhomolog recombination intermediate is processed into a CO or a NCO. Determinants of the CO/NCO-decision are less well studied, but the DNA helicase/translocase FANCM (Fml1 in Sz. pombe) has been shown to limit CO formation in fission yeast and Arabidopsis12,13. RecQ-type DNA helicases perform a wide range of regulatory roles in homologous recombination, and one of which probably is the promotion of NCO formation during meiosis in various organisms14,15,16,17,18.
Here, we employ a series of meiotic recombination assays featuring intragenic markers at differently sized intragenic intervals and flanking intergenic markers to identify and characterize intrinsic determinants of template choice and CO/NCO-decision in fission yeast. We show that the relative positions of DNA sequence polymorphisms between homologs have a strong impact on recombination outcome, not only locally in the form of intragenic recombination (gene conversion), but also on the CO frequency between an up- and a downstream marker. The anti-recombinogenic activity of MutSα-MutLα factors, and of the DNA helicases Fml1 and Rqh1 modulate recombination outcome differentially when comparing various intragenic intervals.
Results and Discussion
Rationale of the meiotic recombination assay
Our meiotic recombination assay features intragenic markers (point mutations in the ade6 gene) and flanking intergenic markers (ura4+-aim2 and his3+-aim) (Fig. 1). This assay allows us to monitor various recombination outcomes: (I) intragenic recombination (gene conversion) events producing Ade+ recombinants, (II) crossovers (COs) between the flanking intergenic markers (ura4+-aim2 and his3+-aim), and (III) the ratio of COs vs. non-crossovers (NCOs) among intragenic ade6+ recombination events (Fig. 1A). Changes in gene conversion and overall CO frequencies observed in this assay can be explained by an altered frequency of DSB formation at a given ade6 mutant allele, or a change in repair template usage. The percentages of COs and NCOs among intragenic ade6+ recombination events are the genetic readout for the CO/NCO-decision, representing recombination intermediate processing after successful strand exchange between homologs. The intragenic events are most likely the result of gene conversions associated with COs or NCOs (non-reciprocal exchange of hereditary information).

Meiotic recombination assay composed of ade6 heteroalleles flanked by artificially introduced markers ura4+-aim2 & his3+-aim. (A) Schematic showing the meiotic recombination assay at ade6 (yellow) and its common outcomes. Ade+ recombinants can arise via gene conversion (GC) associated with a crossover (GC-CO) or a non-crossover (GC-NCO). The positions of ade6, and the artificially introduced markers ura4+-aim2 (green) and his3+-aim (light blue) on chromosome 3 are indicated [in bps]. Positions of point mutations are shown as ▼ and ×. (B) Schematic of the ade6 coding sequence indicating the point mutations and their positions (approximately to scale) used in the recombination assays, hotspots are indicated in red, and non-hotspots in light blue. The distance between the sequence polymorphisms across the homologs is indicated in relation to the given hotspot of each cross [in bp].
The physical distance between point mutations of heteroalleles defines the frequency of intragenic recombination events and their associated CO/NCO ratio
Apart from absolute DSB levels, intragenic recombination frequency is also influenced by the distance between point mutations in a given chromosomal region5,19,20,21. Intragenic recombination in our assays (Fig. 1A) has so far been monitored using point mutations within the ade6 coding sequence, which are at least 1 kb apart12,22,23. We wondered whether the level of COs among intragenic recombination events also changes, when the distance between point mutations was decreased. Therefore, we selected a series of point mutations, which cover almost the complete length of the ade6 coding sequence (Fig. 1B, Supplementary Table S1). These point mutants include the strong meiotic recombination hotspots ade6-M26, -3074, -3083, at the 5′ end of the gene and -3049 at the 3′ prime end of the gene, and the non-hotspot alleles ade6-M216, -M375, -704, -52, -149, -51, and -469 (Fig. 1B, Supplementary Table S1). All hotspots used here mimic a cAMP-response element, which creates a binding site for the Atf1-Pcr1 transcription factor; this in turn generates open chromatin3,5. It can be safely assumed that a given hotspot will receive the same amount of breakage independent of the ade6 allele present on the homolog. This means that the differences seen in the combinations of one specific hotspot with various ade6 alleles will depend on processes downstream of DSB formation. Indeed, the frequency of intragenic recombination positively correlates with the distance between the ade6 alleles, when the same hotspot is used (Fig. 2A, black and grey lines). Recombination at the ade6-M375 allele, which is at a similar position as the strong hotspot alleles ade6-3074 & ade6-3083, is induced at an overall much lower level (Fig. 2A, green line), but appears to be the acceptor of genetic information when crossed to ade6-469 (Fig. 2E), indicating that ade6-M375 is somewhat more recombinogenic than ade6-469. Intragenic recombination frequency at ade6-M375 shows a similar correlation with respect to distance between the DNA polymorphisms as crosses involving hotspots (Fig. 2A). Intragenic intervals of similar size containing the meiotic recombination hotspot alleles, ade6-3083, ade6-3074, or ade6-3049, and a non-hotspot allele produce equivalent intragenic recombination levels (Fig. 2A). Therefore, these hotspot alleles behave similarly in determining intragenic recombination frequency.

