
Abstract
There is a paucity of genetic characterization in people with Parkinson’s disease (PD) of Latino and Afro-Caribbean descent. Screening LRRK2 and GBA variants in 32 New Yorkers of Puerto Rican ethnicity with PD and in 119 non-Hispanic-non-Jewish European PD cases revealed that Puerto Rican participants were more likely to harbor the LRRK2-p.G2019S variant (15.6% vs. 4.2%, respectively). Additionally, whole exome sequencing of twelve Puerto Rican and Dominican PD participants was performed as an exploratory study.
Similar content being viewed by others


With the advent of personalized medicine emerging in the neurology landscape, genetic testing is playing a greater role in counseling individual patients with Parkinson’s disease (PD). LRRK2 and GBA are the most prevalent genetic risk factors for PD1,2. Mutations in both LRRK2 and GBA occur worldwide, and founder effects have been described in some populations3,4. LRRK2 and GBA variant carriers with PD can be enrolled in neuroprotection trials5,6; knowledge of genetic status is required to determine trial eligibility7,8. Thus, study recruitment may be biased toward better studied populations with known higher mutation frequency9,10. Despite suggestion that LRRK2 variants may be increased in Puerto Ricans10,11, mutation rates in individuals with self-reported Puerto Rican ethnicity10,12,13 have not been widely studied.
Understanding the frequencies of potential variants as genetic risk in different populations is especially important as genetic testing shifts to whole -exome and whole -genome sequencing (WES and WGS) whose interpretation rests on these data. However, the majority of exomes in population databases, such as the Genome Aggregation Database (gnomAD v2, https://gnomad.broadinstitute.org/), are from mixed European Caucasians (45% of the >125,000 exomes in gnomAD) while Latino and African origin populations are still underrepresented.
We analyzed the frequencies of the LRRK2-p.G2019S and specific GBA variants in a group of PD participants with self-reported Puerto Rican ethnicity, and also carried out an exploratory WES analysis in a subgroup of Puerto Rican participants as well as a limited number of Dominican PD participants.
Among 32 PD participants with self-reported Puerto Rican ethnicity, and 119 with non-Hispanic, non-Jewish European ancestry, those from Puerto Rico were more likely to harbor the LRRK2-p.G2019S variant than Europeans [Table 1A; 15.6% vs. 4.2%) (OR 4.22 (95% CI: 1.14–15.6)].
Fourteen European participants carried a GBA variant while no Puerto Rican cases had an isolated GBA variant; one carried dual LRRK2-GBA variants (Table 1A).
Additionally, because so few Latino PD patients have been examined for known PD candidate genes at present, as a pilot study, 8 patients of Puerto Rican ethnicity from the initial screening and 4 further patients of Dominican Republic (DR) ethnicity with early age of onset and/or positive family history (FH) underwent WES (as the most readily available tool for detecting potentially relevant coding variants) and standard variant discovery analyses (Table 1B). A search for rare (gnomAD overall allele frequency [AF] <0.01) deleterious variants in 74 disease-associated genes (Supplementary Table 2) identified variants in 8 genes (Table 2), including one in TREM2 (rs2234253) in patients 2 and 5 (Table 1B) both of DR ethnicity with tremor predominant PD; and one in CTSB (rs28605689) in patient 6 (Table 1B) with postural instability and gait disorder (PIGD). Neither of these variants were reported in ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/).
To detect potential risk alleles for PD in the Puerto Rican/Dominican populations, variant AF in our sample was compared to AF in control populations (gnomAD overall, Latino, African, (Non-Finnish) European (NFE)) for variants in the candidate gene list. While this analysis did not reveal a significant enrichment in PD of any single variant, we noted a nominally increased frequency of rs2230570 (EIF4G1) as compared to the Latino population in gnomAD v2 (Supplementary Table 1; p-value 0.008; adjusted p-value 0.052). This variant showed a similar trend in the NFE population (p-value 0.023; adjusted p-value 0.131) but was not significant when compared to the African population (Supplementary Table 1). Although this variant is predicted to be benign, it could be in linkage disequilibrium with some other functional variant in non-coding region not assessed by WES.
