
Abstract
Iron status is known to be associated with the physiology of adipose tissue (AT). We aimed to investigate AT heme and expression of heme exporter (FLVCR1) in association with obesity and type 2 diabetes (T2D). Substantial amounts of FLVCR1 mRNA and protein levels were detected in AT, being significantly increased in subjects with T2D, and positively correlated with fasting glucose, fasting triglycerides and with circulating markers of iron stores (serum ferritin, blood hemoglobin and hematocrit). In both visceral (VAT) and subcutaneous AT (SAT), increased heme levels were found in subjects with T2D. Reinforcing these associations, FLVCR1 mRNA levels were positively linked to fasting glucose in an independent cohort. Longitudianlly, the percent change of FLVCR1 positively correlated with the percent change in fasting glucose (r = 0.52, p = 0.03) after bariatric surgery-induced weight loss. High-fat diet-induced weight gain in rats did not result in significant changes in AT Flvcr1 mRNA but, remarkably, the expression of this gene positively correlated with fasting glucose and negatively with insulin sensitivity (QUICKI). Altogether, these findings showed a direct association between FLVCR1 mRNA levels and hyperglycemia, suggesting that increased adipose tissue heme exportation might disrupt, or is the consequence of, impaired systemic glucose metabolism during the progression to T2D.
Similar content being viewed by others

Introduction
Heme constitutes a relevant form of functional iron in the human body, as well as two-thirds of the average person’s iron intake in developed countries1. Epidemiological studies have substantiated increased risks of type-2 diabetes associated with high heme intake2,3,4,5,6. Even though non-heme iron is the predominant form of dietary iron, heme iron is more easily absorbed2. In fact, the intake of heme iron, but not of non-heme iron, has been positively associated with body iron stores2, 6, and increased body iron stores have been extensively demonstrated in type 2 diabetes7, 8. Mechanistically, excess of intracellular heme levels are degraded by heme oxygenase resulting in increased free iron9. In fact, increased heme oxygenase activity was found in patients with type 2 diabetes in association with body iron stores9. Furthermore, intracellular free heme excess catalyzes the formation of reactive oxygen species (ROS), promoting oxidative stress10, 11. Increased tissue iron deposition and iron-induced oxidative stress might result in increased beta-cells apoptosis, hepatic dysfunction and insulin resistance, and in consequence, to promote the progression of type 2 diabetes6.
Feline Leukemia Virus subgroup C Receptor 1 (FLVCR1) is a plasma membrane heme exporter that ensures the maintenance of appropriate intracellular heme concentration12,13,14. The depletion of FLVCR1 led to excess heme content in erythroid progenitors, and in consequence, increased cytoplasmic ROS and apoptosis, whereas restoring FLVCR1 gene expression returned to normal erythropoiesis, demonstrating that excess of intracellular heme levels disrupted this cellular process12,13,14. Recent studies demonstrated the importance of FLVCR1 in the maintenance of heme homeostasis in other tissues, including intestine15 and liver16. Interestingly, the disruption of FLVCR1 in these tissues resulted in increased tissue heme accumulation in parallel to increased heme oxygenase 1 (HMOX1) and markers of oxidative stress and iron accumulations (ferroportin and ferritin levels)15, 16.
Increased iron17,18,19, markers of iron accumulation17, 20,21,22 and HMOX1 mRNA and protein levels23,24,25 have been demonstrated in mice and human adipose tissue in association with adipose tissue dysfunction (increased inflammation and decreased adipogenesis). In vitro experiments in mouse cells revealed increasing heme biosynthesis during adipocyte differentiation26. Increased intracellular heme accumulation resulted in oxidative stress and adipocyte hypertrophy27. In fact, heme-induced HMOX1 activity decreased adipocyte differentiation of human preadipocytes, and attenuated glucose uptake, mitochondrial function in parallel to increased inflammation and markers of oxidative stress and iron accumulation in human adipocytes25.
To the best of our knowledge, FLVCR1 and heme levels have not been previously explored in human adipose tissue. Considering these intriguing studies17,18,19,20,21,22,23,24,25,26,27 and the importance of FLVCR1 in the maintenance of tissue heme homeostasis12,13,14,15,16, we aimed to investigate cross-sectionally (in two cohorts) the expression of FLVCR1 according to obesity and type 2 diabetes (T2D), and longitudinally the effects of weight gain (in rats) and weight loss (bariatric surgery of morbidly obese subjects) on AT FLVCR1 mRNA levels. Adipose tissue heme levels were also examined.