Physical distance between heteroalleles in ade6 influences frequency of gene conversion (GC) and associated crossovers (COs). (A) Frequency of GC and (B) frequency of CO among GC events at ade6 in wild type over distance between point mutations: crosses involving hotspot ade6-3083 as black solid line, UoA110 × UoA100 (ade6-3083 × ade6-M216) (n = 12), ALP733 × UoA115 (ade6-3083 × ade6-704) (n = 12), ALP733 × UoA119 (ade6-3083 × ade6-52) (n = 5), ALP733 × ALP731 (ade6-3083 × ade6-469) (n = 20); crosses involving hotspot ade6-3074 as black dashed line, UoA106 × UoA100 (ade6-3074 × ade6-M216) (n = 12), UoA104 × UoA115 (ade6-3074 × ade6-704) (n = 12), UoA104 × UoA119 (ade6-3074 × ade6-52) (n = 6), UoA104 × ALP731 (ade6-3074 × ade6-469) (n = 10); crosses involving hotspot ade6-3049 as grey line, UoA122 × UoA497 (ade6-3049 × ade6-149) (n = 6), UoA120 × UoA463 (ade6-3049 × ade6-51) (n = 6), UoA120 × ALP731 (ade6-3049 × ade6-469) (n = 31), UoA116 × UoA123 (ade6-3049 × ade6-52) (n = 12), UoA112 × UoA123 (ade6-3049 × ade6-704) (n = 12), ALP1541 × UoA123 (ade6-3049 × ade6-M375) (n = 12), UoA99 × UoA123 (ade6-3049 × ade6-M216) (n = 12); and crosses involving non-hotspot ade6-M375 as green line – needs to be read from the green secondary y-axis in (A), UoA861 × UoA100 (ade6-M375 × ade6-M216) (n = 6), ALP1541 × UoA119 (ade6-M375 × ade6-52) (n = 6), ALP1541 × ALP731 (ade6-M375 × ade6-469) (n = 16). (C) Frequency of GC and (D) frequency of CO among GC events at ade6 in wild type crosses involving hotspot alleles only: FO1285 × UoA123 (ade6-M26 × ade6-3049) (n = 12), UoA104 × UoA123 (ade6-3074 × ade6-3049) (n = 9), and ALP733 × UoA123 (ade6-3083 × ade6-3049) (n = 9); the hotspot × non-hotspot cross UoA99 × UoA123 (ade6-3049 × ade6-M216) (n = 12) is shown for comparison. (E) Distribution of non-crossover (NCO; Ura+ His− & Ura− His+) and crossover (CO; Ura+ His+ & Ura− His−) classes among Ade+ GC events in wild type (percentages in each class are shown as means ± Std. Dev.); ALP1541 × ALP731 (n = 16), ALP733 × ALP731 (n = 20), ALP1541 × UoA123 (n = 12), ALP733 × UoA123 (n = 9). n indicates the number of independent crosses. For details of data see Supplementary Table S2.
Intriguingly, these observations are also largely true for CO frequency among intragenic recombination events: The shorter an intragenic distance between polymorphisms is, the more likely an intragenic recombination event is resolved as a NCO (Fig. 2B). For crosses involving the hotspot alleles ade6-3083 or ade6-3074 the effect apparently tails off at intragenic distances >600 bp (Fig. 2B). Combining hotspot alleles on both homologs within a cross results in increased overall intragenic recombination rate compared with a hotspot × non-hotspot cross covering a similar intragenic distance between point mutations (Fig. 2C), in line with previous reports24. However, there is no notable difference in COs among intragenic recombination events (Fig. 2D). This indicates that the frequency of CO among intragenic recombination events is primarily a function of the distance between the ade6 heteroalleles on the homologs.
The distribution of different NCO/CO classes amongst intragenic recombination events follows a pattern consistent with intragenic NCOs more likely being associated with the hotter allele. This means that the allele more likely to receive a DSB is the recipient of genetic information in the overwhelming majority of cases, which might represent a bona fide gene conversion event, e.g. the vast majority of Ade+ NCO events in the ade6-3083 × ade6-469 cross are Ura+ His−, because the ade6-3083 allele is linked to the ura4+-aim2 marker (Fig. 2E). If comparable hotspots are combined in a cross the two intragenic NCO classes occur with roughly equal frequency (Fig. 2E, compare cross ade6-3083 × ade6-3049 to crosses ade6-3083 × ade6-469 & ade6-M375 × ade6-3049).
