
Abstract
Trichopria drosophilae (Hymenoptera: Diapriidae) is an important pupal endoparasitoid of Drosophila melanogaster Meigen (Diptera: Drosophilidae) and some other fruit fly species, such as D. suzukii, a very important invasive and economic pest. Studies of T. drosophilae suggest that this could be a good biological control agent for fruit fly pests. In this research, we compared the parasitic characteristics of T. drosophilae reared in D. melanogaster (TDm) with those reared in D. hydei (TDh). TDh had a larger size than TDm. The number of maximum mature eggs of a female TDh was 133.6 ± 6.9, compared with the significantly lower value of 104.8 ± 11.4 for TDm. Mated TDh female wasp continuously produced female offspring up to 6 days after mating, compared with only 3 days for TDm. In addition, the offspring female ratio of TDh, i.e., 82.32%, was significantly higher than that of TDm, i.e., 61.37%. Under starvation treatment, TDh survived longer than TDm. TDh also survived longer than TDm at high temperatures, such as 37 °C, although they both survived well at low temperatures, such as 18 °C and 4 °C. Old-age TDh females maintained a high parasitism rate and offspring female ratio, while they were declined in old-age TDm. Overall, TDh had an advantage in terms of body size, fecundity, stress resistance ability and the parasitism rate compared with TDm. Therefore, T. drosophilae from D. hydei could improve biocontrol efficacy with enormous economic benefits in the field, especially in the control of many frugivorous Drosophilidae species worldwide.
Similar content being viewed by others


Introduction
Drosophila is a genus of flies belonging to the family Drosophilidae. Some species of Drosophila (also called fruit flies), particularly D. melanogaster, have been widely used in the research of genetics, developmental biology and human diseases1,2. However, some Drosophila species are destructive pests of agriculture, especially damaging soft fruits such as berries, cherries and wine grapes3. Fruit flies generally lay eggs in decaying fruits, and the larvae feed and develop with the fruits, which causes health risks and economic losses. Traditional chemical control methods for fruit flies have low efficiency and are harmful to public health4. Consequently, biological control with parasitoids is more sustainable and is urgently needed. Parasitic wasps constitute a major class of natural enemies of many agriculture pests and have tremendous value as biocontrol agents. Most known parasitoid wasp species attack the egg, larval or pupal stages of their hosts and they carry virulence and some other parasitic factors to modify hosts’ physiology and immunity, to change hosts’ metabolism, to destruct hosts’ endocrine and reproductive structures, and finally kill the hosts for their own development5,6,7,8. Many parasitoids are reported to attack various Drosophilidae species, and the majority of them are larval parasitoids, such as Leptopilina heterotoma, L. boulardi and Asobara tabida9,10. Recently, Trichopria drosophilae (Hymenoptera: Diapriidae), an important pupal endoparasitoid of D. melanogaster and some other fruit fly species, has been found to be an ideal natural enemy to constrain the fruit fly population because it has extremely high parasitism efficiency11,12,13,14. The life history and biological characteristics of T. drosophilae have been well studied by several groups. In 2012, Chabert et al. found that T. drosophilae was effective against many fruit fly species, including D. suzukii, a well-known invasive pest12. Female T. drosophilae emerged with a relatively high number of mature eggs, and the egg numbers increased during their first four days after eclosion. This indicates that T. drosophilae might maximize reproduction during early adult life14. Moreover, the parasitism rate of T. drosophilae is higher than that of another well-known cosmopolitan pupal parasitoid, Pachycrepoideus vindemmiae (Diptera: Pteromalidae)13. Although T. drosophilae is reported to be effective against Drosophila species under laboratory conditions, it is necessary to find the parasitoids that have the highest parasitism rate, highest female offspring numbers and longest adult longevity and which are resistant to certain stress conditions, such as food deprivation and extreme weather conditions, for the biological control purpose of augmentative release in the field.
To increase the effectiveness of parasitoids as natural enemies, female adult wasps are supplied with extra nutrient sources, such as sugars, to enhance their longevity and fecundity and subsequently, the biocontrol efficacy15,16. However, host quality can also have a major influence on the fitness and parasitic efficiency of offspring17. Lampson et al. found that different sizes of the same parasitoid had an effect on several biological characteristics, suggesting that larger parasitoids have a longer life span and greater competitiveness18. Another comparative study on the parasitism of P. vindemmiae hatching from housefly and fruit fly pupae showed a positive correlation between the size of the host and the size of the emerged offspring, as well as the longevity, the oviposition duration and other parasitic attributes19.
