
Abstract
The genus Schima includes about 20 species and is distributed only in southern China and adjacent areas of Asia. The previous molecular phylogenetic analysis suggested Schima is in the tribe Gordoniae, along with Gordonia and Franklinia. However, because few fossils have been reported, the biogeographic origin of Schima is still poorly known. In this paper mummified fossil fruits of Schima are described from the upper Oligocene Yongning Formation of the Nanning Basin, Guangxi, South China. In gross morphology, the new fossil species, Schima kwangsiensis, is similar to the extant S. superba by its pentacarpellate, loculicidally dehiscent capsules, 5 imbricate sepals, pedicels with bracteoles and marginally winged seeds. Due to its excellent preservation, the new species may provide sufficient details for understanding the early evolutionary and phytogeographic history of the genus. Morphological clustering analysis shows that the new fossil species is closely related to two extant species (S. wallichii and S. superba) in the genus, implying that they may belong to an ancient taxon that occurs earlier than the others. More importantly, this discovery represents the earliest record of this genus in Asia and it explicitly moves the fossil record back to the late Oligocene in this region.
Similar content being viewed by others


Introduction
Schima, a genus within the Theaceae in Asia, inhabits subtropical to tropical regions across southern and southeastern Asia, from the eastern Himalaya of Nepal and eastern India across Indochina, southern China, Taiwan, and the Ryukyu Islands1. There are about 20 species, including six species endemic to China2, 3. It is mainly characterized by marginally winged seeds and capsules with persistent columella. Traditionally, the genus belongs to the subfamily Theoideae and is usually placed in the tribe Gordoniae with Gordonia, Franklinia and Laplacea 4,5,6, or in the tribe Schimeae with Franklinia and Apterasperma 7,8,9. Recent molecular phylogenetic analysis showed Schima is closely related to Gordonia and Franklinia, forming the monophyletic tribe of Gordoniae 10,11,12,13 (Fig. 1).

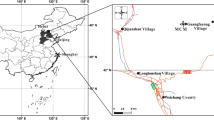
Fruit characters are considered to be of crucial systematic importance in the modern Theaceae8, but fossil fruits of Schima are very rare14, 15. Here, we describe numerous mummified Schima fruits collected from a plant fossil Konservat Lagerstätte reported recently16 in the late Oligocene Yongning Formation of Nanning Basin of Guangxi Province, South China (22°52′50′′ N, 108°25′2′′E, Fig. 2).

Geographic map of the Nanning Basin, Guangxi Province, generated by X. G. Shi, using Arcgis 9.3 (ESRI, Redlands, CA, USA).
The Nanning Basin is located in Nanning City, Guangxi Province and it mainly consists of three formations: the Ducun Formation, the Yongjiang Formation and the Yongning Formation17. According to their respective lithic facies, the Yongning Formation is subdivided into the upper, middle and lower parts16. Of these, the upper part is rich in mummified fossils, and is mainly composed of bluish gray clayey mudstone, interlined with a few coal seams and thin sandstones. Based on uncovered mammal fossils of Anthracotherium chanlingensis Zhao, Anthracokeryx kwangsiensis Qiu, and Heothema sp. from Changlin, Xiaoji, and Guzang in the same basin, the age of the upper part is determined as the late Oligocene16, 17. Therefore, this occurrence is the earliest definite record of the genus Schima in Asia and it moves explicitly the fossil record back to the late Oligocene in Asia. In addition, the new finding with exceptional preservation is the first fossil fruit with a pedicel in the genus. The purpose of this paper is to evaluate the new fossil record of Schima and further discuss systematics and phytogeographical history of the genus based on integrated evidence from megafossils and morphological clustering analysis.
Results
Systematics
Class Magnoliopsida Brongn.
Order Ericales Bercht. & J. Presl.
Family Theaceae Mirbel.
Genus Schima Reinwardt ex Blume.
Species Schima kwangsiensis X. G. Shi, C. Quan et J. H. Jin sp. nov.
Holotype
NNF-070 (Fig. 3A,C) (designated here).