Evaluating loss-of-function variants exome-wide, we discovered a novel stop-gain mutation in exon 10 (of 13) of the COBL gene (p.Glu777*) in patient 1 (Table 1B) who is affected with mild parkinsonism and positive FH. Follow-up analysis revealed a rare deleterious missense variant (rs202117145, Table 2) in the same gene in patient 8 (Table 1B) with gait onset and rest tremor. Neither variant is reported in ClinVar.
Studies on the genetics and clinical spectrum of PD in non-European, especially Latino and Afro-Caribbean, populations are limited and mainly focusing on one or a few PD- associated genes14,15. Such studies are sorely needed, as the demand for direct-to-consumer testing supports the patients´ interest in determining the genetic contribution to their PD. There is also emerging evidence that awareness of genotype could guide therapeutic decisions7,8. Trials for some genes have emerged and multiple are in progress, thus far focused on LRRK2 and GBA variants16,17.
We have previously reported an excess of the LRRK2-p.G2019S variant10 in Puerto Ricans, however, the numbers of screened samples have been relatively small. Here, the increased frequency of LRRK2-p.G2019S in our New York Puerto Rican population was confirmed (15.6%), suggesting that testing in individuals of Puerto Rican ethnicity who are potentially eligible for trials should be considered as it will improve access and equity. However, Nuytemans et al. reported that only 2/37 individuals (5.4%) of Puerto Rican ethnicity ascertained from Manati and Miami harbored p.G2019S mutations13. Local ancestry analysis showed that these two patients had likely European ancestry, suggesting that p.G2019S was introduced to the Latino population through their European ancestor. Further studies of larger cohorts are thus still needed to obtain better frequency estimates. Lower frequencies of p.G2019 were also observed in the PD populations of Argentina (3.2%), Brazil (1.4%), Colombia (1.5%), Ecuador (1.2%), Peru (0.2%), and Uruguay (4.2%)18 and this raises important questions about why even in the Nuytemans study, the rates were slightly increased among Puerto Ricans and whether this is attributable to a greater frequency in the European than the Amerindian ancestry. As the Puerto Rican population represents an admixture of native Taino, African and Hispanic European, thus ancestry informative markers might further help guide such data. Admixtures of European, African, and Amerindian ancestries have similarly been reported in the Puerto Rican PD patients from Miami and Manatí13.
We carried out principal component analysis (PCA) for ancestry related markers for the 12 patients who were included in the WES section. This analysis showed that these patients have admixed genetic background with contributions from European, American (Hispanic) and African global populations (Supplementary Fig. 1). Whether the Puerto Rican ethnicity contains higher proportion of European (non-Ashkenazi) ancestry remains to be determined.
As other genetic variants may be contributing to PD in the Puerto Rican population, and with the advent of WES and WGS, it is important to extend beyond LRRK2-p.G2019S and more frequently assessed GBA variants, and to identify variants that may be overrepresented in understudied populations. In 2021 three studies reported the genetic variants in the Latino population13,19,20, including that of Nuytemans et al. that identified novel LRRK2 and GBA variants in Latino PD patients from the Caribbean13.
Our exploratory WES analysis identified four genes that carried variants with elevated AF in our patient group. COBL is a constrained gene (LOEUF 0.47, Table 2) related to actin cytoskeletal organization during neuronal morphogenesis and its expression is enriched in brain21. The role of EIF4G1 in the PD-related neurodegeneration is still debated22. TREM2, in contrast, has been confirmed to represent a candidate gene for PD susceptibility and progression, and soluble TREM2 (sTREM2) expression in cerebrospinal fluid was increased in PD patients, suggesting sTREM2 in CSF as a substitute immune biomarker for PD neuron injury23. CTSB belongs to the lysosomal proteases, and associations between the lysosomal pathway and PD pathogenesis has been described broadly. Lack of CTSB was shown to impair lysosomal trafficking during neural development24. However, because of our limited sample size, these results should be treated as a starting point for further studies in a much larger set of Latino PD cases.
While our study highlights the urgency to study genetics of these and other underrepresented populations, our WES included a small number of patients, therefore, we did not have statistical power to identify unique clinical differences or major clinical trends. Additional studies are needed, that not only evaluate known variants, but increase the WES/WGS data to identify population-specific risk variants in not only Puerto Rican, and Dominican, but also other Afro-Caribbean populations. This work will be aided by the Latin American Research Consortium on the Genetics of Parkinson’s Disease (LARGE-PD, Mata PI).
