

Abstract
Bacteria communicate and coordinate their behaviour at the intra- and interspecies levels by producing and sensing diverse extracellular small molecules called autoinducers. Autoinducer 2 (AI-2) is produced and detected by a variety of bacteria and thus plays an important role in interspecies communication and chemotaxis. Although AI-2 is a major autoinducer molecule present in the mammalian gut and can influence the composition of the murine gut microbiota, its role in bacteria–bacteria and bacteria–host interactions during gut colonization remains unclear. Combining competitive infections in C57BL/6 mice with microscopy and bioinformatic approaches, we show that chemotaxis (cheY) and AI-2 signalling (via lsrB) promote gut colonization by Escherichia coli, which is in turn connected to the ability of the bacteria to utilize fructoselysine (frl operon). We further show that the genomic diversity of E. coli strains with respect to AI-2 signalling allows ecological niche segregation and stable co-existence of different E. coli strains in the mammalian gut.






Similar content being viewed by others

Data availability
The publicly available E. coli genome database and the associated files (annotations, phylogenetic tree) used and reprocessed in the present study can be found under: https://microbiology.figshare.com/articles/dataset/A_comprehensive_and_high-quality_collection_of_E_coli_genomes_and_their_genes/13270073. Any additional data can be requested from the corresponding author. Source data are available for Figs. 1–5 and Extended Data Figs. 1–10.
Code availability
Customized code used to produce the results presented in the present study is available at https://github.com/lukasmalfi/E_Coli.
References
Colin, R., Sourjik, V. & R Colin, V. S. Emergent properties of bacterial chemotaxis pathway. Curr. Opin. Microbiol. 39, 24–33 (2017).
Milo, R., Jorgensen, P., Moran, U., Weber, G. & Springer, M. BioNumbers—the database of key numbers in molecular and cell biology. Nucleic Acids Res. 38, D750–D753 (2010).
Ni, B., Colin, R., Link, H., Endres, R. G. & Sourjik, V. Growth-rate dependent resource investment in bacterial motile behavior quantitatively follows potential benefit of chemotaxis. Proc. Natl Acad. Sci. USA 117, 595–601 (2020).
Matilla, M. A. & Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 42, 40–67 (2018).
Matilla, M. A. et al. Chemotaxis of the human pathogen Pseudomonas aeruginosa to the neurotransmitter acetylcholine. mBio https://doi.org/10.1128/MBIO.03458-21 (2022).
Lopes, J. G. & Sourjik, V. Chemotaxis of Escherichia coli to major hormones and polyamines present in human gut. ISME J. 12, 2736 (2018).
Yang, J. et al. Biphasic chemotaxis of Escherichia coli to the microbiota metabolite indole. Proc. Natl Acad. Sci. USA 117, 6114–6120 (2020).
Colin, R., Ni, B., Laganenka, L. & Sourjik, V. Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiol. Rev. 45, 1–19 (2021).
Keegstra, J. M., Carrara, F. & Stocker, R. The ecological roles of bacterial chemotaxis. Nat. Rev. Microbiol. 20, 491–504 (2022).
Liou, M. J. et al. Host cells subdivide nutrient niches into discrete biogeographical microhabitats for gut microbes. Cell Host Microbe https://doi.org/10.1016/J.CHOM.2022.04.012 (2022).
McCormick, B. A., Laux, D. C. & Cohen, P. S. Neither motility nor chemotaxis plays a role in the ability of Escherichia coli F-18 to colonize the streptomycin-treated mouse large intestine. Infect. Immun. 58, 2957 (1990).
Leatham, M. P. et al. Mouse intestine selects nonmotile flhDC mutants of Escherichia coli MG1655 with increased colonizing ability and better utilization of carbon sources. Infect. Immun. 73, 8039–8049 (2005).
Song, S. & Wood, T. K. The primary physiological roles of autoinducer 2 in Escherichia coli are chemotaxis and biofilm formation. Microorganisms 9, 386 (2021).
