

Abstract
The vertebrate brain emerged more than ~500 million years ago in common evolutionary ancestors. To systematically trace its cellular and molecular origins, we established a spatially resolved cell type atlas of the entire brain of the sea lamprey—a jawless species whose phylogenetic position affords the reconstruction of ancestral vertebrate traits—based on extensive single-cell RNA-seq and in situ sequencing data. Comparisons of this atlas to neural data from the mouse and other jawed vertebrates unveiled various shared features that enabled the reconstruction of cell types, tissue structures and gene expression programs of the ancestral vertebrate brain. However, our analyses also revealed key tissues and cell types that arose later in evolution. For example, the ancestral brain was probably devoid of cerebellar cell types and oligodendrocytes (myelinating cells); our data suggest that the latter emerged from astrocyte-like evolutionary precursors in the jawed vertebrate lineage. Altogether, our work illuminates the cellular and molecular architecture of the ancestral vertebrate brain and provides a foundation for exploring its diversification during evolution.
Similar content being viewed by others
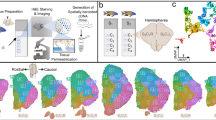

Main
The vertebrate brain is a structurally complex and preeminent organ because of its central functions in the body. Its most fundamental divisions are the forebrain (prosencephalon, traditionally divided into the telencephalon and diencephalon), the midbrain (mesencephalon) and the hindbrain (rhombencephalon) (Fig. 1a). This regionalization is shared across all extant jawed vertebrates and is present even in jawless vertebrates (that is, the extant cyclostomes: lampreys and hagfishes), the sister lineage of jawed vertebrates (gnathostomes)1 (Fig. 1a), which have overall less complex brains than jawed vertebrates2. While a basic molecular regionalization has been described for the substantially simpler central nervous systems (CNSs) of the closest evolutionary relatives of vertebrates (urochordates and cephalochordates)3,4,5, the anatomical complexity of the four major divisions of the vertebrate brain evolved in common vertebrate ancestors ~515–645 million years ago (Ma)6 (Fig. 1a), probably as part of the cephalic expansion that commenced around the emergence of this animal lineage (the ‘new head’ hypothesis)7.

a, Top: phylogenetic tree displaying the main vertebrate lineages and their approximate brain anatomies; the blue bar indicates the estimated confidence interval for the divergence time of cyclostomes and gnathostomes6. Bottom: schematic of the adult sea lamprey brain showing the different regions dissected for this study. b, UMAP of brain cells (all scRNA-seq data combined) coloured according to their corresponding cell type groups. c, Dendrogram describing the relationships between the identified cell types. The coloured boxes correspond to the highlighted cell type groups in b. Top: expression of terminal selector marker genes within each cell type; the circle sizes are proportional to the number of cells expressing the gene. Bottom: binary expression (presence/absence, based on whether a given gene is differentially expressed in the corresponding cell type; Methods) of effector genes (neurotransmitters for neuronal types). PF, posterior forebrain; SC, spinal cord; 1, PNS glia; 2, erythrocytes. d–f, Sagittal sections (same orientation as in a) of the adult brain showing ISS maps of genes marking neurons (STMN4), ependymoglia (SLC1A2) and meningeal fibroblasts (PAH and PDGFRA) (d); anterior forebrain versus posterior forebrain and midbrain neuronal factors (e); and neurotransmitter genes (f). The dashed lines separate the main four brain regions illustrated in a. See Supplementary Fig. 2 for the ISS section schemes. Scale bars, 500 μm. The lamprey gene symbols throughout this study are based on the corresponding mouse orthologue names. When a lamprey gene corresponds to multiple mouse genes (one-to-many orthologous relationships), both gene names are indicated, using a slash (/) for separation.
Previous anatomical and molecular studies of the vertebrate brain have yielded intriguing insights and hypotheses pertaining to its structural and functional evolution8,9. However, its ancestral cellular composition and underlying gene expression programs, as well as its subsequent diversification, have not been systematically explored.
To fill this critical gap, we generated a comprehensive cell type atlas of the adult and larval (ammocoete) brain of the sea lamprey (Petromyzon marinus), based on extensive transcriptomic and spatial expression data at single-cell resolution (https://lampreybrain.kaessmannlab.org/). Integrated comparative analyses of this atlas unveiled details of the cell type repertoire and molecular architecture of the ancestral vertebrate brain but also revealed distinct cell types, gene expression programs and tissue structures that emerged during the evolution of the brain in jawed and jawless vertebrates.
Cellular and molecular organization of the lamprey brain
We generated single-cell RNA-sequencing (scRNA-seq) data (21 libraries in total) for whole adult and ammocoete brains, as well as separately for their four major anatomical regions (telencephalon, diencephalon, mesencephalon and rhombencephalon), to facilitate cell type assignments (Fig. 1a and Supplementary Tables 1 and 2). To ensure optimal scRNA-seq read mapping, we substantially refined and extended previous annotations of the lamprey germline genome10 (Extended Data Fig. 1 and Supplementary Data 1) on the basis of 63 deeply sequenced RNA-seq libraries covering six major organs, including different brain regions (Supplementary Tables 1 and 2 and Methods). After quality control and data filtering (Methods), we obtained transcriptomes for a total of 159,381 high-quality cells (72,810 for adults and 86,571 for ammocoetes). Using a detailed clustering approach and an iterative marker-gene-based annotation procedure (Methods), we identified 151 (95 neuronal) distinct cell types in the adult dataset and 120 (92 neuronal) in the larval dataset (Supplementary Table 3; see the online atlas). To spatially localize cell types across the brain, we generated in situ sequencing (ISS)11 data for 93 selected marker genes in both lamprey life stages and single-molecule RNA fluorescence in situ hybridization (smRNA-FISH) images for four genes in the larval stage (Supplementary Table 4 and Supplementary Data 2).
Overall, neural cell type compositions are similar between the two stages (Extended Data Figs. 2–4a). However, we noted a generally higher cell type specificity of gene expression patterns in adults than in ammocoetes (Extended Data Fig. 4c)—a result that is robust to controls for technical differences between datasets (Extended Data Fig. 4b–d and Methods).
A cell type tree derived from the datasets for the adult lamprey, which is thought to be better suited for the inference of ancestral vertebrate traits than ammocoetes12, reflects cell type relationships based on gene expression distances (Fig. 1b,c). This tree unveils the hierarchical organization of cell types in the lamprey brain (Fig. 1c). The primary division is between neuronal and non-neuronal cell types, which are in turn split into ependymoglial cells (that is, neural-tube-derived glia) and other cells (that is, vascular cells, meningeal fibroblasts, blood cells and glial cells from the peripheral nervous system (PNS)). Our spatial ISS data illustrate that these three major cell type classes occupy very distinct areas of the brain (Fig. 1d).
At a secondary hierarchical level, non-neuronal cells are organized according to their cell class identity (for example, astrocytes, ependymal cells, erythrocytes and immune cells), in agreement with their molecular phenotype (Fig. 1c). By contrast, the organization of neuronal types primarily reflects their anatomical origin. A first separation is thus evident between telencephalic, anterior diencephalic (that is, hypothalamus and pre-thalamus), pineal and habenular neurons on one side of the neuronal clade, and posterior diencephalic (that is, thalamus and pre-tectum), mesencephalic and rhombencephalic/spinal cord neurons on the other. Within each developmental subdivision, neurons are organized according to their neurotransmitter phenotype (Fig. 1c).
The overall hierarchical cell type organization of the lamprey brain is supported by the expression patterns of terminal selectors (that is, sets of transcription factors (TFs) that determine and maintain cell type identity13,14) and effector genes (that is, sets of genes that characterize the molecular phenotype of cells) (Fig. 1c). Inhibitory neurons, for instance, are regulated mainly by DLX genes in the anterior forebrain15 but by GATA2/3, OTX2 and TAL genes in the posterior forebrain, midbrain, hindbrain and spinal cord16,17 (Fig. 1c and Extended Data Fig. 5a; gene names are based on the respective names of the mouse orthologue(s)—see Methods for details regarding the gene nomenclature used in this study). Our ISS data confirm this strict compartmentalization of neuronal regulators (Fig. 1e and Extended Data Fig. 5h,i,l,m). Conversely, neurotransmitter-related genes are expressed across different brain regions (Fig. 1c,f).
The hierarchical relationship of cell types in the lamprey brain is very similar to that observed for a reference mammalian brain atlas (that is, that of the mouse18), which suggests that all vertebrates share a common general cellular and molecular organization of neural tissues that was established during the evolution of the vertebrate stem lineage.
Vertebrate cell type families
To illuminate the cell type composition and molecular architecture of the ancestral vertebrate brain and to uncover differences between the CNSs of cyclostomes and gnathostomes, we performed detailed comparative analyses of our adult lamprey atlas with a corresponding atlas established for the mouse18. The neuronal and non-neuronal cells of the two atlases were contrasted separately using a dedicated method for homologous cell type detection (self-assembling manifold mapping (SAMap))19 and a correlation-based analysis of gene expression that also considers paralogous genes and was adapted from a previous approach20 (Methods).
The SAMap results show a great degree of correspondence between the two species for groups of cell types belonging to the same class (for example, vascular cells, astrocytes and excitatory neurons of the telencephalon), as indicated by the uniform manifold approximation and projection (UMAP) of the inter-species manifold (Fig. 2a,b) and the distribution of mapping scores between the two atlases (Fig. 2e,f, Extended Data Fig. 6 and Supplementary Table 5). This high-level similarity is confirmed by cross-species dendrograms based on the correlation approach applied to orthologous TF genes (Fig. 2c,d and Extended Data Fig. 7). These observations suggest that many of the corresponding cell classes share evolutionarily related gene expression programs (Supplementary Fig. 1). We propose that the matching groups of cell types uncovered in these analyses might constitute homologous cell type ‘families’ (ref. 21) that were already present in the brain of the last common ancestor of jawless and jawed vertebrates more than ~515–645 Ma6.

a,b, SAMap results displaying UMAPs of non-neuronal (a) and neuronal (b) cells from both species. Erythrocytes and oligodendrocytes were removed from the lamprey and mouse datasets, respectively. c,d, Dendrograms reporting gene expression distance (Pearson’s r) of TF genes of non-neuronal (c) and neuronal (d) cell type groups from the two species. BS, bootstrap support (n = 1,000). e,f, Sankey diagrams relating non-neuronal (e) and neuronal (f) cell type groups between the two species based on SAMap mapping scores (min = 0.1; max = 0.65). The link width is proportional to the mapping score. DG, dentate gyrus; SVZ, sub-ventricular zone.