Fruits with pedicels of Schima kwangsiensis sp. nov. and extant Schima superba. (A) The fossil fruit showing the globose capsule, and the apically thickened and recurved pedicel (holotype, specimen number NNF-070). (B) A modern fruit of the species Schima supberba, collected from the Heishiding Nature Reserve, Guangdong Province. (C) The top surface of A showing six loculicidally dehiscent capsule. (D) Five loculicidally dehiscent capsules, paratype, specimen number NNF-074. (E) A capsule and its long pedicel with bracteole scar (green arrowhead), paratype, specimen number NNF-075. (F) Dehiscent capsule showing semiorbicular sepals (green arrowhead), paratype, specimen number NNF-073. Scale bar = 2 mm. Processed and drawn by X. G. Shi using Adobe Photoshop CS5.
Paratypes
NNF-072 (Fig. 4A); NNF-073 (Fig. 3F); NNF-074 (Fig. 3D); NNF-075 (Fig. 3E); NNF-076 (Fig. 4G); NNF-1490 (Fig. 4D); NNF-1491 (Fig. 4E); NNF-1493 (Fig. 4B); NNF-1495 (Fig. 4C); NNF-1499 (Fig. 4F) (designated here).

Fruit structure of Schima kwangsiensis sp. nov. and extant Schima superba. (A–G) Fossil fruits: (A) Dehiscent capsule showing semiorbicular sepals (green arrowhead), specimen number NNF-072. (B) Dehiscent capsule showing splits running down the upper 2/3 of the capsule length, specimen number NNF-1493. (C) Dehiscent capsule with five ventral sutures, specimen number NNF-1495. (D) Dehiscent capsule showing 5 valves and dissepiments, specimen number NNF-1490. (E) Dehiscent capsule showing five locules and the persistent five-angled columella (green arrowhead), specimen number NNF-1491. (F) Half capsule showing the persistent columella (green arrowhead) and reniform seed (red arrowhead), specimen number NNF-1499. (G) Dehiscent capsule with a deciduous valve, specimen number NNF-076. (H,I) Modern fruits of the species Schima supberba: (H) Partially dehiscent capsule showing reniform seeds. (I) Pentacarpellate, loculicidally dehiscent capsule showing apically 5-angled columella (green arrowhead). Scale bar = 2 mm. Processed and drawn by X. G. Shi using Adobe Photoshop CS5.
Locality
Santang Town of Nanning City, Guangxi Province, South China.
Stratigraphic Horizon
Yongning Formation, late Oligocene.
Repository
The Museum of Biology of Sun Yat-sen University (SYS), Guangzhou, China.
Etymology
The epithet “kwangsiensis” means the fossil is collected in Kwangsi (Guangxi) Province, China.
Specific Diagnosis
Pedicel 4–5 cm long; bracteoles 2–3; sepals 5, imbricate, persistent, slightly connate at base; capsule globose or depressed globose, pericarp woody, loculicidally splitting into 5–6 valves; columella persistent, stout, extending for 2/3 of locule length, apically 5-angled; seed reniform and flat, with a marginally membranous wing.
Description
Pedicel slender, apically thickened and recurved, bracteoles 2–3, caduous, away from sepals. Sepals 5, persistent, imbricate, basally slightly connate, semiorbicular, 2–3 mm in diameter. Capsule globose, 2–2.5 cm in diameter, pericarp woody, splitting for 1/2–2/3 length into 5–6 valves (Figs 3A,C–F and 4A–C); columella persistent, stout, extending for 2/3 or more of locule length, apically 5-angled with an enlarged face inside each of the five locules (Fig. 4D–E). Seeds small, reniform, flat, with a clear marginal membranous wings, 7.5–8 mm long, 4–5 mm wide including wing, sub-campylotropous attached to the columella. Hilum linear and short (Figs 4F–G and 5A,B). Seed coat with conspicuous protruding curved ridges that form irregularly reticulate ornamentations (Fig. 5C).

Seed and seed coat structures of Schima kwangsiensis sp. nov. and extant Schima superba. (A,B) The two sides of a reniform fossil seed and its wing, specimen number NNF-076. Scale bar = 1 mm. (C) Details of the seed coat structure of A, showing irregular reticulate epidermal ornamentation, Scale bar = 100 μm. (D,E) The two sides of a modern seed of the species Schima superba, Scale bar = 1 mm. (F) Details of the seed coat structure of D, showing regular reticulate epidermal ornamentation, Scale bar = 100 μm. Processed and drawn by X. G. Shi using Adobe Photoshop CS5.
