

Abstract
The three-dimensional genome structure organized by CTCF is required for development. Clinically identified mutations in CTCF have been linked to adverse developmental outcomes. Nevertheless, the underlying mechanism remains elusive. In this investigation, we explore the regulatory roles of a clinically relevant R567W point mutation, located within the 11th zinc finger of CTCF, by introducing this mutation into both murine models and human embryonic stem cell-derived cortical organoid models. Mice with homozygous CTCFR567W mutation exhibit growth impediments, resulting in postnatal mortality, and deviations in brain, heart, and lung development at the pathological and single-cell transcriptome levels. This mutation induces premature stem-like cell exhaustion, accelerates the maturation of GABAergic neurons, and disrupts neurodevelopmental and synaptic pathways. Additionally, it specifically hinders CTCF binding to peripheral motifs upstream to the core consensus site, causing alterations in local chromatin structure and gene expression, particularly at the clustered protocadherin locus. Comparative analysis using human cortical organoids mirrors the consequences induced by this mutation. In summary, this study elucidates the influence of the CTCFR567W mutation on human neurodevelopmental disorders, paving the way for potential therapeutic interventions.
Similar content being viewed by others

Introduction
There is a growing body of evidence indicating a close association between the disruption of chromatin’s three-dimensional structure and the onset and progression of various diseases, including developmental disorders and cancers1,2,3,4,5. Playing a crucial role as the master weaver of the genome6, CCCTC-binding factor (CTCF) actively participates in the regulation of Topologically Associating Domains (TADs) and chromatin loops7,8,9. Alterations in the expression level or insufficient dosage of CTCF at the cellular level can result in shifts in cell fate, potentially leading to cellular demise7,10,11,12. Notably, maternal depletion of Ctcf halts early embryonic development13, elevating zygotic lethality14. In excitatory neurons, a conditional knockout of Ctcf induces neuronal cell death and gliosis15.
Moreover, Ctcf proves indispensable for early forebrain development, contributing to a delicate balance between neural progenitor cell proliferation and differentiation16. Recent discoveries in clinical and targeted sequencing studies have unveiled cases of heterozygous CTCF mutations, encompassing deletions, frameshift, and missense mutations. These variations are highly correlated with potential symptoms subsumed under the classification of intellectual developmental disorder, autosomal dominant 21 (OMIM 615502), which can range from global developmental delay, intellectual disability, and short stature to autistic behaviors and symptoms resembling congenital heart disease17,18,19,20,21. While mouse models utilizing tissue-specific Ctcf conditional knockout have been explored to mirror the pertinent phenotypic traits16,22,23,24,25, the impact of germline-origin Ctcf point mutations on individual development and the underlying mechanisms in human diseases remain unclear.
Building on the observation that a clinically heterozygous CTCF mutation at c.1699 C > T (p.Arg567 > Trp) induces severe phenotypes, including intellectual disability, microcephaly, hypotonia, growth deficiency, delayed development, short stature, delayed bone age, and feeding difficulties17,19,20,26,27, we establish a Ctcf mutant mouse model and a human embryonic stem cell (hESC)-derived cortical organoid model harboring the CTCFR567W mutation to explore its mechanistic involvement. In the mouse model, homozygous CTCFR567W-mutant offspring experience early postnatal mortality, while their heterozygous counterparts exhibit a survival rate comparable to wild-type mice, displaying only a lean phenotype during the juvenile stage. Additionally, at embryonic-day-18.5 (E18.5), mice with homozygous CTCFR567W mutation manifest abnormal neurological and cardiopulmonary development.
Specifically, within the mouse cortex, the CTCFR567W mutation accelerates the depletion of stem-like cells, hastens the maturation of GABAergic neurons, and induces irregularities in both neurodevelopmental and synaptic pathways. The CTCFR567W mutation leads to a reduction in CTCF binding to its upstream motifs, causing a partial reorganization of chromatin interactions, and subsequently influencing gene expression, particularly within the clustered protocadherin (cPcdh) locus. Consistent with the mouse model, the hESC-derived cortical organoid model reveals that the homozygous CTCFR567W mutation hinders self-organization during differentiation, while the heterozygous mutation causes an imbalance in the differentiation of stem-like cells and the maturation of neurons. This imbalance, induced by the CTCFR567W mutation, impacts pathways crucial for neural development and affects genes implicated in neurodevelopmental disorders.
Results
Construction and phenotypic characterization of CTCFR567W-mutant mice
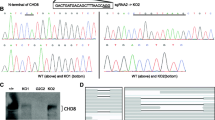
The R567 residue of CTCF, a conserved residue across most known orthologues (Fig. 1a), is located proximal to the splice donor site in exon 9. Notably, this residue, situated within the 11th zinc finger (ZF) domain of the CTCF protein (Fig. 1b), was predicted to specifically contact DNA17,28. To investigate the impact of the CTCFR567W mutation on developmental phenotypes, we generated a mouse model harboring the CTCFR567W mutation using CRISPR/Cas9-mediated genome editing (Fig. 1b). Through multiple rounds of breeding and genotyping, we observed that heterozygous mutant mice (Ctcf+/R567W) exhibited a birth state similar to that of wild-type mice (Ctcf+/+), while homozygous mutant mice (CtcfR567W/R567W) experienced respiratory distress and succumbed within 30 minutes (min) after birth (Fig. 1c and Supplementary Fig. 1a).

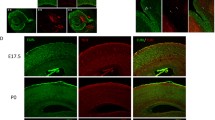
a The amino acid (aa) conservation of CTCF around R567 across species. CTCF ZF11 residues annotated in UniProt are marked with a green line, DNA-binding residues (residues 565–568) with a blue line, and Zn2+ coordinating residues with a yellow band. R567 is highlighted in red. b CRISPR/Cas9 knock-in of the CTCFR567W mutation diagram. The sgRNA targeting sequence (Ctcf exon 9) and the mutation site R567W (R, Arg, CGG; W, Trp, TGG) are underlined. The mutated base and the PAM sequence NGG are in red. The aa sequence around the mutation site is highlighted with a pink box. c Representative photograph and Sanger sequencing results of neonatal mice. Scale bar, 10 mm. The mutation base C > T is highlighted in blue. d Box and whisker plots showing the body weight (95% confidence interval) of E18.5 mice (Ctcf+/+: n = 19, Ctcf+/R567W: n = 22, CtcfR567W/R567W: n = 17). e Two-week-old Ctcf+/+ and Ctcf+/R567W mice images. f Weight curves of postnatal day 9 (P9), P16, P23, and P30 Ctcf+/+ and Ctcf+/R567W mice (Ctcf+/+: n = 44, Ctcf+/R567W: n = 39). g Box and whisker plots showing the body weight (95% confidence interval) of P16 mice for different sexes of Ctcf+/+ and Ctcf+/R567W mice (Ctcf+/+: M, n = 33, F, n = 25, Ctcf+/R567W: M, n = 27, F, n = 32; M: male, F: female). h, i Sagittal brain and heart paraffin H&E staining images of Ctcf+/+, Ctcf+/R567W, and CtcfR567W/R567W mice at E18.5. Enlarged views of the cortex, hippocampus, and left ventricular wall are shown on the right. Scale bars, 200 μm. Quantitative data are presented as mean ± SD. p-values were obtained by one-way ANOVA with Dunnett’s multiple comparisons test adjustment (d), two-way ANOVA with Šídák’s multiple comparisons test adjustment (f), and two-tailed unpaired t-test (g) are indicated in the graphs. Experiments in (h, i) were repeated independently three times with similar results. All Box and whisker plots show lower and upper quartiles (box limits), median (center line) and minimum to maximum values (whiskers). Source data are provided as a Source Data file.
Upon analyzing the genotyping results of numerous embryos at E18.5, we noted that most CtcfR567W/R567W mice could developed into intact individuals, albeit considerably smaller than their wild-type counterparts (0.815 ± 0.107 g vs 1.157 ± 0.095 g, p = 1 × 10−15; Fig. 1d). Additionally, based on a 1-month recording, Ctcf+/R567W mice demonstrated a relatively thin physique with no discernible difference in body weight by sex (Fig. 1e–g), consistent with the lean phenotype reported in previous studies on Ctcf hemizygous mice9 and clinical cases of CTCF mutation11,17,19,29. The lean phenotype in Ctcf+/R567W mice might be attributed to reduced levels of growth hormone (GH), as indicated by enzyme-linked immunosorbent assay (ELISA) (16.14 ± 3.78 mg/mL vs 13.1 ± 2.38 mg/mL, p = 0.043) and quantitative RT-PCR (RT-qPCR) (Supplementary Fig. 1b, c). Despite this, the lifespan of Ctcf+/R567W mice was comparable to that of Ctcf+/+ mice (Supplementary Fig. 1d). Furthermore, we noted a statistically significant increase in Ctcf expression in neuron and lung tissues, as revealed by RT-qPCR, due to the CTCFR567W mutation (Supplementary Fig. 1e). However, this mutation did not significantly alter CTCF splicing or distribution across tissues, as assessed by Western blot and immunohistochemistry (Supplementary Fig. 1f, g).
In investigating pathological phenotypes following Ctcf mutation, we focused on the brain, heart, and lung tissues due to the birth lethal phenotype observed in CtcfR567W/R567W mice and the prevalence of neurodevelopmental disorders in clinical populations. Our observations revealed normalcy in all three tissues of both Ctcf+/+ and Ctcf+/R567W mice, with no apparent pathological changes (Fig. 1h, i and Supplementary Fig. 1h and 2a–c). However, CtcfR567W/R567W mice exhibited abnormal histopathology in their hearts, characterized by a decreased proportion of ventricles, resembling the noncompaction cardiomyopathy phenotype, including a thin ventricular wall, noncompaction of the ventricular myocardium, and trabecular hyperplasia (Fig. 1i). Additionally, we observed a thickened alveolar interstitium and dense, noninflated alveoli in the lungs of E18.5 and postnatal day 0 (P0) CtcfR567W/R567W mice (Supplementary Fig. 1h). While these pathological changes could potentially contribute to respiratory distress, they might also represent secondary effects of the animals’ demise rather than direct consequences of Ctcf homozygous mutation.
To investigate whether the Ctcf+/R567W-induced phenotype in mice could replicate clinical neurodevelopmental disorders and autism-like traits, we conducted a series of behavioral experiments, including the open-field test (OFT), novel object recognition (NOR) test, three-chamber test, elevated plus maze (EPM) test, morris water maze test, and rotarod test. No significant abnormalities were observed in Ctcf+/R567W adult mice compared with wild-type mice (Supplementary Fig. 2d–q), except for a lower resistance to fatigue in the rotarod test (Supplementary Fig. 2r). These findings suggest that, unlike Ctcf hemizygous mice which exhibit loss of Ctcf expression in the brain30, Ctcf+/R567W mice maintain mostly normal brain function, given that Ctcf expression is preserved. Discrepancies between mice and humans may be attributed to the complexities of the human nervous system. In summary, our results indicate that the homozygous CTCFR567W mutation leads to birth lethality and developmental delay in mice, somewhat mirroring known clinical phenotypes manifested as overall developmental disorders.