The observed distribution patterns also suggest that, at these long intragenic intervals, a subset of CO events could stem from the processing of one joint molecule, presumably a single Holliday junction25 or its precursors, positioned between the two ade6 point mutations; in contrast to a gene conversion event being resolved as a CO. This idea makes the following prediction: If CO events among Ade+ recombinants (mostly Ura− His− genotypes) are created by processing of a joint molecule situated between the two ade6 point mutations, then reciprocal Ura+ Ade− His+ recombinants carrying the mutations of both ade6 heteroalleles must exist. To test this, we sequenced the ade6 locus from 32 Ura+ Ade− His+ colonies from an ade6-3083 × ade6-469 cross. Based on the frequency of 0.677% Ura− Ade+ His− events among the total viable progeny in such a cross representing 8.375% of recombinants among all Ura− His− colonies (240 Ura− His− colonies among 2,969 total viable progeny, 8.083%), we would expect that 2-3 of the 32 Ura+ Ade− His+ carry both the 3083 and the 469 mutation within the ade6 locus, if all these events were generated by CO processing of a recombination internediate between the two heteroalleles. We did not observe any instances in which the ade6 locus of Ura+ Ade− His+ progeny harbored both mutations (Supplementary Fig. S1). Intragenic COs, if arising at all, are thus potentially only a minor cause in such progeny among gene conversions, which are already relatively rare events. Rather, it is simple gene conversions at single loci, which are primarily generated by mismatch repair or DNA synthesis during DSB repair26, that are responsible.
MutSα and MutLα are strong negative modulators of recombination frequency specifically at short intragenic intervals
Potential candidates for genetic pathways modulating recombination frequency at intragenic intervals of different lengths are MutS-MutL complexes, which bind to heteroduplex DNA and repair mismatches11. Sz. pombe has a streamlined nuclear mismatch repair system consisting of MutSα (Msh2-Msh6), MutSβ (Msh2-Msh3), and a single MutL (MutLα, Mlh1-Pms1); there is also a mitochondrial MutS protein called Msh127. Importantly, the meiotic pro-crossover factors MutSγ (Msh4-Msh5), the meiosis-specific MutLγ component Mlh3, and Mlh2 – a MutLβ-homolog and a modulator of meiotic gene conversion tract length – are all missing in fission yeast28,29. This suggests that Sz. pombe is a suitable model to study the role of MutSα/β-MutLα during meiosis without potential crosstalk from MutSγ-MutLγ pro-crossover factors30.
At small intragenic intervals the absence of MutSα and/or MutLα causes a substantial increase in intragenic recombination frequency (Fig. 3A, Supplementary Fig. S2). This relationship shows an inverse correlation, i.e. the shorter the intragenic interval the higher the increase. This ranges from a ~70-fold increase at the ade6-149 × ade6-3049 (33 bp) interval, via a ~35-fold one at ade6-3049 × ade6-51 (53 bp), to a ~10-fold augmentation at the ade6-M216 × ade6-3083 (84 bp) interval (Fig. 3A, Supplementary Fig. S2). The mutSα mutants (msh2-30, msh6Δ) and the mutLα mutants (mlh1Δ, pms1-16) displayed similar frequencies of intragenic recombination to each other, and the msh2-30 mlh1Δ double mutant is not discernible from either single mutant (Fig. 3A), indicating that MutSα and MutLα work in the same pathway. Deleting mutSβ (msh3) is of no consequence at the ade6-M216 × ade6-3083 interval (Fig. 3A; p = 0.613 against wild type, two-tailed Mann-Whitney U), likely because all the ade6 mutations tested are substitution mutations, and MutSβ only recognizes insertion/deletion loop mismatches larger than 2 nucleotides11. At larger intragenic intervals, there seems to be little or no role for MutSα-MutLα in limiting recombination events. A moderate, but mostly non-significant, tendency of lower intragenic recombination frequency can be observed (Fig. 3B, Supplementary Fig. S2). Altogether, these data show that MutSα-MutLα has a strong anti-recombinogenic role at small intragenic intervals, but seemingly no substantial role in determining recombination outcome at large intragenic intervals.

MutSα and MutLα, but not MutSβ, are major modulators of the gene conversion (GC) rate, and of the crossover (CO) frequency among GC events. (A,B) Frequency of GC in wild type (WT), msh2, msh3, msh6, mlh1, and pms1 mutants (A) at the intragenic 84 bp interval ade6-M216 × ade6-3083: UoA110 × UoA100 (WT, n = 12), UoA478 × UoA476 (msh2-30, n = 6), UoA494 × UoA492 (msh3Δ, n = 6), UoA482 × UoA480 (msh6Δ, n = 6), UoA364 × UoA361 (mlh1Δ, n = 8), UoA407 × UoA405 (pms1-16, n = 5), UoA828 × UoA830 (msh2-30 mlh1Δ, n = 6); (B) at the intragenic 1,320 bp interval ade6-3083 × ade6-469: ALP733 × ALP731 (WT, n = 20), UoA477 × UoA479 (msh2-30, n = 6), UoA493 × UoA495 (msh3Δ, n = 6), UoA481 × UoA483 (msh6Δ, n = 6), UoA362 × UoA371 (mlh1Δ, n = 11), UoA406 × UoA410 (pms1-16, n = 6), UoA827 × UoA829 (msh2-30 mlh1Δ, n = 6). (C,D) Frequency of CO between his3+-aim and ura4+-aim2 associated with GC events at ade6 in wild type (WT), msh2, msh3, msh6, mlh1, and pms1 mutants (C) at the intragenic 84 bp interval ade6-M216 × ade6-3083: strains as in (A); (D) at the intragenic 1,320 bp interval ade6-3083 × ade6-469: strains as in (B). n indicates the number of independent crosses. For details of data see Supplementary Table S3.