Based on the results of previous studies14, T. drosophilae reared on a larger sized host could be more advantageous for further biological control. Here, we used D. hydei as a substitute host, of which the pupae are significantly larger than those of D. melanogaster. Then, we compared the body size, fecundity, stress resistance ability and parasitism efficiency between the two parasitoid populations that emerged from the different hosts.
Results
The parasitoid and host size measurements
The respective pupal length and width were 4.05 ± 0.13 mm and 1.27 ± 0.04 mm for D. hydei (n = 18) and 2.93 ± 0.14 mm and 0.99 ± 0.06 mm for D. melanogaster (n = 37). The size of D. hydei was significantly larger than that of D. melanogaster (length: t = 28.57, df = 53, P < 0.01, width: t = 18.68, df = 53, P < 0.01). To investigate whether there was a correlation between the size of the hosts and their offspring, T. drosophilae was used to parasitize D. melanogaster and D. hydei pupae. The measurements indicated that the body length of TDh was significantly longer than that of TDm, in both females and males (Fig. 1A–C). The body length of female TDh was 2.41 ± 0.12 mm (n = 10), compared with 2.12 ± 0.11 mm (n = 12) for female TDm (t = 5.50, df = 20, P < 0.01). The length of male TDh was 2.21 ± 0.07 mm (n = 16), compared with 1.92 ± 0.11 mm (n = 10) for male TDm (t = 7.94, df = 24, P < 0.01). These results showed that the size of TDh was much larger than that of TDm.

(A) Body length measurements for TDh and TDm. The body length of female TDh was 2.41 ± 0.12 mm (n = 10), compared with 2.12 ± 0.11 mm (n = 12) for female TDm (t = 5.50, df = 20, P < 0.01). The body length of male TDh was 2.21 ± 0.07 mm (n = 16), compared with 1.92 ± 0.11 mm (n = 10) for male TDm (t = 7.94, df = 24, P < 0.01). Significant differences based on Student’s t-test at P < 0.05 are indicated by asterisks. (B) Photographs of male TDh and TDm. The body length of parasitoids was measured as the length from the head to the tip of the abdomen. (C) Photographs of female TDh and TDm.
Parasitism rate and offspring female ratio comparison
The results showed that this local 4-day old T. drosophilae females had an extremely high parasitism rate. Approximately 97% of the D. melanogaster pupae were successfully parasitized by TDm, and no significant difference in the parasitism rate was found between TDh and TDm (t = 1.67, df = 4, P > 0.05) females. However, the offspring female ratio of TDh, which averaged 82.32%, was significantly higher than that of TDm, which averaged 61.37% (t = 8.96, df = 4, P < 0.01) (Table 1).
The fecundity of T. drosophilae
The number of mature eggs in the ovaries of TDh and TDm females was compared among different ages (Fig. 2). The results showed that the number of mature eggs was affected by the female age for TDh (X2 = 69.06, df = 7, P < 0.01) and TDm (X2 = 51.84, df = 7, P < 0.01). The mean number was further compared among different age classes using ANOVA between two T. drosophilae groups. Interestingly, the number of mature eggs of female TDh and TDm increased until the TDm females were 96 hours old, whereas this increase persisted for an additional 48 hours for TDh. Thus, the maximum number of mature eggs of TDh (133.60 ± 6.87) was observed 144 h after emergence, while that of TDm (104.80 ± 11.44) was observed 96 h after emergence (t = 4.279, df = 7, P < 0.01; Fig. 2).

Egg maturation dynamics of female T. drosophilae when reared in two different hosts. Values are the means ± SEM. Significant differences at P < 0.05 are indicated by asterisks.
We observed that mated TDh continuously produced female offspring until 6 days after mating, compared with approximately 3 days for TDm (Fig. 3A,B). In this assay, TDm female wasps could survive 10.40 ± 0.91 days (Fig. 3B), whereas TDh female wasps survived 23.81 ± 2.40 days (Fig. 3A). The lifetime number of emerged progeny was higher for the TDh population (199.38 ± 20.94 offspring per female, n = 8) than for the TDm population (104.50 ± 20.91 offspring per female, n = 8) (t = 9.07, df = 14, P < 0.01). The number of female progeny was 73.25 ± 5.47 for TDh and 27.25 ± 7.69 for TDm (t = 13.79, df = 14, P < 0.01) (Fig. 3C).