Blood, vascular and PNS cells
The blood cells found in the lamprey brain can be classified into erythrocytes, characterized by the massive expression of haemoglobin and haeme-related genes (for example, ALAS1/2), and immune cells, which are mainly composed of microglia/macrophages and lymphocytes (Extended Data Fig. 8a). The microglia/macrophage cell types are highly correlated to mammalian perivascular macrophages and microglia (Fig. 2c,e and Supplementary Table 5) and express genes that are typically related to the non-specific immune response (for example, GRN, CSF1R and HCK/LYN; Extended Data Fig. 8a–c) both outside (macrophages) and inside (microglia) the brain (Extended Data Fig. 8d). We also identified a lymphocytic cell population (type: Lympho2) expressing one of the two known cyclostome-specific variable lymphocyte receptor genes (VLRA; Extended Data Fig. 8a), which is part of a distinct adaptive immune system that emerged in the cyclostome lineage in parallel to that of gnathostomes22.
We identified several vascular cell types, corresponding to endothelial cells/pericytes, which express typical vascular markers (for example, EPAS1 and KDR; Extended Data Fig. 8a–c) and are principally localized at the innermost meningeal layer (Extended Data Fig. 8d), forming the perineural vascular plexus23. The inner and outer leptomeningeal layers are populated by fibroblast-like cells (type: Fibro1) that are probably homologous to the meningeal vascular fibroblasts described in the mouse brain18,24, given their high respective homology mapping scores (Fig. 2c,e and Supplementary Table 5) and the expression of key orthologous marker genes (for example, PDGFRA, FOXC1 and LUM; Extended Data Fig. 8a–d). A second fibroblast type (Fibro2) occupies the space between the leptomeningeal boundaries (Extended Data Fig. 8d) and is characterized by the expression of genes involved in the metabolism of glucose (G6PC)25, fatty acids (FABP3), cholesterol (SOAT1/2) and aromatic amino acids (PAH) (Extended Data Fig. 8a) This cell type might correspond to meningeal round cells, which form a metabolically active tissue typical of lamprey that is not present in the meninges of other vertebrates23,25.
PNS glia are represented by a small cluster (n = 53) expressing the orthologues of the mouse TF genes Sox10 and Sox9 (denominated SOXE2 and SOXE3 in lamprey, respectively26); they co-localize with cranial nerve roots (Extended Data Fig. 8a,e). This group of cells, which most likely corresponds to the previously described peripheral ensheathing glia27, expresses some markers whose mouse orthologues are characteristic of satellite glia (SOXE2) and Schwann cells and their precursors (EGR2/3/4 and PMP22/EMP3) (Extended Data Fig. 8a). However, they lack the expression of key peripheral myelin constituent genes such as MPZ and PMP2, confirming the absence of actual myelin in the lamprey PNS28. Together with the co-clustering of this cell type with mouse satellite glia and Schwann cells (Fig. 2c), our observations strongly support and extend the hypothesis that lamprey PNS ensheathing glia are homologous to mammalian Schwann cells/precursors. The co-localization of this cell type with meningeal fibroblasts and vascular smooth muscle cells in the cell type tree (Extended Data Fig. 8a) probably reflects their common developmental origin from the neural crest18.
Ependymoglial cells and the origin of myelination
Our analyses revealed that ependymoglial cells (that is, CNS glia) in lamprey are divided into two main, developmentally related, cell classes: ependymal-like cells and astrocyte-like cells, referred to as ‘ependymal’ and ‘astrocytes’ hereafter, given the observations described below. Ependymal cells are ciliated, epithelial-like cells that populate the ventricular system of the brain, the circumventricular organs29 and the choroid plexuses30 and are characterized by the expression of the ciliogenesis-related TF FOXJ1 and the extracellular matrix component CCN2/3/5 (Extended Data Fig. 9a,e,f,i). We identified two types of specialized secretory ependymal types in the lamprey brain: choroid plexus epithelial cells (OTX2+), responsible for the production of cerebrospinal fluid (CSF), and hypendymal cells of the sub-commissural organ (SCO), which massively express the main Reissner’s fibres component SCO-spondin (SSPO)31 (Extended Data Fig. 9a,b,g,i). Two additional types of specialized ependymal cells are the pigmented pineal epithelial cells, defined by markers that are common to the retina pigment epithelium (for example, RPE65 and RRH; Extended Data Fig. 9a), and the KERA-expressing ependymal cells of the hindbrain and spinal cord (types: ReEpen1 and ReEpen3; Extended Data Fig. 9a,c,d). The large number of detected ependymal cells and cell types in the adult dataset (Extended Data Fig. 3b) probably reflects the large relative sizes of the ventricles and choroid plexuses of the lamprey brain (Extended Data Fig. 9i)32.
Notably, lamprey astrocytes are highly comparable to those from mouse in terms of their overall transcriptome signature (Fig. 2c,e). They share key marker genes that are fundamental for the development and function of astrocytes, such as SOXE3 (Sox9), HES5 and SLC1A2 (Fig. 3a and Extended Data Fig. 9a). However, like in other anamniotes (for example, fishes and amphibians), lamprey astrocytes are mainly localized around the ventricles (Fig. 3 and Extended Data Fig. 9h), forming the so-called ependymo-radial glia33.

a,b, UMAPs showing the expression of astrocyte-specific (a) and oligodendrocyte-specific (b) orthologous genes in the mouse (top) and lamprey (middle) atlases. Bottom: ISS maps of the adult lamprey brain for the same genes, showing coronal sections of the telencephalon (a) and sagittal sections of the whole brain (b; same orientation as in Fig. 1a). See Supplementary Fig. 2 for the ISS section schemes. Scale bars, 500 μm.
Like in the PNS, lamprey CNS axons are not myelinated28, consistent with the absence of key master regulators of oligodendrocyte identity (OLIG1 and OPALIN) and myelin-specific genes (MOBP and TSPAN2) in its genome. Other myelin-related genes are present in the genome, but they are not expressed in glial cells (for example, PDGFRA and NKX6-1/2 are expressed in meningeal fibroblasts; Extended Data Fig. 8a). Notably, despite the lack of myelination, lamprey astrocytes express several oligodendrocyte-specific genes, such as the TFs NKX2-2 and SOXE2 (Sox10)34 (Fig. 3b and Extended Data Fig. 9a), the proteolipid gene PLP1/GPM6B (orthologous to the myelin components Plp1 and Gpm6b) (Fig. 3b and Extended Data Fig. 9a), and the extracellular matrix glycoproteins TNR and HEPACAM (Extended Data Fig. 9a,j,k). Given the expression of crucial TFs of oligodendrocyte identity and the presence of myelin-related genes within lamprey astrocytes, our findings lend strong support to the hypothesis that oligodendrocytes originated from astrocyte-like glia in gnathostome ancestors27.
Neuronal diversity across brain regions
Finally, we scrutinized neuronal cell types across the different brain regions. Hindbrain and spinal cord neurons are defined by the expression of several HOX genes (HOXA/B3, HOXA/B4 and HOXA/B5; Extended Data Fig. 5a,b). Two types of hindbrain glycinergic cells (ReInh5 and ReInh6), probably corresponding to inhibitory reticulospinal neurons35, are highly correlated to reticular neurons of the medulla in mouse18 (Supplementary Table 5) and express related markers (SLC6A5, SLC32A1b and EBF2/3; Extended Data Fig. 5a,b,f,g). Cholinergic neurons expressing the TF gene TBX6/20 show very localized expression within the hindbrain, probably corresponding to afferent nuclei of cranial nerves36 (Extended Data Fig. 5c–e). None of the detected midbrain/hindbrain clusters specifically express markers related to Purkinje (for example, ALDOC, PCP2, SLC1A6 and CAR8) or granule (for example, NEUROD1, CBLN1 and GABRA6) neurons of the cerebellum, nor are these markers expressed in the dorsal isthmic region (Supplementary Data 2). We also did not detect the expression of marker genes in this region that are associated with neurons of inferred ancestral cerebellar nuclei37, which were shown to have diversified in the gnathostome lineage through duplications37. These observations confirm the absence of proper cerebellar nuclei in the lamprey brain37,38. Within the rostral spinal cord, we identified two types of GABAergic CSF-contacting cells39 (ReInh1 and ReInh2); these are ciliated neurons that are homologous to the gnathostome CSF-contacting neurons of the spinal cord central canal and express genes coding for channels that respond to changes in CSF pH (PKD2L1 and PKD2L2) and for proteins that remove toxic oxidative compounds from the CSF (AMBP) (Extended Data Fig. 5a,b).
Thalamic, pre-tectal and tectal neurons are divided into excitatory and inhibitory classes (Extended Data Fig. 5a) and express TFs that are typical of homologous anatomical regions in mouse (that is, thalamus, pre-tectum and superior colliculus)18. In fact, like in the murine brain, glutamatergic neurons are characterized by the expression of SHOX2, EBF1 and EBF2/3, whereas GABAergic neurons express GATA2/3a, GATA2/3b, TAL1 and OTX2 (Figs. 1e and 4a and Extended Data Fig. 5a,f–m).

a–e, ISS maps of selected neuronal marker genes across caudal and rostral coronal sections through the diencephalon (a,b) and telencephalon (c–e). f, UMAP showing LGE-derived and MGE-derived GABAergic neurons of the telencephalon. g, Dendrogram showing the relationships between the clusters in f and the expression of selected marker genes. Dhy, dorsal hypothalamus; LH, left habenula; Pal, pallium; Pald, pallidum; PpT, parapineal tract; PS, pineal stalk; MH, medial habenula; OT, optic tectum; PCN, postoptic commissure nucleus; PTN, posterior tubercle nucleus; RH, right habenula; RPa, rostral paraventricular area; St, striatum; VHy, ventral hypothalamus. See Supplementary Fig. 2 for ISS section schemes; scale bars, 500 μm.
Epithalamic neurons (that is, neurons stemming from the dorsal-most region of the diencephalon) are divided into habenular types and pineal/parapineal photoreceptors, like in the gnathostome brain40. All habenular neurons express the same TFs (NR4A2, ETV1 and IRX2/5; Extended Data Fig. 10a,e), with the medial and lateral nuclei showing very distinct expression patterns for several genes (for example, MYO9A, PRKCQ, GNG2 and TMEM64; Fig. 4b and Extended Data Fig. 10a,d). The medial habenula is occupied by glutamatergic, nitrergic and cholinergic neurons41,42 (Fig. 1c and Extended Data Fig. 10a,d), with a cell type expressing neuropeptide Y (NPY; Extended Data Fig. 10a,d). The lateral habenulae are molecularly related to each other; they co-express several markers (GNG2, TMEM64 and SLC1A3/6a; Fig. 4b and Extended Data Fig. 10a,d) and can be distinguished by the differential expression of two neuropeptide genes: proenkephalin (PENK; right) and cholecystokinin-like (CCK-like; left) (Extended Data Fig. 10f).