Laganenka, L., Colin, R. & Sourjik, V. Chemotaxis towards autoinducer 2 mediates autoaggregation in Escherichia coli. Nat. Commun. 7, 12984 (2016).
Jani, S., Seely, A. L., Peabody V, G. L., Jayaraman, A. & Manson, M. D. Chemotaxis to self-generated AI-2 promotes biofilm formation in Escherichia coli. Microbiology https://doi.org/10.1099/mic.0.000567 (2017).
Pereira, C. S., Thompson, J. A. & Xavier, K. B. AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 37, 156–181 (2013).
Ismail, A. S., Valastyan, J. S. & Bassler, B. L. A host-produced autoinducer-2 mimic activates bacterial quorum sensing. Cell Host Microbe 19, 470–480 (2016).
Valastyan, J. S., Kraml, C. M., Pelczer, I., Ferrante, T. & Bassler, B. L. Saccharomyces cerevisiae requires cff1 to produce 4-hydroxy-5-methylfuran-3(2h)-one, a mimic of the bacterial quorum-sensing autoinducer AI-2. mBio 12, 1–17 (2021).
Zhang, L. et al. Sensing of autoinducer-2 by functionally distinct receptors in prokaryotes. Nat. Commun. 11, 1–13 (2020).
Laganenka, L. & Sourjik, V. Autoinducer 2-dependent Escherichia coli biofilm formation is enhanced in a dual-species coculture. Appl. Environ. Microbiol. 84, e02638-17 (2018).
Thompson, J. A., Oliveira, R. A., Ubeda, C., Xavier, K. B. & Djukovic, A. Manipulation of the quorum sensing signal AI-2 affects the antibiotic-treated gut microbiota. Cell Rep. 10, 1861–1871 (2015).
Hsiao, A. et al. Members of the human gut microbiota involved in recovery from Vibrio cholerae infection. Nature 515, 423–426 (2014).
Wotzka, S. Y. et al. Microbiota stability in healthy individuals after single-dose lactulose challenge—a randomized controlled study. PLoS ONE 13, e0206214 (2018).
Jensen, K. F. The Escherichia coli K-12 ‘wild types’ W3110 and MG1655 have an rph frameshift mutation that leads to pyrimidine starvation due to low pyrE expression levels. J. Bacteriol. 175, 3401–3407 (1993).
Blattner, F. R. et al. The complete genome sequence of Escherichia coli K-12. Science 277, 1453–1462 (1997).
Soupene, E. et al. Physiological studies of Escherichia coli strain MG1655: growth defects and apparent cross-regulation of gene expression. J. Bacteriol. 185, 5611–5626 (2003).
Hobman, J. L., Penn, C. W. & Pallen, M. J. Laboratory strains of Escherichia coli: model citizens or deceitful delinquents growing old disgracefully? Mol. Microbiol. 64, 881–885 (2007).
Leatham, M. P. et al. Precolonized human commensal Escherichia coli strains serve as a barrier to E. coli O157:H7 growth in the streptomycin-treated mouse intestine. Infect. Immun. 77, 2876–2886 (2009).
Stecher, B. et al. Flagella and chemotaxis are required for efficient induction of Salmonella enterica serovar Typhimurium colitis in streptomycin-pretreated mice. Infect. Immun. 72, 4138–4150 (2004).
Stecher, B. et al. Motility allows S. typhimurium to benefit from the mucosal defence. Cell Microbiol. 10, 1166–1180 (2008).
Thompson, J. A., Oliveira, R. A. & Xavier, K. B. Chemical conversations in the gut microbiota. Gut Microbes 7, 163–170 (2016).
González Barrios, A. F. et al. Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022). J. Bacteriol. 188, 305–316 (2006).