The offspring number of single TDh and TDm female wasps. (A) The number of offspring each day (red) was calculated by adding the male (black) and female (purple) offspring numbers. (B) The number of offspring each day (blue) was calculated by adding the male (black) and female (purple) offspring numbers. (C) The total number of offspring for a single TDh and TDm female. In total, 8 TDh and 8 TDm female wasps were used in this experiment, respectively. Values are the means ± SEM. Significant differences using Student’s t-test at P < 0.05 are indicated by asterisks.
The stress resistance ability of T. drosophilae
To determine T. drosophilae stress resistance ability, TDh and TDm were treated with different environmental stresses, including starvation and high and low temperatures. Under food deprivation, the starved TDm wasps had a maximum life span of 192 hours, and half of the wasps survived 120 hours, whereas the TDh wasps had a maximum life span of 288 hours, and half of the wasps could survive at least 216 hours. The TDh wasps had a longer lifespan than the TDm wasps under starvation treatment (Log-rank test X2 = 744.30, df = 1, P < 0.01) (Fig. 4).

The survival rate of TDh and TDm during the starvation treatment. TDh had a longer survival lifespan than TDm under the starvation treatment (X2 = 744.30, df = 1, P < 0.01). Significant differences based on log-rank test (Mantel-Cox) analysis.
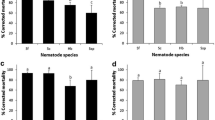
To determine how different temperatures affect T. drosophilae survival, we placed TDh and TDm into incubators at 4 °C, 18 °C, 25 °C and 37 °C. The results showed that almost all TDh and TDm wasps survived well at lower temperatures (4 °C and 18 °C). However, the survival rates of TDh were higher than those of TDm at 25 °C or 37 °C (25 °C: Log-rank test X2 = 23.09, df = 1, P < 0.01; 37 °C: Log-rank test X2 = 14.79, df = 1, P < 0.01) (Fig. 5A,B).

The survival rates of TDm (A) and TDh (B) at 4 °C, 18 °C, 25 °C and 37 °C. The results showed that almost all TDh and TDm wasps survived well at 4 °C and 18 °C. However, the survival rates of TDh were higher than those of TDm at 25 °C or 37 °C (25 °C: X2 = 23.09, df = 1, P < 0.01; 37 °C: X2 = 14.79, df = 1, P < 0.01). Significant differences were based on log-rank test (Mantel-Cox) analysis.
T. drosophilae parasitism efficiency related to age
In order to evaluate the influence of T. drosophilae age on the parasitism rate, 1-, 5-, 10-, 15-, 20-, 25-, 30- and 40-day-old wasps were used to parasitize the hosts. The results showed that both TDh and TDm had an extremely high parasitism rate at all time points; however, a significant decrease in the parasitism rate was observed for the 40-day-old TDm parasitoids compared with the 40-day-old TDh parasitoids (t = 4.94, df = 4, P < 0.01) (Fig. 6A). In accordance with the results of our fecundity experiment (Table 1, Fig. 3C), the offspring female ratio of TDh was slightly higher than that of TDm; significant differences were found between TDh and TDm at 5 days (t = 3.32, df = 4, P < 0.05), 10 days (t = 3.43, df = 4, P < 0.05) and 40 days (t = 6.87, df = 4, P < 0.01) after eclosion (Fig. 6B).

(A) The parasitic rate of TDh and TDm at different ages. There was a significant increase in the parasitism rate for the 40-day-old TDh parasitoids compared to TDm (t = 4.94, df = 4, P < 0.01) (B) The offspring female ratio of TDh and TDm at different ages. The offspring female ratio of TDh was slightly higher than that of TDm; however, significant differences were found between TDh and TDm at 5 days (t = 3.32, df = 4, P < 0.05), 10 days (t = 3.43, df = 4, P < 0.05) and 40 days (t = 6.87, df = 4, P < 0.01) after eclosion. Values are the means ± SEM. Significant differences based on Student’s t-test at P < 0.05 are indicated by asterisks.