The pineal and parapineal of the lamprey are directly photosensitive organs with neuroendocrine outputs43. We detected signatures of both aspects of these organs at the molecular level by the expression of the genes CRX (necessary for the differentiation of photoreceptors), GUCA1B (involved in visual phototransduction), LHX3/4 and ISL1/2 (required for the development of retinal photoreceptors44,45, as well as of neuroendocrine cells in the mammalian anterior pituitary46) in all detected cell types (Extended Data Fig. 10a,o,r). We could assign four clusters to these organs: predominantly in the pineal, we detected cone opsin-expressing (type: Photo1) and rod opsin-expressing (Photo2) cells defined by their expression of marker genes commonly associated with cones and rods, including arrestin, phosphodiesterase and GRKs. The pineal and parapineal organs differ in their expression of RCVRN and genes involved in the biosynthesis of melatonin (Extended Data Fig. 10a,n,p,q and the online atlas). More prominent in the parapineal organ, types Photo3 and Photo4 express the non-visual opsin gene parietopsin and the neuropeptide gene TAC1 (Extended Data Fig. 10a). Unlike the pineal stalk, characterized by the expression of pineal markers, the parapineal ganglion and tract cells are marked by genes also detected in the habenulae (for example, PPP1R14A/B/C and GNG2; Extended Data Fig. 10k,n,o), consistent with reports that the secondary (downstream) neurons of the parapineal are segregated away from the photoreceptors of the parapineal vesicle47.
Nearly all monoaminergic neurons, identified by the expression of monoamine transport (SLC18A1a and SLC18A1b) and metabolic (TH and TPH1/2) genes, form a unique taxon within the cell type tree (Fig. 1c and Extended Data Fig. 10a), which includes serotoninergic and dopaminergic neurons of the hindbrain, midbrain and hypothalamus. Dopaminergic neurons of the posterior tubercle nucleus of the hypothalamus (type: MeDopa1) co-express dopamine-related and glutamate-related genes48 (Extended Data Fig. 10a,c) and are considered homologues of the dopaminergic neurons of the substantia nigra pars compacta of amniotes49, an important component of basal ganglia. These cells are located next to NTS-producing neurons50 (a modulator of dopaminergic activity51; Fig. 4a) and express the TF PROX1 (Fig. 1c), which is crucial for the development of dopaminergic posterior tubercle nucleus cells in zebrafish52.
Like in the mouse brain atlas18, most hypothalamic peptidergic neurons co-cluster with monoaminergic cells (Fig. 1c and Extended Data Fig. 10a). Neurons of the ventral hypothalamus and postoptic commissure nucleus express the neuropeptide genes CCK53 and pro-opiomelanocortins (POMCa and POMCb) (Fig. 4b and Extended Data Fig. 10e,h), as well as the circadian-rhythm-related genes SIX3/6a and PER1/2 (also expressed in the pineal complex; see the online atlas). Other neuropeptides expressed in the hypothalamus are galanin (GAL)54, somatostatins (SSTa and SSTc)55, NPY56, neurotensin (NTS), vasotocin (VAT), PENK, prepronociceptin (PNOC), gonadotropin-releasing hormones (GNRH1 and GNRH2), prolactin-releasing hormone (PRLH) and FAM237A/B (Extended Data Fig. 10a,d–h). Additional peptidergic neurons cluster with inhibitory neurons of the pallium/sub-pallium (Extended Data Fig. 10a); these include GAL+ neurons of the septum and preoptic area (type: DePep10) and glutamatergic neurons expressing VAT, GNRH1 and NEUROD2/6a (type: DePep9) located in the rostral paraventricular area of the preoptic area (Fig. 4c,d and Extended Data Fig. 10a,n).
Inhibitory neurons of the telencephalon are classified into olfactory bulb (OB) and pallium/sub-pallium cell types and are all enriched for typical forebrain GABAergic markers (GAD1/2, DLX1a, DLX1b and DLX2/3/5; Extended Data Figs. 5l,m and 10a). OB neurons can be recognized by (1) the conserved expression of several TFs that are characteristic of the anterior forebrain and placodes in chordates3 (for example, SP8/9, PAX6, FOXG1 and ETV1; Extended Data Fig. 10a,s), (2) the unique expression of PRDM12 (expressed in pain-sensing nerve cells and V1 interneurons in gnathostomes57,58; Extended Data Fig. 10a,t), and (3) the presence of dopaminergic cells (type: TeDopa1; Extended Data Fig. 10a).
SP8/9+ neurons are also present in the sub-pallium (type: TeInh4), within a region traditionally considered to correspond to the medial preoptic nucleus (MPO)59 (Fig. 4d), where they co-express ISL1/2 and TAC1, both markers of striatal projection neurons in gnathostomes (Fig. 4g and Extended Data Fig. 10a). The presence of SP8/9+–ISL1/2+ and SP8/9+–ETV1+ neurons in the sub-pallium and OB, respectively, is already known for mammals, where they originate from the lateral ganglionic eminence (LGE)60, suggesting that these two cell populations share the same developmental origin and migratory patterns across vertebrates.
Another important sub-pallial progenitor zone in jawed vertebrates is the medial ganglionic eminence (MGE). We identified neurons (type: TeInh5) expressing LHX6/8 and NKX2-1/4a (both markers of MGE-derived cells in mammals) (Fig. 4g) within two sub-pallial regions: (1) dorsal to the MPO (Fig. 4d), in a region traditionally called ‘striatum’ (ref. 61), and (2) the putative pallidum62, a nucleus located ventrolateral to the thalamic eminences (Fig. 4e). Recursive clustering revealed the presence of subtypes that express markers that are typical of MGE-derived neurons of the sub-pallial amygdala (SPA) and pallidum in jawed vertebrates63 (for example, TACR1, GBX1 and SOX6; Fig. 4g).
The presence of DLX+ GABAergic neurons expressing LGE-related and MGE-related markers in the pallium (Extended Data Fig. 10j,o) implies that migration from progenitor zones of the sub-pallium also occurs in lamprey. Many of these neurons express the neuropeptide genes PENK and SSTc, which mark GABAergic interneuron types in the pallium of several gnathostome species64,65,66 (Extended Data Fig. 10n,o,q,r). The vasoactive intestinal peptide (a marker of a subpopulation of cortical GABAergic interneurons in amniotes) is also present in the lamprey pallium, but, contrary to gnathostomes, it is expressed exclusively in glutamatergic neurons (Fig. 4c and Extended Data Fig. 10a,p).
The expression programs of excitatory neurons of the lamprey telencephalon are overall highly correlated to those of the corresponding cell types in mouse (Fig. 2f). This similarity is confirmed by the expression of marker genes typical of mammalian cortical glutamatergic neurons within the lamprey pallium67 (previously denoted ‘lateral pallium’ (ref. 59); see also the discussion below) and, partially, OB (for example, TBR1, EMX1/2a, EMX1/2b, RTN4R, LHX2/9, BCL11B and IKZF1/3; Fig. 5b–d and Extended Data Figs. 5h,i and 10a,g–i,k,l,q,v). We identified eight distinct cell types populating four different regions of the lamprey dorsal telencephalon and anterior diencephalon: (1) dorsomedial telencephalic nucleus (DMTN; type: TeExc1), (2) anterior pre-thalamic eminence (a region previously believed to correspond to the ‘medial pallium’ (ref. 2); see also the discussion below) (PThE; type: TeExc4), (3) pallial extended amygdala (PEA; type: TeExc3) and (4) pallium (types: TeExc2 and TeExc5–8) (Fig. 5a,b,f and Supplementary Table 3; see the online atlas). DMTN is a relay nucleus that is innervated by tufted-like cells of the OB68 and is located at the interface between the pallium and OB, of which it constitutes the caudal-most portion. Like the OB, the DMTN displays a layered structure with outer glutamatergic neurons, which share the same expression profile with cells of the OB glomerular layer (for example, EBF1) and inner GABAergic (PRDM12+) neurons (Extended Data Fig. 10p,r–t,v,w). PThE and PEA neurons express the TFs OTX2 and NR2F1/6a and are defined by the expression of EBF1, SSTc (TeExc4) and C1QL3, PNOC (TeExc3) (Fig. 5b and Extended Data Fig. 10a,i,o,s). We found that pallial neurons form a three-layered cortex with an inner GABAergic/glutamatergic layer, a middle glutamatergic layer and an external molecular, fibre-rich layer, in accord with previous work69 (Fig. 5g). They all express multiple genes associated with cortical projection neurons in amniotes (for example, FOXP1/2/4, MEIS2, LAMP5, RORB and TCAP; Extended Data Fig. 10a). However, contrary to what is known for amniotes and, since recently, also for amphibians (that is, for tetrapods in general)70, we did not observe any regional specification of gene expression patterns among these neurons (for example, dorsal, lateral or ventral) that could be related to known, functionally distinct areas of the pallium (for example, somatosensory, visual, motor or olfactory), as previously observed on the basis of connectivity data71,72.

a, UMAP of excitatory neurons of the telencephalon highlighting the different regions. b, Dendrogram showing the relationships between the clusters in a and the expression of selected marker genes. c,d, Spatial localization of the clusters of a and b and the expression (ISS) of selected marker genes (highlighted in bold in b) in caudal (c) and rostral (d) coronal sections through the telencephalon. e, Section scheme of c, d, f and g. f, Spatial expression (ISS) of selected marker genes within the dorsal telencephalon. g, Top left: spatial expression of glutamatergic (RTN4R) and GABAergic (DLX1b) marker genes in the telencephalon. Top right: magnification from the dashed square showing the layered organization of the pallium; each neuronal class is highlighted by plotting the expression of multiple specific marker genes: GABA (GAD1/2, DLX1a, DLX1b and DLX2/3/5) and Glut (SLC17A/6/7/8 and TBR1). Bottom: spatial scatter plots highlighting the positions of GABAergic (right) and glutamatergic (left) neurons within the pallium. PTh, pre-thalamus. Scale bars, 500 μm.
Discussion
In this study, we used extensive scRNA-seq and targeted spatial transcriptomics data to create a neural cell type atlas for a cyclostome representative: the sea lamprey (https://lampreybrain.kaessmannlab.org/). Our cell type tree analyses revealed that lampreys and gnathostomes share a common fundamental cellular and molecular organization of the brain that emerged in the vertebrate stem lineage more than ~515–645 Ma. This finding is in line with previous studies, such as the shared broad brain regionalization (Figs. 1a and 6a) and previously described patterning mechanisms across vertebrates1. Our comparisons of lamprey and mouse cell types revealed homologous relationships for many cell type families; that is, we identified groups of cell types partly sharing the same gene expression programs. These cell type families probably constituted the core of the ancestral vertebrate cell type repertoire.

a, Vertebrate phylogenetic tree as in Fig. 1a showing key brain innovations (in red) as indicated, shown or confirmed by our study. b,c, Our interpretation of lamprey telencephalic regions; the names in parentheses indicate the original designation of the region before this study. The diagrams represent caudal (b) and rostral (c) coronal sections through the telencephalon. d, Scheme for sections in b and c. LPal, lateral pallium.