Bansal, T., Jesudhasan, P., Pillai, S., Wood, T. K. & Jayaraman, A. Temporal regulation of enterohemorrhagic Escherichia coli virulence mediated by autoinducer-2. Appl. Microbiol. Biotechnol. 78, 811–819 (2008).
Xavier, K. B. & Bassler, B. L. Regulation of uptake and processing of the quorum-sensing autoinducer AI-2 in Escherichia coli. J. Bacteriol. 187, 238–248 (2005).
Xavier, K. B. et al. Phosphorylation and processing of the quorum-sensing molecule autoinducer-2 in enteric bacteria. ACS Chem. Biol. 2, 128–136 (2007).
Hegde, M. et al. Chemotaxis to the quorum-sensing signal AI-2 requires the Tsr chemoreceptor and the periplasmic LsrB AI-2-binding protein. J. Bacteriol. 193, 768–773 (2011).
Neumann, S., Hansen, C. H., Wingreen, N. S. & Sourjik, V. Differences in signalling by directly and indirectly binding ligands in bacterial chemotaxis. EMBO J. 29, 3484–3495 (2010).
Oliveira, R. A. et al. Klebsiella michiganensis transmission enhances resistance to Enterobacteriaceae gut invasion by nutrition competition. Nat. Microbiol. 5, 630–641 (2020).
Luo, C. et al. ConStrains identifies microbial strains in metagenomic datasets. Nat. Biotechnol. 33, 1045–1052 (2015).
Tyakht, A. V. et al. Genetic diversity of Escherichia coli in gut microbiota of patients with Crohn’s disease discovered using metagenomic and genomic analyses. BMC Genom. 19, 1–14 (2018).
Pereira, F. C. & Berry, D. Microbial nutrient niches in the gut. Environ. Microbiol. 19, 1366–1378 (2017).
Conway, T. & Cohen, P. S. Commensal and pathogenic Escherichia coli metabolism in the gut. Microbiol. Spectr. https://doi.org/10.1128/microbiolspec.MBP-0006-2014 (2015).
Meador, J. P., Caldwell, M. E., Cohen, P. S. & Conway, T. Escherichia coli pathotypes occupy distinct niches in the mouse intestine. Infect. Immun. 82, 1931–1938 (2014).
Brito, P. H., Rocha, E. P. C., Xavier, K. B. & Gordo, I. Natural genome diversity of AI-2 quorum sensing in Escherichia coli: conserved signal production but labile signal reception. Genome Biol. Evol. 5, 16–30 (2013).
Stecher, B. et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl Acad. Sci. USA 109, 1269–1274 (2012).
Riley, M. A. & Gordon, D. M. The ecological role of bacteriocins in bacterial competition. Trends Microbiol. 7, 129–133 (1999).
Marques, J. C. et al. LsrF, a coenzyme A-dependent thiolase, catalyzes the terminal step in processing the quorum sensing signal autoinducer-2. Proc. Natl Acad. Sci. USA 111, 14235–14240 (2014).
Laganenka, L. et al. Quorum sensing and metabolic state of the host control lysogeny-lysis switch of bacteriophage T1. mBio https://doi.org/10.1128/mBio.01884-19 (2019).
Schembri, M. A., Hjerrild, L., Gjermansen, M. & Klemm, P. Differential expression of the Escherichia coli autoaggregation factor antigen 43. J. Bacteriol. 185, 2236–2242 (2003).
Horesh, G. et al. A comprehensive and high-quality collection of Escherichia coli genomes and their genes. Microb. Genom. 7, 1–15 (2021).
Wiame, E., Delpierre, G., Collard, F. & Van Schaftingen, E. Identification of a pathway for the utilization of the Amadori product fructoselysine in Escherichia coli. J. Biol. Chem. 277, 42523–42529 (2002).
Erbersdobler, H. F. & Faist, V. Metabolic transit of Amadori products. Mol. Nutr. Food Res. 45, 177–181 (2001).