Comparison
The new fossil species is characterized by its 5-loculed loculicidally dehiscent capsules, 5 imbricate sepals and pedicels with bracteoles. These features are consistent with the subfamily Theoideae of Theaceae and readily distinguish the fossils from fruits of other angiosperm families18. Modern Theoideae are divided into two main groups according to seed characters: one is composed of taxa bearing wingless seeds and includes Camellia, Tutcheria, Pyrenaria, Parapyrenaria, Apterosperma and Franklinia; the other encompasses taxa with winged seeds such as Gordonia, Laplacea, Schima, Hartia and Stewartia 6. Furthermore, all the genera with winged seeds can be further divided into two groups based on seeds having either apical or marginal wings. Those genera with marginally winged seeds mainly include Schima, Stewartia and Hartia, but the columella in fruit of Stewartia and Hartia is incomplete or only extends ca. 1/2 the length of the locule, easily distinguishing these genera. As for Franklinia, its immature seeds sometimes have diminished wings, but the dehiscence mode of the capsules in Franklinia is unique that they have both loculicidal dehiscence and septicidal dehiscence, and the capsule valves rarely adheres to the central part of the columella8, 9. Therefore, compared with other related genera, these fossil fruits belong to Schima because they have marginally winged seeds and persistent columellae extending most of the length of the locules.
Initially, the genus Schima was established in 1823, based on the type species S. noronhae Reinwardt ex Blume from Indonesia; the species has long pedicels, two caducous bracteoles, five suborbicular, ca.5 mm sepals, globose 5-loculed capsules and marginally winged seeds19. Since then, additional Schima species have been reported from China and adjacent areas. At present, it is generally acknowledged that there are a total of 20 species belonging to Schima, and 13 of them are distributed in China3. Compared with extant species of Schima, the new fossil species is distinguished by its longer pedicels, smaller sepals and larger globose fruits. Among extant Schima plants in China, the sepals are normally larger than 5 mm in diameter, and there are only two species (S. argentea and S. superba) with sepals measuring 2 mm in diam.; but they both have shorter pedicels (1–2 cm). Moreover, the size of the fruits is different. In fruit gross morphology S. superba can be viewed as the closest relative to the new fossil species.
To date, three fossil fruits of Schima have been reported, among which two species, S. macrocalycalis Mai and S. lignitica (Menzel) Mai, are from Germany14; another one, namely S. nanlinensis Li, Awasthi, Yang and Li., from China15. The fruit of S. lignitica is ovoid to oblong which can easily be distinguished from our fossils. The difference between the new species and S. macrocalycalis is that the latter has larger and suborbicular sepals. S. nanlinensis differs from the new fossil species in having depressed globose, and smaller capsules (0.8–1.1 cm). Detailed morphological comparison of the new species and similar species along within Schima is summarized in Table 1.
Systematic implication
Theaceae is a large and complex family in angiosperms that includes approximately 19 genera and 600 species3, 6. The classification of Theaceae has been disputed since the family was established. Traditionally, it is made up of two subfamilies Theoideae and Ternstroemioideae5, 20,21,22. However, a series of molecular studies suggested that the two subfamilies should be regarded as separate families10, 23,24,25. In APG III system of 2009, Ternstroemioideae was removed from the Theaceae to form the Ternstroemiaceae together with Pentaphylax (Pentaphylacaceae)26. Nevertheless, evolutionary trends within the Theaceae s.s. ( = Theoideae) remain controversial, which was shown by quite different classification systems6, 27. Specifically, the evolutionary significance of pedicel length has been subject to intense debate during the past few decades6,7,8. Keng (1980) even suggested that species with a long pedicel are more primitive than those with a short pedicel28, but the hypothesis is controversial due to the lack of definitive cladistic and fossil evidence. Hence, the mummified fossil fruits with intact pedicels described here could shed light on the early evolution of the genus Schima, and even of the family Theaceae.
In order to gain a clearer taxonomic position of Schima kwangsiensis within the genus, a clustering analysis based on eight fruit characters (Tables 2, 3) for two fossil species and 13 extant species of the genus in China was performed using a modification of Ward’s method in R package29. The result shows that the species within Schima, including both the fossil and extant species, may be divided into four groups. As shown in the dendrogram (Fig. 6), clade A consists of only two species S. brevipedicellata and S. multibracteata, and is distinguished from the others by having short pedicels and large sepals. Clade B, represented by S. sinensis, S. sericans, S. noronhae, S. remotiserrata, S. villosa, S. crenata, and S. khasiana, mainly bears long pedicels and large sepals. Clade C, including the new fossil S. kwangsiensis and two extant species S. superba, S. wallichii, is characterized by having long pedicels and small sepals (there is one exception). The last clade, comprising one fossil species S. nanlinensis, and two extant species, S. argentea and S. parviflora, has small sepals and short pedicels. Furthermore, among 13 extant species, eight taxa have long pedicels and five taxa have short pedicels. It is obvious that the two morphological characters, pedicel and sepal size, are tightly correlated with each other in the genus Schima (Fig. 6). In general, the cluster analysis shows that the new fossil species is closely similar in pedicel and calyx size to two extant species (S. wallichii and S. superba) in the genus, implying that they may belong to an ancient taxon that predated the others.