Our analyses of non-neuronal cells revealed the presence of two distinct cell types within the lamprey ependymoglia that are probably homologous to ependymal cells and astrocytes of gnathostomes (Fig. 1c), suggesting that two of the three main macroglial cell types (astrocytes, ependyma and oligodendrocytes) were already established in the common vertebrate ancestor. Notably, however, our work confirms the absence of oligodendrocytes and sheds new light on their origination. We found that lamprey astrocytes express several oligodendrocyte-specific genes, including master regulators and effector genes (Fig. 3b). Our observations suggest that key components of the molecular machinery of oligodendrocytes were present in astrocyte-like cells of the vertebrate ancestor, and indicate that oligodendrocytes originated from these evolutionary precursors in the gnathostome lineage (Fig. 6a). Our work thus extends previous studies, which showed that lamprey axons seem to be physically associated with astrocytes27 and that key aspects of the regulatory program required for oligodendrocyte differentiation in gnathostomes are present during lamprey gliogenesis34. While our data also confirm the absence of actual myelin in the lamprey PNS3, our study lends strong support to the previous hypothesis4 that lamprey PNS glia are homologous to both mammalian Schwann and satellite cells.
Furthermore, our analyses did not provide evidence for the presence of granule or Purkinje cells in the rostral hindbrain, strongly supporting the notion that the mature lamprey brain lacks a proper cerebellum. We note, however, that a recent study detected the expression of granule and Purkinje cell TFs in the dorsal rhombomere 1 of lamprey embryos73. A targeted prospective analysis of the dorsal isthmic region in the adult and developmental lamprey brain might thus reveal the presence of potential rare homologues of cerebellar cell types.
The discovery of both LGE-derived and MGE-derived inhibitory neurons in the lamprey telencephalon confirms that the two main GABAergic progenitor zones of the sub-pallium were already present in the common vertebrate ancestor8,70 (Figs. 4f and 6a). Our findings challenge the traditional neuroanatomy view regarding the localization of the main sub-pallial regions of lampreys. Previous studies2,59 used to locate the striatum dorsal to the MPO and ventrolateral to the pallium (Fig. 4d). In this study, however, we identified a group of LGE-derived neurons (type: TeInh4), located in the MPO, that express the genes ISL1/2, TAC1 and PENK (Fig. 4g), whose orthologues are typical markers of projection neurons of the dorsal striatum in jawed vertebrates (medium spiny neurons). This evidence indicates that the MPO of lampreys is in fact homologous to the dorsal striatum of jawed vertebrates and that it should be renamed accordingly (Fig. 6b). The region traditionally considered to correspond to the striatum, in contrast, is populated by MGE-derived cells (type: TeInh5) that express the markers LHX6/8, GAL, GBX1 and SOX6 (Fig. 4c,d,g), whose orthologues are expressed in the same combination in the SPA of jawed vertebrates. We therefore propose that this region corresponds to the SPA and not to the striatum, as previously believed2,59 (Fig. 6b). MGE-derived cells can also be found caudal to the SPA, where they form the pallidum (Figs. 4e and 6c). Outside the lamprey sub-pallium, LGE-derived and MGE-derived cells also contribute to GABAergic interneurons of the OB and pallium, indicating that their migratory patterns are conserved across vertebrates (Fig. 6a).
Our analysis of the dorsal telencephalon confirms the hypothesis that the region previously denoted ‘medial pallium’ in lamprey is actually a rostral enlargement of the PThE74 (that is, it is part of the diencephalon). This notion is supported by the expression of genes that are typically expressed in pre-thalamic excitatory neurons in both lampreys and jawed vertebrates (EBF1 and EBF2/3) in the corresponding lamprey cell type (TeExc4), located in the previously denoted medial pallium, and by the absence of expression of marker genes that are typically associated with excitatory neurons of the pallium (FOXG1 and EMX1/2) (Fig. 5b–d,f). Our data also indicate the presence of a PEA in lamprey, in support of a previous hypothesis74, and show that it is located dorsal to the SPA. This region is populated by cells (type: TeExc3) that express markers of the extended amygdala in mouse18,75 (LMO3 and PNOC) (Figs. 5b–d,f and 6b).
Within the region previously denoted as ‘lateral pallium’ (ref. 2,59), we identified groups of cell types that are probably homologous to glutamatergic mammalian cortical neurons, supporting the hypothesis that the core cell types composing cortical/nuclear circuits across jawed vertebrates emerged in common vertebrate ancestors72,76. These neurons express genes that are associated with different projection modalities (for example, input, intratelencephalic or output) (Fig. 5a,b), but not in the same combinations as observed in jawed vertebrates77. Altogether, our observations indicate that only the evaginated (that is, lateral) portion of the lamprey telencephalon should be considered a bona fide pallium (Fig. 6b,c), which—in terms of cell type expression signatures—is homologous to all subdivisions (dorsal, ventral, lateral and medial) of the pallium of tetrapods. This suggests that the regional specification of gene expression patterns among pallial neurons evolved during gnathostome evolution in the lineage leading to tetrapods70,78. Future work may illuminate the timing and mechanisms underlying this regionalization.
Altogether, our study provides a global view of the cellular composition and molecular architecture of the ancestral vertebrate brain and provides the groundwork for investigating its extensive cellular and structural diversification during vertebrate evolution.
Methods
Sea lamprey samples
Sea lamprey (Petromyzon marinus) samples were dissected from specimens obtained from three different sources (Supplementary Table 1). The sampled animals were euthanized by submersion in 0.1% MS-222 (Sigma, A5040-25G), unless specified otherwise, followed by decapitation according to local guidelines. Tissue samples from larvae (that is, ammocoetes, between 90 and 130 mm in body length), juveniles (Youson stages 6–7) and adults used for bulk tissue RNA-seq and genome annotation (see below) were collected from freshwater streams in Maine, USA, and held in large, aerated tanks with sand and freshwater until being sacrificed. All procedures were approved by the University of Colorado, Boulder, Institutional Animal Care and Use Committee as described in protocol 2392. Larvae (between 70 and 120 mm in body length) used for scRNA-seq, smRNA-FISH and Cartana experiments were collected form the River Ulla in Galicia, Spain, and kept at the Interfaculty Biomedical Research Facility of Heidelberg University in freshwater aerated tanks with river sediment and appropriate temperature conditions (~15 °C) until used for tissue collection. All animal procedures were performed in accordance with European Union and German ethical guidelines on animal care and experimentation and were approved by the local animal welfare authorities (Regierungspräsidium Karlsruhe). Upstream migrating mature adults used for the scRNA-seq experiments were obtained from a commercial supplier (Novas Y Mar, Galicia, Spain) and were processed immediately upon their arrival at the laboratory. All procedures were approved by the Bioethics Committee of the University of Santiago de Compostela and the Xunta de Galicia Government and conformed to European Union and Spanish regulations for the care and handling of animals in research. Adult specimens used for the Cartana experiments were obtained from the US Fish and Wildlife Service and Department of the Interior and were euthanized by immersion in 0.25% MS-222, followed by decapitation. All procedures were approved by the California Institute of Technology Institutional Animal Care and Use Committee protocol 1436.
RNA extraction and sequencing of bulk tissue samples
In total, 63 sea lamprey tissue samples from six organs (brain, heart, liver, kidney, ovary and testis) were dissected from larval, juvenile and adult specimens. Total RNA was extracted using different extraction protocols (Supplementary Table 1); RNA quality was inspected using the Fragment Analyzer (Advanced Analytical Technologies), and its concentration was determined using a NanoDrop (Thermo Fisher Scientific). Strand-specific RNA-seq libraries were generated using the Illumina TruSeq Stranded mRNA Library protocol. Each library was sequenced on Illumina HiSeq 2500 platforms (100 nucleotides, single-end) at the Lausanne Genomic Technologies Facility (https://www.unil.ch/gtf).
Sea lamprey genome annotation
Bulk tissue RNA-seq reads were mapped to the sea lamprey germline genome10 using GSNAP79 (v.2018-03-01) with the option to find known and new splice junctions in individual reads activated (novelsplicing, 1). The resulting BAM files for each stage and tissue were merged before being used for transcriptome assembly with StringTie80 (v.1.3.4d). Each resulting GTF file was filtered for putative assembly artefacts using GffRead81 (v.0.9.9) by discarding single-exon transcripts and multi-exon mRNAs that have any intron with a non-canonical splice site consensus (that is, not GT-AG, GC-AG or AT-AC). Individual annotated transcriptomes were then merged together with the already available set of annotated protein-coding genes from the germline genome study10 to obtain a non-redundant set of transcripts. Genome annotation was further refined using TransDecoder (v.5.3.0; https://github.com/TransDecoder/TransDecoder) to identify candidate coding regions within the transcript sequences; this process involves identifying the longest putative open reading frame within each transcript and then searching the corresponding peptides against SwissProt (https://uniprot.org) using BlastP82 (v.2.5.0+) and Pfam (https://pfam.xfam.org) using HMMER83 (v.3.2). Annotation quality was assessed by comparing the number of reads mapping to exonic, intronic and intergenic regions of the genome (Extended Data Fig. 1). Annotation completeness was also estimated using BUSCO84 (v.3) by comparing the set of translated longest coding sequences from each transcript against a set of metazoan-conserved single-copy orthologues from OrthoDB85 (Supplementary Table 6).
Orthology assignment and gene nomenclature
Homology information for the set of annotated genes was retrieved by applying the OrthoFinder86 (v.2.3.11) pipeline against a group of selected chordates: vase tunicate (Ciona intestinalis)87, inshore hagfish (Eptatretus burgeri; permission to use unpublished genome data was given exclusively for the purposes of the present study; personal communication), Australian ghostshark (Callorhinchus milii)88, spotted gar (Lepisosteus oculatus)89, zebrafish (Danio rerio)90, West Indian Ocean coelacanth (Latimeria chalumnae)91, western clawed frog (Xenopus tropicalis)92, red junglefowl (Gallus gallus)93, house mouse (Mus musculus) and human (Homo sapiens). By reconstructing a complete set of rooted gene trees among the analysed species, this tool allows us to establish all orthology relationships among all genes and to infer duplication events and cross-reference them to the corresponding nodes on the gene and species trees. Proteomes were downloaded from Ensembl94 (remaining species; v.97) databases and used for a BlastP Best Reciprocal Hit analysis; to avoid redundancies in the blast results, only the peptides coming from the longest isoform within each gene were used. Rooted gene trees from the inferred orthogroups—that is, groups of genes descended from a single gene in the last common ancestor—were obtained using multiple sequence alignments (MAFFT95 v.7.455) with IQ-TREE96 (v.1.6.12; 1,000 bootstrap replicates) and STRIDE97. Orthology relationships can be explored in our online atlas. Throughout this work, we use mouse orthologue names to indicate lamprey gene names. This choice is justified by the fact that most lamprey genes lack a clear and consistent nomenclature and the fact that mouse is used as the main reference in our study. In cases where multiple mouse genes correspond to one lamprey gene (one-to-many relationships), we append all orthologue names, separated by slashes. We made an exception for SOX genes, where we used the well-established cyclostome annotation (SOXA, SOXB,…) and lamprey reference work.