Wolf, A. R. et al. Bioremediation of a common product of food processing by a human gut bacterium. Cell Host Microbe 26, 463–477.e8 (2019).
Barroso-Batista, J. et al. Specific eco-evolutionary contexts in the mouse gut reveal Escherichia coli metabolic versatility. Curr. Biol. 30, 1049–1062.e7 (2020).
Frazão, N., Sousa, A., Lässig, M. & Gordo, I. Horizontal gene transfer overrides mutation in Escherichia coli colonizing the mammalian gut. Proc. Natl Acad. Sci. USA 116, 17906–17915 (2019).
Sourjik, V. & Berg, H. C. Functional interactions between receptors in bacterial chemotaxis. Nature 428, 1–4 (2004).
Sourjik, V. & Berg, H. C. Receptor sensitivity in bacterial chemotaxis. Proc. Natl Acad. Sci. USA 99, 123–127 (2002).
Sourjik, V., Vaknin, A., Shimizu, T. S. & Berg, H. C. In vivo measurement by FRET of pathway activity in bacterial chemotaxis. Methods Enzymol. 423, 365 (2007).
Laganenka, L., López, M. E., Colin, R. & Sourjik, V. Flagellum-mediated mechanosensing and RflP control motility state of pathogenic Escherichia coli. mBio https://doi.org/10.1128/mBio.02269-19 (2020).
Somavanshi, R., Ghosh, B. & Sourjik, V. Sugar influx sensing by the phosphotransferase system of Escherichia coli. PLoS Biol. 14, e2000074 (2016).
Ortega, Á., Zhulin, I. B. & Krell, T. Sensory repertoire of bacterial chemoreceptors. Microbiol. Mol. Biol. Rev. 81, e00033-17 (2017).
Wang, L., Hashimoto, D., Tsao, C. Y., Valdes, J. J. & Bentley, W. E. Cyclic AMP (cAMP) and cAMP receptor protein influence both synthesis and uptake of extracellular autoinducer 2 in Escherichia coli. J. Bacteriol. 187, 2066–2076 (2005).
Ha, J.-H. et al. Evidence of link between quorum sensing and sugar metabolism in Escherichia coli revealed via cocrystal structures of LsrK and HPr. Sci. Adv. 4, eaar7063 (2018).
Graf von Armansperg, B. et al. Transcriptional regulation of the Nε-fructoselysine metabolism in Escherichia coli by global and substrate-specific cues. Mol. Microbiol. 115, 175–190 (2021).
Adler, J. Chemotaxis in bacteria. Science 153, 708–716 (1966).
Koster, D. A., Mayo, A., Bren, A. & Alon, U. Surface growth of a motile bacterial population resembles growth in a chemostat. J. Mol. Biol. 424, 180–191 (2012).
Laganenka, L. & Sourjik, V. Autoinducer 2-dependent Escherichia coli biofilm formation is enhanced in a dual-species co-culture. Appl. Environ. Microbiol. https://doi.org/10.1128/AEM.02638-17 (2017).
Gauger, E. J. et al. Role of motility and the flhDC Operon in Escherichia coli MG1655 colonization of the mouse intestine. Infect. Immun. 75, 3315–3324 (2007).
de Paepe, M. et al. Trade-off between bile resistance and nutritional competence drives Escherichia coli diversification in the mouse gut. PLoS Genet. https://doi.org/10.1371/journal.pgen.1002107 (2011).
Monday, S. R., Minnich, S. A. & Feng, P. C. H. A 12-base-pair deletion in the flagellar master control gene flhC causes nonmotility of the pathogenic German sorbitol-fermenting Escherichia coli O157:H− strains. J. Bacteriol. 186, 2319–2327 (2004).
Conway, T. & Cohen, P. S. Applying the restaurant hypothesis to intestinal microbiota: anaerobes in mixed biofilms degrade polysaccharides, sharing locally prepared sugars with facultative anaerobes that also colonize the intestine. Microbe 10, 324–328 (2015).