Discussion
Assessing the capacity of the T. drosophilae parasitoid to attack Drosophilidae species and enhancing its ability to adapt to extreme environments are two of the most important steps for the release of T. drosophilae as a biological control agent. In this study, we showed that local T. drosophilae was able to successfully attack D. melanogaster and D. hydei under laboratory conditions. A previous study reported that T. drosophilae offspring reared in large hosts such as D suzukii were larger than those reared in D. melanogaster14. Because D. hydei had a larger size than D. melanogaster, we compared the offspring size that emerged from the two different hosts, and found that the size of TDh was much larger than that of TDm.
Parasitoids reared in substitute hosts would help to increase the availability of biocontrol agents20,21,22. It has also been proven that large parasitoids of the same species have longer life spans, and large females produce approximately twice as many eggs as small females18. Thus, we evaluated the different parasitic characteristics of T. drosophilae reared in D. hydei and D. melanogaster pupae. Compared to T. drosophilae populations from California14, TDm females in our experiments had a similar number of mature eggs, and the egg load increased during the first four days. However, the number of TDh mature eggs was significantly higher than that of TDm and increased during the first six days. Fecundity is the maximum potential reproductive output of a parasitoid female over its lifetime and represents one of the major parasitic characteristics. Under the test conditions, the daily fecundity of TDm and TDh decreased with increasing female age, and when provided only with D. melanogaster pupae, the adult female TDm only survived for 10 days, which is shorter than the reported T. drosophilae lifespan13. However, TDh survived for 26 days and produced more female offspring than TDm. Another interesting phenomenon was that female TDh produced female offspring for 6 days after one mating event, compared with only 3 days for TDm. T. drosophilae has a sex-determination system in which males develop from unfertilized eggs and are haploid, whereas females develop from fertilized eggs and are diploid23,24. The results suggested that size differences of T. drosophilae between males or females from different hosts may influence sperm production or storage. In mosquitos, male size does correlate with total numbers of sperm within a male and the number transferred to females25,26.
Stress resistance ability is an important factor in evaluating parasitoid fitness and biocontrol efficacy in the field. A larger sized host may provide more nutrients that are vital for parasitoid development, which may be the reason why TDh survived longer than TDm in the starvation experiments. Additionally, our data indicated that both TDh and TDm wasps survived for a long time at lower temperatures (4 °C and 18 °C). The reason for this is that the lower temperature will slow the metabolism of the wasps and can even extend their lifespan27.
During the last 10 years, D. suzukii, also known spotted wing drosophila, has become widely distributed from Asia to Europe and North and South America28,29,30,31. D. suzukii has spread rapidly to become a serious pest that economically damages soft and thin-skinned fruits in the major fruit production areas32,33,34. Extensive applications of chemical insecticides will lead to a number of problems, such as pest resistance and chemical residue. Therefore, non-toxic and environmentally friendly biological control methods are urgently needed. Some entomopathogenic nematodes and fungi have been used to kill D. suzukii adults30,35. However, control of D. suzukii populations is very limited. So far, 50 hymenopteran parasitoids are reported to infect various drosophila species which belong to four families including two larval parasitoids, Braconidae and Eucoilidae, and two pupal parasitoids, Pteromalidae and Diapriidae9. Some studies have shown that most of these larval parasitoids cannot develop in D. suzukii because of its strong immune response12. T. drosophilae is a highly effective pupal parasitoid that can attack D. suzukii and has been proven to be a potential agent for biological control14,36,37. Our study demonstrates that D. hydei-reared parasitoids show more beneficial parasitic characteristics than D. melanogaster-reared parasitoids. D. hydei has a worldwide distribution and is easy to raise in large numbers. Therefore, rearing of T. drosophilae in D. hydei pupae could be a successful biocontrol strategy, especially for the aim of reducing D. suzukii infestation.
Methods
Insect collection and rearing
D. melanogaster, D. hydei and T. drosophilae were collected from traps baited with grape fruits in May 2016 at Zijingang Campus (30.29°N, 120.08°E), Zhejiang University, Hangzhou, China, and were maintained in our laboratory at a temperature of 25 ± 1 °C, relative humidity of 50–60%, and a photoperiod of 16 h: 8 h (L: D) inside plastic bottles (approximately 10 cm in length and 5 cm in diameter). Both D. melanogaster and D. hydei were maintained on a standard cornmeal/molasses/agar medium38. T. drosophilae colonies were maintained on D. melanogaster pupae, and the adult wasps were provided with apple juice/agar medium (27 g agar, 33 g brown sugar and 330 ml pure apple juice in 1000 ml diluted water).