Mutating mutSα-mutLα genes increases CO frequency among gene conversion events (Fig. 3C,D, Supplementary Fig. S3) and/or changes the distribution of recombinant classes (Supplementary Fig. S4). Both long and short intragenic intervals involving the ade6-3083 allele showed increases in associated CO frequency in comparison to wild type, albeit this trend was not statistically significant in all cases (Fig. 3C,D, Supplementary Fig. S3). This trend makes the share of COs among gene conversion events independent of the length of the interval (compare Fig. 2B with Fig. 3C,D, Supplementary Fig. S3).
Interestingly, there is also a substantial shift in CO classes among gene conversion events from mostly Ura− His− to mainly Ura+ His+ in mutSα-mutLα mutants at the short intervals ade6-M216 × ade6-3083 and ade6-149 × ade6-3049, but not at the short ade6-51 × ade6-3049 interval (Supplementary Fig. S4A–C). At long intervals (ade6-3083 × ade6-469, ade6-M375 × ade6-469, ade6-M216 × ade6-3049) this shift is not observed (Supplementary Fig. S4D–F). The change in CO classes among gene conversion events at the short intervals (ade6-M216 × ade6-3083 and ade6-149 × ade6-3049) is not a consequence of selective survival or the formation of diploid or disomic spores, because mutSα-mutLα mutants have a spore viability similar to wild type, and the extent of the phenotype is the same in several different mutants (Supplementary Table S3). As with intragenic recombination frequency, the mutSβ-deletion msh3Δ behaves just like wild type for CO outcome (Fig. 3C,D; p = 0.439 against wild type, two-tailed Mann-Whitney U; Supplementary Figs S4A and S4D).
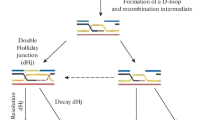
The observed effects of different parental and recombinant classes amongst progeny having undergone a gene conversion event can be explained by envisioning a DSB 5′ or 3′ of a point mutation leading to a recombination intermediate (D-loop, Holliday junction), which will then be processed immediately at the break site, or ends up somewhat removed from the initial break site by multiple consecutive invasion steps, by branch migration, or both31,32,33. The genetic makeup of the progeny is, therefore, a compound result of processing distinct recombination intermediates in different ways. The genetic composition of wild-type and mutant progeny resulting from the meiotic recombination assays can be explained as different combinations of scenarios suggested previously34. For example, recombination between ade6-3083 and ade6-M216, which gives rise to mainly Ura− Ade+ His+ NCOs and Ura− Ade+ His− COs, may be explained by the model in Fig. 4A. In this model, a bias in favour of Ura− Ade+ His− COs stems from strand exchange/branch migration being constrained to within the region defined by the ade6-3083 – ade6-M216 interval and resolution of the recombination intermediate occurring by D-loop cleavage (Fig. 4A,C). Ura− Ade+ His+ NCOs and additional Ura− Ade+ His− COs come from HJ resolution (Fig. 4A,C).

Possible scenarios for CO/NCO recombination events creating Ade+ progeny from crosses with different ade6 heteroalleles and ura4+-aim2 and his3+-aim as flanking markers. (A,B) The two black lines represent double-stranded DNA of one chromatid; chromatids not involved in the depicted recombination event are omitted for clarity. Positions of the hotspot and non-hotspot alleles are indicated in red and light blue, respectively. (A) Predominant situation in wild type, where Ade+ CO recombinants are mostly Ura− His−. (B) Situation explaining the Ura+ Ade+ His+ progeny observed in some mutSα-mutLα mutant crosses. Extensive branch migration and/or multiple invasion events could cause the D-loop or Holliday Junction (HJ) eventually being established left of the non-hotspot allele. Subsequent processing will generate Ura+ Ade+ His+ CO progeny at a high frequency. (C) Frequency of possible recombination outcomes in crosses involving two ade6 heteroalleles and flanking markers (ura4+-aim2 and his3+-aim) as shown in (A) and (B).
We also considered whether this alteration of recombination outcome at ade6-M216 × ade6-3083 in mutSα-mutLα mutants, which leads to relatively few Ade+ His− Ura− COs and a big increase in the proportion of Ade+ His+ Ura+ COs (Fig. 3, Supplementary Fig. S4A), might have something to do with the complexity of the ade6-3083 allele. This allele consists of multiple substitution mutations and can potentially form a C/C-mismatch in the heteroduplex DNA during strand exchange that is less efficiently repaired during meiosis than other mismatches35. However, as mentioned above, a moderate shift of CO recombinant classes among intragenic events can also be seen at another small interval, ade6-149 × ade6-3049 (Supplementary Fig. S4B). Unlike ade6-3083, ade6-3049 contains only a single nucleotide difference (Supplementary Table S1) and, therefore, the complexity of a given ade6 allele is unlikely to be the critical factor affecting the shift in CO recombinant class. This is complicated by the fact that a third small interval, ade6-51 × ade6-3049, does not show this shift between CO recombinant categories, similar to long intervals (Supplementary Fig. S4C–F). We think that a deficit in heteroduplex rejection and mismatch repair, caused by loss of msh2, could results in strand exchange/branch migration extending beyond the non-hotspot mutation (i.e. ade6-M216 or ade6-149) prior to D-loop cleavage/HJ resolution, with the base-pair mismatches in the recombinant chromosomes remaining unrepaired. Together, these altered features could explain the increase in Ade+ His+ Ura+ COs at the ade6-M216 × ade6-3083 and ade6-149 × ade6-3049 intervals in mutSα-mutLα mutant crosses (Fig. 4B,C). However, why ade6-51 × ade6-3049 would not show this behavior remains unclear; potentially the positioning of the DSBs in relation to the hotspot mutations could play a role here.