Cell dissociation and scRNA-seq data generation
Larval and adult heads were air-dissected, and the brains were placed in 1× HBSS (Life Technologies, 14185052) for cleaning and removal of the meninges. Once cleaned, the brains were further treated as a whole sample or, for the second set of experiments, divided into regions (telencephalon, diencephalon, mesencephalon and rhombencephalon). Brain tissue was dissociated using the Papain Dissociation System (Worthington, LK003150), according to the manufacturer’s protocol, with the following modifications: the tissue was incubated in papain solution (volume adjusted for tissue size, 100–300 µl) at 28 °C for 15 min under constant agitation. Then, the tissue was gently triturated by pipetting up and down and collected by centrifugation for 1 min at 300 g. This step was followed by a second incubation in fresh papain solution and a final trituration, performed as described above. The dissociated cells were spun down at 300 g for 5 min and resuspended in the inhibitor solution (prepared following the Papain Dissociation System specifications). The suspension was filtered using a 40 µM falcon strainer (Sigma-Aldrich, CLS431750-50EA), and, immediately afterwards, a discontinuous density gradient was performed. The cells were then resuspended in Leibovitz’s L-15 Medium (Life Technologies, 21083027), reaching a final volume between 50 and 100 µl, depending on the original tissue size. The cells were examined for viability and counted using a trypan blue staining and a Neubauer counting chamber (Assistent).
After ensuring a cell viability greater than 90% and a concentration equal or higher than 300 cells per µl, cell suspensions (~15,000 cells per reaction) were loaded onto the Chromium system (10x Genomics). Complementary DNA amplification and scRNA-seq libraries were constructed using Single-Cell 3′ Gel Bead and Library v.2 (for larvae) and v.3 kits (for adults and larvae), following the instructions of the manufacturer. For three larval whole brains, we additionally produced libraries using the v.3 kit (SN580, SN582 and SN588) for adequate technical comparisons between the larval and adult datasets (main text and Extended Data Fig. 4b–d). Complementary DNA libraries were amplified using 12 or 13 PCR cycles and quantified on a Qubit Fluorometer (Thermo Fisher Scientific). Average fragment size was determined on a Fragment Analyzer (Agilent). The libraries were sequenced using the NextSeq 500/550 High Output Kit v.2.5 on the Illumina NextSeq 550 system (28 cycles for Read 1, 56 cycles for Read 2, 8 cycles for i7 index and 0 cycles for i5 index).
Single-cell RNA-seq data processing
The scRNA-seq reads were mapped to the reference genome10 with our extended annotation (see above), and unique molecular identifier (UMI) count matrices were produced using CellRanger v.3.0.2 (10x Genomics). Cell-containing droplets were obtained from the CellRanger calling algorithm and validated by checking (1) the cumulative distribution of UMIs, (2) the distribution of UMIs coming from mitochondrial genes and (3) the distribution of the proportion of UMIs coming from intronic regions. Putative multiplets (that is, droplets containing more than one cell) were identified using DoubletFinder98 and Scrublet99; droplets labelled as multiplets by either of the two methods were removed from the count matrices.
The obtained count matrices were analysed using Seurat v.3.1.5 (ref. 100) and pre-processed by keeping only genes expressed in at least five cells and by removing cells containing fewer than 200 UMIs and more than 5% (ammocoete) or 10% (adult) mitochondrial UMIs. The raw UMI counts were then normalized using the SCTransform method101, and the top 3,000 highly variable genes (HVGs) across all cells were used for subsequent analyses. Principal component analysis (PCA) was applied to the normalized HVG matrices, and the resulting 75 most significant PCs were used for building a shared nearest-neighbour graph that was then clustered using the Louvain method with different resolution values (0.5–10). Differential expression analysis was run to find potential marker genes from all clusters across all resolution values (Wilcoxon rank sum test: logFC ≥ 0.25; min.pct = 0.1; Bonferroni-adjusted P < 0.01). The PCA-transformed matrices were finally embedded in two-dimensional space using UMAP and t-distributed stochastic neighbour embedding (t-SNE) dimensionality reduction techniques.
The clustered cells were further manually inspected to identify and then remove spurious clusters (that is, clusters composed by damaged/stressed cells or multiplets/empty droplets that escaped the previous filtering steps). Cell types/states were annotated on top of the clusters obtained using the highest resolution value (10); a putative phenotype/function was assigned to each cluster by allocating marker genes to any of the following Gene Ontology102 categories: transcription (co-)factor, neurotransmitter metabolism, neurotransmitter transport, neurotransmitter receptor, neuropeptide103, neuropeptide receptor103, immune response, erythrocyte differentiation, blood vessel development, neurogenesis or gliogenesis. Annotated clusters that were contiguous on the UMAP and t-SNE embeddings were manually inspected and joined together if they were showing similar expression patterns among their respective marker genes. Additional functional information was added by comparing the annotated clusters to published vertebrate neural single-cell datasets18,104.
Datasets coming from different samples were integrated using integrative non-negative matrix factorization as implemented in LIGER v.0.5.0 (ref. 105). The datasets were integrated at two levels: (1) integration of replicates coming from the same brain region (that is, telencephalon, diencephalon, mesencephalon, rhombencephalon and whole brain) and stage (that is, ammocoete and adult), and (2) integration, within each stage, of all replicates together in the same dataset encompassing all sampled regions. Each integrated dataset was then imported to Seurat to perform shared nearest-neighbour graph construction, clustering, differential expression analysis, 2D embedding and cluster annotation as described above.
We noticed that the number of UMIs and expressed genes per cell was consistently lower for the larval dataset (produced using Chromium kit v.2) than for the adult one (produced using Chromium kit v.3) (Supplementary Tables 1 and 2). To establish whether this difference reflected an actual biological property of the two stages, we produced three larval datasets using the v.3 kit and compared their number of expressed genes per cell to the larval v.2 and adult v.3 datasets (Extended Data Fig. 4b) (see also ‘Cell dissociation and scRNA-seq data generation’). We also compared the distributions of cell-type-specific gene expression signals across datasets based on gene specificity indices calculated using the method developed by Tosches and colleagues64. Briefly, to obtain the specificity index with this method, the mean of normalized scRNA-seq read counts of each gene (gc) is calculated for each cell type (C) and then divided by its mean across all cells:
Lamprey–mouse comparisons
To find cross-vertebrate similarities and differences in neural cell types, the adult integrated brain atlas was compared against a published juvenile mouse nervous system atlas18. The two datasets were first compared via a correlation-based approach. That is, the raw UMI count matrices were extracted from both species datasets, and orthology information for the corresponding gene IDs was added; orthology relationships between mouse and lamprey were obtained from the OrthoFinder analysis (see above; Supplementary Table 7). The UMI counts coming from paralogues in the respective species were summed (‘meta-gene’ method20), and the species-specific gene IDs were replaced by numeric indices (1.n, where n is the number of orthology groups between the mouse and lamprey) shared by the two species. The new meta-gene count matrices were then normalized using SCTransform, filtered for HVGs and averaged across all annotated clusters. The expression levels were finally transformed to specificity indices (see above), which were then used for Pearson correlation analyses. Dendrograms relating cell-type families between lamprey and mouse were constructed using the pvclust106 R package with complete hierarchical clustering and 1,000 replicates.
In addition, the two datasets were compared using the SAMap (v.0.2.3) algorithm19, a method that enables mapping single-cell transcriptomic atlases between phylogenetically distant species. A gene–gene bipartite graph with cross-species edges connecting homologous gene pairs was constructed by performing reciprocal BlastP searches between the two proteomes of the two species. The graph was used in a second step to project the two datasets into a joint, lower-dimensional manifold representation, where the expression correlation between homologous genes was iteratively used to update the homology graph connecting the two atlases. After the analysis was run, a mapping score (ranging from 0 to 1) was computed among all possible cross-species cluster pairs. The full list of all lamprey gene names used in this study with their respective gene IDs is reported in Supplementary Table 8.
ISS
Whole brains (adults) and heads (larvae) were embedded in OCT mounting medium and then flash-frozen by laying them on isopentane, previously cooled on liquid nitrogen. Adult tissues were rinsed with ice-cold PBS before being frozen. The tissues were cryosectioned in 10 µm coronal and sagittal sections and stored at −80 °C until further use. Sections were processed for ISS using the High Sensitivity Library Preparation Kit from CARTANA AB (10x Genomics). The method and data processing are described by Ke and colleagues11. Processing of sections was done following CARTANA’s protocol with minor modifications. In brief, sections on SuperFrost Plus glass slides (Thermo Fisher Scientific) were air-dried for 5 min. Afterwards, the sections were fixed by 3.7% (v/v) paraformaldehyde in UltraPure distilled water (DNase/RNase-Free, Thermo Fisher Scientific, 10977035) for 7 min and washed in PBS (Thermo Fisher Scientific, 70011036; diluted in UltraPure distilled water), followed by 0.1 N HCl treatment for 5 min and a wash with PBS. The sections were then dehydrated with ethanol and air-dried before being covered with SecureSeal hybridization chambers (Grace Bio-Labs, 10910000). All subsequent steps, including probe hybridization and ligation, amplification, fluorescent labelling and quality control imaging, followed the manufacturer’s specifications. Finally, the mounted sections were shipped to CARTANA’s facility (Solna, Sweden) for ISS.
Single-molecule RNA-FISH
Larval whole heads were snap-frozen and cryosectioned (horizontal sections) as described above. This time, however, the sections were collected on coverslips (22 mm × 22 mm) previously pretreated with a silanization solution (0.3% (v/v) bind-silane (GE Healthcare Life Sciences, 17-1330-01), 0.1% (v/v) acetic acid and 99.6% (v/v) ethanol).