Methods
Participants
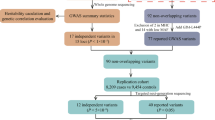
Evaluation of the frequency of LRRK2-p.G2019S and GBA variants was carried out in 151 participants in total: 32 with self-reported Puerto Rican ethnicity, and 119 with non-Hispanic-non-Jewish European ancestry from MSBI, SUNY Downstate and James J. Peters Veterans Affairs Medical Center (Bronx). Subsequently, 8 participants of Puerto Rican and 4 of Dominican ethnicity who were negative in the LRRK2 or GBA testing, and who had higher likelihood of genetic etiology because of younger age of onset and/or positive FH, were selected for WES. We chose 57 years as cut-off for age of onset as this was the median age in our overall sample, and included patients with any FH, but did not limit to first-degree relatives. One Puerto Rican participant was subsequently excluded from the AF analyses due to discovery of biallelic PRKN deletions on an independent clinical testing including MLPA.
All participants met criteria for PD and signed informed consents from their respective institutions. The study was approved by and conforms with all Ethical Regulations of the Mount Sinai Internal Review Board and SUNY Downstate Internal Review Board.
Genotyping for GBA variants
Genomic DNA (50–80 ng) was used to PCR amplify 5 fragments suitable for Sanger sequencing. Fragments were designed to include 10 common GBA variants (84GG, IVS2 + 1, p.E326K, p.T369M, p.N370S, p.V394L, p.D409G, p.L444P, p.A456P, and p.R496H). Primer sequences for PCR fragments were chosen to avoid amplification of the GBA pseudogene and are available upon request. Following PCR amplification, fragments were cleaned enzymatically with ExoSAP-IT (Affymetrix, Santa Clara, CA) according to manufacturer instructions. Samples were sequenced using ABI BigDye Terminator chemistry (v1.1) and visualized on an ABI 3730xl DNA analyzer. ABI sequencer files for each sample were compared to GBA consensus sequence to identify variants using Mutation Surveyor software (SoftGenetics, State College, PA). In addition to DNA sequencing, a PCR amplification assay was designed to assess presence of the RecNciI recombinant allele (primers and conditions available upon request).
Evaluation of LRRK2 and GBA variant frequencies
The LRRK2-p.G2019S and 11 variants in GBA (84GG, IVS2 + 1, p.E326K, p.T369M, p.N370S, p.V394L, p.D409G, p.L444P, p.A456P, RecNciI, and p.R496H) were analysed as described previously25,26 and Supplementary Methods]. Logistic regression models were used to evaluate the association between the self-reported ethnicity and the LRRK2-p.G2019S variant status, and the GBA variant status, respectively (STATA16, Texas). Two models were run: (i) with and (ii) without including “age of onset” as a co-variate. No difference in results were observed when adjusting vs not adjusting for this co-variate (data not shown). T-tests and Mann–Whitney tests were used to assess group differences.
Whole exome sequencing
We performed WES on 12 samples (8 PR and 4 DR) to detect functional rare variants in 74 candidate genes (see Supplementary Methods for details on WES, Supplementary Table 2 for candidate genes), chosen based on a literature review for genes previously associated with PD and other movement disorders with phenotypic overlap including frontotemporal dementia (FTD), dystonia, essential tremor (ET), chorea-acanthocytosis (ChAc), McLeod Syndrome (MLS), Huntington disease-like 2 (HDL2), neurodegeneration with brain iron accumulation disorders (NBIA) and Alzheimer’s disease (AD). Additionally, we searched for loss-of-function mutations exome-wide. To test for the enrichment of more common variants in candidate genes in the PD population, Fisher’s tests were performed using allele counts in our WES cohort vs. published allele counts in a population database (gnomad v2).