CTCFR567W homozygous mutation hampers neural development and electrical activity
To explore the potential impact of the CTCFR567W mutation on neurodevelopment, we conducted follow-up experiments using E18.5 fetal mouse brain tissues and primary neurons. As the phenotypes of Ctcf+/R567W mice were generally consistent with those of Ctcf+/+ mice according to preliminary observations and tests, therefore, we focused on examining the effects of the homozygous mutation on neural development and activity in subsequent studies.
We initially labeled cultured primary neurons with an anti-TUJ1 antibody to evaluate neurite complexity. Both primary neurons and stem cell-derived neural clusters from CtcfR567W/R567W mice exhibited significantly less complex neuronal protrusions compared to cells from Ctcf+/+ mice (Fig. 2a and Supplementary Fig. 3a). Despite no differences in soma size across genotypes, morphological analysis revealed fewer primary neurites in neurons from CtcfR567W/R567W mice (4.524 ± 1.504 vs 5.864 ± 1.552, p = 0.0127; Fig. 2b). Total neurite length was also significantly decreased in CtcfR567W/R567W mice (171.2 ± 59.38 vs 339.3 ± 109.9 μm, p = 8.4 × 10−7; Fig. 2b), indicating that the Ctcf homozygous mutation results in neurite abnormalities.

a TUJ1 (green) immunostaining and Sholl analysis of primary neurons cultured for 5 days from E18.5 mouse forebrains (n = 3 for each genotype). Nuclei were stained with DAPI (blue). Scale bars, 50 μm. b Quantification of soma sizes (left), the number of primary neurites (middle) and total neurite length (right) of primary neurons from (a) (n = 3 for each genotype, the n of neurons counted are indicated in the graph). c The raster image of MEA experiments showing electrical activity of cultured neurons from three genotypes over a time period (60 s) on day 14. Black and blue bars represent spikes and bursts, respectively. The top of the raster plot shows the spike histogram. d–h Quantification of MEA metrics of Ctcf+/+ and CtcfR567W/R567W cultured neurons. Statistics of electrical activity for 15 min, including the number of spikes per min (d) the number of bursts per min (e) the number of network bursts (f) the mean firing rate (g) and the synchrony index (h) are provided (Ctcf+/+: n = 4, CtcfR567W/R567W: n = 2; the n of cultured wells recorded are indicated in the graph). i Golgi staining of brain slices at the cortex (top) or thalamus (bottom) from Ctcf+/+ and CtcfR567W/R567W mice at E18.5. Scale bars, 100 μm. Areas of interest are indicated by red rectangles. j, k Calcium influx after depolarization from in vitro-cultured cortical neurons on day 10 for Ctcf+/+ and CtcfR567W/R567W mice (n = 3 for each genotype) (RFU: relative fluorescence units). Data from the Sholl analysis are presented as mean ± SEM, and other quantitative data are presented as mean ± SD. p-values by two-way ANOVA with repeated measures (a), one-way ANOVA with Dunnett’s multiple comparisons test adjustment (b), and two-tailed unpaired t-test (d–h and k) are indicated in the graphs. Experiments were repeated three times independently with similar results. All Box and whisker plots show lower and upper quartiles (box limits), median (center line) and minimum to maximum values (whiskers), representing 95% confidence intervals. Source data are provided as a Source Data file.
We employed microelectrode array (MEA) to assess E18.5 primary neurons and explore whether the CTCFR567W mutation affects neuroelectric activity and neural network formation. While all three genotypes exhibited spontaneous firing activity, CtcfR567W/R567W neurons displayed significantly lower neural network activity, evident through reduced numbers of spikes, bursts and network bursts, along with decreases in mean firing rate and synchronization index (Fig. 2c–h and Supplementary Fig. 3b–e). No significant variation in these metrics was observed between Ctcf+/+ and Ctcf+/R567W neurons (Fig. 2c and Supplementary Fig. 3f–i).
Golgi staining further revealed fewer stained neurons and neurites in the thalamus and cortex of CtcfR567W/R567W mice compared to Ctcf+/+ mice (Fig. 2i). Calcium (Ca2+) imaging experiments also demonstrated significantly attenuated Ca2+ signals in cultured cortical and hippocampal neurons of the CtcfR567W/R567W group (Fig. 2j, k and Supplementary Fig. 3j, k). In conclusion, our results suggest that the Ctcf homozygous mutation at R567 impairs neural activity and network function.
CTCFR567W mutation induces premature depletion of progenitor cells and accelerates the maturation of GABAergic neurons
CTCF plays a crucial role in transcriptional regulation31,32,33. To assess the impact of the homozygous CTCFR567W mutation on gene expression in brain tissue, we conducted RNA sequencing (RNA-seq) on whole brains from E18.5 fetal mice. Principal component analysis (PCA) revealed a subtle deviation in CtcfR567W/R567W samples from Ctcf+/+ and Ctcf+/R567W samples, suggesting a certain degree of heterogeneity among the samples (Supplementary Fig. 4a). Differentially expressed gene (DEG) analysis showed minimal differences between Ctcf+/+ and Ctcf+/R567W mice, while a limited number of DEGs were identified in CtcfR567W/R567W mice (Supplementary Fig. 4b). Considering the diverse cell types in the brain, we isolated neurons from E18.5 fetal brain cortices, cultured them in vitro, and conducted RNA-seq experiments. The results indicated a downregulation of more DEGs in neurons, most of which were consistent with the RNA-seq data from the whole brain (Fig. 3a, b). Notably, these co-downregulated genes were enriched in interferon pathways and cPcdh-mediated cell adhesion molecules (Fig. 3c). The cPcdh gene clusters, consisting of three tandem gene clusters (Pcdhα, Pcdhβ, and Pcdhγ), are associated with neuronal identity, synaptic organization, axonal guidance, and neural circuit assembly34,35,36.

a, b Scatter plot showing DEGs detected by RNA-seq in whole brains and primary cultured neurons of E18.5 Ctcf+/+ and CtcfR567W/R567W mice. Significantly upregulated genes are shown in red, and significantly downregulated genes are shown in blue (a). Co-downregulated genes are shown in blue (b). c Network showing the GO analysis results of co-downregulated genes in (b). d UMAP visualization of snRNA-seq data and cell clustering results of cortex samples from E18.5 Ctcf+/+ and CtcfR567W/R567W mice. e Proportion of different cell types in the cortex of Ctcf+/+ and CtcfR567W/R567W mice. f Number of DEGs for each cell type in cortex samples of E18.5 Ctcf+/+ and CtcfR567W/R567W mice from snRNA-seq data. g GO analysis of DEGs for indicated cell clusters from cortex snRNA-seq in CtcfR567W/R567W mice compared to Ctcf+/+ mice. GO analysis was carried out by clusterProfiler software with two-sided hypergeometric test. h UMAP visualization of snRNA-seq data and cell clustering results of heart samples from E18.5 Ctcf+/+ and CtcfR567W/R567W mice. Plots are colored by cell type. mCM, mature cardiomyocytes; dCM, developing cardiomyocytes; EC, endothelial cells. i GO network of DEGs for 7 myocardial-related cell types (CM1, CM2, dCM1, dCM2, dCM3, dCM4, epicardium). Sizes of the circles represent the level of enrichment of the GO term. A greener color indicates more cell types enriched in that term. j Average expression levels of indicated gene sets in different cell types of heart snRNA-seq data. k Correlation heatmap of DEGs from the indicated cell types based on the Jaccard score.
To assess the impact of the CTCFR567W mutation on neurodevelopment at the single-cell level, we performed single-nucleus RNA seq (snRNA-seq) on brain cortices from E18.5 Ctcf+/+ and CtcfR567W/R567W mice. Following quality control and data preprocessing, we obtained a total of 13,137 and 15,661 cells from wild-type and mutant samples, respectively. Utilizing the ScType platform for annotation37, cluster analysis identified nine cell populations, including radial glial cells, neural progenitor cells, neuroblasts, immature neurons, glutamatergic neurons, GABAergic neurons, oligodendrocyte precursor cells, microglia, and endothelial cells (Fig. 3d and Supplementary Fig. 4c). Our data revealed that the CTCFR567W mutation had no significant effect on the cellular phenotype or landscape of the mouse cortex but led to noticeable shifts in specific cohorts. Specifically, the mutant mice exhibited a reduction in the proportion of stem-like neural progenitor cells and radial glial cells, as well as immature neurons, coupled with an increase in postmitotic neuroblasts and inhibitory GABAergic neurons (Fig. 3e). Additionally, a marginal decrease in excitatory glutamatergic neurons was observed (Fig. 3e).
The DEGs identified through functional annotation of main stem/progenitor cells and neuronal clusters revealed that the CTCFR567W mutation impacts crucial pathways guiding neural development, synaptic structuring, and cell-cell adhesion. However, the major pathways affected were not consistently observed across different cell subgroups (Fig. 3f, g). Notably, GABAergic neurons exhibited enriched differential pathways associated with locomotory behavior, cognition, and synaptic transmission. In contrast, glutamatergic neurons displayed enrichment in pathways related to the regulation of membrane potential, neuron migration, and learning processes (Fig. 3g). Furthermore, an overlap was observed between DEGs and risk genes identified in genome-wide association studies (GWAS) of autism spectrum disorder (ASD) (Supplementary Fig. 4d). Additionally, there was a downregulation of Pcdhα and Pcdhβ genes and an upward trend in the expression of Pcdhγ genes, demonstrating consistent expression changes across all subgroups (Supplementary Fig. 4e). Collectively, the CTCFR567W mutation disrupts mouse cortical neurodevelopment, leading to premature depletion of the progenitor pool and accelerated maturation of GABAergic neurons. These findings provide partial elucidation of the phenotypic manifestations observed in previous neuronal experiments (Fig. 2), establishing a tangible link to intellectual disabilities and autism-like phenotypes in clinical developmental disorders associated with CTCF mutation.
CTCFR567W mutation has little effect on cell type proportions but leads to noticeable DEGs in E18.5 heart and lung tissues
Further investigation into the effect of the CTCFR567W mutation on gene expression in E18.5 heart and lung tissues, guided by snRNA-seq and focusing on cardiopulmonary pathological phenotypes in CtcfR567W/R567W mice (Fig. 1i and Supplementary Fig. 1h), revealed distinct patterns. We classified 11 clusters, with 7 associated with cardiomyocytes (CMs), for cardiac tissue (Fig. 3h and Supplementary Fig. 5a), and 14 clusters for lung tissue cell types (Supplementary Fig. 5b, c). The mutation did not significantly alter the proportions of most cell types (Supplementary Fig. 5d, e). DEG analysis showed that CTCFR567W resulted in a limited number of DEGs per cell type (Supplementary Fig. 5f, g). Notably, in cardiac cells, genes linked to energy metabolism and myocardial contraction were significantly downregulated in CtcfR567W/R567W mice, while genes associated with muscle tissue development were upregulated (Fig. 3i, j). In the lungs, irregular expression of genes related to vascular structures and cell morphogenesis was observed, with DEGs varying across all clusters (Supplementary Fig. 5h, i). These gene pathways might correlate with the previously detected abnormal heart and lung morphology phenotypes and could contribute to potential respiratory failure at birth in these mutant mice (Fig. 1i and Supplementary Fig. 1h). Joint analysis of DEGs across heart, brain, and lung tissues indicated that the DEGs induced by CTCFR567W were specific to both tissues and cell types (Fig. 3k).