A cluster dendrogram of fossil and extant species of Schima in China based on fruit characters, generated and drawn by X. G. Shi using R (http://www.r-project.org/) and Adobe Photoshop CS5.
Paleogeography and Paleoecology
The origin and biogeographic history of Schima is little known because of the paucity of the fossil records. Up to now, only five definitive fossil occurrences have been reported in the world. Three fossil species are recognized on the basis of fruits, including Schima macrocalycalis from the middle Eocene of Germany, Schima lignitica from the late Miocene of Germany14, and Schima nanlinensis from the Miocene of Yunnan, China15. One wood fossil, Schima protowallichii occurs in the Miocene of Japan30, and a leaf fossil with well preserved cuticle was described as Schima mataschensis, is from the late Miocene of Styria, Austria31. In addition, one seed fossil, Schima euryoidesonce was reported from the late Eocene of Germany. However, its identity is doubtful as it has horseshoe-shaped embryos, a feature not found in Schima 32.
The Schima fruits reported here are the earliest fossils of this genus in Asia, and imply that this genus probably first appeared in Asia, its modern distribution area, by the late Oligocene. Moreover, the new fossil record is important because of the excellent preservation. Extant Schima has a small and lightweight seed with a marginal wing and the special structure of seeds aids long distance dispersal by wind6. The seeds of Schima fossils are morphologically similar to those of the extant genus implying that little change has occurred in the shape of seeds’ wing during the past 23 million years.
In addition to the fossils considered above, four species of Schimoxylon–a wood genus resembling Schima were reported, among which Schimoxylon dachelense was from the Upper Cretaceous of Egypt33, Schimoxylon g ordonioides was from the Tertiary deposits of Borneo of uncertain age34, Schimoxylon altingioides was from the Eocene of Germany35, and Schimoxylon benderi was from the late middle Eocene of Myanmar36 (Fig. 7). According to these fossil records, Schima possibly appeared by Late Cretaceous and achieved a widespread distribution from low-latitudes to middle latitudes during the Tertiary and was more widespread in the Northern Hemisphere than today. It suggests the ancient species of Schima probably originated in Northern Africa or Western Europe and further dispersed to the regions of Asia. However, the genus apparently experienced subsequent extinction both in Europe and in North Africa later and is now confined to subtropical and tropical zones within South China and Southeast Asia.

Map showing modern and fossil distribution of Schima and related fossil taxa, generated by X. G. Shi, using Arcgis 9.3 (ESRI, Redlands, CA, USA).
Extant Schima superba, which has similar fruits, is a tree up to ca. 30 m tall and occurs in evergreen broadleaved forests with a wide distribution in the subtropical and tropical montane areas at altitudes of 100–2500 m37. It is likely that Schima kwangsiensis may have been tall trees and grown in a similar environment. Additionally, preliminary investigations suggest that the mummified flora reported here mainly comprises Fagaceae, Theaceae, Bambusoideae, Anacardiaceae, Annonaceae, and Nyssaceae representing flowering plants, and Polyporaceae of the higher fungi16. These fossil taxa provide a glimpse of ancient ecological environments in Guangxi, indicating there has been a typical montane evergreen broad-leaved forest in southern China since at least the late Oligocene. The dominant species were mainly Schima kwangsiensis and some groups of Fagaceae in the tree layer, which grew under a warm moist forest environment.