WES was performed using the Agilent SureSelect Human All Exon V4 + UTR (71 Mb) library and an Illumina HiSeq 2000 with a paired-end module. Burrows–Wheeler Aligner (bwa-mem)27 was used to align reads to the reference sequence GRCh37/hg19, after which quality control, duplicate read removal, base quality recalibration, and variant calling was performed using Haplotypecaller in Genome Analysis ToolKit (GATK) version 3.328. The mean per-base coverage for called variants ranged from 25-34X across samples. Single nucleotide variants and small in/dels were annotated with in silico predictions using the Variant Effect Predictor module from Ensembl (http://useast.ensembl.org/info/docs/tools/vep/index.html?redirect=no), in particular, the CADD (https://cadd.gs.washington.edu/), SIFT (https://sift.bii.a-star.edu.sg/), and Polyphen (http://genetics.bwh.harvard.edu/pph2/) scores, and via MACIE scoring29, (protein score > 0.80), which was eventually used to include variants of interest based on an estimate of the probability of a functional variant at the locus.
Stop and splice-acceptor/donor mutations detected in canonical transcripts genome-wide were filtered for quality using LOFTEE (Loss of Function Transcript Estimator) scores from the VEP (Variant Effect Predictor) plugin of Ensembl. As such, based on criteria set by the program, variants that are near the end of the transcript are filtered out30.
In testing for the enrichment of variants in the PD population, Fisher’s tests were limited to alleles found in >3 out of 11 individuals in our cohort, and those that had a higher variant alle frequency in PD (“risk alleles”). p-values were corrected for multiple testing using the false discovery rate method.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The IRBs of the participating institutions do not allow direct access to datasets used and/or analysed during the current study. Datasets are available directly from from Laurie Ozelius (laurie.ozelius@mgh.harvard.edu) or from Rachel Saunders-Pullman (Rachel.Saunders-Pullman@mountsinai.org) upon request with approval of a data use agreement.
References
Cook Shukla, L., et al. Parkinson Disease Overview. 2004 May 25 [Updated 2019 Jul 25]. in GeneReviews® [Internet] (eds Adam, M. P., Ardinger, H. H., Pagon, R. A., et al.) (University of Washington, 1993–2021).
Ortega, R. A. et al. Association of dual LRRK2 G2019S and GBA variations with Parkinson disease progression. JAMA Netw. Open 4, e215845 (2021).
Nishioka, K. et al. Glucocerebrosidase mutations are not a common risk factor for Parkinson disease in North Africa. Neurosci. Lett. 477, 57–60 (2010).
Ruiz-Martínez, J. et al. Penetrance in Parkinson’s disease related to the LRRK2 R1441G mutation in the Basque Country (Spain). Mov. Disord. 25, 2340–2345 (2010).
Cruz-Vicente, P., Passarinha, L. A., Silvestre, S. & Gallardo, E. Recent developments in new therapeutic agents against Alzheimer and Parkinson diseases: in-silico approaches. Molecules. 26, 2193 (2021).
Vuletić, V., Rački, V., Papić, E. & Peterlin, B. A systematic review of Parkinson’s disease pharmacogenomics: is there time for translation into the clinics? Int. J. Mol. Sci. 22, 7213 (2021).
Garofalo, A. W. et al. Selective inhibitors of G2019S-LRRK2 kinase activity. J. Med. Chem. 63, 14821–14839 (2020).
Kelly, K. & West, A. B. Pharmacodynamic biomarkers for emerging LRRK2 therapeutics. Front. Neurosci. 14, 807 (2020).
Vinagre-Aragón, A. et al. A more homogeneous phenotype in Parkinson’s disease related to R1441G mutation in the LRRK2 gene. Front. Neurol. 12, 635396 (2021).
Saunders-Pullman, R. et al. LRRK2 G2019S mutations may be increased in Puerto Ricans. Mov. Disord. 26, 1772–1773 (2011).
Alcalay, R. N. et al. Frequency of known mutations in early-onset Parkinson disease: implication for genetic counseling: the consortium on risk for early onset Parkinson disease study. Arch Neurol 67, 1116–1122 (2010).
Markopoulou, K. et al. Longitudinal monitoring of Parkinson’s disease in different ethnic cohorts: the DodoNA and LONG-PD study. Front. Neurol. 11, 548 (2020).
Nuytemans, K. et al. Novel variants in LRRK2 and GBA identified in Latino Parkinson disease cohort enriched for Caribbean origin. Front. Neurol. 11, 573733 (2020).