Results
Heme levels and heme exporter-related gene expression in human AT
Anthropometrical and clinical parameters of the study subjects are shown in Table 1. Substantial amounts of heme exporter (FLVCR1) mRNA and protein levels were detected in both VAT and SAT (Fig. 1a–c). FLVCR1 mRNA was positively correlated with FLVCR1 protein levels (r = 0.56, p = 0.05; Fig. 1d). Adipose tissue fraction analysis indicated increased FLVCR1 mRNA levels in stroma vascular fraction (SVF) in comparison to adipocytes (Fig. 1e,f) in both VAT and SAT.

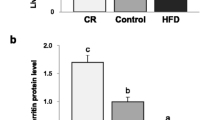
FLVCR1 gene expression in VAT (a) and SAT (b) according to fasting glucose status and type 2 diabetes in cohort 1 [NFG (n = 80), IFG (n = 29) and T2D (n = 19)]. *p < 0.05 and **p < 0.01 compared with NFG participants. †p < 0.05 and ††p < 0.01 compared with IFG participants. RU, relative units of gene expression. Gene expression was calculated by 2−∆∆Ct method. (c) Representative immunoblot of FLVCR1 protein levels in VAT (n = 7) and SAT (n = 5) from NFG participants. (d) Bivariate correlation between FLVCR1 gene expression and FLVCR1 protein levels (normalized by β-actin) (n = 12). (e,f) FLVCR1 gene expression in adipose tissue fractions (SVF vs adipocytes) in VAT (e) and SAT (f) (n = 5). **p < 0.01 compared with SVF.
In both VAT (Fig. 1a) and SAT (Fig. 1b), FLVCR1 gene expression was significantly increased in patients with T2D when compared with AT form patients with impaired fasting glucose or with normal fasting glucose levels, and expression of this gene was positively correlated with fasting glucose (Table 2). Expression of the FLVCR1 gene was associated with other T2D-associated metabolic traits, including positive correlations with fasting triglycerides in both SAT and VAT, and circulating markers of iron stores (serum ferritin, blood hemoglobin and hematocrit) only in VAT (Table 2). Negative correlations with HDL-cholesterol in SAT and expression of adipogenesis (ADIPOQ) and insulin signaling-related genes (SLC2A4, IRS1) (Table 2) in VAT were also observed.
To investigate a possible association between adipose tissue FLVCR1 and heme levels, adipose tissue heme levels were measured in a subgroup of 41 [32 NFG and 9 T2D (Table 3)] participants from cohort 1. In both SAT and VAT, adipose tissue heme levels were detected at substantial amounts and were significantly increased in patients with T2D (Fig. 2a,b). Heme levels positively correlated with fasting glucose (r = 0.51, p = 0.001 in VAT; r = 0.34, p = 0.03 in SAT, Fig. 2c,d). VAT and SAT heme levels were positively correlated with FLVCR1 mRNA levels (r = 0.34, p = 0.04 in VAT; r = 0.37, p = 0.03 in SAT, Table 2).

Hemin levels in VAT (a) and SAT (b) according to type 2 diabetes in cohort 1 [NFG (n = 32) and T2D (n = 9)]. *p < 0.05 and **p < 0.01 compared with NFG participants. (c,d) Bivariate correlation between adipose tissue hemin levels and fasting glucose (n = 41) in both VAT (c) and SAT (d).
Since heme oxygenase 1 (HMOX1) gene expression is known to be strongly induced by heme levels [23–25], HMOX1 mRNA levels were also analyzed in this subgroup from cohort 1 (Table 3). Adipose tissue heme levels positively correlated with HMOX1 gene expression (r = 0.32, p = 0.04 in VAT; r = 0.36, p = 0.02 in SAT, Suppl Fig. 1A,B).
Replication in an independent cohort
Anthropometrical and clinical parameters of these subjects are shown in Table 1. Reinforcing the above described associations, FLVCR1 gene expression was positively correlated with fasting glucose (r = 0.31, p = 0.02 in VAT; r = 0.27, p = 0.04 in SAT) (Table 2).