To reduce tissue autofluorescence, the sections were embedded in polyacrylamide (PA) gel, RNAs were anchored to the gel by LabelX treatment, and cellular proteins and lipids were cleared as previously described107,108, with modifications. LabelX solution was prepared by reacting Label-IT (Mirus Bio) with Acryloyl X, SE (Thermo Fischer Scientific), as described by Chen and colleagues108. Specifically, sections were air-dried for 15–20 min and fixed in 3.7% paraformaldehyde in PBS for 10–15 min, followed by a 2 min incubation in 4% SDS in PBS and washes with PBS. The fixed sections were then incubated in 70% ethanol at 4 °C for at least 16 h. Next, sections on coverslips were washed twice with PBS and once with 1× MOPS pH 7.7 (Sigma-Aldrich, M9381) and incubated with LabelX (diluted to a concentration of 0.006 mg ml−1 in 1× MOPS) at room temperature for 4 h, followed by two PBS washes. To anchor LabelX-modified RNAs, the sections were embedded in thin 4% PA gels. First, the coverslips were washed for 2 min with a PA solution, consisting of 4% (v/v) of 19:1 acrylamide/bis-acrylamide (Sigma-Aldrich, A9926-5), 60 mM Tris⋅HCl pH 8 and 0.3 M NaCl. The coverslips were then washed for 2 min with the PA solution supplemented with ammonium persulfate (Sigma-Aldrich, 7727-54-0) and TEMED (Sigma-Aldrich, T7024) at final concentrations of 0.03% (w/v) and 0.15% (v/v), respectively. To cast the gel, 75 µl of the PA solution (supplemented with the polymerizing agents) was added to glass slides previously treated with Repel Silane (GE Healthcare Life Sciences, 17-1332-01) and washed with ethanol. Each coverslip was then layered on top of a slide, with one drop of PA solution, ensuring that a thin PA layer formed between the slide and the coverslip. The gel was allowed to cast at room temperature for 1.5 h. The coverslips and slides were gently separated, leaving coverslips with sections embedded into the PA gel. The coverslips were then washed with digestion buffer consisting of 0.8 M guanidine-HCl, 50 mM Tris⋅HCl pH 8, 1 mM EDTA and 0.5% (v/v) Triton X-100. The coverslips were incubated with digestion buffer supplemented with 8 U ml−1 proteinase K (Sigma-Aldrich, P2308) at 37 °C for 2–3 h.
After background reduction, the sections were hybridized with HuluFISH probes, designed and developed by PixelBiotech. The hybridization protocol followed the manufacturer’s recommendations. Briefly, the coverslips were washed twice with HuluWash buffer (PixelBiotech GmbH) and incubated in 50 µl of probe solution, consisting of each probe diluted in hybridization buffer at a concentration of 1:100. The coverslips were incubated at 37 °C for 12 h, inside a light-protected humidified chamber. Afterwards, the coverslips were washed four times with HuluWash buffer. Each wash lasted 10 min and was done at room temperature. The last wash was supplemented with Hoechst 33342 (Thermo Fisher Scientific, H3569). The coverslips were then mounted in two drops of Prolong Diamond mounting medium (Thermo Fisher Scientific, P36961). The mounted sections were allowed to cure at room temperature for 24 hours.
All sections were imaged on a Leica TCS-SP5, a confocal laser scanning microscope controlled by the Leica Application Suite. All images shown are the projections of mosaics built by stitching individual z-stacks. Each z-stack consisted of individual images (50 images for SSPOa, VAT and GNRH1a; 15 images for ZFP704) taken by setting a range of 10–15 µm and a step size below 0.8 µm. The images were captured with a ×63 immersion oil objective and sequentially excited by a 405 nm diode laser (for the Hoechst 33342 staining), followed by the laser required for each probe (561 nm DPSS laser for SSPOa, ZFP704 and VAT; and 633 nm HeNe laser for GNRH1a). Projections of the z-stacks were performed in Fiji v.2 (ref. 109) by using the average intensity projection. Further processing (only when required) involved contrast enhancing (saturated pixels between 0.1% and 0.3%) and background subtraction for noise reduction (rolling ball with a radius of 50 pixels).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The raw and processed bulk and scRNA-seq data have been deposited to ArrayExpress with the accession numbers E-MTAB-11085 (bulk) and E-MTAB-11087 (single cell) (https://www.ebi.ac.uk/arrayexpress/). The genome annotation files and in situ images have been deposited to Zenodo110 (https://doi.org/10.5281/zenodo.5903844). Information about gene expression, cell type annotation and gene orthology relationships across species can be visualized using the online atlas (https://lampreybrain.kaessmannlab.org/).
Code availability
All code underlying the published atlas is available on GitHub (https://github.com/f-lamanna/LampreyBrainAtlas/) and Zenodo111 (https://doi.org/10.5281/zenodo.8113793) together with detailed instructions about its usage.
Change history
25 September 2023
A Correction to this paper has been published: https://doi.org/10.1038/s41559-023-02227-1
References
Sugahara, F., Murakami, Y., Pascual-Anaya, J. & Kuratani, S. Reconstructing the ancestral vertebrate brain. Dev. Growth Differ. 59, 163–174 (2017).
Nieuwenhuys, R. & Nicholson, C. in The Central Nervous System of Vertebrates (eds Nieuwenhuys, R. et al.) 397–495 (Springer, 1998).
Cao, C. et al. Comprehensive single-cell transcriptome lineages of a proto-vertebrate. Nature 571, 350–354 (2019).
Benito-Gutiérrez, È. et al. The dorsoanterior brain of adult amphioxus shares similarities in expression profile and neuronal composition with the vertebrate telencephalon. BMC Biol. 19, 110 (2021).
Albuixech-Crespo, B. et al. Molecular regionalization of the developing amphioxus neural tube challenges major partitions of the vertebrate brain. PLoS Biol. 15, e2001573 (2017).
Kumar, S., Stecher, G., Suleski, M. & Hedges, S. B. TimeTree: a resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. 34, 1812–1819 (2017).
Gans, C. & Northcutt, R. G. Neural crest and the origin of vertebrates: a new head. Science 220, 268–273 (1983).
Sugahara, F. et al. Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain. Nature 531, 97–100 (2016).
Grillner, S. Evolution of the vertebrate motor system—from forebrain to spinal cord. Curr. Opin. Neurobiol. 71, 11–18 (2021).
Smith, J. J. et al. The sea lamprey germline genome provides insights into programmed genome rearrangement and vertebrate evolution. Nat. Genet. 50, 270–277 (2018).
Ke, R. et al. In situ sequencing for RNA analysis in preserved tissue and cells. Nat. Methods 10, 857–860 (2013).
Miyashita, T., Gess, R. W., Tietjen, K. & Coates, M. I. Non-ammocoete larvae of Palaeozoic stem lampreys. Nature 591, 408–412 (2021).
Arendt, D. et al. The origin and evolution of cell types. Nat. Rev. Genet. 17, 744–757 (2016).
Hobert, O. Terminal selectors of neuronal identity. Curr. Top. Dev. Biol. 116, 455–475 (2016).
Martínez-de-la-Torre, M., Pombal, M. A. & Puelles, L. Distal-less-like protein distribution in the larval lamprey forebrain. Neuroscience 178, 270–284 (2011).
Kala, K. et al. Gata2 is a tissue-specific post-mitotic selector gene for midbrain GABAergic neurons. Development 136, 253–262 (2009).
Achim, K. et al. The role of Tal2 and Tal1 in the differentiation of midbrain GABAergic neuron precursors. Biol. Open 2, 990–997 (2013).
Zeisel, A. et al. Molecular architecture of the mouse nervous system. Cell 174, 999–1014.e22 (2018).
Tarashansky, A. J. et al. Mapping single-cell atlases throughout Metazoa unravels cell type evolution. eLife 10, e66747 (2021).
Geirsdottir, L. et al. Cross-species single-cell analysis reveals divergence of the primate microglia program. Cell 179, 1609–1622.e16 (2019).
Arendt, D., Bertucci, P. Y., Achim, K. & Musser, J. M. Evolution of neuronal types and families. Curr. Opin. Neurobiol. 56, 144–152 (2019).
Pamcer, Z. et al. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature 430, 174–180 (2004).
Nakao, T. Electron microscopic studies on the lamprey meninges. J. Comp. Neurol. 183, 429–453 (1979).
Vanlandewijck, M. et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature 554, 475–480 (2018).
Rovainen, C. M. Glucose production by lamprey meninges. Science 167, 889–890 (1970).
Lee, E. M. et al. Functional constraints on SoxE proteins in neural crest development: the importance of differential expression for evolution of protein activity. Dev. Biol. 418, 166–178 (2016).
Weil, M.-T. et al. Axonal ensheathment in the nervous system of lamprey: implications for the evolution of myelinating glia. J. Neurosci. 38, 6586–6596 (2018).
Bullock, T. H., Moore, J. K. & Fields, R. D. Evolution of myelin sheaths: both lamprey and hagfish lack myelin. Neurosci. Lett. 48, 145–148 (1984).
Joly, J.-S. et al. Windows of the brain: towards a developmental biology of circumventricular and other neurohemal organs. Semin. Cell Dev. Biol. 18, 512–524 (2007).
Bundgaard, M. & Van Deurs, B. Brain barrier systems in the lamprey. II. Ultrastructure and permeability of the choroid plexus. Brain Res. 240, 65–75 (1982).
Barreiro-Iglesias, A., Villar-Cerviño, V., Anadón, R. & Rodicio, M. C. A monoclonal antibody as a tool to study the subcommissural organ and Reissner’s fibre of the sea lamprey: an immunofluorescence study before and after a spinal cord transection. Neurosci. Lett. 464, 34–38 (2009).
Cserr, H. F., Bundgaard, M., Ashby, J. K. & Murray, M. On the anatomic relation of choroid plexus to brain: a comparative study. Am. J. Physiol. Regul. Integr. Comp. Physiol. 7, 76–81 (1980).
Becker, C. G. & Becker, T. Neuronal regeneration from ependymo-radial glial cells: cook, little pot, cook! Dev. Cell 32, 516–527 (2015).
Yuan, T., York, J. R. & McCauley, D. W. Gliogenesis in lampreys shares gene regulatory interactions with oligodendrocyte development in jawed vertebrates. Dev. Biol. 441, 176–190 (2018).
Wannier, T., Orlovsky, G. & Grillner, S. Reticulospinal neurones provide monosynaptic glycinergic inhibition of spinal neurones in lamprey. NeuroReport 6, 1597–1600 (1995).
Pombal, M. A., Marín, O. & Gonzlez, A. Distribution of choline acetyltransferase-immunoreactive structures in the lamprey brain. J. Comp. Neurol. 431, 105–126 (2001).
Kebschul, J. M. et al. Cerebellar nuclei evolved by repeatedly duplicating a conserved cell-type set. Science 370, eabd5059 (2020).
Lannoo, M. J. & Hawkes, R. A search for primitive Purkinje cells: zebrin II expression in sea lampreys (Petromyzon marinus). Neurosci. Lett. 237, 53–55 (1997).
Jalalvand, E., Robertson, B., Wallén, P. & Grillner, S. Ciliated neurons lining the central canal sense both fluid movement and pH through ASIC3. Nat. Commun. 7, 10002 (2016).
Oppel, A. Lehrbuch der Vergleichenden Mikroskopischen Anatomie der Wirbeltiere (Gustav Fischer, Jena, 1905).
Stephenson-Jones, M., Floros, O., Robertson, B. & Grillner, S. Evolutionary conservation of the habenular nuclei and their circuitry controlling the dopamine and 5-hydroxytryptophan (5-HT) systems. Proc. Natl Acad. Sci. USA 109, 164–173 (2012).
Grillner, S., von Twickel, A. & Robertson, B. The blueprint of the vertebrate forebrain—with special reference to the habenulae. Semin. Cell Dev. Biol. 78, 103–106 (2018).