Recombination outcome in a msh2Δ in Saccharomyces cerevisiae has also been shown to be more complex than in wild type36,37. Intriguingly, in S. cerevisiae the action of Msh2 seems to be restricted to class I COs, which are subjected to CO interference, whereas Mus81-dependent class II COs are unchanged in msh2Δ37. Sz. pombe operates only a class II CO pathway via Mus81-processing, completely lacking a class I CO pathway. Nevertheless, the absence of Msh2 in fission yeast has a profound effect on CO frequency, and the way recombination intermediates are processed (Fig. 3, Supplementary Fig. S4).
Fml1 is a negative modulator of CO frequency among gene conversion events independent of the distance between point mutations
The DNA helicases, Fml1 and Rqh1, are also prime candidates for modulating recombination frequency at intragenic intervals of different lengths12,38. However, Fml1 apparently does not modulate gene conversion levels, as at all intragenic intervals tested, fml1Δ is similar to wild type (Fig. 5A,B, Supplementary Fig. S5A). In contrast, the RecQ-family DNA helicase Rqh1 is required for wild-type levels of gene conversion12. The deletion of rqh1 reduces gene conversion frequency to about a third of wild-type percentage at short (ade6-M216 × ade6-3083, ade6-3049 × ade6-469) intervals, and to about a tenth of wild-type frequency at the long ade6-3083 × ade6-469 interval (Fig. 5A,B, Supplementary Fig. S5).

The RecQ-family helicase Rqh1, but not the FANCM-type helicase Fml1, is a major modulator of the gene conversion (GC) rate. Rqh1 and Fml1 are major modulators of crossover (CO) frequency among GC events. Frequency of GC in WT, fml1, and rqh1 deletions (A) at the intragenic 84 bp interval ade6-M216 × ade6-3083: UoA110 × UoA100 (WT, n = 12), UoA450 × UoA447 (fml1Δ, n = 9), UoA502 × UoA499 (rqh1Δ, n = 6); (B) at the intragenic 1,320 bp interval ade6-3083 × ade6-469: ALP733 × ALP731 (WT, n = 20), ALP1133 × MCW4718 (fml1Δ, n = 15), ALP781 × ALP780 (rqh1Δ, n = 10). Frequency of CO between his3+-aim and ura4+-aim2 associated with GC events at ade6 in WT, fml1, and rqh1 deletions (C) at the intragenic 84 bp interval ade6-M216 × ade6-3083: strains as in (A); (D) at the intragenic 1,320 bp interval ade6-3083 × ade6-469: strains as in (B). n indicates the number of independent crosses. For details of data see Supplementary Table S3.
As with long intervals12, fml1Δ results in a ~10 percentage point increase of CO frequency among gene conversion events at short intervals (Fig. 5C,D, Supplementary Fig. S5). The absence of Rqh1 induces moderate increases in CO levels among gene conversion events at the 84 bp ade6-M216 × ade6-3083 and the 1,320 bp ade6-3083 × ade6-469 interval, which are not statistically significant (Fig. 5C,D). However, at the 254 bp ade6-3049 × ade6-469 interval CO frequency among ade6+ events is raised by 17 percentage points in rqh1Δ (p = 3.72 × 10−9 against wild type, two-tailed Mann-Whitney U) (Supplementary Fig. S5). The ade6-3083 allele contains multiple point mutations and thus represents a more complex situation than the ade6-3049 allele, which only harbors a single point mutation. Fml1 can seemingly drive NCO pathway(s) independently of the complexity of the underlying DNA sequence, because it has the same effect in crosses with complex and single-mutation alleles. In contrast, Rqh1 can apparently fulfill this role only at the simple ade6-3049 allele.
In Sz. pombe Fml1 has been shown to specifically limit CO formation during the late CO/NCO-decision12. Fml1 acts as a promotor of NCOs, likely by driving late recombination intermediates into the SDSA pathway, after strand invasion and DNA synthesis has happened. In accordance with this, absence of fml1 leads to an increase in CO among Ade+ gene conversion events, but has little effect on intragenic recombination itself (Fig. 5, Supplementary Fig. S5)12. This role is independent of the size of the intragenic interval, with Fml1 driving 10–12% of NCO recombination in any case.