Effects of bariatric surgery-induced weight loss
Bariatric surgery-induced weight loss did not result in significant changes in FLVCR1 gene expression (Suppl Table 1). However, the percent change of FLVCR1 gene expression was positively correlated with the percent change of fasting glucose (r = 0.52, p = 0.03, Fig. 3a). Fasting glucose decreased (95.2 ± 7.1 vs 86.3 ± 5.2, p = 0.002) in 11 participants and increased (88.8 ± 10.1 vs 95.4 ± 12.1, p = 0.01) in 5 subjects two years after surgery.

(a) Bivariate correlation between the change of FLVCR1 gene expression and the change of fasting glucose 2 years after bariatric surgery (Cohort 3, n = 16)). (b) The effect of high-fat diet (HFD) on eWAT Flvcr1 gene expression in rat experiments [normal diet (n = 10) and high fat diet (n = 10)]. (c,d) Bivariate correlation between eWAT Flvcr1 gene expression and fasting glucose (c) or QUICKI (d) (n = 20).
Effects of weight gain in rats
In rats, comparing those fed with HFD vs normal diet (Suppl Fig. 1C–E), no significant differences on Flvcr1 gene expression in epididymal white adipose tissue (eWAT) were observed (Fig. 3b). Nonetheless, expression of Flvcr1 gene was positively correlated with fasting glucose (r = 0.47, p = 0.04; Fig. 3c) and negatively with QUICKI (r = −0.45, p = 0.04; Fig. 3d). Flvcr1 was not significantly correlated with Il6 (r = 0.29, p = 0.2) and Slc2a4 (r = −0.21, p = 0.3) mRNA levels in eWAT.
Discussion
This study demonstrated substantial amounts of heme and FLVCR1 mRNA and protein levels in both VAT and SAT. The main finding of the present study was that adipose tissue heme and FLVCR1 mRNA levels were increased in AT from patients with type 2 diabetes in direct proportion to fasting glucose levels, but not in association with BMI. Taking into account the well-demonstrated relationship between high diet heme iron uptake and increased risk of type 2 diabetes2,3,4,5,6, this study suggests a relationship between adipose tissue export of heme iron and type 2 diabetes. In line with these results, the change of FLVCR1 mRNA levels ran in parallel with the change of fasting glucose independently of weight loss. Fasting glucose decreased in some participants and increased in others 2 years after surgery. These discrepancies could be explained by the fact that no patient with type 2 diabetes was included in the current study. It is known that the effects of bariatric surgery on fasting glucose reduction are more pronounced in patients with type 2 diabetes28. After weight gain, Flvcr1 mRNA levels were positively associated with fasting glucose and negatively with insulin sensitivity (QUICKI) in rats, but not with fatness. In cohort 1, both SAT and VAT FLVCR1 gene expression was also associated with fasting triglycerides, and with serum ferritin and decreased mRNA levels of adipogenic markers in VAT. In line with this, increased susceptibility to iron-induced insulin resistance and dysfunctional adipogenesis has been demonstrated in VAT20, 29. Increased intracellular heme accumulation is known to result in dysfunctional and hypertrophic adipocytes27, suggesting that adipocyte heme excess might contribute to the link between adipocyte hypertrophy and type 2 diabetes30,31,32,33. In addition FLVCR1 mRNA was negatively correlated with expression of insulin signaling-related genes (SLC2A4 and IRS1). Decreased SLC2A4 and IRS1 gene expression in adipose tissue are associated with insulin resistance34.
In adipose tissue fraction, FLVCR1 mRNA levels were increased in SVF vs adipocytes. Considering that FLVCR1 is required for T cell development and survival35, highly expressed in macrophages13, 36, and decreased under inflammatory conditions36, the relationship between adipose tissue FLVCR1 and markers of macrophages [ITGAX 37], regulatory T lymphocytes [FOXP3 38] or inflammation (TNF, IL6) was explored. Of note, adipose tissue FLVCR1 were not associated with ITGAX, FOXP3, TNF or IL6 mRNA levels, suggesting that adipose tissue FLVCR1 mRNA was not attributed to inflammatory cells.
Adipose tissue heme levels were confirmed using HMOX1 mRNA levels, another surrogate marker of intracellular heme levels39. By different ways, HMOX1 (heme degradation)23,24,25 and FLVCR1 (heme exportation)13,14,15,16 might impact on intracellular heme levels. Increased HMOX1 gene and protein expression has been recently demonstrate in obese participants, but not in association with type 2 diabetes24, 25, 40, 41, whereas FLVCR1 was increased specifically in patients with type 2 diabetes (current findings).