Methods
The fossil fruits were exceptionally well-preserved, and include intact pedicels and seeds. The specimens were thoroughly washed in water and dried in air. They were observed and photographed using a stereoscopic microscope (Zeiss Stereo Discovery V20). The seed micromorphology was investigated using a scanning electronic microscope (SEM Quanta 400 F). Next, the resulting images were processed with Adobe Photoshop CS5 (Adobe Inc., San Jose, California, USA). The extant fruits of Schima superba for comparison were collected from Heishiding Nature Reserve, Guangdong Province. To prevent the potential fracturing on drying, all the megafossil specimens used herein are preserved in a mixed solution of 50% alcohol and 100% glycerol with the volume ratio of 10:1 and deposited at the Museum of Biology of Sun Yat-sen University (SYS), Guangzhou, China. The terminology used to describe the fruit and seed follows Keng8, and Min & Bartholomew3.
Clustering analysis was performed using R (http://www.r-project.org/). The Excel file containing the binary data was imported to R package and the 0/1 matrix was used to calculate Euclidean distance. The resultant distance matrix was employed to construct dendrograms using hierarchical cluster analysis with Ward’s algorithm to infer genetic relationships38.
References
Wu, Z. Y. The areal-types of Chinese Genera of seed plants. Acta BotanicaYunnanica, supplement IV, 1-139(1991).
Keng, H. Florae Malesianae precursors: 58. Part four, the genus Schima (Theaceae) in Malesia, Gardens’Bullentin. Singapore. 46(1), 77–87 (1994).
Ming, T. L., Bartholomew, B. Theaceae. In: Wu ZY, Raven PH, editor. Flora of China. Vol 12. Beijing:Science Press and Missouri Botanical Garden Press, St. Louis (2007).
Melchior, H. T. In: Engler A. Prantl E. eds. Die naturlichen pflanzenfamilien. 2nd ed. Leipzig: Wilhelm Engelmann. 21,109-154 (1925).
Takhtajan, A. Diversity and classification of flowering plants. New York: Columbia university Press (1997).
Chang, H. T., Ren, S. X. Theacae, in: Flora Reipublicae popularis Sinicae. Beijing: Science Press. 1–251 (1998).
Sealy J. R. A revision of the genus Camellia. London: Royal Horticultural Society (1958).
Keng, H. Comparative morphological studies in Theaceae. University of California Publications in Botany. 33, 269–384 (1962).
Ye, C. X. A discussion on relationship among genera in Theoideae (Theaceae). Acta Scientiarum Naturallium Universitatis Sunyatseni. 29, 74–81 (1990).
Prince, L. M. & Parks, C. R. Phylogenetic relationships of Theaceae inferred from chloroplast DNA sequence data. American Journal of Botany. 88, 2309–2320 (2001).
Yang, J. B. et al. Phylogenetic relationships of Theaceae inferred from mitochondrial matR gene sequence data. Acta BotanicaYunnanica. 28, 29–36 (2006).
Li, M. M., Li, J. H., Tredici, P. D., Corajod, J. & Fu, C. X. Phylogenetics and biogeography of Theaceae based on sequences of plastid genes. Journal of Systematics and Evolution. 51(4), 396–404 (2013).
Zhang, W., Kan, S. L., Zhao, H., Li, Z. Y. & Wang, X. Q. Molecular phylogeny of Tribe Theeae (Theaceaes.s.) and Its Implications for Generic Delimitation. Plos One 9(5), e98133, doi:10.1371/journal.pone.0098133 (2014).
Mai, D. H. Uber fossile Lauraceae und Theaceae in Mitteleuropa. Feddes Repertorium. 82(5), 313–342 (1971).
Li, Y., Awasthi, N., Yang, J. & Li, C. S. Fruits of Schima (Theaceae) and seeds of Toddalia (Rutaceae) from the Miocene of Yunnan Province, China. Review of Palaeobotany and Palynology. 193, 119–127 (2013).
Quan, C. et al. First Oligocene mummified plant Lagerstätte at the low latitudes of East Asia. Science China Earth Sciences. 59(3), 445–448 (2016).
Zhao, Z. New Anthracothere materials from the Paleogene of Guangxi. Verteb PalAsia. 31, 13–190 (1993).
Cronquist, A. An integrated system of classification of flowering plants. New York: Columbia University Press (1981).
Bentham, G., Hooker, J. D. Genera Plantarum. Vol. I.185 (1862).
Airy-Shaw, H. K. Notes on the genus Schima and on the classification of the Theaecae-Camellioideae. Kew Bulletin. 496-500 (1936).
Hutchison, J. Evolution and phylogeny of flowering plants. London: Academic Press (1969).
Thorne, R. F. Classfication and geography of flowering plants. Bot. Rev. 58, 225–348 (1992).