The parasitoid and host size measurements
D. melanogaster and D. hydei pupae, as the different hosts, were parasitized by T. drosophilae. For convenience, T. drosophilae that emerged from D. hydei and D. melanogaster pupae were called TDh and TDm, respectively. The TDh and TDm adults and the pupae of their hosts were imaged using a KEYENCE VHX-2000C digital microscope system (Osaka, Japan). The body length and width of 18 D. hydei pupae and 37 D. melanogaster pupae were measured using KEYENCE VHX-2000C software. The length of the hind tibia or the length of the whole body is usually used as a proxy for the size of parasitoid wasps14,39. Here, body lengths of 10 female and 16 male TDh and 12 female and 10 male TDm were measured.
Parasitism rate and offspring female ratio comparison
To compare the parasitism rate and offspring female ratio of TDh and TDm, D. melanogaster pupae were parasitized by 4-day-old TDh and TDm similar to a previous study14 at a wasp/host ratio of 1:10 for 24 hours. This experiment was performed three times, and 200, 120 and 120 D. melanogaster host pupae were exposed to TDh and TDm. The same approach was applied to compare TDh and TDm at different ages. After eclosion, TDh and TDm adult females were maintained on apple juice wasp food at 25 °C in an incubator without hosts. Then, 1-, 5-, 10-, 15-, 20-, 25-, 30- and 40-day-old TDh and TDm female wasps were collected to parasitize D. melanogaster pupae after fully mating with young TDh and TDm males, respectively, for 24 hours. Three replicates were performed for the experiments, and 5 females and 30 host pupae were used in each experiment. After being infected, the host pupae were kept in a 25 °C incubator until the wasps emerged. The parasitism rate and offspring female ratio of the wasps were calculated using the following formulas: parasitism rate = (the number of hosts − the number of emerged flies)/the number of hosts; offspring female ratio = the number of female parasitoids/the number of total emerged parasitoids.
The fecundity and stress resistance ability of T. drosophilae
The egg load of a female parasitoid wasp
The newly emerged male and female wasps were collected and placed in plastic bottles containing apple juice wasp food without hosts. To compare the maximum egg load between TDh and TDm, ovaries of 12-, 24-, 48-, 72-, 96-, 144-, 192- and 240-h-old female T. drosophilae adults were dissected in 1 × PBS buffer, pH 7.4. Ten female wasps for each category were dissected, and the mature eggs were counted at each time point. An egg was considered mature based on criteria used in a previous study14: the chorion of a mature egg is smooth, thin and transparent, and the developing embryo is visible, while immature eggs lack these characteristics and are attached to each another.
The offspring of a single female wasp
To compare the offspring numbers of TDh and TDm, a fully mated female was allowed to parasitize 150 two-day old D. melanogaster pupae for 24 hours at 25 °C. Then, the host pupae were replaced by a new batch of 150 pupae the following day until the female adult died. The total number of offspring from single females was counted as the number of emerged wasps, including males and females. In total, 8 TDh and 8 TDm female wasps were used in this experiment, respectively.
Starvation and high and low temperature tolerances
One hundred newly emerged wasps of TDh and TDm (50 females, 50 males) were reared in an empty plastic bottle without any food at 18 °C for the starvation treatment. For the high and low temperature tolerance experiment, 100 newly emerged wasps of TDh and TDm (50 females, 50 males) were reared on apple juice wasp food in incubators at 4 °C, 18 °C, 25 °C and 37 °C. The survival rate (the number of surviving wasps/100) was calculated every 12 hours for the starvation treatment and daily for the high and low temperature tolerance analysis. Three replicates were performed for each experiment.
Data analysis and statistics
The effects of female age on the number of mature eggs were analysed using a generalized linear model (GLM) and the mean number of mature eggs in different age classes were further compared using analysis of variance (ANOVA). Log-rank tests (Mantel-Cox) were performed to analyse trends in the survival rate during the environmental stresses, i.e., starvation and high and low temperatures. Student’s t-test was used to compare the body length or body width of parasitoids and hosts, the parasitism rate and offspring female ratio, as well as the fecundity of female parasitoid wasps. Statistical analyses were performed using GraphPad Prism version 7.0a (Graphpad Software, San Diego, CA) and SPSS software 25.0 (SPSS Inc., Chicago, IL). Error bars indicate the standard error of the mean (SEM), and all data sets are expressed as the mean ± SEM. Significant differences between groups were determined by the P-value and are marked with one asterisk for P < 0.05 and two asterisks for P < 0.01.