Heme biosynthesis is required for adipocyte differentiation26. Increased fat accretion results in raised heme levels in adipose tissue, enhanced heme-induced HMOX1 expression and activity, free iron accumulation25, proinflammatory activity and, in consequence, insulin resistance and hyperglycemia. In bone marrow, depletion of FLVCR1 (i.e. lack of heme exportation), with the attendant heme accumulation, leads to cell damage and apoptosis42. Efficient heme exportation is also crucial in maintaining intestinal homeostasis15. The situation regarding adipose tissue, reported for the first time in the present manuscript, seems to be different from these two cellular models. The current study has shown how impaired glucose metabolism was associated with increased heme levels in adipose tissue in parallel to raised expression of FLVCR1 and HMOX1. These data suggest that heme exportation and degradation in adipose tissue is possibly induced in diabetogenic conditions to counteract intracellular heme levels, preventing their negative effects on free iron-induced oxidative stress and cellular damage10, 11. However, further functional experiments are necessary to clarify the relevance of FLVCR1 in adipocyte physiology
In conclusion, intracellular heme excess in human adipocytes might result in enhanced export of heme iron, and in alterations in systemic glucose metabolism. Possibly, iron accumulation in other tissues [liver19, muscle43 and brain44] would also worsen hyperglycemia and type 2 diabetes. The current study is focused on adipose tissue, but considering the importance of other insulin-dependent tissues (including liver and skeletal muscle) on glucose metabolism, additional studies exploring the relationship between heme exportation and glucose metabolism in liver and skeletal muscle are required. Further molecular and cellular studies should be designed to decipher the mechanism underlying the relationship between heme exportation and glucose metabolism.
Methods
Human adipose tissue samples
Adipose tissue samples were obtained from two independent cohorts. In cohort 1 (N = 251), visceral (VAT, N = 123) and subcutaneous (SAT, N = 128) adipose tissue samples from participants with normal body weight and different degrees of obesity, body mass index (BMI) ranging from 20 to 68 kg/m2, recruited at the Endocrinology Service of the Hospital Universitari Dr. Josep Trueta (Girona, Spain) were analyzed. In a second cohort of non-diabetic morbidly obese (cohort 2, BMI > 35 kg/m2) subjects, 50 paired SAT and VAT samples were studied. All subjects were of Caucasian origin and reported that their body weight had been stable for at least three months before the study. Liver and renal diseases were specifically excluded by biochemical work-up. All subjects gave written informed consent, validated and approved by the ethical committee of the Hospital Universitari Dr. Josep Trueta after the purpose of the study was explained to them. All methods were performed in accordance with the relevant guidelines and regulations. Adipose tissue samples were obtained from SAT and VAT depots during elective surgical procedures (cholecystectomy, surgery of abdominal hernia and gastric bypass surgery). Adipose tissue samples were washed, fragmented and immediately flash-frozen in liquid nitrogen before being stored at −80 °C.
The isolation of adipocyte and stromal vascular fraction cells (SVF) was performed from 6 SAT (n = 5) and VAT (n = 5) non-frozen adipose tissue samples as described previously22. These samples were washed three to four times with phosphate-buffered saline (PBS) and suspended in an equal volume of PBS supplemented with 1% penicillin-streptomycin and 0.1% collagenase type I warmed to 37 °C. The tissue was placed in a shaking water bath at 37 °C with continuous agitation for 60 minutes and centrifuged for 5 minutes at 300 to 500 g at room temperature. The supernatant, containing mature adipocytes, was recollected. The pellet was identified as the SVF. Isolated mature adipocytes and SVF were stored at −80 °C for gene expression analysis.
Study of the effects of fat mass reduction induced by bariatric surgery
Sixteen subjects who underwent bariatric surgery via Roux-en-Y gastric bypass (RYGB) at the Hospital Universitari Dr. Josep Trueta (Girona, Spain) were part of an ongoing study21, 25. Inclusion criteria were age between 30 and 60 years, BMI ≥ 35 kg/m2 and ability to understand the study protocol. Exclusion criteria were: (i) baseline T2D; (ii) use of medications able to interfere with insulin action; (iii) acute infection process 4 weeks prior to sample collection; (iv) history of a chronic systemic disease or other serious chronic associated illness. Adipose tissue samples from the SAT depot were obtained during bariatric surgery. Postoperative samples of SAT were obtained by subcutaneous biopsy at the mesogastric level 2 years after surgery. Fasting blood samples were obtained at the same day of the biopsy. All subjects gave written informed consent, validated and approved by the ethical committee of the Hospital Universitari Dr. Josep Trueta, after the purpose of the study was explained to them. All methods were performed in accordance with the relevant guidelines and regulations.