Long, Z., Quaife, B., Salman, H. & Oltvai, Z. N. Cell-cell communication enhances bacterial chemotaxis toward external attractants. Sci. Rep. 7, 1–12 (2017).
Molloy, M. J. et al. Intraluminal containment of commensal outgrowth in the gut during infection-induced dysbiosis. Cell Host Microbe 14, 318–328 (2013).
Haag, L. M. et al. Intestinal microbiota shifts towards elevated commensal Escherichia coli loads abrogate colonization resistance against Campylobacter jejuni in mice. PLoS ONE https://doi.org/10.1371/journal.pone.0035988 (2012).
Spees, A. M. et al. Streptomycin-induced inflammation enhances Escherichia coli gut colonization through nitrate respiration. mBio 4, e00430-13 (2013).
Carvalho, F. A. et al. Transient inability to manage proteobacteria promotes chronic gut inflammation in TLR5-deficient mice. Cell Host Microbe 12, 139–152 (2012).
Datsenko, K. A. & Wanner, B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl Acad. Sci. USA 97, 6640–6645 (2000).
Cherepanov, P. P. & Wackernagel, W. Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene 158, 9–14 (1995).
Furter, M., Sellin, M. E., Hansson, G. C. & Hardt, W. D. Mucus architecture and near-surface swimming affect distinct Salmonella Typhimurium infection patterns along the murine intestinal tract. Cell Rep. 27, 2665–2678.e3 (2019).
Valdivia, R. H. & Falkow, S. Bacterial genetics by flow cytometry: rapid isolation of Salmonella Typhimurium acid-inducible promoters by differential fluorescence induction. Mol. Microbiol. 22, 367–378 (1996).
Barthel, M. et al. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect. Immun. 71, 2839–2858 (2003).
Miller, K. A., Phillips, R. S., Kilgore, P. B., Smith, G. L. & Hoover, T. R. A mannose family phosphotransferase system permease and associated enzymes are required for utilization of fructoselysine and glucoselysine in Salmonella enterica serovar Typhimurium. J. Bacteriol. 197, 2831–2839 (2015).
Johnson, M. et al. NCBI BLAST: a better web interface. Nucleic Acids Res. 36, W5–W9 (2008).
Mckinney, W. Pandas: a foundational python library for data analysis and statistics. Python High Perform. Sci. Comput. 14, 9 (2011).
Szklarczyk, D. et al. The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 49, D605–D612 (2021).
Huerta-Cepas, J., Serra, F. & Bork, P. ETE 3: reconstruction, analysis, and visualization of phylogenomic data. Mol. Biol. Evol. 33, 1635–1638 (2016).
Menardo, F. et al. Treemmer: a tool to reduce large phylogenetic datasets with minimal loss of diversity. BMC Bioinform. 19, 1–8 (2018).
Harris, C. R. et al. Array programming with NumPy. Nature 585, 357–362 (2020).
Pereira, C. S. et al. Phosphoenolpyruvate phosphotransferase system regulates detection and processing of the quorum sensing signal autoinducer-2. Mol. Microbiol. 84, 93–104 (2012).
Acknowledgements
We thank K. Xavier (Instituto Gulbenkian de Ciência, Oeiras, Portugal) for generously providing the E. coli ARO071 strain and for helpful discussions. We also thank the EPIC RCHCI staff for support of the animal work. L.L. is supported by a grant (no. LA 4572/1-1) from the Deutsche Forschungsgemeinschaft. This work has been further funded by grants from the Swiss National Science Foundation (SNF; grant nos. 310030B_173338 and 310030_192567, NCCR Microbiomes) to W.-D.H. V.S. acknowledges support by the Hessian Ministry of Higher Education, Research, and the Arts–LOEWE research cluster ‘Diffusible Signals’ subproject A1. J.W.L. was supported by a grant (no. NRF-2019R1A6A3A03031885) from the National Research Foundation, Republic of Korea. C.v.M. is supported by the Swiss NSF (grant no. 310030_192569). C.L.D. and J.P. are supported by a grant (no. SNF 205321L_10724) from the Swiss NSF.