CTCFR567W weakens CTCF binding to sites with upstream motifs
To explore the impact of the CTCFR567W mutation on the manifestation of the associated disease phenotype through alterations in specific CTCF binding, we scrutinized the chromatin binding characteristics of CTCFR567W. This investigation is crucial, considering that CTCF exerts its biological functions by binding to specific DNA motifs via its 11 ZFs. Specifically, ZFs 4–7 engage the M1/core (C) motif, which represents the core consensus, and ZFs 9–11 engage the M2/upstream (U) motif, corresponding to peripheral motifs upstream to the core consensus site28,38,39,40. The crystal structure of CTCF ZFs (6–11) in complex with its DNA motif (PDB: 5YEL) has been resolved, and it is established that amino acids 565–568 within ZF11 of CTCF are suggested to form hydrogen bonds with DNA, corresponding to the CTCF U motif (Fig. 4a, b)28. Structural modeling indicates that the R567W mutation causes a shift of the ZF away from the DNA’s phosphate backbone, thereby disrupting the critical hydrogen bond between R567 and the DNA backbone. Furthermore, the W567 residue may impede the formation of the essential hydrogen bond between the neighboring R566 residue and DNA bases, further compromising ZF11 binding to the U motif (Fig. 4b, c).

a Crystal structure of the CTCF ZFs6–11-gb7CSE (CTCF-binding conserved sequence element within Pcdhγb7 promoter) complex obtained from PDB: 5YEL28 showing Zn2+ ions in cyan, DNA in gray, ZFs6–11 in green, and the R567 residue in red. Corresponding DNA sequences are shown at the bottom. b Molecular dynamic simulations of the CTCF-DNA complex. Wild-type and R567W mutant structures with R567/W567 residues are shown in blue and with R566 residues in green. Structures also show hydrogen bonding (yellow dashed lines) between R566/R567 and DNA bases C21, G23, and G4 (gray). C: cytosine; G: guanine. c Overlay of wild-type (red) and R567W mutant (white) CTCF-DNA structures showing changes in hydrogen bonding (yellow dashed lines) between the R567/W567 residue and surrounding DNA bases and phosphate backbone upon mutation. Structures of (a–c) were visualized using the PyMoL Molecular Graphics System91. Hydrogen bonds are shown as dashed lines. d Comparison of normalized CTCF binding strength for brain tissues between Ctcf+/+ and CtcfR567W/R567W mice. e CTCF U motifs of downregulated CTCF sites derived from Supplementary Fig. 7f. f Overlap analysis of downregulated CTCF binding sites (log2(fold change of Ctcf+/+/CtcfR567W/R567W) > 1) in brain, heart and lung tissues (left). Average CTCF binding strengths (middle) and ratios of different types of U motifs (right) were analyzed for each set of overlapping peaks. g Comparison of genome-wide insulation scores in brain tissues between Ctcf+/+ and CtcfR567W/R567W mice. The color in the plot represents the density of points. h Models depicting different types of TADs caused by the CTCFR567W mutation. i Bar plot showing proportions of different types of TADs in brain, heart, and lung tissues. j Hi-C and CTCF ChIP-seq tracks illustrating the stable and variant TADs among brain, heart and lung tissues between Ctcf+/+ and CtcfR567W/R567W mice. TAD boundaries identified for each tissue are shown at the bottom.
To validate the hypothesis, we synthesized oligonucleotide probes containing the CTCF C motif (M1) and U motif (M2), respectively, based on the previous studies40. We then purified recombinant fusion proteins for Glutathione S-Transferase (GST)-tagged wild-type (wt) CTCF ZF domain (ZF1–11), ZF1–11 with R567W mutation (mut), and ZF1–11 with R567W/R566C double mutation (dmut) (Supplementary Fig. 6a). In vitro electrophoretic mobility shift assay (EMSA) results indicated that CTCF mutation diminished migration of both U and C motif probes, as quantified by the bound/free ratios (Supplementary Fig. 6b, c). Additionally, GST pull-down assays followed by dot blot analysis revealed reduced enrichment of both R567W and R567W/R566C mutant proteins for all tested probes, including the U, C, U + C and mutant U + C motif probes (Supplementary Fig. 6d, e). Collectively, these in vitro findings confirm that the CTCFR567W-mutant ZFs exhibit a diminished capacity to bind DNA motif probes compared to wild-type ZFs.
To assess the alterations in chromatin binding affinity of the CTCFR567W-mutant protein in vivo, we conducted adjusted chromatin immunoprecipitation sequencing (ChIP-seq) experiments on brain, heart, and lung tissues in Ctcf+/+ and CtcfR567W/R567W mice using anti-CTCF antibody (Supplementary Fig. 7a). Surprisingly, we observed a reduction in CTCF binding at numerous sites in the CTCFR567W mutant group across all tissues (Fig. 4d and Supplementary Fig. 7b, c), suggesting weakened CTCF chromatin binding by CTCFR567W. Additionally, a greater reduction in CTCF binding corresponded to an increased enrichment of U CTCF motifs, specifically the ZFs 9–11 binding motifs, while the core consensus sequence remained invariant (Supplementary Fig. 7d).
To scrutinize the detailed binding properties affected by CTCFR567W, we performed motif enrichment analysis on our ChIP-seq data using a previously described method41. Using brain tissues as an example, we categorized CTCF sites into four groups based on alterations in CTCF binding strength following the CTCFR567W mutation. For each group, we extracted 20 bp upstream sequences of CTCF core consensus binding motif, which were then divided into 10 subclusters (Supplementary Fig. 7e). The sequence logo for each subcluster indicated that sites with a greater decrease in CTCF binding upon mutation tended to contain a higher percentage of the U motif (group 1: 3.81%, group 2: 8.57%, group 3: 27.79%, group 4: 31.69%) (Supplementary Fig. 7e). In addition to the canonical CTCF U motif, we identified several U motif-like sequence logos, termed noncanonical U motifs, at the downregulated CTCF sites following the mutation. To comprehensively assess the CTCF U motif, clustering analysis was performed using the upstream sequences of all downregulated CTCF sites in three tissues, revealing seven types of upstream motifs labeled from U1 to U7, with U1 corresponding to the canonical CTCF U motif (Fig. 4e and Supplementary Fig. 7f). Collectively, CTCF sites containing U motifs accounted for ~60% of downregulated CTCF sites (Supplementary Fig. 7g), and about 20–30% of total CTCF sites in each tissue, respectively (Supplementary Fig. 7h). The reduced CTCF sites post-CTCFR567W mutation in different tissues contained similar proportions of different types of U motifs (Fig. 4f). These results suggest that CTCFR567W partially reduces CTCF binding, potentially achieved by weakening its ability to bind the U motif.
CTCFR567W leads to local reorganization of higher-order chromatin structures
CTCF mediates long-range chromatin interactions42,43. To investigate whether CTCFR567W disrupts chromatin organization, Bridge Linker-Hi-C (BL-Hi-C) was performed in the brain, heart, and lung tissues of E18.5 Ctcf+/+ and CtcfR567W/R567W mice. Quality control analysis revealed nearly identical distance decay curves and good reproducibility between replicates in each tissue (Supplementary Fig. 8a, b). The Ctcf mutation had little effect on compartments (Supplementary Fig. 8c) but noticeably altered the insulation score, implying potential changes to TADs (Fig. 4g and Supplementary Fig. 8d). Further analysis classified TADs into stable, fusion, separation, and confusion types (Fig. 4h). While most TADs remained unchanged (Fig. 4i), disturbed TADs were observed only in certain regions, exhibiting a tissue-specific alteration pattern (Fig. 4j). These results indicate that CTCFR567W reduces CTCF binding and locally alters chromatin structure, which may be linked to the disruption of gene expression.
CTCFR567W mutation contributes to differential binding and disruption of enhancer-promoter interactions at the Pcdhβ locus
In our investigation into the connection between the CTCFR567W mutation-mediated chromatin structural variation and gene expression, we identified DEGs within TADs and invariant TAD boundaries. Notably, a majority of DEGs were located within stable TADs (Fig. 5a). Further analysis revealed a significant number of DEGs existed within the same TAD, with a particular focus on the cPcdh clusters, encompassing over 30 DEGs on chromosome 18 (Chr18) (Fig. 5b). Visualization of this locus exhibited a subdivision into two subTADs, one corresponding to Pcdhα and the other to Pcdhβ and Pcdhγ. Intriguingly, the subTAD containing Pcdhβ and Pcdhγ further fragmented into two subTADs post-CTCFR567W mutation (Fig. 5c).

a Correlation of DEGs positions with TAD boundaries and four types of TADs post CTCFR567W mutation in different tissues. b Heatmap showing the number of DEGs located in one TAD or TAD boundary. The genomic location (Mb) of selected TADs or TAD boundaries are shown on the right with the associated number of DEGs. c Tracks showing Hi-C maps, insulation scores with different window sizes and normalized CTCF ChIP-seq signals in brain tissues of Ctcf+/+ and CtcfR567W/R567W mice at the cPcdh locus. d Fold change in cPcdh gene expression in brain tissue RNA-seq data (top) and corresponding CTCF binding strength (bottom) in the promoter of each gene in brain tissues of Ctcf+/+ and CtcfR567W/R567W mice. Pcdhα-E: enhancer for Pcdhα; Pcdhβγ-E: enhancer for Pcdhβγ. e 4C tracks showing the interactions between promoters and distal enhancers of cPcdh in brain tissues of Ctcf+/+ and CtcfR567W/R567W mice. SE: super enhancer. f Schematics summarizing the CTCFR567W mutation in regulating higher-order chromatin structure at the cPcdh locus.
A detailed examination of cPcdhs expression indicated a prevailing downregulation of genes within this locus, although some remained unchanged or exhibited upregulation (Fig. 5d). The genes at the cPcdh locus were characterized by CTCF binding sites in their promoters, interacting with CTCF on the distal enhancer to facilitate long-range interactions44. Strikingly, changes in CTCF binding strength at the cPcdh promoters precisely corresponded with the changes in expression of cPcdh genes, while CTCF binding only slightly reduced at the two enhancer elements (Pcdhα-E and Pcdhβγ-E) (Fig. 5d and Supplementary Fig. 8e). Considering the cooperative role of CTCF and cohesin in mediating chromatin interactions45, our examination of cohesin subunit RAD21 occupancy via ChIP-qPCR at the cPcdh locus in the brain tissue of Ctcf+/+ and CtcfR567W/R567W mice yielded consistent results with CTCF. This included decreased RAD21 at Pcdhβ promoters and enhancers, accompanied by an increased binding at Pcdhγa5, b4, and b5, respectively, indicating a downstream shift of chromatin loops away from the Pcdhβ locus (Supplementary Fig. 8f, g). QHR-4C experiments46 demonstrated a significant weakening of the connections between the distal enhancer (SE-f) and the promoters of Pcdhβ genes in CtcfR567W/R567W mice (Fig. 5e), suggesting the loss of multiple long-range interactions resulted in a noticeable alteration in the TAD structure of this locus, complementing the observed TAD splitting and isolation of the Pcdhβ locus into a subTAD (Fig. 5c, f).