References
Mackay, T. F. C. Mutations and quantitative genetic variation: lessons from Drosophila. Philos. Trans. R. Soc. Lond B Biol. Sci. 365, 1229–1239 (2010).
Hales, K. G., Korey, C. A., Larracuente, A. M. & Roberts, D. M. Genetics on the fly: a primer on the Drosophila model system. Genetics 201, 815–842 (2015).
Lee, J. C. et al. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manage. Sci. 67, 1358–1367 (2011).
Bruck, D. J. et al. Laboratory and field comparisons of insecticides to reduce infestation of Drosophila suzukii in berry crops. Pest Manage. Sci. 67, 1375–1385 (2011).
Pennacchio, F. & Strand, M. R. Evolution of developmental strategies in parasitic hymenoptera. Annu. Rev. Entomol. pp. 233–258 (2006).
Shi, M. & Chen, X. X. Progress in study on regulation of insect host physiology by parasitoids in China. Chin. J. Biol. Control 31, 620–637 (2015).
Zhao, W. et al. Comparative transcriptome analysis of venom glands from Cotesia vestalis and Diadromus collaris, two endoparasitoids of the host Plutella xylostella. Sci. Rep. 7, 1298, https://doi.org/10.1038/s41598-017-01383-2 (2017).
Gao, F. et al. Cotesia vestalis teratocytes express a diversity of genes and exhibit novel immune functions in parasitism. Sci. Rep. 6, 26967, https://doi.org/10.1038/srep26967 (2016).
Carton, Y., Boule’treau, B., Van Alphen, J. J. M. & Van Lenteren, J. C. The Drosophila parasitic wasps. In: Ashburner, M., Carson, H. L. & Thompson, J. N. (eds) The genetics and biology of Drosophila, vol 3. Academic Press, London, pp 347–394 (1986).
Fleury, F., Gibert, P., Ris, N. & Allemand, R. Ecology and life history evolution of frugivorous Drosophila parasitoids. In: Prevost, G. (Ed.), Advances in Parasitology, Vol 70: Parasitoids of Drosophila, pp. 3–44 (2009).
Schlenke, T. A., Morales, J., Govind, S. & Clark, A. G. Contrasting infection strategies in Generalist and specialist wasp parasitoids of Drosophila melanogaster. PLoS Path. 3, 1486–1501 (2007).
Chabert, S., Allemand, R., Poyet, M., Eslin, P. & Gibert, P. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest. Drosophila suzukii. Biol Control 63, 40–47 (2012).
Rossi Stacconi, M. V. et al. Host stage preference, efficacy and fecundity of parasitoids attacking Drosophila suzukii in newly invaded areas. Biol. Control 84, 28–35 (2015).
Wang, X. G., Kacar, G., Biondi, A. & Daane, K. M. Life-history and host preference of Trichopria drosophilae, a pupal parasitoid of spotted wing drosophila. Bio Control 61, 387–397 (2016).
Tian, J. C. et al. Different Performance of Two Trichogramma (Hymenoptera: Trichogrammatidae) Species Feeding on Sugars. Environ Entomol. 45, 1316–1321 (2016).
Harvey, J. A. et al. The effect of different dietary sugars and honey on longevity and fecundity in two hyperparasitoid wasps. J Insect Physiol. 58, 816–23 (2012).
Liu, Z. D., Xu, B. B., Li, L. & Sun, J. H. Host-size mediated trade-off in a parasitoid Sclerodermus harmandi. Plos One 6(8), e23260 (2011).
Lampson, L. J., Morse, J. G. & Luck, R. F. Host selection, sex allocation, and host feeding by Metaphycus helvolus (Hymenoptera: Encyrtidae) on Saissetia oleae (Homoptera: Coccidae) and its effect on parasitoid size, sex, and quality. Environ. Entomol. 25, 283–294 (1996).
Zhou, H. F. et al. Comparative studies on parasitism of Pachycrepoideus vindemmiae (Rondani) reared on housefly and fruit fly pupae. Chin. J. Appl. Entomol. 51, 194–199 (2014).
Powell, W. & Wright, A. F. The abilities of the aphid parasitoids Aphidius ervi Haliday and A. rhopalosiphi De Stefani Perez (Hymenoptera: Braconidae) to transfer between different known host species and the implications for the use of alternative hosts in pest control strategies. Bull. Entomol. Res. 78, 683–693 (1988).