Author information
Authors and Affiliations
Contributions
L.L., W.-D.H. and V.S. conceived and designed the experiments. L.L. and J.-W.L. performed the experiments. L.M. and C.v.M. performed bioinformatic analysis. C.L.D., L.F. and J.P. synthesized fructoselysine. All authors contributed to data analysis and writing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Microbiology thanks Mariana Byndloss and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 E. coli Z1331 colonizes ampicillin-pretreated SPF mice without causing inflammation.
a, c.f.u. of E. coli Z1331 WT (yidX-bla, ampr) detected in faeces (F) and caecal content (CC) of ampicillin-pretreated SPF mice at different time points of a 72 h infection. Lines indicate median values (mice n = 9, from ≥2 independent animal experiments). The slight drop of fecal E. coli densities between 48 h and 72 h.p.i. is likely due to the regrowth of microbiota. Dashed line indicates the detection limit. b, Lipocalin-2 levels in faeces (F) and caecal content (CC) of E. coli-infected mice as measured by ELISA. Lines represent median values (mice n = 5, from ≥2 independent animal experiments). Dashed line indicates approximate threshold of lipocalin-2 concentration marking a shift from non-inflamed to the inflamed gut, as observed in the streptomycin mouse model for Salmonella diarrhea. Note that gut colonization by wild type S. Typhimurium yields lipocalin-2 levels of 104 ng/g faeces during full-blown gut inflammation1. c, Competitive indices (C.I.) for chemotaxis-deficient ΔcheY strains from different phylogroups in competition against the respective WT strains in SPF ampicillin-pretreated mice. F, faeces. CC, caecal content. Lines indicate median values (minimum mice n = 5, at least two independent replicates). Dashed line indicates C.I. value of 1.
Extended Data Fig. 2 E. coli Z1331 ΔcheY has no colonization defect in single-strain infection.
a, c.f.u. of E. coli Z1331 WT and ΔcheY detected in faeces (F) and caecal content (CC) of ampicillin-pretreated SPF mice at different time points of a 72 h infection. Lines indicate median values (mice n = 4, two independent replicates). b, Number of aggregates formed by WT and ΔcheY cells in a single-strain infection normalized to the number of detected cells in a tissue section as seen below (two-tailed Mann-Whitney test, **P < 0.005). Lines indicate median values (image sample n = 11, tissue sections from two independent experiments were analyzed). c, Caecal tissue sections of mice infected either with E. coli WT (mCherry-positive, shown in orange) or ΔcheY (GFP-positive, shown in green) at 72 h.p.i. Actin filaments (red) and DNA (blue) were stained with phalloidin and DAPI, respectively. Scale bars, 50 µm. d, An example of image segmentation and analysis of bacterial aggregates (as seen above) using ImageJ. Detected particles are indicated in red, with aggregates (at least 50 px2 in size) outlined in yellow. Particles of non-bacterial origin (food fibers etc, as seen in ΔcheY panel) were manually excluded from analysis. Scale bars, 50 µm.
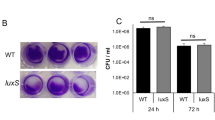
Extended Data Fig. 3 Increased luminal AI-2 levels abolish fitness advantage of wild-type E. coli in ΔlsrB/WT competitive infection.