The deletion of rqh1 has a very strong meiotic phenotype, leading to reductions in intragenic recombination, CO, and spore viability (Fig. 5, Supplementary Fig. S5). This on its own would indicate an early role in promoting strand exchange and/or DSB resection, but then Rqh1 additionally is capable of promoting NCO formation among ade6+ events at some intragenic intervals during later stages of recombination (Fig. 5, Supplementary Fig. S5). Most likely this is due to Rqh1 actually performing the following functions: (I) promotion of interhomolog recombination events, probably in cooperation with Rad55–57 and Rlp1-Rdl1-Sws1, but independently of Sfr1-Swi5,34 potentially also by providing longer resection tracts39; (II) dismantling D-loops, this enables the release of break ends to search for homology elsewhere, starts cycles of multiple consecutive invasion steps, and provides opportunities for Fml1 to drive NCO formation via SDSA; and (III) branch migration of established D-loops and Holliday junctions, thereby promoting heteroduplex DNA formation further away from the break site38.
Overall, these data show that Fml1 has likely no role in modulating gene conversion levels, but drives NCO formation downstream after successful strand invasion and DNA synthesis. Rqh1 promotes intragenic recombination, but also has moderate anti-recombinogenic activity in CO formation among gene conversion events.
In conclusion, factors directly involved in generating CO and NCO recombinants during meiosis have been identified and characterized in recent years12,13,14,15,22, and several inroads have been made in understanding how template choice is regulated and executed during meiotic recombination10,34. However, we still only have a basic understanding of how underlying DNA sequence polymorphisms influence meiotic recombination outcomes. This is critically important for understanding recombination event distribution in natural populations, where any two parental genomes will be littered with sequence polymorphisms. Here, we demonstrate that specific DNA sequence differences between the two homologs strongly impact on which outcome is achieved, and that this is largely driven by the action of the MutS-MutL complex. This highlights the importance of the interplay between cis- and trans-factors in shaping the genetic diversity of a given population.
Material and Methods
Bacterial and yeast strains and culture conditions
E. coli strains were grown on LB and SOC media – where appropriate containing 100 µg/ml Ampicillin40. Competent cells of E. coli strains NEB10®-beta (New England BioLabs Inc., Ipswich, MA, USA), and XL1-blue (Agilent Technologies, Santa Clara, CA, USA) were transformed following the protocols provided by the manufacturers. Sz. pombe strains used for this study are listed in Supplementary Table S4. Yeast cells were cultured on yeast extract (YE), and on yeast nitrogen base glutamate (YNG) agar plates containing the required supplements (concentration 250 mg/l on YE, 75 mg/l on YNG). Crosses were performed on malt extract (ME) agar containing supplements at a final concentration of 50 mg/l41.
Different ade6 hotspot and non-hotspot sequences (Supplementary Table S1) were introduced by crossing the respective mutant ade6 strain with ade6+ strains carrying the ura4+ and his3+ artificially introduced markers (aim) (UoA95, UoA96, UoA97, UoA98)22. The point mutations in the ade6 alleles were verified by Sanger DNA sequencing (Source BioScience, Nottingham, UK) (Supplementary Table S1).
Using an established marker swap protocol42 the natMX6-marked rqh1Δ-G1 was derived from an existing rqh1Δ::kanMX6 allele43, creation of the natMX6-marked pms1-16 insertion mutant allele has been described previously44.
Marker cassettes to delete msh3, and msh6, and to partially delete msh2 were constructed by cloning targeting sequences of these genes into pFA6a-kanMX6, pAG25 (natMX4), and pAG32 (hphMX4), respectively, up- and downstream of the dominant drug resistance marker45,46. The targeting cassettes were released from plasmids (pALo130, pALo132, pALo134) generated for this purpose by a restriction digest, and transformed into the strains FO652 (msh2 and msh6) and ALP729 (msh3). For specifics of strain and plasmid construction, please refer to Supplementary Materials. Plasmid sequences are available on figshare (https://dx.doi.org/10.6084/m9.figshare.6949274).
Transformation of yeast strains was performed using an established lithium-acetate procedure47. All plasmid constructs were verified by DNA sequencing (Source BioScience plc, Nottingham, UK).
All DNA modifying enzymes (high-fidelity DNA polymerase Q5, restriction endonucleases, T4 DNA ligase) were supplied by New England BioLabs. Oligonucleotides were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA).
Genetic and molecular assays
Determination of spore viability by random spore analysis and the meiotic recombination assay have been previously described in detail22,41.
To test whether intragenic COs exist, genomic DNA of Ura+ Ade− His+ progeny from an ade6-3083 × ade6-469 (ALP733 × ALP731) cross was used to PCR-amplify the ade6 locus (oligonucleotides oUA219 5′-AAAGTTGCATTTCACAATGC-3′ and oUA66 5′-GTCTATGGTCGCCTATGC-3′) for Sanger sequencing (Eurofins Scientific, Brussels, Belgium) with oUA219, oUA66, or nested oligonucleotides oUA779 5′-CTCATTAAGCTGAGCTGCC-3′ and oUA780 5′-AAGCTCTCCATAGCAGCC-3′.
Data presentation and statistics
Raw data is available on figshare (https://doi.org/10.6084/m9.figshare.6949274). Line graphs were produced using Microsoft Excel 2016 (version 16.0.4638.1000, 32-bit). Box-and-whisker plots were created in R (version i386, 3.0.1) (http://www.r-project.org/) using the standard settings of the boxplot() function34. The lower and upper ‘hinges’ of the box represent the first and third quartile, and the bar within the box indicates the median (=second quartile). The ‘whiskers’ represent the minimum and maximum of the range, unless they differ more than 1.5-times the interquartile distance from the median. In the latter case, the borders of the 1.5-times interquartile distance around the median are indicated by the ‘whiskers’ and values outside this range (‘outliers’) are shown as open circles. R was also used to compute Kruskal-Wallis test and Tukey’s Honest Significant Differences employing the kruskal.test() and TukeyHSD() functions, respectively. Mann-Whitney U tests were performed as previously described34.