Soltis, D. E. et al. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal of the Linnean Society. 133, 381–461 (2000).
Anderberg, A. A., Rydin, C. & Kallersjo, M. Phylogenetic relationships in the order Ericaless. l: analyses of molecular data from five genes from the plastid and mitochondrial genomes. American Journal of Botany. 89(4), 677–687 (2002).
Schonenerger, J., Anderberg, A. A. & Sytsma, K. J. Molecular phylogenetics and patterns of floral evolution in the Ericales. International journal of plant Sciences. 166(2), 265–288 (2005).
APG. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society. 161, 105–121 (2009).
Ming, T. L. Studies on the genus Camellia in the world. Kunming, Yunnan Science and Technology Press (2000).
Keng, H. On the unification of Laplacea and Gordonia (Theaceae). Garden’s Bulletin, Singapore. 33, 303–311 (1980).
Murtagh, F. & Legendre, P. Ward’s hierarchical agglomerative clustering method: which algorithms implement Ward’s criterion? Journal of Classification. 31(3), 274–295 (2014).
Choi, S. K. et al. Fossil woods from the Miocene in the Yamagata Prefecture, Japan. IAWA J 31, 95–117 (2010).
Kovar-Ender, J. & Hably, L. The flora of Mataschen—a unique plant assemblage from the late Miocene of eastern Styria (Austria). Acta Palaeobotanica. 46(2), 157–233 (2006).
Grote, P. J. & Dilcher, D. L. Investigations of angiosperms from the Eocene of North America: A new genus of the Theaceae based on fruit and seed remains. Botanical Gazette. 152, 190–206 (1989).
Kramer, K. D. T.-H. Sudost-Asiens (unter Ausschluss der Dipterocarpaceae) 2. Teil. Palaeontographica Abteilung B. 145, 1–150 (1974).
Bande, M. B. & Prakash, U. The Tertiary flora of Southeast Asia with remarks on its palaeoenvironment and phytogeography of the Indo-Malayan region. Review of Palaeobotany and Palynology. 49, 203–233 (1986).
Gottwald, H. P. J. Hölzeraus Marinen Sanden des OberenEozän von Helmstedt (Niedersachsen). Palaeontogr. Abt. B. 225, 27–103 (1992).
Licht, A. et al. Fossil woods from the late middle Eocene Pondaung Foration, Myanmar. Review of Palaeobotany and Palynology. 202, 29–46 (2014).
Yang, S. X., Deng, H. & Li, M. S. Manganese uptake and accumulation in a woody hyperaccumulator, Schima superb. Plant, Soil and Environment. 10, 441–446 (2008).
Joe, H. Ward, Jr. Hierarchical Grouping to Optimize an Objective Function. Journal of the American Statistical Association. 58, 236–244 (1963).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant Nos. 41372002, 41210001, 41528201), the Joint Project of the National Natural Science Foundation of China and the Russian Foundation for Basic Research (Grant Nos. 41611130044, 16–55–53007), and the Fundamental Research Funds for the Central Universities (Grant no. 16lgjc28). We thank graduate students majoring in Plant Science at Sun Yat-sen University for participating in the field. We also offer our sincere gratitude to Prof. Robert A. Spicer (The Open University, UK) for improving English.
Author information
Authors and Affiliations
Contributions
C.Q., Q.Y.F. and J.H.J. did the field work. C.Q. and J.H.J. designed the research. Q.Y.F. took photos for the specimens. X.G.S., J.H.J. and C.Q. wrote the paper. All authors read and agreed with the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shi, XG., Fu, QY., Jin, JH. et al. Mummified Oligocene fruits of Schima (Theaceae) and their systematic and biogeographic implications. Sci Rep 7, 4009 (2017). https://doi.org/10.1038/s41598-017-04349-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-04349-6
- Springer Nature Limited
This article is cited by
-
The earliest large carpenter bee (Xylocopa) and its adhering pollen (Araliaceae, Theaceae)
Palaeobiodiversity and Palaeoenvironments (2024)
-
Incompatible pollen abortion and late-acting self-incompatibility in Schima superba
Scientific Reports (2022)
-
Species richness, forest types and regeneration of Schima in the subtropical forest ecosystem of Yunnan, southwestern China
Forest Ecosystems (2020)
-
Putrescine modifies the pollen tube growth of tea (Camellia sinensis) by affecting actin organization and cell wall structure
Protoplasma (2020)