Cole, W. C. & Youson, J. H. Morphology of the pineal complex of the anadromous sea lamprey, Petromyzon marinus L. Am. J. Anat. 165, 131–163 (1982).
Fischer, A. J., Foster, S., Scott, M. A. & Sherwood, P. Transient expression of LIM-domain transcription factors is coincident with delayed maturation of photoreceptors in the chicken retina. J. Comp. Neurol. 506, 584–603 (2008).
Lu, Y. et al. Single-cell analysis of human retina identifies evolutionarily conserved and species-specific mechanisms controlling development. Dev. Cell 53, 473–491.e9 (2020).
Mullen, R. D. et al. Roles of the LHX3 and LHX4 LIM-homeodomain factors in pituitary development. Mol. Cell. Endocrinol. 265–266, 190–195 (2007).
Yáñez, J., Pombal, M. A. & Anadón, R. Afferent and efferent connections of the parapineal organ in lampreys: a tract tracing and immunocytochemical study. J. Comp. Neurol. 403, 171–189 (1999).
Fernández-López, B., Sobrido-Cameán, D., Anadón, R., Rodicio, M. C. & Barreiro-Iglesias, A. Restricted co-localization of glutamate and dopamine in neurons of the adult sea lamprey brain. J. Anat. 231, 776–784 (2017).
von Twickel, A. et al. Individual dopaminergic neurons of lamprey SNc/VTA project to both the striatum and optic tectum but restrict co-release of glutamate to striatum only. Curr. Biol. 29, 677–685.e6 (2019).
Brodin, L. et al. Neurotensin‐like peptides in the CNS of lampreys: chromatographic characterization and immunohistochemical localization with reference to aminergic markers. Eur. J. Neurosci. 2, 1095–1109 (1990).
St.-Gelais, F., Jomphe, C. & Trudeau, L. É. The role of neurotensin in central nervous system pathophysiology: what is the evidence? J. Psychiatry Neurosci. 31, 229–245 (2006).
Pistocchi, A. et al. Crucial role of zebrafish prox1 in hypothalamic catecholaminergic neurons development. BMC Dev. Biol. 8, 27 (2008).
Sobrido-Cameán, D. et al. Cholecystokinin in the central nervous system of the sea lamprey Petromyzon marinus: precursor identification and neuroanatomical relationships with other neuronal signalling systems. Brain Struct. Funct. 225, 249–284 (2020).
Sobrido-Cameán, D. et al. Galanin in an agnathan: precursor identification and localisation of expression in the brain of the sea lamprey Petromyzon marinus. Front. Neuroanat. 13, 83 (2019).
Sobrido-Cameán, D. et al. Differential expression of somatostatin genes in the central nervous system of the sea lamprey. Brain Struct. Funct. 226, 1031–1052 (2021).
Barreiro-Iglesias, A., Anadón, R. & Rodicio, M. C. New insights on the neuropeptide Y system in the larval lamprey brain: neuropeptide Y immunoreactive neurons, descending spinal projections and comparison with tyrosine hydroxylase and GABA immunoreactivities. Neuroscience 167, 396–413 (2010).
Chen, Y. C. et al. Transcriptional regulator PRDM12 is essential for human pain perception. Nat. Genet. 47, 803–808 (2015).
Thélie, A. et al. Prdm12 specifies V1 interneurons through cross-repressive interactions with Dbx1 and Nkx6 genes in Xenopus. Development 142, 3416–3428 (2015).
Pombal, M. A., Alvarez-Otero, R., Pérez-Fernández, J., Solveira, C. & Megías, M. Development and organization of the lamprey telencephalon with special reference to the GABAergic system. Front. Neuroanat. 5, 20 (2011).
Stenman, J., Toresson, H. & Campbell, K. Identification of two distinct progenitor populations in the lateral ganglionic eminence: implications for striatal and olfactory bulb neurogenesis. J. Neurosci. 23, 167–174 (2003).
Pombal, M. A., El Manira, A. & Grillner, S. Organization of the lamprey striatum—transmitters and projections. Brain Res. 766, 249–254 (1997).
Stephenson-Jones, M., Samuelsson, E., Ericsson, J., Robertson, B. & Grillner, S. Evolutionary conservation of the basal ganglia as a common vertebrate mechanism for action selection. Curr. Biol. 21, 1081–1091 (2011).
García-López, M. et al. Histogenetic compartments of the mouse centromedial and extended amygdala based on gene expression patterns during development. J. Comp. Neurol. 506, 46–74 (2008).
Tosches, M. A. et al. Evolution of pallium, hippocampus, and cortical cell types revealed by single-cell transcriptomics in reptiles. Science 360, 881–888 (2018).
Tosches, M. A. & Laurent, G. Evolution of neuronal identity in the cerebral cortex. Curr. Opin. Neurobiol. 56, 199–208 (2019).
Wei, X. et al. Single-cell Stereo-seq reveals induced progenitor cells involved in axolotl brain regeneration. Science 377, eabp9444 (2022).
Villar-Cerviño, V., Barreiro-Iglesias, A., Mazan, S., Rodicio, M. C. & Anadón, R. Glutamatergic neuronal populations in the forebrain of the sea lamprey, Petromyzon marinus: an in situ hybridization and immunocytochemical study. J. Comp. Neurol. 519, 1712–1735 (2011).
Suryanarayana, S. M., Pérez-Fernández, J., Robertson, B. & Grillner, S. Olfaction in lamprey pallium revisited—dual projections of mitral and tufted cells. Cell Rep. 34, 108596 (2021).
Suryanarayana, S. M., Robertson, B., Wallén, P. & Grillner, S. The lamprey pallium provides a blueprint of the mammalian layered cortex. Curr. Biol. 27, 3264–3277.e5 (2017).
Woych, J. et al. Cell-type profiling in salamanders identifies innovations in vertebrate forebrain evolution. Science 377, eabp9186 (2022).
Ocaña, F. M. et al. The lamprey pallium provides a blueprint of the mammalian motor projections from cortex. Curr. Biol. 25, 413–423 (2015).
Suryanarayana, S. M., Pérez-Fernández, J., Robertson, B. & Grillner, S. The evolutionary origin of visual and somatosensory representation in the vertebrate pallium. Nat. Ecol. Evol. 4, 639–651 (2020).
Sugahara, F., Pascual-Anaya, J., Kuraku, S., Kuratani, S. & Murakami, Y. Genetic mechanism for the cyclostome cerebellar neurons reveals early evolution of the vertebrate cerebellum. Front. Cell Dev. Biol. 9, 700860 (2021).
Pombal, M. A., Megías, M., Bardet, S. M. & Puelles, L. New and old thoughts on the segmental organization of the forebrain in lampreys. Brain Behav. Evol. 74, 7–19 (2009).
Rodriguez-Romaguera, J. et al. Prepronociceptin-expressing neurons in the extended amygdala encode and promote rapid arousal responses to motivationally salient stimuli. Cell Rep. 33, 108362 (2020).
Briscoe, S. D. & Ragsdale, C. W. Evolution of the chordate telencephalon. Curr. Biol. 29, R647–R662 (2019).
Briscoe, S. D. & Ragsdale, C. W. Homology, neocortex, and the evolution of developmental mechanisms. Science 362, 190–193 (2018).
Striedter, G. F. & Northcutt, R. G. The independent evolution of dorsal pallia in multiple vertebrate lineages. Brain Behav. Evol. 96, 200–211 (2022).
Wu, T. D. & Nacu, S. Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics 26, 873–881 (2010).
Pertea, M. et al. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290–295 (2015).
Pertea, G. & Pertea, M. GFF Utilities: GffRead and GffCompare. F1000Research 9, 304 (2020).
Camacho, C. et al. BLAST+: architecture and applications. BMC Bioinform. 10, 421 (2009).
Eddy, S. R. Accelerated profile HMM searches. PLoS Comput. Biol. 7, e1002195 (2011).
Simão, F. A., Waterhouse, R. M., Ioannidis, P. & Kriventseva, E. V. BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 19, 3210–3212 (2015).
Zdobnov, E. M. et al. OrthoDB v9.1: cataloging evolutionary and functional annotations for animal, fungal, plant, archaeal, bacterial and viral orthologs. Nucleic Acids Res. 45, D744–D749 (2017).
Emms, D. M. & Kelly, S. OrthoFinder: phylogenetic orthology inference for comparative genomics. Genome Biol. 20, 238 (2019).
Satou, Y. et al. Improved genome assembly and evidence-based global gene model set for the chordate Ciona intestinalis: new insight into intron and operon populations. Genome Biol. 9, R152 (2008).
Venkatesh, B. et al. Elephant shark genome provides unique insights into gnathostome evolution. Nature 505, 174–179 (2014).
Braasch, I. et al. The spotted gar genome illuminates vertebrate evolution and facilitates human–teleost comparisons. Nat. Genet. 48, 427–437 (2016).
Howe, K. et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 496, 498–503 (2013).
Amemiya, C. T. et al. The African coelacanth genome provides insights into tetrapod evolution. Nature 496, 311–316 (2013).
Hellsten, U. et al. The genome of the western clawed frog Xenopus tropicalis. Science 328, 633–636 (2010).
Hillier, L. W. et al. Sequencing and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 432, 695–716 (2004).
Yates, A. D. et al. Ensembl 2020. Nucleic Acids Res. 48, D682–D688 (2020).
Katoh, K. & Standley, D. M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780 (2013).
Nguyen, L. T., Schmidt, H. A., Von Haeseler, A. & Minh, B. Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274 (2015).
Emms, D. M. & Kelly, S. STRIDE: species tree root inference from gene duplication events. Mol. Biol. Evol. 34, 3267–3278 (2017).
McGinnis, C. S., Murrow, L. M. & Gartner, Z. J. DoubletFinder: doublet detection in single-cell RNA sequencing data using artificial nearest neighbors. Cell Syst. 8, 352484 (2018).
Wolock, S. L., Lopez, R. & Klein, A. M. Scrublet: computational identification of cell doublets in single-cell transcriptomic data. Cell Syst. 8, 281–291.e9 (2019).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902.e21 (2019).
Hafemeister, C. & Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296 (2019).
The Gene Ontology Consortium. Gene Ontology: tool for the unification of biology. Nat. Genet. 25, 25–29 (2000).
Wang, Y. et al. NeuroPep: a comprehensive resource of neuropeptides. Database 2015, bav038 (2015).
Raj, B. et al. Simultaneous single-cell profiling of lineages and cell types in the vertebrate brain. Nat. Biotechnol. 36, 442–450 (2018).
Welch, J. D. et al. Single-cell multi-omic integration compares and contrasts features of brain cell identity. Cell 177, 1873–1887.e17 (2019).
Suzuki, R. & Shimodaira, H. Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 22, 1540–1542 (2006).
Moffitt, J. R. et al. High-performance multiplexed fluorescence in situ hybridization in culture and tissue with matrix imprinting and clearing. Proc. Natl Acad. Sci. USA 113, 14456–14461 (2016).