References
Lam, I. & Keeney, S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb. Perspect. Biol. 7, a016634 (2015).
Wahls, W. P. & Davidson, M. K. New paradigms for conserved, multifactorial, cis-acting regulation of meiotic recombination. Nucleic Acids Res. 40, 9983–9989 (2012).
Kon, N., Krawchuk, M. D., Warren, B. G., Smith, G. R. & Wahls, W. P. Transcription factor Mts1/Mts2 (Atf1/Pcr1, Gad7/Pcr1) activates the M26 meiotic recombination hotspot in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 94, 13765–13770 (1997).
Yamada, S. et al. Correlation of meiotic DSB formation and transcription initiation around fission yeast recombination hotspots. Genetics 206, 801–809 (2017).
Steiner, W. W. & Smith, G. R. Optimizing the nucleotide sequence of a meiotic recombination hotspot in Schizosaccharomyces pombe. Genetics 169, 1973–1983 (2005).
Yamada, S., Ohta, K. & Yamada, T. Acetylated Histone H3K9 is associated with meiotic recombination hotspots, and plays a role in recombination redundantly with other factors including the H3K4 methylase Set1 in fission yeast. Nucleic Acids Res. 41, 3504–3517 (2013).
Hunter, N. Meiotic recombination: the essence of heredity. Cold Spring Harb. Perspect. Biol. 7, a016618 (2015).
Humphryes, N. & Hochwagen, A. A non-sister act: Recombination template choice during meiosis. Exp. Cell Res. 329, 53–60 (2014).
Lorenz, A. Modulation of meiotic homologous recombination by DNA helicases. Yeast 34, 195–203 (2017).
Hong, S. et al. The logic and mechanism of homologous recombination partner choice. Mol. Cell 51, 440–453 (2013).
Surtees, J. A., Argueso, J. L. & Alani, E. Mismatch repair proteins: Key regulators of genetic recombination. Cytogenet. Genome Res. 107, 146–159 (2004).
Lorenz, A. et al. The fission yeast FANCM ortholog directs non-crossover recombination during meiosis. Science 336, 1585–1588 (2012).
Crismani, W. et al. FANCM limits meiotic crossovers. Science 336, 1588–1590 (2012).
De Muyt, A. et al. BLM helicase ortholog Sgs1 is a central regulator of meiotic recombination intermediate metabolism. Mol. Cell 46, 43–53 (2012).
Lukaszewicz, A., Howard-Till, R. A. & Loidl, J. Mus81 nuclease and Sgs1 helicase are essential for meiotic recombination in a protist lacking a synaptonemal complex. Nucleic Acids Res. 41, 9296–9309 (2013).
Agostinho, A. et al. Combinatorial regulation of meiotic holliday junction resolution in C. elegans by HIM-6 (BLM) helicase, SLX-4, and the SLX-1, MUS-81 and XPF-1 nucleases. PLoS Genet. 9, e1003591 (2013).
Schvarzstein, M. et al. DNA helicase HIM-6/BLM both promotes MutSγ-dependent crossovers and antagonizes MutSγ-independent interhomolog associations during Caenorhabditis elegans meiosis. Genetics 198, 193–207 (2014).
Hatkevich, T. et al. Bloom syndrome helicase promotes meiotic crossover patterning and homolog disjunction. Curr. Biol. 27, 96–102 (2017).
Gutz, H. Site specific induction of gene conversion in Schizosaccharomyces pombe. Genetics 69, 317–337 (1971).
Zahn-Zabal, M. & Kohli, J. The distance-dependence of the fission yeast ade6-M26 marker effect in two-factor crosses. Curr. Genet. 29, 530–536 (1996).
Fox, M. E., Virgin, J. B., Metzger, J. & Smith, G. R. Position- and orientation-independent activity of the Schizosaccharomyces pombe meiotic recombination hot spot. M26. Proc. Natl. Acad. Sci. USA 94, 7446–7451 (1997).
Osman, F., Dixon, J., Doe, C. L. & Whitby, M. C. Generating crossovers by resolution of nicked Holliday junctions: a role for Mus81-Eme1 in meiosis. Mol. Cell 12, 761–774 (2003).
Lorenz, A., West, S. C. & Whitby, M. C. The human Holliday junction resolvase GEN1 rescues the meiotic phenotype of a Schizosaccharomyces pombe mus81 mutant. Nucleic Acids Res. 38, 1866–1873 (2010).
Hyppa, R. W. & Smith, G. R. Crossover invariance determined by partner choice for meiotic DNA break repair. Cell 142, 243–255 (2010).
Cromie, G. A. et al. Single Holliday junctions are intermediates of meiotic recombination. Cell 127, 1167–1178 (2006).