The CTCF binding changes induced by the CTCFR567W mutation in the cPcdh locus were further elucidated. Notably, the overall binding strength of CTCF at the cPcdh locus was significantly weaker than the genome-wide average (Supplementary Fig. 8h), implying weak CTCF motifs. Motif analysis of CTCF core motifs and flanking 20 bp sequences revealed that all these CTCF sites possessed weak CTCF core motifs, and sites with decreased CTCF binding at the cPcdh locus were highly enriched with U motifs, while unchanged or increased CTCF binding sites were not (Supplementary Fig. 8i).
Together, the CTCFR567W mutation selectively reduces CTCF binding in the promoters of Pcdhβ genes and alters the chromatin structure of the cPcdh locus by decreasing EP interactions. This leads to a downregulation of cPcdh genes, potentially correlating with impairments of neural function.
CTCFR567W mutation leads to developmental anomalies of hESC-derived cortical organoids
To evaluate the functional consequences of the CTCFR567W mutation within the human cell system, we employed CRISPR/Cas9 technology to introduce the mutation into hESCs (Supplementary Fig. 9a). Two clones each for wild-type, heterozygous, and homozygous cells were successfully generated, confirmed through PCR and Sanger sequencing of both genomic DNA and cDNA (Supplementary Fig. 9b–d; cloning and validation details were shown in the Methods section). Notably, our investigation revealed that the CTCFR567W mutation had no effect on CTCF expression or splicing patterns, aligning with our observations in the mouse model across diverse tissues (Supplementary Fig. 9e, f).
Utilizing wild-type and CTCFR567W hESCs, we initiated the generation of cerebral cortical organoids following the established protocols47,48. In contrast to wild-type organoids, the homozygous (CTCFR567W/R567W) organoids displayed significant developmental defects, manifesting as poor organization, reduced survival rates beyond 2 months, and substantially smaller sizes during differentiation, as evidenced by images captured at days 20, 30, and 60, respectively (Fig. 6a, c, d). Conversely, the heterozygous (CTCF+/R567W) organoids initially exhibited smaller sizes but reached dimensions comparable to those of wild-type organoids by days 60 and 90 (Fig. 6a–d).

a, b Representative photographs (a: days 20, 30, 60; b: day 90) of the CTCF+/+, CTCF+/R567W, and CTCFR567W/R567W organoids. Scale bars in (a): 500 μm, scale bars in (b): 5 mm. c, d Survival rate (c) and size quantification (d) (95% confidence interval) of CTCF+/+, CTCF+/R567W and CTCFR567W/R567W organoids on days 20, 30 and 60, respectively. e UMAP visualization of scRNA-seq data integrated from cortical organoids and fetal brain cortices, colored by cell type. f, g UMAP visualization of cortical organoid scRNA-seq data after integration, colored by predicted cell type (f) and sample (g). h UMAP visualization of the log-fold change in cell proportions between CTCF+/+ and CTCF+/R567W cells in 1 month cortical organoid scRNA-seq data. i Distribution of cell type proportions among cortical organoid scRNA-seq data from 1 month and 2 month CTCF+/+ and CTCF+/R567W samples. j GO analysis of DEGs for each cell type from scRNA-seq in CTCF+/R567W organoids compared to scRNA-seq in CTCF+/+ organoids. The starting number of organoids was 32 for each genotype (a–d), each genotype contains two clones. Quantitative data are presented as mean ± SD. p-values from one-way ANOVA with Dunnett’s multiple comparisons test adjustment are indicated in the graphs. Experiments in (a–d) were repeated independently three times with similar results. All Box and whisker plots show lower and upper quartiles (box limits), median (center line) and minimum to maximum values (whiskers). Source data are provided as a Source Data file.
Furthermore, CTCFR567W/R567W organoids exhibited reductions in the levels of both the neural progenitor marker SOX2 after 1 month of differentiation and the projected neuronal marker SATB2 at 3 months, indicative of early depletion of stem cells/progenitor cells and abnormalities in neurodevelopment (Supplementary Fig. 9g). Notably, excitatory neuronal marker genes (e.g. VGLUT1, VGLUT2, and TBR1) were abnormally downregulated (Supplementary Fig. 9h), while inhibitory neuronal marker genes (including GAD1, GAD2, VGAT, etc.) were abnormally upregulated in CTCF+/R567W organoids compared with wild-type organoids (Supplementary Fig. 9i), mirroring findings observed in single-cell data from the mouse cortex.
Concurrent with the observed developmental anomalies, we performed RNA-seq experiments on CTCF+/+, CTCF+/R567W, and CTCFR567W/R567W hESC-derived organoids on day 35 (Supplementary Fig. 10a). Unlike the limited number of DEGs observed in mouse models, both CTCF+/R567W and CTCFR567W/R567W cortical organoids exhibited hundreds of DEGs compared with the wild-type organoids (Supplementary Fig. 10b–d). Co-upregulated genes in both mutant organoids were associated with mitochondrial metabolism-related pathway, while co-downregulated genes primarily participated in developmental processes, including forebrain development, axonogenesis, and stem cell population maintenance (Supplementary Fig. 10e). These downregulated pathways elucidate the phenotypes of small size, stem cell depletion, and abnormal neurodevelopment in the mutant organoids, consistent with the clinical manifestations of growth retardation and intellectual impairment in heterozygous CTCF mutant cases.
Moreover, in line with the data presented in the mouse model, we observed extensive downregulation of cPCDH genes in both CTCF+/R567W and CTCFR567W/R567W organoids (Supplementary Fig. 10f), underscoring the conservation of the effects induced by the CTCFR567W mutation at this locus. These findings indicate that hESC-derived CTCFR567W heterozygous cerebral cortical organoids may also exhibit developmental irregularities, supporting the involvement of the R567W mutation in CTCF-related clinical developmental disorders.
CTCFR567W heterozygous mutation expedites depletion of stem-like cells and prematurely drives development of hESC-derived cortical organoids
To assess if organoids with solely CTCFR567W heterozygous mutation manifested comparable functional consequences to those observed in the snRNA-seq data of the mouse cortex, we conducted scRNA-seq on CTCF+/+ and CTCF+/R567W cortical organoids at 1- and 2 -month differentiation intervals, encompassing a total of 71,955 individual cells. In order to contextualize our organoid data, we integrated the scRNA-seq data information with the human fetal developmental brain scRNA dataset from the Allen Brain Atlas. Notably, our organoid data aligned well with the data from the fetal brain cortex (Supplementary Fig. 10g, h). Subsequently, we categorized the cells in the integrated organoid dataset into distinct subgroups such as erythrocytes, fibroblasts, glioblasts, immune cells, neural crest cells, neuroblasts, neurons, neuronal intermediate progenitor cells (IPCs), oligos, radial glia, and vascular cells (Fig. 6e–g). Cells of unidentified subtypes were further annotated based on specific cell markers, utilizing INA (a neuronal marker gene) and SLC32A1 (a GABAergic marker gene) (Supplementary Fig. 10i).
Our results unveiled significant differences in cell type proportions between the CTCF+/+ and CTCF+/R567W organoids at the 1 month mark. A notable rise in GABAergic neurons and a simultaneous decline in radial glial cells were observed (Fig. 6h). This trend was consistent with the visualization of the CTCF+/+/CTCF+/R567W cell proportion for each cell type in the 1 month organoids (Fig. 6i). Intriguingly, even at 1 month, CTCF+/R567W organoids exhibited a substantial number of GABAergic cells, with proportions intermediate between the 1 month and 2 month CTCF+/+ organoids. By 2 months, the cell type proportions between the two groups began to converge (Fig. 6i). Our differential gene expression analysis between CTCF+/+ and CTCF+/R567W cell types revealed a decreased in the number of DEGs during differentiation from the 1 month to the 2 month, indicating that the gene expression differences are more pronounced in the early stages of organoid differentiation (Supplementary Fig. 10j). Enrichment analysis of the of 1 month data revealed that the CTCF heterozygous mutation affects numerous neural development pathways, including forebrain development, axonogenesis, and neuron projection guidance (Fig. 6j). We identified 77 genes that overlapped with risk genes associated with brain development disorders in GWAS, including the RBFOX1 and RUNX1T1 genes, which were differentially expressed in all cell types and associated with ASD (Supplementary Fig. 10k, l).
Our findings establish that the CTCFR567W heterozygous mutation in hESC-derived cortical organoids accelerates the depletion of stem-like cells and triggers premature development of GABAergic neurons, mirroring the effects observed in the mouse cortex at the single-cell level. These perturbations in cellular composition and neural development-related pathways offer crucial insights for understanding specific clinical neurodevelopmental disorders.
Discussion
In this study, we established a murine model to simulate clinical developmental disorders associated with the CTCFR567W mutation, revealing the indispensability of CTCF-R567 in normal development. Given the presence of the CTCFR567W mutation in clinical cases of neurodevelopmental disorders and the respiratory distress observed in our homozygous mice, our focus centered on investigating the effects of CTCFR567W on brain and cardiopulmonary tissues. Our results suggest that the impact of CTCFR567W on genes is cell-type specific, implying a broad influence on neurodevelopment and respiratory function, with potential implications for hematopoiesis, immunity, and other systems requiring further investigation.
The lack of homozygous individuals with CTCF mutations may be attributed to the dose-dependent function of CTCF, leading to the inability of homozygotes to survive. This supposition is congruent with the severe defects in hESC-mediated cortical organoids containing the homozygous mutation. Furthermore, the suboptimal performance of heterozygous mice on the rotarod hinted at minor abnormalities in cardiorespiratory function, aligning with the phenotype observed in some human patients with CTCF heterozygous mutations17,18,20,21. However, direct evidence linking the CTCFR567W mutation to cardiac irregularities remains unclear and requires further investigation. Concurrently, despite our heterozygous mice could not fully phenocopy the clinical neurodevelopmental disorders, the observed cellular imbalance in heterozygous hESC-derived cortical organoids may offer essential insights into these specific disorders.
CTCFR567W heterozygous organoids displayed an early depletion of stem-like cells and an increase in GABAergic neurons compared to wild-type organoids. This aligns with our findings in snRNA-seq data of homozygous mutant mice, suggesting that CTCFR567W mutation may disrupt the differentiation process of stem-like cells into specific neurons in both human and mouse systems. This finding corresponds with previous reports indicating that CTCF is crucial for maintaining a balance between neural progenitor cell proliferation and differentiation during early forebrain development16. Recent scRNA-seq data on cortical organoids of autism risk genes also identified an increase in GABAergic neurons48, suggesting that CTCF mutations, similar to autism risk factors, may induce neurodevelopmental disorders through shared cellular development pathways.