Bribosia, E., Bylemans, D., Migon, M. & Van Impe, G. In-field production of parasitoids of Dysaphis plantaginea by using the rowan aphid Dysaphis sorbi as substitute host. Bio Control 50, 601–610 (2005).
Tunca, H., Venard, M., Colombel, E.-A. & Tabone, E. A new substitute host and its effects on some biological properties of Ooencyrtus kuvanae. Bull. Entomol. Res. 107, 742–748 (2017).
Weiblen, G. D. How to be a fig wasp. Annu. Rev. Entomol. 47, 299–330 (2002).
Beukeboom, L. W. & van de Zande, L. Genetics of sex determination in the haplodiploid wasp Nasonia vitripennis (Hymenoptera: Chalcidoidea). J. Genet. 89, 333–339 (2010).
Ponlawat, A. & Harrington, L. C. Age and body size influence male sperm capacity of the dengue vector Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 44, 422–426 (2007).
Ponlawat, A. & Harrington, L. C. Factors associated with male mating success of the dengue vector mosquito, Aedes aegypti. Am. J. Trop. Med. Hyg. 80, 395–400 (2009).
Colinet, H., Boivin, G. & Hance, T. Manipulation of parasitoid size using the temperature-size rule: fitness consequences. Oecologia 152, 425–433 (2007).
Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manage. Sci. 67, 1352–1357 (2011).
Cini, A. et al. Tracking the invasion of the alien fruit pest Drosophila suzukii in Europe. J. Pest Sci. 87, 559–566 (2014).
Woltz, J. M., Donahue, K. M., Bruck, D. J. & Lee, J. C. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 139, 759–770 (2015).
Asplen, M. K. et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): a global perspective and future priorities. J. Pest Sci. 88, 469–494 (2015).
Rota-Stabelli, O., Blaxter, M. & Anfora, G. Drosophila suzukii. Curr. Biol. 23, R8–R9 (2013).
Atallah, J., Teixeira, L., Salazar, R., Zaragoza, G. & Kopp, A. The making of a pest: the evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. Biol. Sci. 281, 20132840 (2014).
Hamby, K. A. et al. Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 89, 605–619 (2016).
Naranjo-Lazaro, J. M. et al. Susceptibility of Drosophila suzukii Matsumura (Diptera: Drosophilidae) to entomophatogenic fungi. Southwest. Entomol. 39, 201–203 (2014).
Mazzetto, F. et al. Drosophila parasitoids in northern Italy and their potential to attack the exotic pest Drosophila suzukii. J. Pest Sci. 89, 837–850 (2016).
Rossi Stacconi, M. V. et al. Comparative life history traits of indigenous Italian parasitoids of Drosophila suzukii and their effectiveness at different temperatures. Biol. Control 112, 20–27 (2017).
Huang, J., Reilein, A. & Kalderon, D. Yorkie and Hedgehog independently restrict BMP production in escort cells to permit germline differentiation in the Drosophila ovary. Development. 144, 2584–2594 (2017).
Gao, S. et al. Relationships between Body Size and Parasitic Fitness and Offspring Performance of Sclerodermus pupariae Yang et Yao (Hymenoptera: Bethylidae). PLoS One. 11, e0156831 (2016).
Acknowledgements
We thank the anonymous reviewers for many useful comments. This research was supported by the National Key R&D Program of China (2017YFD0200400), the National Science Fund for Excellent Young Scholars (31622048), the National Science Foundation of China (31672079) and the National Thousand-Young-Talents Program of China.
Author information
Authors and Affiliations
Contributions
J.N.C. and J.H.H. conceived and designed the experiments; J.N.C., S.C.Z., Y.W. and M.S. performed the experiments and analysed the data; J.N.C., J.H.H. and X.X.C. wrote and revised the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, J., Zhou, S., Wang, Y. et al. Biocontrol characteristics of the fruit fly pupal parasitoid Trichopria drosophilae (Hymenoptera: Diapriidae) emerging from different hosts. Sci Rep 8, 13323 (2018). https://doi.org/10.1038/s41598-018-31718-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-31718-6
- Springer Nature Limited
This article is cited by
-
Preliminary evidence of Drosophila suzukii parasitism in Southeast England
Parasitology Research (2023)
-
Search performance and octopamine neuronal signaling mediate parasitoid induced changes in Drosophila oviposition behavior
Nature Communications (2022)