Holliday, R. A mechanism for gene conversion in fungi. Genet. Res. 5, 282–304 (2007).
Marti, T. M., Kunz, C. & Fleck, O. DNA mismatch repair and mutation avoidance pathways. J. Cell. Physiol. 191, 28–41 (2002).
Manhart, C. M. & Alani, E. Roles for mismatch repair family proteins in promoting meiotic crossing over. DNA Repair (Amst). 38, 84–93 (2016).
Duroc, Y. et al. Concerted action of the MutLβ heterodimer and Mer3 helicase regulates the global extent of meiotic gene conversion. Elife 6, e21900 (2017).
Rogacheva, M. V. et al. Mlh1-Mlh3, a meiotic crossover and DNA mismatch repair factor,is a Msh2-Msh3-stimulated endonuclease. J. Biol. Chem. 289, 5664–5673 (2014).
Farah, J. A., Cromie, G. A. & Smith, G. R. Ctp1 and Exonuclease 1, alternative nucleases regulated by the MRN complex, are required for efficient meiotic recombination. Proc. Natl. Acad. Sci. USA 106, 9356–9361 (2009).
Piazza, A., Wright, W. D. & Heyer, W. D. Multi-invasions are recombination byproducts that induce chromosomal rearrangements. Cell 170, 760–773 (2017).
Marsolier-Kergoat, M. C., Khan, M. M., Schott, J., Zhu, X. & Llorente, B. Mechanistic view and genetic control of DNA recombination during meiosis. Mol. Cell 70, 9–20 (2018).
Lorenz, A., Mehats, A., Osman, F. & Whitby, M. C. Rad51/Dmc1 paralogs and mediators oppose DNA helicases to limit hybrid DNA formation and promote crossovers during meiotic recombination. Nucleic Acids Res. 42, 13723–13735 (2014).
Schär, P. & Kohli, J. Marker effects of G to C transversions on intragenic recombination and mismatch repair in Schizosaccharomyces pombe. Genetics 133, 825–835 (1993).
Martini, E. et al. Genome-wide analysis of heteroduplex DNA in mismatch repair-deficient yeast cells reveals novel properties of meiotic recombination pathways. PLoS Genet. 7, e1002305 (2011).
Cooper, T. J. et al. Mismatch repair impedes meiotic crossover interference. bioRxiv 480418, https://doi.org/10.1101/480418 (2018).
Cromie, G. A., Hyppa, R. W. & Smith, G. R. The fission yeast BLM homolog Rqh1 promotes meiotic recombination. Genetics 179, 1157–1167 (2008).
Osman, F., Ahn, J. S., Lorenz, A. & Whitby, M. C. The RecQ DNA helicase Rqh1 constrains Exonuclease 1-dependent recombination at stalled replication forks. Sci. Rep. 6, 22837 (2016).
Sambrook, J. F. & Russell, D. W. Molecular Cloning: A Laboratory Manual. (Cold Spring Harbor Laboratory Press, 2000).
Sabatinos, S. A. & Forsburg, S. L. Molecular genetics of Schizosaccharomyces pombe. Methods Enzymol. 470, 759–795 (2010).
Sato, M., Dhut, S. & Toda, T. New drug-resistant cassettes for gene disruption and epitope tagging in Schizosaccharomyces pombe. Yeast 22, 583–591 (2005).
Doe, C. L., Ahn, J. S., Dixon, J. & Whitby, M. C. Mus81-Eme1 and Rqh1 involvement in processing stalled and collapsed replication forks. J. Biol. Chem. 277, 32753–32759 (2002).
Lorenz, A. New cassettes for single-step drug resistance and prototrophic marker switching in fission yeast. Yeast 32, 703–710 (2015).
Bähler, J. et al. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14, 943–951 (1998).
Goldstein, A. L. & McCusker, J. H. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15, 1541–1553 (1999).
Brown, S. D. & Lorenz, A. Single-step marker switching in Schizosaccharomyces pombe using a lithium acetate transformation protocol. Bio-protocol 6, e2075 (2016).
Acknowledgements
We are grateful to Jürg Bähler, Edgar Hartsuiker, Franz Klein, Jürg Kohli, Josef Loidl, Kim Nasmyth, Fekret Osman, Gerald R. Smith, Walter W. Steiner, and the National BioResource Project (NBRP) Japan for providing materials, and to C. Bryer, A. Mehats, and H. Rickman for technical assistance. This work was supported by the Biotechnology and Biological Sciences Research Council UK (BBSRC) [grant numbers BB/F016964/1, BB/M010996/1], the University of Aberdeen (College of Life Sciences and Medicine Start-up grant to AL), and the Wellcome Trust (Programme grant to MCW) [grant number 090767/Z/09/Z].
Author information
Authors and Affiliations
Contributions
A.L. and M.C.W. conceived this study. A.L., S.D.B., S.J.M., M.N.A. and M.J. conducted the experiments. A.L. drafted the manuscript. All authors read, revised and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brown, S.D., Mpaulo, S.J., Asogwa, M.N. et al. DNA sequence differences are determinants of meiotic recombination outcome. Sci Rep 9, 16446 (2019). https://doi.org/10.1038/s41598-019-52907-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-52907-x
- Springer Nature Limited