The single amino acid mutation, R567W within ZF11 of CTCF, significantly impacts CTCF binding to chromatin, particularly at sites containing the upstream motif. This finding is consistent with earlier findings indicating that mutations within ZF11 of CTCF primarily affect sites with this motif, yet over 60% of binding sites are maintained39,49. In our study, we took advantage of adjusted ChIP-seq methods for CTCF in mouse tissues and found that a single point mutation could significantly alter the chromatin functions of CTCF, which had the similar effect as observed after ZF11 deletion. It’s important to note that individual heterogeneity and variations in methodologies and systems used across different studies could also lead to the certain biases. The altered binding specificity of CTCFR567W at sites with the U motif can be attributed to the essential functions of ZFs 9–11 in recognizing this motif and flanking sequences involved in boundary insulation50. This finding underscores the requirement of R567 for sequence-specific DNA-binding activity and the chromatin function of CTCF, which ultimately influences specific biological phenotypes. Similar to other CTCF ZF mutations, alterations in DNA binding at specific loci can potentially lead to a spectrum of phenotypes, ranging from loss to gain-of-function effects51.
CTCFR567W exerts a broad and diverse impact on gene expression across distinct cell types, with DEGs potentially influenced by direct or indirect downstream effects of the CTCFR567W mutation. These effects may accumulate and become more evident at E18.5 during extended embryonic tissue development. Notably, among these DEGs, a substantial downregulation is observed in most cPcdh genes, both in human and mouse models, consistent with previous findings in Ctcf-conditional-knockout mice23,24. Dysregulation of cPcdh is strongly associated with abnormal neural functions and deletion of this cluster proves fatal at birth in mice35,52,53.
In the mouse model, CTCFR567W leads to a global reduction in binding at the promoters of Pcdhβ cluster genes and induces changes in local 3D organization by dividing one subTAD covering the Pcdhβ and Pcdhγ regions into two subTADs. Transcriptome data from human organoids also reveal a significant downregulation of PCDHΒ genes. Given the highly conserved regulatory role of CTCF at this locus in humans and mice, we opted not to explore further the potential regulatory mechanisms in the complex organoid system. It is important to note that cPcdh encodes neuronal identity and exhibits single cell-specific expression pattern34. The congruence of regulation and chromatin conformational changes mediated by CTCFR567W at the individual neuron level with patterns found in bulk cells warrants further investigation. Our analysis primarily focused on the cPcdh locus in the nervous system at the regulatory mechanism level. However, the impact of the CTCFR567W mutation on the regulatory mechanisms of other tissue-specific differentially expressed genes, particularly those in the heart and lung tissues, remains an avenue for future study.
In conclusion, successful engagement of peripheral CTCF ZFs such as ZF11 containing DNA-binding residue R567 is essential for normal development across various tissue types. This study provides valuable disease models for further exploration into the pathogenesis of clinically relevant CTCF mutations. Furthermore, these models hold potential for aiding in the development of therapeutic interventions by manipulating the 3D chromatin structure.
Methods
The animal experiments were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Animal Experiments of Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences (IACUC: 2023085). The use of human H1 ESCs (WiCell, WA01) in this study is compliant with the Guidance of the Ministry of Science and Technology for the Review and Approval of Human Genetic Resources and was approved by the Life Science and Medical Ethics Committee of the Guangzhou Institutes of Biomedicine and Health under license number GIBH-LMEC2023-024-01 (AL).
Mice
Mice were accommodated in a specific pathogen-free (SPF) facility, maintained under controlled conditions of 20 °C temperature and 50% humidity, with a 12 hour (h) dark/light cycle. Food and water were provided ad libitum. C57BL/6 N mice (Cyagen Biosciences, Guangzhou, China) harboring the CTCFR567W mutation were generated via CRISPR/Cas9-mediated genome editing. The single guide RNA (sgRNA) targeting Ctcf was inserted into the gRNA-spCas9 cloning vector. Single-strand oligodeoxynucleotides (ssODNs) with targeting sequences and mutation sites were synthesized and flanked by 60–80 bp homologous sequences. Ctcf gRNA and Cas9 mRNA were produced by in vitro transcription and coinjected into fertilized eggs with ssODN donors to obtain mutant mice. The mice were identified by PCR and DNA sequencing for genotyping. Mice carrying the Ctcf mutation allele were mated with wild-type C57BL/6 N mice to obtain heterozygous offspring, which were then crossed to generate CtcfR567W/R567W mice. Great efforts have been made to ensure that genetics and environment are comparable among groups in this study. When the number of mice needed for an experiment was small enough to be obtained from a single litter, littermate mice were used for comparisons between genotypes. For experiments requiring larger numbers of mice, pups were selected from two or more litters bred during the same period and housed them under identical conditions. The targeted sgRNA and ssODN sequences for genome editing and the genotyping primers are listed in Supplementary Tables 1 and 2.
Plasmid construction
For construction of the sgRNA-Cas9 vector for gene editing, sgRNA targeting human CTCF was synthesized and constructed into the pX459 vector (pSpCas9(BB)-2A-Puro, Addgene Cat#62988). For construction of the prokaryotic expression vector, the sequence for CTCF ZF fragments was amplified from the cDNA of mESCs and then cloned into the pGEX-4T-2 vector, and the desired mutations were introduced with site-directed mutagenesis to construct the pGEX-ZF-wt, pGEX-ZF-mut (R567W) and pGEX-ZF-dmut (R566C, R567W) vectors.
Tissue processing and cell culture
Pregnant mice were anesthetized with 1.2% v/v avertin (2,2,2-tribromoethanol) (Sigma, Cat# T48402) and sacrificed by cervical dislocation. The embryos were removed, and the desired fetal mouse tissues were taken immediately after decapitation. For brain tissue retrieval and neural cell culture, the whole brain of the fetal mouse was removed by dissection, the meninges were stripped, and the cerebral cortex and hippocampus were separated. Tissues were digested by using the papain dissociation system (Worthington, Cat# LK003160) according to the manufacturer’s instructions. Cells were filtered through a 70 μm filter membrane, counted and inoculated in culture dishes coated with poly-D-lysine (PDL) (Sigma, Cat# P6407). Primary neurons were first cultured in medium containing DMEM/F12 (Gibco, Cat# 11130032) supplemented with 10% v/v fetal bovine serum (FBS) (Vistech, SE100-B) and 1% v/v penicillin/streptomycin (Gibco, Cat# 15140122) at 37 °C in a 5% CO2 incubator. The medium was replaced after 3–4 h with neurobasal medium (Gibco, Cat# 21103049) supplemented with 2% v/v B27 (Gibco, Cat# 17504044), 1% v/v GlutaMAX (Gibco, Cat# 35050061) and 1% v/v penicillin/streptomycin. The medium was changed every other day. HEK293T cells were cultured in high-glucose DMEM (HyClone, Cat# SH30022.01) supplemented with 10% v/v FBS. Feeder-free H1 hESCs (WA01) (WiCell) were cultured in PGM1 human pluripotent stem cell culture medium (Cellapy, Cat# CA1007500) in Matrigel (Corning, Cat# 354277)-coated culture plates. Then, the cells were digested into clumps with 0.5 mM EDTA for normal culture or digested into single cells with Accutase for passaging (STEMCELL Technologies, Cat# 07920) and cultured with medium containing 10 μM ROCK inhibitor Y-27632 (Sigma, Cat# SCM075) for one day; then the inhibitor was withdrawn, and cells were cultured in normal medium to form individual clones.
Generation of CTCFR567W point mutation-containing hESC lines
Human H1 cells harboring the CTCFR567W mutation were constructed via CRISPR/Cas9. sgRNAs were designed with an online tool (http://benchling.com). The ssODN containing the desired mutation and enzymatic cleavage site was used as a template for homologous repair. The sgRNA primers were synthesized, annealed, cloned and inserted into the pX459 vector. The Cas9-sgRNA vector and ssODN were transfected into cells for gene editing using FuGENE HD (Promega, Cat# E2311) transfection reagent. Since Cas9-sgRNA vector contained puromycin resistant gene, the cells were screened with medium containing 1 μg/mL puromycin (Gibco, Cat# A11138-03). After 48 h of screening, the surviving cells were digested into single cells using Accutase for passaging into Matrigel-coated 6-well plates. After ~10 days of culture, the single cells were successfully expanded into individual clones. Over one hundred clones were observed under a microscope and manually picked out using a micropipette. The selected clones were transferred to 96-well plates and further cultured for ~3–5 days until they reached a sufficient size. Subsequently, the clones were digested using 0.5 mM EDTA in PBS to detach them from the culture plates for passaging and amplification. Once the clones reached a certain quantity, a portion of the cells was collected for subsequent genomic DNA extraction and PCR genotyping, followed by enzyme digestion and Sanger sequencing for identification of correctly edited clones.
Detailed information on clones and editing strategies: The first round of editing in wild-type cells using sgRNA1 (target: AACATTTACACGTCGGGTAA) and ssODN1 (introducing the EcoR I site) yielded several heterozygous clones, which were verified by genotyping to have one allele that was correctly mutated and the other allele with a 6 bp deletion. These clones were referred to as heterozygous clone 1. Several wild-type clones were also retained after transfection and were selected as editing control wild-type clones. In the second round of editing, sgRNA2 (target: TGGGAAAACATTTACACGTC) and ssODN2 (introducing the Mlu I site) or ssODN3 (introducing the EcoR I site) were used to edit heterozygous clone 1 and wild-type cells, respectively. This process yielded homozygous clones 1 and 2, and heterozygous clone 2. The clones were verified by genomic PCR, enzyme digestion, and Sanger sequencing. cDNA PCR (with primers for exons 8–10 [aa 469–644]) and sequencing also confirmed that the clones were correct (Supplementary Fig. 9a–d). The sequences of the targeted sgRNAs, ssODNs and genotyping primers used are listed in Supplementary Tables 1 and 2.
RNA extraction and RT-qPCR
Total RNA was extracted from cells and tissues with an RaPure Total RNA Micro Kit (Magen, Cat# R4012-03) according to the manufacturer’s instructions. For cells, 1 × 105 – 1 × 106 cells were counted and collected. For tissues, the isolated mouse tissues were immediately crushed with a homogenizer. RNA quantification and reverse transcription were performed with HiScript III RT SuperMix for qPCR (+ gDNA wiper) (Vazyme Biotech, Cat# R323-01). Real-time PCR was performed with SYBR Green mix (Genstar, Cat# A301-01) on a CFX96 real-time PCR system (Bio-Rad) according to the manufacturer’s instructions. Gapdh mRNA was used as an internal control for normalization of gene expression. The primers used in this study are listed in Supplementary Table 3.