Marder, K. S. et al. Predictors of parkin mutations in early-onset Parkinson disease: the consortium on risk for early-onset Parkinson disease study. Arch. Neurol. 67, 731–738 (2010).
Duque, A. F. et al. Analysis of the LRRK2 p.G2019S mutation in Colombian Parkinson’s disease patients. Colomb. Med. 46, 117–121 (2015).
Jennings, D. et al. LRRK2 inhibition by BIIB122 in healthy participants and patients with Parkinson’s disease. Mov. Disord. 38, 386–398 (2023).
Vollstedt, E. J. et al. MJFF global genetic Parkinson’s disease study group. Embracing monogenic Parkinson’s disease: the MJFF global genetic PD cohort. Mov. Disord. 38, 286–303 (2023).
Cornejo-Olivas, M. et al. Variable frequency of LRRK2 variants in the Latin American research consortium on the genetics of Parkinson’s disease (LARGE-PD), a case of ancestry. NPJ Parkinsons Dis. 3, 19 (2017).
Torrealba-Acosta, G. et al. Clinical and genetic analysis of Costa Rican patients with Parkinson’s disease. Front. Neurol. 12, 656342 (2021).
Loesch, D. P. et al. Latin American Research Consortium on the genetics of Parkinson’s disease (LARGE-PD). Characterizing the genetic architecture of Parkinson’s disease in Latinos. Ann. Neurol. 90, 353–365 (2021).
Ahuja, R. et al. Cordon-bleu is an actin nucleation factor and controls neuronal morphology. Cell. 131, 337–350 (2007).
Deng, H., Wu, Y. & Jankovic, J. The EIF4G1 gene and Parkinson’s disease. Acta Neurol. Scand 132, 73–78 (2015).
Peng, G. et al. Analysis of cerebrospinal fluid soluble TREM2 and polymorphisms in sporadic Parkinson’s disease in a Chinese population. J. Mol. Neurosci. 70, 294–301 (2020).
Jiang, M. et al. Cathepsin B inhibition blocks neurite outgrowth in cultured neurons by regulating lysosomal trafficking and remodeling. J. Neurochem. 155, 300–312 (2020).
Nichols, W. C. et al. Parkinson study group-PROGENI investigators. Genetic screening for a single common LRRK2 mutation in familial Parkinson’s disease. Lancet 365, 410–412 (2005).
Moran, E. E. et al. Cognitive functioning of glucocerebrosidase (GBA) non-manifesting carriers. Front. Neurol. 12, 635958 (2021).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
McKenna, A. et al. The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303 (2010).
Li, X. et al. A multi-dimensional integrative scoring framework for predicting functional variants in the human genome. Am. J. Hum. Genet. 109, 446–456 (2022).
Karczewski, K. J. Genome Aggregation Database Consortium et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 581, 434–443 (2020).
Acknowledgements
We would like to thank participants and family members for generously contributing their time and effort to this study. Côté Clinical Genetics Initiative Award from the Parkinson’s Foundation, Empire Clinical Research Investigator Program, National Institute of Neurological Disorders and Stroke (NINDS) (U01NS107016-01A1 and U01NS094148-01), and the Bigglesworth Family Foundation.
Author information
Authors and Affiliations
Contributions
G.M.M. drafted and revised the manuscript for intellectual content; analyzed and interpreted the data. R.O., A.D., D.R., and L.O. had a major role in the acquisition of data; analyzed and interpreted the data, revised the manuscript. R.Y. and A.N. interpreted the data. V.K., N.U., M.S., V.S., J.M., R.H.W., S.B., and J.C. had a major role in the acquisition of data and revised the manuscript. R.S.-P. designed and conceptualized study, drafted and revised the manuscript for intellectual content; had a major role in the acquisition of data; analyzed and interpreted the data.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Miltenberger-Miltenyi, G., Ortega, R.A., Domingo, A. et al. Genetic risk variants in New Yorkers of Puerto Rican and Dominican Republic heritage with Parkinson’s disease. npj Parkinsons Dis. 9, 160 (2023). https://doi.org/10.1038/s41531-023-00599-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41531-023-00599-6
- Springer Nature Limited