Heme levels measurement
In a subgroup of 41 [32 with normal fasting glucose (NFG) and 9 with T2D)] participants from cohort 1, adipose tissue heme levels were measured using the hemin assay kit (ab65332, Abcam plc, Cambridge, UK) according to the manufacturer’s instructions, with a coefficient of variation <10%.
Analytical methods
Serum glucose concentrations were measured in duplicate by the glucose oxidase method using a Beckman glucose analyser II (Beckman Instruments, Brea, California). Intraassay and interassay coefficients of variation were less than 4% for all these tests. HDL-cholesterol was quantified by an homogeneous enzymatic colorimetric assay through the cholesterol esterase/cholesterol oxidase/peroxidase reaction (Cobas HDLC3). Total serum triglycerides were measured by an enzymatic, colorimetric method with glycerol phosphate oxidase and peroxidase (Cobas TRIGL). We used a Roche Hitachi Cobas c 711 instrument to perform the determinations. Serum ferritin was determined by Microparticle Enzyme ImmunoAssay (AXSYMTM; Abbot Laboratories, Abbott Park, IL), with a coefficient of variation intra- and interassay <6%.
Effects of weight gain in rats
Four-week-old male Wistar rats (n = 20) (breeding house of the University of Navarra) were housed in a room with controlled temperature (22 ± 2 °C), relative humidity (50 ± 10%), ventilation (at least 15 complete changes of air/h), and 12:12 light-dark cycle (lights on at 8:00 am). Rats were housed in individual cages and were fed ad libitum during an average of 6 months either a normal chow diet [ND (12.1 kJ/g: 4% fat, 48% carbohydrate and 14% protein; diet 2014, Harlan, Teklad Global Diets, Harlan Laboratories Inc., Barcelona, Spain); n = 10] for comparative purposes or a high-fat diet [HFD (23.0 kJ/g: 60% fat, 27% carbohydrate and 14% protein; diet F3282, Bio-Serv, Frenchtown, NJ, USA); n = 10] to induce obesity. Body weight and food intake were recorded on a regular basis to monitor progression of diet-induced obesity. After an overnight fast, rats were sacrificed by decapitation and the epididymal white adipose tissue (eWAT) depots were carefully dissected out, weighed, frozen in liquid nitrogen, and stored at –80 °C. All experimental procedures conformed to the European Guidelines for the Care and Use of Laboratory Animals (directive 2010/63/EU) and were approved by the Ethical Committee for Animal Experimentation of the University of Navarra (049/10). All methods were performed in accordance with the relevant guidelines and regulations.
RNA expression
RNA purification was performed using RNeasy Lipid Tissue Mini Kit (QIAgen, Izasa SA, Barcelona, Spain) and the integrity was checked by Agilent Bioanalyzer (Agilent Technologies, Palo Alto, CA). Gene expression was assessed by real time PCR using an LightCycler® 480 Real-Time PCR System (Roche Diagnostics SL, Barcelona, Spain), using TaqMan® technology suitable for relative genetic expression quantification. The RT-PCR reaction was performed in a final volume of 12 μL. The cycle program consisted of an initial denaturing of 10 min at 95 °C then 40 cycles of 15 sec denaturizing phase at 95 °C and 1 min annealing and extension phase at 60 °C. A threshold cycle (Ct value) was obtained for each amplification curve and a ∆Ct value was first calculated by subtracting the Ct value for human cyclophilin A (PPIA) RNA from the Ct value for each sample. Fold changes compared with the endogenous control were then determined by calculating 2−∆Ct, so gene expression results are expressed as expression ratio relative to PPIA gene expression according to manufacturers’ guidelines. Primer/probe sets used were: feline leukemia virus subgroup C cellular receptor 1 (FLVCR1, Hs01067777_m1); tumor necrosis factor (TNF, Hs00174128_m1); adiponectin (ADIPOQ, Hs00605917_m1); solute carrier family 2 (facilitated glucose transporter), member 4 (SLC2A4 or GLUT4, Hs00168966_m1); Insulin receptor substrate 1 (IRS1, Hs00178563_m1); integrin subunit alpha X (ITGAX or CD11C, Hs00174217_m1); forkhead box P3 (FOXP3, Hs01085834_m1); Interleukin 6 (IL6, Hs00985639_m1); heme oxygenase 1 (HMOX1, Hs01110250_m1); and Peptidylprolyl isomerase A (cyclophilin A) (4333763, PPIA as endogenous control). In rats, primer/probe sets used were feline leukemia virus subgroup C cellular receptor 1, ( Flvcr1, Rn01411620_m1), interleukin 6 (Il6, Rn01410330_m1), solute carrier family 2 (facilitated glucose transporter), member 4 (Slc2a4 or Glut4, Rn00562597_m1) and Eukaryotic 18S rRNA (4331182) as endogenous control.