a, AI-2 levels of AI-2 in faeces of SPF mice before (SPF Amp−) and 24 h after (SPF Amp+) treatment with 20 mg ampicillin. Mean fluorescence of a plasmid-based AI-2 reporter strain was measured by flow cytometry and plotted in arbitrary units (a.u.). Lines indicate median values (mice n = 4, at least two independent replicates). P values were calculated using two-tailed Mann-Whitney test (*P < 0.05). b, AI-2 levels of AI-2 in faeces (F) of SPF ampicillin-pretreated mice infected with E. coli Z1331 WT (-ARO071) or with 1:1 mix of E. coli Z1331 WT and E. coli ARO071. Mean fluorescence of a plasmid-based AI-2 reporter strain was measured by flow cytometry and plotted in arbitrary units (a.u.). Lines indicate median values (mice n = 6, at least two independent replicates). P values were calculated using two-tailed Mann-Whitney test (**P < 0.005). c, c.f.u. data for the experiment shown in Fig. 2d. F, faeces, CC, caecal content. Lines indicate median values (mice n = 6, at least two independent replicates). P values were calculated using two-tailed Mann-Whitney test (**P < 0.005; ns, not significant). The dashed line indicates the detection limit. Note that the total c.f.u. loads can differ between caecum and faeces due to yet unidentified reasons.
Extended Data Fig. 4 Competitive indices (C.I.) for ΔlsrB strains of lsr-positive E. coli W3110 and 8550 in competition against the respective WT strains in SPF ampicillin-pretreated mice.
F, faeces. CC, caecal content. Lines indicate median values (minimum mice n = 5, at least two independent replicates). Dashed line indicates C.I. value of 1.
Extended Data Fig. 5 CheY and LsrB belong to the same regulatory pathway.
E. coli Z1331 ΔcheY and ΔcheY ΔlsrB knockout strains were competed against the wild-type strain. Additionally competitive indices (C.I.) of ΔlsrB and ΔcheY mutants were analyzed in ΔcheY and ΔlsrB backgrounds, respectively. F, faeces, CC, caecal content. Lines indicate median values (minimum mice n = 5, from at least two independent infection experiments). P values were analyzed using two-tailed Mann-Whitney test (**P < 0.005; *P < 0.05; ns, not significant).
Extended Data Fig. 6 Self-produced AI-2 enhances gut colonization by E. coli.
a, Experimental scheme of competitive infection in germ-free (GF) mice. C57BL/6 J GF mice were orally infected with 5×107 c.f.u. E. coli W3110 WT and ΔlsrB or ΔlsrB ΔluxS and ΔluxS at a 1:1 ratio. Faeces were collected 24, 48 h.p.i. and mice were euthanized at 72 h.p.i. b, C.I. of non-AI-2 chemotactic ΔlsrB mutant in WT and ΔluxS background strains in the GF mouse infection model. F, faeces, CC, caecal content. Lines indicate median values (mice n = 9, from least two independent experiments). P values were calculated using two-tailed Mann-Whitney test (****P < 0.0001). Dashed line indicates C.I. value of 1. c, Experimental scheme of competitive infection in SPF mice. C57BL/6 J SPF mice were pretreated with 20 mg ampicillin by oral gavage 24 h prior to infection with E. coli W3110 WT and ΔlsrB or ΔlsrB ΔluxS and ΔluxS at 1:1 ratio. Faeces were collected at 24, 48 h.p.i and mice were euthanized at 72 h.p.i. d, C.I. of non-AI-2 chemotactic ΔlsrB mutant in WT and ΔluxS background strains in SPF ampicillin-pretreated mouse infection model. F, faeces, CC, caecal content. Lines indicate median values (minimum mice n = 5, from at least two independent experiments). P values were calculated using two-tailed Mann-Whitney test (**P < 0.005). Dashed line indicates C.I. value of 1.