Protein extraction and immunoblotting
Isolated mouse tissues or cultured cells were lysed with RIPA buffer (1% v/v Triton X-100, 0.1% w/v sodium dodecyl sulfate (SDS), 150 mM KCl, 50 mM Tris-HCl (pH 7.4), 1 mM EDTA, 1% w/v deoxycholic acid (sodium salt)) supplemented with protease inhibitors (PIC) (Bimake, Cat# B14001) followed by sonication. The total protein supernatant was obtained by high-speed centrifugation, and the samples were separated on SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Cat# IPVH00010). The membranes were blocked with 5% w/v skim milk and incubated with primary and secondary antibodies before being washed with TBS-T buffer (20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.1% v/v Tween-20). Finally, the membranes were assessed with an enhanced chemiluminescence (ECL) Kit (NCM Biotech, Cat# P10300). The antibodies used in this study are listed in Supplementary Table 6.
Histology, immunofluorescence, and image acquisition
Intact mouse tissues were fixed with 4% w/v paraformaldehyde (Beyotime Biotechnology, Cat# P0099) for 24 h and then made into 3 μm paraffin sections per standard protocols. Hematoxylin and eosin (H&E) staining was performed according to the manufacturer’s instructions (Servicebio, Cat# G1005). For immunohistochemistry, sections were treated with 0.3% v/v H2O2 for 15 min, washed with PBS, blocked with 1% w/v BSA containing 0.3% v/v Triton X-100 for 1 h, and incubated with antibodies at 4 °C. After washing 5 times with PBS, the sections were incubated with secondary antibodies and then stained with DAB and hematoxylin. Images were acquired with a Pannoramic MIDI slice scanner (3D HISTECH).
For immunofluorescence, samples were rinsed with PBS for 3 min, and fixed in 4% w/v paraformaldehyde for 10 min. Following PBS rinses and permeabilization in 0.5% v/v Triton X-100 for 15 min, samples were blocked in 1% w/v BSA/PBS for 1 h. Next, primary antibodies diluted in 1% w/v BSA/PBS were added and incubated for 2 h at room temperature (RT). Following the removal of the primary antibody and subsequently wash with PBS, secondary antibodies were added at a 1:500 dilution in 1% BSA/PBS and incubated for 1 h at RT. After washing with PBS three times for 5 min each time, the samples were stained with DAPI/PBS (1:1000) for 10 min and washed with PBS. The antibodies used in this study are listed in Supplementary Table 6.
For image acquisition, the samples were analyzed by using a Pannoramic MIDI slice scanner (3D HISTECH, CaseViewer 2.4.0, 20x objective), a light microscope (OLYMPUS IX73, cellSens software, 40x objective) or a LSM 710 NLO confocal laser scanning microscope (Carl Zeiss Technology, ZEN 2012 software, 40x objective). Sholl analysis54 was performed on individual neurons by using ImageJ software to quantify their morphological characteristics.
Golgi staining
E18.5 mice were delivered by cesarean section as described above. Golgi staining experiments were conducted using an FD Rapid GolgiStain™ Kit (FD Neuro Technologies, Cat# PK401-50) according to the kit instructions. Briefly, the brains of E18.5 mice were removed as quickly as possible, soaked in an equal-volume mixture of solutions A and B while protected from light at RT for 2 weeks and then transferred to solution C for another 3 days. The impregnated brains were sliced using a vibratome (Leica VT1200S) at a thickness of 100 μm. The sections were immersed in a mixture of solutions D and E for 10 min, dehydrated, cleaned with xylene and then sealed with resin sealing tablets. Images of brain regions of interest were taken by bright-field microscopy.
Intracellular Ca2+ imaging
Neurons from the cortex or hippocampus were loaded with Fluo-4-AM (4 μM) (Yeasen, Cat# 40704ES72) in culture medium at 37 °C for 30 min. Afterward, the neurons were washed with PBS three times, and culture medium was added for the following measurements. The relative fluorescence intensity was measured with a fluorescence microscopic system equipped with a FITC filter. During the measurements, KCl was added to a final concentration of 30 mM to depolarize the neurons and evoke Ca2+ influx. The recorded images were quantified with ImageJ software.
Multielectrode array (MEA) measurements
MEA experiments were recorded using the Maestro Pro system (Axion Biosystems). Briefly, brain tissues were isolated and digested as described above. Approximately 1 × 105 primary neurons were inoculated into PDL-coated 48-well MEA plates (Axion Biosystems, Cat# M768-tMEA-48W), and primary neural cells were cultured as described above. The spontaneous neural electrical activity of the cultured cells was recorded weekly, and the medium was changed 24 h prior to recording. The plates were loaded into the Maestro Pro instrument, and electrical activity was recorded by AxIS Navigator software for 15 min, measuring the number of active electrodes (electrodes with an average of 5 spikes per min), the number of spikes, the number of bursts (individual electrodes with > 5 spikes in 100 ms), the number of network bursts (35% of electrodes with > 50 spikes in 100 ms simultaneously), the synchronization index (with a synchrony window of 20 ms) and other metrics. The data were exported using the Neural Metric Tool and analyzed with GraphPad Prism 9 software.
Mouse behavioral tests
Mice were group-housed (5–8 animals per cage) in standard filter-top cages with ready access to water and rodent food and maintained on a 12 h light/dark cycle with 50–60% relative humidity. Mouse behavioral experiments were performed using 8–12 week-old male mice as described previously55,56,57. During data collection, investigators were blinded to the group allocation of the mice to ensure the unbiased results. Each apparatus was cleaned with 75% v/v alcohol spray prior to testing each animal to prevent bias due to olfactory cues.
Open-field test (OFT)
Each mouse was placed in an open-field arena (44.5 cm × 44.5 cm × 44.5 cm) and allowed to explore freely for 15 min. The activity of the mice in the box was automatically recorded with a camera and the animal behavior analysis software EthoVision XT 13 (Noldus), and each mouse was tested only once.
Novel Object Recognition test (NOR)
NOR experiments were performed in the same open-field arena (44.5 cm × 44.5 cm × 44.5 cm) as the OFT. During the habituation phase, mice were allowed to freely explore the arena and two identical objects for 5 min. The mice were then placed back in the home cage for 1 h as a time interval sufficient for detection of memory in the next phase. During the testing phase, one of the objects was replaced with a new object, and mice were allowed to explore freely for 5 min. The exploration time and times as well as the distance from each object (touching the object with the mouth or nose and approaching the object within ~2–3 cm were considered as exploration of the object) were automatically recorded with a camera and the animal behavior analysis software EthoVision XT 13 (Noldus).
Three-chamber test for social interaction and novelty behavior
The test apparatus consisted of a rectangular three-chambered box. Each chamber measured 20 × 40 × 22 cm and was evenly separated by walls with two retractable doors (each door 3 × 5 cm), allowing mice access to each chamber. An unfamiliar male mouse (stranger 1), which had no prior contact with the subject mice, was placed in one of the side chambers. This unfamiliar mouse was enclosed in a small, round wire cage that enabled nose contact between the bars but prevented fighting. The wire cage was 11 cm high and 9 cm in diameter at the bottom, with vertical bars spaced 0.5 cm apart. Subject mice were first placed in the middle of the chamber and allowed to freely explore throughout the test box for 10 min. The time spent around the cage was measured with the aid of a camera mounted on top of the box to quantify of social preference for stranger 1. A second unfamiliar male mouse (stranger 2) was placed in an identical small wire cage on the other side of the chamber. Thus, the test mice could choose between the first unfamiliar mouse that had already been studied (stranger 1) and a new unfamiliar mouse (stranger 2). As before, the test mice were free to move around in the three chambers for 10 min. The animal behavior analysis software EthoVision XT 13 (Noldus) automatically recorded the time and number of mutual contacts between the test mice and Stranger 1 and Stranger 2.
Elevated plus maze test (EPM)
The EPM apparatus consisted of two open arms and two closed arms (each 9.5 cm wide and 40 cm long) arranged in a cruciform pattern, with the intersection being the central zone, which was 55 cm above the floor. To assess anxiety, mice were placed in the central area, facing an open arm and allowed to explore freely for 5 min. The time and number of explorations in the open and closed arms were automatically recorded by EthoVision XT 13 (Noldus).
Tail suspension test
A cylindrical tube was placed over the tail of the mouse to prevent the mouse from holding its leg during suspension. One end of the tape was stuck to the tail of the mouse, and the other end was fixed to the hanging tail support. The distance from the head of each mouse to the platform was ensured to be the same. The suspension lasted for 6 min, and the immobility time during the last 4 min was recorded with a video camera and analyzed.
Forced swim test
Mice were placed in a 25 cm high, 10 cm diameter clear glass cylinder filled with water to a depth of 10 cm at 22 °C. A 6 min swimming test was performed on each mouse, and the last 4 min of immobility time was recorded for further processing.
Morris water maze
Mice were placed in a circular tank (137 cm in diameter) filled with water (22 ± 1 °C) and divided into quadrants with visual cues for spatial orientation. The circular platform (10 cm in diameter) was adjusted to be submerged 1 cm below the water surface, and the position of the platform was unchanged. Each mouse entered the water from one of the four quadrants at a time. Each mouse was gently placed into the water facing the wall of the tank and each trial lasted 1 min; the trial ended when the mouse climbed onto and stayed on the hidden platform for 10 s. The software automatically recorded the time taken by the mouse to reach the platform as the latency; if the mouse failed to find the hidden platform within 1 min, the mouse was guided to the platform and allowed to stay on the platform for 15 s. The mouse was removed and dried with a dry towel. Four trials were performed per mouse per day at intervals of 30 min or more for 6 consecutive days of training. On day 7, the platform was removed, and each mouse was placed in the water in the quadrant opposite the target quadrant. The software was used to record the number of times each mouse crossed the platform as well as the time and distance it moved in the target quadrant within 1 min. The probe trial was performed twice.
Rotarod test
The rotarod test involved 3 training sessions per day for 4 days. The first day began with acclimatization of all experimental mice at a lower speed (4 r/min) to eliminate fear, with each mouse trained 3 times for 5 min each at intervals of 30 min or more. The next 3 days, the rotarod was set to accelerate from 4–25 rpm for 300 s and the time spent by each mouse on the rotarod was recorded.
Protein purification
BL21(DE3) chemically competent cells (TransGen Biotech, Cat# CD601) were transformed with pGEX-4T-2, pGEX-ZF-wt, pGEX-ZF-mut (R567W) and pGEX-ZF-dmut (R566C, R567W) plasmids and grown to an OD600 of 0.6–0.8 at 37 °C in LB medium containing 100 μg/mL ampicillin. The cells were induced by adding 1 mM IPTG and 200 μM ZnSO4 at 28 °C for 6–8 h. The cells were collected by centrifugation at 4 °C and resuspended in appropriate lysis buffer (PBS with 1 mM DTT, 0.5 mM EDTA) supplemented with 1 × PIC before being sonicated. The cell debris was removed by centrifugation at 20,000 × g. The supernatant was filtered through a 0.45 μm membrane and combined with prewashed glutathione agarose resin (Yeasen Biotech, Cat# 20507ES10) via incubation at 4 °C for 2–4 h. The GST fusion proteins were then washed with PBS-T buffer (PBS with 1% v/v Triton X-100 and 1 × PIC) and eluted with glutathione elution buffer (50 mM Tris-HCl (pH 8.0), 10 mM reduced glutathione, 2 mM DTT, 150 mM NaCl and 1% v/v Triton X-100). The eluted proteins were quantified by the Bradford method and concentrated for subsequent use.