Protein preparation and western blot
To explore whether FLVCR1 protein is detected in human adipose tissue in proportion to FLVCR1 mRNA levels, FLVCR1 protein was analysed in 12 samples according to the amount of available adipose tissue. Adipose tissue proteins were extracted directly in radio immnunoprecipitation assay (RIPA) buffer (0.1% SDS, 0.5% sodium deoxycholate, 1% Nonidet P-40, 150 mmol/L NaCl, and 50 mmol/L Tris-HCl, pH 8.00), supplemented with protease inhibitors (1 mmol/L phenylmethylsulfonyl fluoride). Cellular debris and lipids were eliminated by centrifugation of the solubilized samples at 14000 g for 10 min at 4 °C, recovering the soluble fraction. Protein concentration was determined using the RC/DC Protein Assay (Bio-Rad Laboratories, Hercules, CA). RIPA protein extracts (25 μg) were separated by SDS-PAGE and transferred to nitrocellulose membranes by conventional procedures. Membranes were immunoblotted with anti-FLVCR antibody (ab70838, Abcam plc, Cambridge, UK) and β-actin antibodies (sc-47778, Santa Cruz Biotechnology, CA, USA). Anti-rabbit IgG and anti-mouse IgG coupled to horseradish peroxidase was used as a secondary antibody. Horseradish peroxidase activity was detected by chemiluminescence, and quantification of protein expression was performed using Scion Image software.
Statistical analyses
Statistical analyses were performed using the SPSS 12.0 software. Unless otherwise stated, descriptive results of continuous variables are expressed as mean and SD for Gaussian variables or median and interquartile range for non-Gaussian variables. Parameters that did not fulfill normal distribution criteria were log transformed to improve symmetry for subsequent analyses. The relation between variables was analyzed by simple correlation (using Spearman’s and Pearson’s tests). ANOVA and unpaired Student’s t-tests were used to compare clinical variables, heme levels and FLVCR1 gene expression relative to obesity and T2D.
Availability of materials and data
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Hooda, J., Shah, A. & Zhang, L. Heme, an essential nutrient from dietary proteins, critically impacts diverse physiological and pathological processes. Nutrients 6, 1080–1102 (2014).
Bao, W., Rong, Y., Rong, S. & Liu, L. Dietary iron intake, body iron stores, and the risk of type 2 diabetes: a systematic review and meta-analysis. BMC Med. 10, 119 (2012).
Fernandez-Cao, J. C. et al. Heme iron intake and risk of new-onset diabetes in a Mediterranean population at high risk of cardiovascular disease: an observational cohort analysis. BMC Public Health. 13, 1042 (2013).
Pan, A. et al. Changes in red meat consumption and subsequent risk of type 2 diabetes mellitus: three cohorts of US men and women. JAMA Intern Med. 173, 1328–1335 (2013).
de Oliveira Otto, M. C. et al. Dietary intakes of zinc and heme iron from red meat, but not from other sources, are associated with greater risk of metabolic syndrome and cardiovascular disease. J Nutr. 142, 526–533 (2012).
Zhao, Z. et al. Body iron stores and heme-iron intake in relation to risk of type 2 diabetes: a systematic review and meta-analysis. PLoS One 7, e41641 (2012).
Fernández-Real, J. M., McClain, D. & Manco, M. Mechanisms Linking Glucose Homeostasis and Iron Metabolism Toward the Onset and Progression of Type 2 Diabetes. Diabetes Care. 38, 2169–2176 (2015).
Montonen, J. et al. Body iron stores and risk of type 2 diabetes: results from the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Diabetologia. 55, 2613–2621 (2012).
Arredondo, M., Jorquera, D., Carrasco, E., Albala, C. & Hertrampf, E. Microsatellite polymorphism in the heme oxygenase-1 gene promoter is associated with iron status in persons with type 2 diabetes mellitus. Am J Clin Nutr. 86, 1347–1353 (2007).