Cheng, Y. S. et al. Autonomous combinatorial color barcoding for multiplexing single molecule RNA visualization. Preprint at bioRxiv https://doi.org/10.1101/127373 (2017).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Lamanna, F. et al. Genome annotation files and in situ images associated with analyses of the lamprey neural cell type atlas. Zenodo https://doi.org/10.5281/zenodo.5903844 (2023).
Lamanna, F. et al. All code underlying the analyses of the lamprey neural cell type atlas. Zenodo https://doi.org/10.5281/zenodo.8113793 (2023).
Acknowledgements
We thank all members of the Kaessmann group for fruitful discussions; Robert Frömel for his help with library sequencing; E. Panzariello, M. Sanchez-Delgado and N. Trost for the brain and animal illustrations; M. Mall for providing temporary lab space; and M. Cardoso-Moreira for discussions and comments on the manuscript. The computations were performed on the Kaessmann lab server (managed by N. Trost) and the bwForCluster from the Heidelberg University Computational Center (supported by the state of Baden-Württemberg through bwHPC and the German Research Foundation, INST 35/1134-1 FUGG). We thank W. Wang of the Northwestern Polytechnical University of Xi’an, China; Y. Zhang of the Institute of Zoology, Chinese Academy of Sciences; and J. Pascual-Anaya from the University of Málaga, Spain, for granting access to the early draft of the inshore hagfish genome (https://doi.org/10.1101/2023.04.08.536076). This work was supported by grants from the European Research Council (no. 615253, OntoTransEvol), the European Commission (Marie Skłodowska-Curie Actions ITN: EvoCELL) and the Tschira foundation, which funded the Illumina NextSeq machine used for sequencing. D.M.M. and D.J. were supported by grants from the National Science Foundation (IOS 1656843), the National Institutes of Health/National Institute of Dental and Craniofacial Research (RDE025940) and University of Colorado, Boulder RIO Innovative Seed Grant FY21 (all to D.M.M.). D.J. was also supported by a grant from the Scientific Grant Agency of the Slovak Republic (VEGA 1/0450/21). This project has received funding from the European Research Council under the European Union’s Horizon 2020 research and innovation programme (VerteBrain to H.K., grant agreement no. 101019268; and NeuralCellTypeEvo to D.A., grant agreement no. 788921).
Author information
Authors and Affiliations
Contributions
F.L., F.H.-S. and H.K. conceived and organized the study on the basis of H.K.’s original design. F.L., F.H.-S. and H.K. wrote the manuscript with input from all authors. F.L. performed all analyses and developed the brain atlas app. F.H.-S. established and optimized the tissue dissociation protocol and performed all scRNA-seq and in situ experiments with support from A.P.O., J. Schmidt and C.S. and guidance from M.S. F.L. and F.H.-S. annotated and interpreted the data. T.B. prepared the bulk libraries with guidance from K.M. M.S. established the smRNA-FISH protocol. F.H.-S., A.P.O., D.J., D.S.-C., G.N.S.-D., M.L.M., J. Stundl and S.A.G. collected the samples. A.B.-I., D.M.M., M.E.B. and M.C.R. provided the samples. J.J.S. provided early access to genome assemblies and annotations. A.P.O., M.S., F.M., D.S.-C., A.B.-I., D.M.M. and D.A. provided useful feedback and discussions. H.K. supervised the study and provided funding.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Maria Tosches and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Assessment of lamprey genome annotation quality.
Barplots comparing the proportion of reads mapping to exonic, intronic, and intergenic regions between the lamprey genome annotation produced in this study (re-annotated) and the published annotations of lamprey10, chicken, mouse, and human.
Extended Data Fig. 2 Larval (ammocoete) brain dataset.
a, Schematic of the sea lamprey larval brain showing the different regions dissected for this study and UMAP projections for each brain region. Each UMAP projection represents an integration of the biological replicates (as indicated in panel b) for each brain region, respectively. Additional information is available in the interactive atlas (https://lampreybrain.kaessmannlab.org/ammocoete.html). b, Barplots showing the proportions of each cell type group (as reported in a and Fig. 1b, c) for each sample. For whole brain samples, proportions are showed separately for datasets obtained using the v2 and v3 kits. c, Heatmap showing clustered pseudobulk brain regions based on Spearman’s ρ. Median ρ = 0.90.
Extended Data Fig. 3 Adult brain dataset.
a, Schematic of the sea lamprey adult brain showing the different regions dissected in this study and UMAP projections for each brain region. Each UMAP projection represents an integration of the biological replicates (as indicated in panel b) for each brain region, respectively. Additional information available in the interactive atlas (https://lampreybrain.kaessmannlab.org/adult.html). b, Barplots showing the proportions of each cell type group (as reported in a and Fig. 1b, c) for each sample. For whole brain samples, the proportions of neural cell types only are additionally showed. c, Heatmap showing clustered pseudobulk brain regions based on Spearman’s ρ. Median ρ = 0.97.
Extended Data Fig. 4 Differences between larval and adult datasets and between v2 and v3 kits.
a, UMAP of 13,301, 22,950, and 10,557 cells from the larval (v2; left), larval (v3; middle), and adult (right) whole brain datasets, respectively. Dashed lines highlight neuronal cells. b, Distribution of the number of expressed genes per cell for each whole brain sample. c, Distributions of specificity index scores for whole brains (neurons only). d, Same as b with each replicate downsampled to the same number of cells (N = 1,000). Statistical significance between groups in b, c, and d calculated with two-sided Mann-Whitney U test (P = 0.000013). Boxplot annotation: bounds of box, Q1 (25th percentile), Q3 (75th percentile), Q3 - Q1 = IQR (interquartile range); centre, median; minimum, (Q1 – 1.5 * IQR); maximum (Q3 – 1.5 * IQR).
Extended Data Fig. 5 Posterior forebrain, midbrain, and hindbrain neurons.
a, Subtree from the dendrogram of Fig. 1c displaying the expression of selected marker genes for each cell type (gene names mentioned in main text are highlighted in bold). SC, spinal cord. b, Sagittal section (anterior end to the left) of a larval head (white dashed line outlines the brain) showing the expression of SLC6A5, HOXA/B5, and PKD2L1. Dashed lines separate the main brain regions. c, UMAP projection of a larval hindbrain dataset showing the expression of CHAT and TBX6/20. d-m Sagittal sections (anterior end to the left) of the adult brain showing the expression of selected marker genes. Arrows in d and e indicate the putative location of cranial nerve nuclei. See Supplementary Fig. 2 for the ISS section schemes; scale bars, 500 μm.
Extended Data Fig. 6 SAMap scores for all cell type groups.
a, b, Heatmaps of SAMap mapping scores for all groups of non-neuronal and neuronal cell types between mouse and lamprey, including all genes (a) and TF genes only (b).
Extended Data Fig. 7 Correlations between cell type groups.
a, b, Heatmaps showing Pearson’s correlation coefficients of specificity indexes of lamprey and mouse cell type groups for all orthologous genes (a) and for TFs only (b).
Extended Data Fig. 8 Blood, vascular, and PNS cells.
a, Subtree from the dendrogram of Fig. 1c displaying the expression of selected marker genes for each cell type (gene names mentioned in main text are highlighted in bold). b, Expression of marker genes for immune (GRN), meningeal (PDGFRA, PAH), and vascular (KDR) cells. c, Expression of lamprey markers and their mouse orthologs in the respective brain atlases (UMAPs). d, Coronal section of the larval telencephalon showing the spatial expression of the genes shown in b (same color code). Arrows mark leptomeningeal layers. e, Coronal section of the adult isthmic region (mesencephalon/rhombencephalon) showing the expression of SOXE1 and SOXE2 within cranial nerve roots (white arrows). See Supplementary Fig. 2 for ISS section schemes; scale bars, 500 μm.
Extended Data Fig. 9 Ependymoglial cells.
a, Subtree from the dendrogram of Fig. 1c displaying the expression of selected marker genes for each cell type (gene names mentioned in main text are highlighted in bold). b, Expression of OTX2 within the adult telencephalon. White arrows point to choroid plexus. c, Expression of KERA within the adult brain (sagittal section; anterior end to the left), showing its concentration in the hindbrain. d, Violin plot displaying KERA expression among ependymoglial cell types; color code as in a. e, f, Expression of FOXJ1 (e) and CCN1-5 (f) within the adult telencephalon. g, Horizontal section of the larval brain (anterior end to the upper left corner) showing the expression of SSPO (smFISH) around the rostral end of the third ventricle (sub-commissural organ). h, Expression of STMN4 (neurons) and HES5 (astrocytes) within the adult telencephalon showing the periventricular localization of lamprey astrocytes. i, Sagittal section (anterior end to the left) of a larval head (brain enclosed within white dashed line) showing the expression of CCN1-5 and OTX2. White arrows point to choroid plexuses. j, k, Expression of TNR (j), HEPACAM (k), and their mouse orthologs on the respective brain atlases. See Supplementary Fig. 2 for ISS section schemes; scale bars, 500 μm.
Extended Data Fig. 10 Anterior forebrain and epithalamic neurons.
a, Subtree from the dendrogram of Fig. 1c displaying the expression of marker genes for each cell type (gene names mentioned in main text are highlighted in bold). b, Section scheme of the adult brain. c-t, ISS maps of selected neuronal marker genes across various adult brain coronal sections (shown in b). u, Section scheme of the larval brain. v-w, ISS maps of selected neuronal marker genes across various larval brain coronal sections (shown in u). DHy, dorsal hypothalamus; DMTN, dorsomedial telencephalic nucleus; LH, left habenula; MH, medial habenula; MPO, medial preoptic nucleus; OB, olfactory bulb; OT, optic tectum; Pal, pallium; Pald, pallidum; PEA, pallial extended amygdala; PCN, postoptic commissure nucleus; PO, pineal organ; PpO, parapineal organ; PpT, parapineal tract; PS, pineal stalk; PTh, pre-thalamus; PThE, pre-thalamic eminence; PTN, posterior tubercle nucleus; RH, right habenula; RPa, rostral paraventricular area; Sp, septum; St, striatum; Th, thalamus; VHy, ventral hypothalamus. Scale bars, 500 μm.
Supplementary information
Supplementary Information
Supplementary Figs. 1 and 2.
Supplementary Tables
Supplementary Tables 1–8.
Supplementary Data 1
Lamprey genome custom annotation files.
Supplementary Data 2
In situ images produced in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lamanna, F., Hervas-Sotomayor, F., Oel, A.P. et al. A lamprey neural cell type atlas illuminates the origins of the vertebrate brain. Nat Ecol Evol 7, 1714–1728 (2023). https://doi.org/10.1038/s41559-023-02170-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41559-023-02170-1
- Springer Nature Limited
This article is cited by
-
Exciting times for evolutionary biology
Nature Ecology & Evolution (2024)
-
A cell type atlas of the lamprey brain
Nature Ecology & Evolution (2023)