Extended Data Fig. 7 Infection of SPF ampicillin-pretreated mice with E. coli 8178 and 8850 does not cause inflammation.
a, H&E staining of caecal tissue of uninfected mice (PBS) and mice infected with 5×107 c.f.u. of E. coli Z1331 WT + 8178 WT and E. coli Z1331 WT + 8850 WT (1:1000 ratio) at 72 h.p.i (as seen in Fig. 4). Scale bar, 50 µm. b, Histopathology analysis of the caecal tissue section as seen above. 3 sections from 2 mice per group were analyzed. c, Lipocalin-2 levels in faeces (F) and caecal content (CC) of E. coli-infected mice as measured by ELISA. Lines represent median values (mice n = 7, at least two independent animal experiments). Dashed line indicates approximate threshold of lipocalin-2 concentration marking a shift from non-inflamed to the inflamed gut. d, Colonization levels of E. coli 8178 in competition experiments with E. coli Z1331 as seen in Fig. 2. Lines indicate median values (mice n = 7, at least two independent replicates). P values were calculated using two-tailed Mann-Whitney test (ns, not significant). e, Colonization levels of E. coli 8850 in competition experiments with E. coli Z1331 as seen in Fig. 2. Lines indicate median values (minimum mice n = 6, at least two independent replicates). P values were calculated using two-tailed Mann-Whitney test (ns, not significant).
Extended Data Fig. 8 Fructoselysine is an attractant sensed by the Trg chemoreceptor.
Examples of FRET measurements of the response to fructoselysine (reflected by the ratio of YFP/CFP fluorescence) by E. coli W3110 a, wild-type, b, Δtrg, c, Δtsr, d, Δtar, e, Δtap, f, ΔptsI and g, ΔcheA (negative control) knockout strains. Buffer-adapted cells were stimulated with step-like addition and removal of compounds (indicated by downward and upward arrows, respectively). Stimulation with saturating concentration of aspartate or serine, two strong attractants, was used as a positive control. Time traces of fluorescence intensity in the YFP (shown in yellow) and CFP channels (shown in blue) are shown in the right. Opposite changes in two channels indicate specific FRET response. Note that higher concentrations of fructoselysine solution have unspecific effect on fluorescence in both YFP and CFP channels, particularly visible in ΔcheA negative control, but little effect on the YFP/CFP ratio. Residual effect on the YFP/CFP ratio in the negative control was subtracted from all dose-response curves in Fig. 5b.
Extended Data Fig. 9 LsrB and Tsr belong to the same regulatory pathway.
Competitive indices (C.I.) of E. coli Δtsr and Δtsr ΔlsrB mutant strains vs the wild-type strain E. coli Z1331 in SPF ampicillin-pretreated mice. F, faeces, CC, caecal content. Lines indicate median values (minimum mice n = 6, at least two independent replicates). P values were analyzed using two-tailed Mann-Whitney test (ns, not significant).
Extended Data Fig. 10 E. coli Z1331 utilizes fructoselysine as a sole carbon source.
E. coli Z1331 WT, ΔfrlA and ΔptsI strains were grown aerobically for 24 h in M9 minimal medium supplemented with either 1% fructoselysine (FL) or 2% arabinose (non-PTS sugar, used as a control for ΔptsI growth) and NH4Cl as a nitrogen source. Mean optical densities are shown, error bars indicate s.d. (sample n = 6, from at least two independent experiments). P values were calculated using two-tailed Mann-Whitney test (**P < 0.005; ns, not significant).
Supplementary information
Supplementary Information
Supplementary Figs. 1–3 and Information Tables 1 and 2.
Supplementary Tables 1 and 2
Supplementary Table 1: Strains and plasmids used in this study. Supplementary Table 2: Correlation analysis of the E. coli lsrB gene.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Laganenka, L., Lee, JW., Malfertheiner, L. et al. Chemotaxis and autoinducer-2 signalling mediate colonization and contribute to co-existence of Escherichia coli strains in the murine gut. Nat Microbiol 8, 204–217 (2023). https://doi.org/10.1038/s41564-022-01286-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-022-01286-7
- Springer Nature Limited
This article is cited by
-
Spatial structure, chemotaxis and quorum sensing shape bacterial biomass accumulation in complex porous media
Nature Communications (2024)