Chiabrando, D., Vinchi, F., Fiorito, V., Mercurio, S. & Tolosano, E. Heme in pathophysiology: a matter of scavenging, metabolism and trafficking across cell membranes. Front Pharmacol. 5, 61 (2014).
Balla, G., Vercellotti, G. M., Muller-Eberhard, U., Eaton, J. & Jacob, H. S. Exposure of endothelial cells to free heme potentiates damage mediated by granulocytes and toxic oxygen species. Lab Invest. 64, 648–655 (1991).
Quigley, J. G. et al. Identification of a human heme exporter that is essential for erythropoiesis. Cell. 118, 757–766 (2004).
Keel, S. B. et al. A heme export protein is required for red blood cell differentiation and iron homeostasis. Science. 319, 825–828 (2008).
Doty, R. T. et al. Coordinate expression of heme and globin is essential for effective erythropoiesis. J Clin Invest. 125, 4681–4691 (2015).
Fiorito, V., Forni, M., Silengo, L., Altruda, F. & Tolosano, E. Crucial Role of FLVCR1a in the Maintenance of Intestinal Heme Homeostasis. Antioxid Redox Signal. 23, 1410–1423 (2015).
Vinchi, F. et al. Heme exporter FLVCR1a regulates heme synthesis and degradation and controls activity of cytochromes P450. Gastroenterology. 146, 1325–1338 (2014).
Gabrielsen, J. S. et al. Adipocyte iron regulates adiponectin and insulin sensitivity. J Clin Invest. 122, 3529–3540 (2012).
Orr, J. S. et al. Obesity alters adipose tissue macrophage iron content and tissue iron distribution. Diabetes. 63, 421–432 (2014).
Fernández-Real, J. M. et al. Adipose tissue R2* signal is increased in subjects with obesity: A preliminary MRI study. Obesity (Silver Spring). 24, 352–358 (2016).
Dongiovanni, P. et al. Dietary iron overload induces visceral adipose tissue insulin resistance. Am J Pathol. 182, 2254–2263 (2013).
Moreno-Navarrete, J. M. et al. Insulin resistance modulates iron-related proteins in adipose tissue. Diabetes Care. 37, 1092–1100 (2014).
Moreno-Navarrete, J. M., Ortega, F., Moreno, M., Ricart, W. & Fernández-Real, J. M. Fine-tuned iron availability is essential to achieve optimal adipocyte differentiation and mitochondrial biogenesis. Diabetologia. 57, 1957–1967 (2014).
Wagner, G. et al. HO-1 inhibits preadipocyte proliferation and differentiation at the onset of obesity via ROS dependent activation of Akt2. Sci Rep. 7, 40881 (2017).
Jais, A. et al. Heme oxygenase-1 drives metaflammation and insulin resistance in mouse and man. Cell. 158, 25–40 (2014).
Moreno-Navarrete, J. M. et al. HMOX1 as a marker of iron excess-induced adipose tissue dysfunction, affecting glucose uptake and respiratory capacity in human adipocytes. Diabetologia. (in press) (2017).
Kumar, N. et al. Regulation of adipogenesis by natural and synthetic REV-ERB ligands. Endocrinology. 151, 3015–3025 (2010).
Puri, N. et al. Heme induced oxidative stress attenuates sirtuin1 and enhances adipogenesis in mesenchymal stem cells and mouse pre-adipocytes. J Cell Biochem. 113, 1926–1935 (2012).
Camastra, S. et al. Long-term effects of bariatric surgery on meal disposal and β-cell function in diabetic and nondiabetic patients. Diabetes. 62, 3709–3717 (2013).
Moreno-Navarrete, J. M. et al. CISD1 in association with obesity-associated dysfunctional adipogenesis in human visceral adipose tissue. Obesity (Silver Spring) 24, 139–147 (2016).
Acosta, J. R. et al. Increased fat cell size: a major phenotype of subcutaneous white adipose tissue in non-obese individuals with type 2 diabetes. Diabetologia. 59, 560–570 (2016).
Rojas-Rodriguez, R. et al. Human adipose tissue expansion in pregnancy is impaired in gestational diabetes mellitus. Diabetologia 58, 2106–2114 (2015).