Electrophoretic mobility shift assay (EMSA)
EMSA experiments were performed using a chemiluminescent EMSA Kit (Beyotime Biotechnology, Cat# GS009) according to the manufacturer’s instructions. Briefly, probes for the CTCF motif were synthesized and annealed. The probes were incubated with appropriate amounts of purified GST fusion proteins for 10–20 min at RT and then electrophoresed on a 5% v/v native polyacrylamide gel in ice-cold 0.5 × TBE buffer (45 mM Tris-borate, 1 mM EDTA (pH 8.0)). The gels were scanned in a gel imaging system (Azure Biosystems). The bound/free ratios of probes were quantified with ImageJ software. The sequences of EMSA probes of CTCF motifs are listed in Supplementary Table 4.
GST pull-down assay
GST pull-down experiments were performed by incubating appropriate amounts of biotin-modified probes, purified GST fusion protein and glutathione agarose resin for 6–12 h. Then, glutathione agarose was washed with PBS-T buffer (PBS with 1% v/v Triton X-100) supplemented with 1 × PIC. After centrifugation, the precipitated complexes were digested with proteinase K (Tiangen Biotech, Cat# RT403). The enriched probes were extracted with phenol:chloroform:isoamyl alcohol (25:24:1, Sigma-Aldrich, Cat# P3803) followed by ethanol precipitation and purification. The enriched DNA was spotted onto an Amersham Hybond-N+ membrane (GE Healthcare, Cat# RPN203B), UV cross-linked for 10 min and incubated with streptavidin-HRP (Beyotime Biotechnology, Cat# A0303). Finally, the membranes were washed and detected with an ECL chemiluminescence kit. The sequences of GST pull-down probes of CTCF motifs are listed in Supplementary Table 4.
RNA-seq and data analysis
The RNA-seq experiments followed as previously described58. Total RNA was extracted from cells and tissues with an RaPure Total RNA Micro Kit according to the manufacturer’s instructions. The libraries were constructed using the VAHTS Universal V8 RNA-seq Library Prep Kit for Illumina (Vazyme Biotech, Cat# NR605) according to the manufacturer’s instructions. Briefly, total RNA was purified by two rounds of mRNA purification steps with mRNA capture beads to ensure removal of rRNA. The mRNAs were fragmented to 250–450 bp at 85 °C for 6 min. Immediately afterward, cDNA first- and second-strand synthesis, end-repair and adapter ligation reactions were performed. cDNA purification and size selection were performed with AMPure XP magnetic beads (Beckman Coulter, Cat# A63882). The product was used for library amplification and purification followed by sequencing on an Illumina NovaSeq platform (Annoroad Gene Technology Co., Ltd.). For RNA-seq data analysis, paired-end reads were handled with Trim Galore (version 0.6.5) and then mapped to the mouse (mm10) or human (hg38) genome using STAR aligner (version 2.7.0)59. Transcript abundance was quantified by RSEM (version 1.2.22)60. To remove the effect of sex on gene expression in mouse samples, genes originating from ChrX and ChrY were filtered out. DEGs were determined using DESeq2 (version 1.32.0)61 with q-value < 0.05 and fold-change > 1.5. Gene Ontology (GO) analysis was performed using clusterProfiler (version 4.0.0)62. The DEGs of mouse brain tissues and neurons are listed in Supplementary Data 1. The DEGs of human cortical organoids are listed in Supplementary Data 2.
SnRNA-seq and data analysis for mouse brain cortex
Nuclei of E18.5 mouse brain cortex tissues were extracted using lysis buffer (10 mM Tris-HCl (pH 7.4), 10 mM NaCl, 3 mM MgCl2, 0.1% v/v NP 40, 0.01% w/v digitonin, 1% w/v BSA, 0.1% v/v Tween-20, 1 mM DTT, 1 U/μL RNase inhibitor). The suspended nuclei were filtered using a cell strainer and counted using Countess II FL Automated Cell Counter (Thermo Fisher Scientific) for subsequent single-cell capture. SnRNA-seq library was performed by SequMed Biotech Inc. (GuangZhou, China) following the manufacturer’s instructions using BD Rhapsody WTA Amplification kit (BD Biosciences, Cat #633801). Briefly, the cells and barcoded magnetic beads were added to the microwell plate chip of the BD Rhapsody system. Each bead was linked with a cell in a microwell. The next steps involved cell lysis within the microwells, release of mRNA with polyA tails, and its capture by the oligo-dT of the magnetic beads. The captured single-cell mRNA transcriptome was then collected, and reverse transcription was performed to obtain cDNA sequences. The cDNA underwent second-generation sequencing library construction and was subjected to quality control before being sequenced on the Illumina NovaSeq platform in PE 150 mode.
For snRNA-seq data analysis, the BD Rhapsody whole transcriptome analysis (WTA) pipeline was utilized to process FASTQ files, converting raw sequencing reads into gene expression matrix with unique molecular identifier (UMI) counts for each gene in each cell. The Seurat (version 4.0.5) was utilized for further analysis of the read count matrix for each gene/sample. Quality control measures were applied to filter individual cells, retaining only those expressing genes detected in at least 3 cells and cells expressing a minimum of 200 genes. Additionally, cells with gene expression counts ranging from 500–6000, mitochondrial gene expression not exceeding 20%, and a minimum RNA count of 2000 per cell were retained. The analysis yielded 13137 single cells in wild-type sample and 15661 in mutant sample that were used for subsequent clustering and differential expression analyses.
For cell annotation, the ScType package37, which employs machine learning algorithms and reference datasets, was used. This package annotated the cell types based on the gene expression profiles of the single cells, leveraging marker genes or gene expression patterns to assign cell types. To identify genes showing differential expression between CTCF mutated sample and wild type sample, the Seurat package’s FindMarkers function was employed. Differential expression analysis utilized the Wilcox method, which applies the Wilcoxon rank-sum test to compare gene expression levels between groups of cells. Benjamini-Hochberg method was used for multiple testing correction of p-values to ensure the rigor of the statistical results. A corrected p-value < 0.05 with log2 fold change above 0.5 was considered statistically significant. All significant DEGs in each cell type were selected for GO enrichment analysis using clusterProfiler (v3.18.1). The DEGs are listed in Supplementary Data 3.
SnRNA-seq and data analysis for mouse heart and lung tissues
SnRNA-seq of E18.5 heart and lung tissues were performed by Jiayin Biotechnology Ltd. (Shanghai, China) according to the instruction manual of the Chromium Single Cell 3’ Reagent Kit v3 (10x Genomics, Cat# PN-1000075) with the 10x Genomics platform.
For snRNA-seq data analysis, paired-end reads were aligned to the mouse reference genome (mm10) using Cell Ranger (version 6.1.1). Raw count matrices were used to remove ambient RNA with SoupX (version 1.6.1)63 and then analyzed using Seurat (version 4.1.1)64. Poor-quality cells (> 50% mitochondrial genes, < 500 genes and < 500 UMI per cell for heart samples; > 5% mitochondrial genes, < 500 genes and < 1000 UMI per cell for lung samples) were filtered out. Potential doublets were removed with DoubletFinder (version 2.0.3)65. The data were then normalized and integrated using the SCTransform integration workflow by regressing out the cell cycle effect and percentages of mitochondria-expressed genes per cell. PCA was performed (npcs = 50), and dimensionality reduction (uniform manifold approximation and projection, UMAP) and clustering were then carried out (dimensions = 30; cluster resolution = 0.2 for heart samples, 0.4 for lung samples). Cell markers for each cell cluster were determined with the FindAllMarkers function. For heart samples, cell types were annotated based on a reference paper66. For lung samples, cell types were manually verified based on canonical markers. DEGs were identified via FindMarkers with a min. pct = 0.25 and an adjusted p-value < 0.05, and are listed in Supplementary Data 4 and 5. GO networks for DEGs were constructed using BinGo (version 3.0.5)67 and then visualized in Cytoscape (version 3.9.1)68.
ChIP-seq
ChIP experiments were performed as described previously with some modifications69,70. Briefly, an appropriate amount of mouse tissue was crushed with a homogenizer, cross-linked with 1% w/v formaldehyde for 10 min and quenched with 0.125 M glycine. The crosslinked tissues were lysed in ChIP lysis buffer (1% w/v SDS, 10 mM EDTA, 50 mM Tris-HCl (pH 8.0) and 1 × PIC) and then sonicated to obtain chromatin fragments 200–400 bp in size. For normalization, an equal amount of HEK293T chromatin was added to the sample chromatin solution of different groups for subsequent ChIP experiments. Immunoprecipitation was carried out using 5 μg of antibodies for a duration of 12 h incubation at 4 °C, followed by a 2 h incubation with Dynabeads™ protein G (Thermo Fisher Scientific, Cat# 10003D). The immune complexes were subsequently washed with low salt buffer, high salt buffer, LiCl wash buffer and TE buffer. The enriched chromatin DNA was reverse-crosslinked and then purified using the MinElute PCR Purification Kit (Qiagen Cat# 28006) for ChIP-qPCR or ChIP-seq library construction. ChIP-seq libraries were constructed using the VAHTS Universal DNA Library Prep Kit for Illumina® V3 (Vazyme Biotech, Cat# ND607) according to the manufacturer’s instructions. Briefly, 1–5 ng of enriched DNA was end-repaired and adapter-ligated. PCR library amplification was performed using VAHTS Multiplex Oligos Set 4 for Illumina (Vazyme Biotech, Cat# N321). DNA size selection and library purification were performed with AMPure XP Beads followed by sequencing on the Illumina NovaSeq platform (Annoroad Gene Technology Co., Ltd.). The primers of ChIP-qPCR used in this study are listed in Supplementary Table 5.
ChIP-seq data analysis
Raw reads were trimmed to remove adapters using Trim Galore and then mapped to the mouse (mm10) and human (hg38) mixed genomes using Bowtie2 (version 2.2.5)71 with the following parameters: --very-sensitive --end-to-end --no-unal --no-mixed --no-discordant. Only uniquely mapped reads with MAPQ > 30 were retained. Reads mapped to the mouse or human genome were separated and then subjected to peak calling using macs2 (version 2.2.7.1)72 with the default options. The CTCF peaks are listed in Supplementary Data 6.