Henninger, A. M., Eliasson, B., Jenndahl, L. E. & Hammarstedt, A. Adipocyte hypertrophy, inflammation and fibrosis characterize subcutaneous adipose tissue of healthy, non-obese subjects predisposed to type 2 diabetes. PLoS One 9, e105262 (2014).
Rydén, M., Andersson, D. P., Bergström, I. B. & Arner, P. Adipose tissue and metabolic alterations: regional differences in fat cell size and number matter, but differently: a cross-sectional study. J Clin Endocrinol Metab 99, E1870–E1876 (2014).
Hammarstedt, A., Jansson, P. A., Wesslau, C., Yang, X. & Smith, U. Reduced expression of PGC-1 and insulin-signaling molecules in adipose tissue is associated with insulin resistance. Biochem Biophys Res Commun. 301, 578–582 (2003).
Philip, M. et al. Heme exporter FLVCR is required for T cell development and peripheral survival. J Immunol. 194, 1677–1685 (2015).
Philip, M., Chiu, E. Y., Hajjar, A. M. & Abkowitz, J. L. TLR Stimulation Dynamically Regulates Heme and Iron Export Gene Expression in Macrophages. J Immunol Res. 2016, 4039038 (2016).
Song, M. G. et al. Depot-specific differences in angiogenic capacity of adipose tissue in differential susceptibility to diet-induced obesity. Mol Metab. 5, 1113–1120 (2016).
Vasanthakumar, A. et al. The transcriptional regulators IRF4, BATF and IL-33 orchestrate development and maintenance of adipose tissue-resident regulatory T cells. Nat Immunol. 16, 276–285 (2015).
Yang, M. et al. Induction of heme-oxygenase-1 (HO-1) does not enhance adiponectin production in human adipocytes: Evidence against a direct HO-1 - Adiponectin axis. Mol Cell Endocrinol. 413, 209–216 (2015).
Shakeri-Manesch, S. et al. Diminished upregulation of visceral adipose heme oxygenase-1 correlates with waist-to-hip ratio and insulin resistance. Int J Obes (Lond) 33, 1257–1264 (2009).
Mannerås-Holm, L., Benrick, A. & Stener-Victorin, E. Gene expression in subcutaneous adipose tissue differs in women with polycystic ovary syndrome and controls matched pair-wise for age, body weight, and body mass index. Adipocyte. 3, 190–196 (2014).
Byon, J. C., Chen, J., Doty, R. T. & Abkowitz, J. L. FLVCR is necessary for erythroid maturation, may contribute to platelet maturation, but is dispensable for normal hematopoietic stem cell function. Blood. 122, 2903–2910 (2013).
Moreno-Navarrete, J. M. et al. Obesity Is Associated With Gene Expression and Imaging Markers of Iron Accumulation in Skeletal Muscle. J Clin Endocrinol Metab. 101, 1282–1289 (2016).
Blasco, G. et al. Brain iron overload, insulin resistance, and cognitive performance in obese subjects: a preliminary MRI case-control study. Diabetes Care. 37, 3076–3083 (2014).
Acknowledgements
We acknowledge the technical assistance of E. Loshuertos and O. Rovira (both from Endocrinology, IdIBGi, Spain). We are indebted to the IDIBGI Biobank, integrated in the Spanish National Biobank Network, for the sample and data procurement. This work was partially supported by research grants FIS PI12/02631 and PI15/01934 from the Instituto de Salud Carlos III from Spain and was also supported by FEDER funds. CIBEROBN Fisiopatología de la Obesidad y Nutrición is an initiative from the Instituto de Salud Carlos III from Spain.
Author information
Authors and Affiliations
Contributions
J.M.M.-N. and J.M.F.-R. participated in study design and analysis of data. F.O., A.R., J.L., S.B. and M.S.-M. participated in acquisition of data. W.R. and G.F. participated in interpretation of data. J.M.M.-N. and J.M.F.-R. wrote and edited the manuscript. F.O., A.R., J.L., S.B., M.S.-M., W.R. and G.F. revised the manuscript critically for important intellectual content. All authors participated in final approval of the version to be published.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Moreno-Navarrete, J.M., Rodríguez, A., Ortega, F. et al. Increased adipose tissue heme levels and exportation are associated with altered systemic glucose metabolism. Sci Rep 7, 5305 (2017). https://doi.org/10.1038/s41598-017-05597-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-05597-2
- Springer Nature Limited
This article is cited by
-
Unearthing FLVCR1a: tracing the path to a vital cellular transporter
Cellular and Molecular Life Sciences (2024)