To quantitatively compare the CTCF binding strength between wild-type and CTCF-mutated samples, CTCF ChIP signals were normalized according to the method in a previously published paper with minor changes73. Normalization was performed based on the assumption that the average CTCF binding strength from HEK293T cells mixed in different samples should be the same. The peak summit value of the averaged CTCF binding strength from HEK293T cells for each sample was extracted to calculate the scale factor. The scale factor for wild-type samples was considered to be 1. For corresponding CTCF-mutated samples, the scale factor was calculated by summit (wild-type)/summit (mutant). The scale factors for brain, heart and lung samples are shown in Supplementary Fig. 7a. Normalized bigwig files were generated with the bamCoverage tool in deepTools2 (version 3.5.1)74 using RPGC normalization and scale factors.
CTCF upstream motif analysis
The CTCF core motif was first identified for each CTCF site with FIMO (version 5.5.3)75 using the parameter “--thresh 1e-4” with the CTCF position frequency matrix obtained from the HOCOMOCO database, and then the upstream 20 bp sequence for each CTCF site was extracted. The upstream sequences were clustered using hamming distance. Heatmaps were generated with ggmsa (version 1.3.4)76, and sequence logos were generated with ggseqlogo (version 0.1)77.
The CTCF motif in the promoters of Pcdh genes was analyzed as follows. As the CTCF binding strength in most Pcdh promoters was weak (Supplementary Fig. 8h), CTCF binding sites were hardly identified using the default parameters of macs2. Instead, CTCF binding regions in Pcdh promoters were manually selected based on the CTCF ChIP signal. Then, the CTCF core motif sequences and ± 20 bp flanking sequences were extracted, and sequence logos were generated with ggseqlogo.
Bridge Linker-Hi-C (BL-Hi-C)
The BL-Hi-C experiments were conducted following previously established procedures78. In summary, suitable quantities of mouse tissues were crushed using a homogenizer, cross-linked with 1% w/v formaldehyde for 10 min, and subsequently neutralized with 0.2 M glycine. The nuclei were extracted with SDS lysis buffer (1% w/v SDS, 50 mM HEPES-KOH (pH 7.5), 150 mM NaCl, 1 mM EDTA, 1% v/v Triton X-100, 0.1% w/v Sodium Deoxycholate) and then utilized for subsequent procedures: digestion with HaeIII (NEB, Cat# R0108L) at 37 °C for 12 h, end-plus-A treatment with 10 mM dATP (NEB, Cat# N0440S) and Klenow Fragment (exo -) (NEB, Cat# M0212L) at 37 °C for 30 min, along with proximity ligation with biotin-labeled BL-Linker (oligo sequences were listed in Supplementary Table 2.) at 16 °C for a duration of 4 h. The unligated linker was eliminated using Lambda Exonuclease (NEB, Cat# M0262L) and Exonuclease I (NEB, Cat# M0293L) at 37 °C for 1 h. The DNA was then reverse-crosslinked and purified using a phenol:chloroform:isoamyl alcohol (25:24:1) extraction. Subsequently, the DNA was fragmented into an average size of 300 bp through sonication, and the biotin-labeled DNA was enriched using Dynabeads™ M-280 streptavidin (Thermo Fisher Scientific, Cat# 11205D). The enriched bead-bound DNA underwent end repair, adapter ligation, PCR amplification, and DNA library construction, followed by sequencing on the Illumina NovaSeq platform (Annoroad Gene Technology Co., Ltd.).
BL-Hi-C data analysis
The adapters were trimmed from raw paired-end reads with Trim Galore, and the linker sequences were removed with the trimlinker tool from the ChIA-PET2 package (version 0.9.3)79. The trimmed sequences were then mapped to the mouse (mm10) genome and processed using HiC-Pro (version 2.11.1)80. The reproducibility of the Hi-C replicates was assessed with HiCRep (version 0.2.6)81. The valid pair files generated by HiC-Pro were transformed into hic files with KR normalization using juicer tools (version 1.122.01)82.
A/B compartments were analyzed by juicer tools with 100 kb resolution. Insulation scores (ISs) were calculated with 50 kb resolution using FAN-C (version 0.9.20)83. ISs were calculated with a window size of 400 kb for brain samples and a window size of 300 kb for heart and lung samples. TAD boundaries were determined by the “fanc boundaries” function with the parameter -s 0.7. The genomic regions that had no mapped reads and the lowest 5% covered reads were combined as low-coverage regions, which were excluded from the insulation score and TAD boundary analysis. The region between two nearby TAD boundaries was defined as the TAD. The statistics from HiC-Pro processes and TAD information are summarized in Supplementary Data 7.
Quantitative high-resolution chromosome conformation capture copy (QHR-4C)
QHR-4C experiments were performed as previously described46. Briefly, digested suspensions of 1 × 105–1 × 106 cells from tissues were cross-linked with 2% w/v formaldehyde for 10 min, and crosslinking was terminated with 0.2 M glycine. The cell pellet was permeabilized by lysis buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 5 mM EDTA, 0.5% v/v NP-40, 1% v/v Triton X-100), digested with DpnII overnight (NEB, Cat# R0543S) and subjected to proximity ligation by T4 DNA Ligase (NEB, Cat# M0202S). Then, chromatin was reverse-crosslinked and DNA was purified by using MinElute PCR Purification Kit, and was subjected to sonication, resulting in fragmentation into pieces <1000 bp. To enrich ligation events associated with a specific viewpoint, an appropriate amount of sonicated DNA was taken as a template for linear amplification. This process was conducted for 100 cycles using a 5’ biotin-labeled probe of the viewpoint of interest by using Phanta Max Super-Fidelity DNA Polymerase (Vazyme Biotech, Cat# P505). The amplified products were incubated at 95 °C for 5 min, immediately cooled on ice to obtain ssDNA and then enriched with Dynabeads™ M-280 streptavidin. The bead-bound ssDNA was then ligated with adapters. Finally, QHR-4C libraries were constructed with specific primer pairs (forward primers containing Illumina P5 with sequences near a specific viewpoint and reverse primers containing Illumina P7 with an index and sequences matching the adapter) within the system of Phanta Max Super-Fidelity DNA Polymerase and then sequenced on the Illumina NovaSeq platform (Annoroad Gene Technology Co., Ltd.). The primers for QHR-4C used in this study are listed in Supplementary Table 2.
QHR-4C data analysis
Adapter sequences in raw paired-end reads were removed with Trim Galore. The primer sequence at the 5’ end of read 1 was then trimmed with cutadapt (version 3.4)84. Reads that did not contain primer sequences were discarded. Reads were mapped to the mm10 genome using Bowtie2 with the following parameters: --very-sensitive --end-to-end --no-unal -X 2000. Bam files were imported into the r3Cseq package (version 1.38.0)85. Normalized bedgraph files were thus generated and then transformed into bigwig files using the bedGraphToBigWig tool.
Generation of cortical organoids
Cortical organoids differentiated by different genotypes of hESCs were generated as previously described47,48. hESC clones of different genotypes were digested into single cells for subsequent organoid differentiation. Briefly, 9000 cells per well were reaggregated in ultra-low-cell-adhesion 96-well plates with V-bottomed conical wells (Sbio Japan, Cat# MS9096VZ) in cortical differentiation medium (CDM) I. This medium contained Glasgow-MEM (Gibco, Cat# 11710035), 20% v/v Knockout Serum Replacement (KSR) (Gibco, Cat# 10828028), 0.1 mM Minimum Essential Medium non-essential amino acids (MEM-NEAA) (Gibco, Cat# 11140050), 1 mM sodium pyruvate (Gibco, Cat# 11360070), 0.1 mM 2-mercaptoethanol (Life Technologies, Cat# 21985023), and 1% v/v penicillin/streptomycin. From day 0 to day 6, ROCK inhibitor Y-27632 was added to the medium at a final concentration of 20 μM. From day 0–day 18, WNT inhibitor IWR-1 (STEMCELL Technologies, Cat# 72564) and TGF-β inhibitor SB431542 (STEMCELL Technologies, Cat# 72232) were added at concentrations of 3 μM and 5 μM, respectively. From day 18, the floating aggregates were cultured in 6-well ultra-low-attachment culture dishes (Corning, Cat# 3741) on an orbital shaker at 70 rpm in CDM II medium, containing DMEM/F12, 2 mM Glutamax, 1% v/v N2 Supplement (Gibco, Cat# A1370701), 1% v/v Chemically Defined Lipid Concentrate (Gibco, Cat# 11905031), 0.25 μg/mL Amphotericin B (Gibco, Cat#15290026), and 1% v/v penicillin/streptomycin. On day 35, cell aggregates were transferred to the CDM III medium, consisting of CDM II supplemented with 10% v/v FBS, 5 μg/mL heparin (STEMCELL Technologies, Cat# 07980) and 1% v/v Matrigel (Corning, Cat# 356234). From day 70, organoids were cultured on an orbital shaker at 70 rpm in CDM IV medium, consisting of CDM III supplemented with B27 supplement and 2% v/v Matrigel. The cultures were incubated at 37 °C and 5% CO2. And the medium was replaced every 3–5 days. The cortical organoids at different time points of differentiation, were imaged and collected for size quantification, frozen section preparation and RNA-seq experiments, respectively. The organoids were fixed with 4% w/v paraformaldehyde, immersed in 30% w/v sucrose solution, encapsulated in OCT Tissue Freezing Medium, and snap-frozen at -80 °C. Ten-micrometer sections were made using a cryostat microtome (Leica, Germany) at -20 °C for subsequent immunofluorescence staining. Images and area values were obtained with an Olympus inverted microscope. The measured data are provided in the Source Data table. Bulk RNA-seq experiments and data analysis have been described in the previous methods section.
Organoid dissociation and scRNA-seq
A combination of 3–4 similarly sized cortical organoid spheres of the same genotype were subjected to digestion with papain at a concentration of 1 mg/mL for 10 min at 37 °C on a shaker. The digests were gently pipetted and subjected to another 10 min digestion. The resulting cell suspension was filtered through a 40 µm filter and resuspended in a solution of 1% w/v BSA in PBS for cell counting. The scRNA-seq library for cortical organoids was constructed by SequMed Biotech Inc. (GuangZhou, China). Briefly, single-cell suspensions were processed using the DNBelab C Series Single-Cell Library Prep set (MGI, 940-000047-00). Droplet encapsulation, emulsion breakage, mRNA capture, reverse transcription, cDNA amplification, and purification was used to convert the suspensions into barcoded scRNA-seq libraries. After confirmation of the quality, the libraries were sequenced on the DNBSEQ-T7RS platform.
Computational analysis of organoid scRNA-seq data
Cell Ranger (version 6.1.1) with the human reference genome (hg38) was used to generate the output count matrix. Cells with > 20% mitochondrial content, fewer than 300 features, and features expressed in fewer than three cells were excluded from the analysis. We used Scrublet (version 0.2.3)86 and the gene expression patterns (total counts > 40,000) to determine the doublets. The doublet score was calculated independently for each sample with the default settings and with the expected doublet score set at a rate at 10%.
Four organoid scRNA-seq datasets were generated for this study. The cortex region data were extracted from the Allen Brain’s human fetal developmental brain scRNA data, which were used as reference data for cell type annotation. SCANPY (version 1.9.1)87 and Seurat (version 4.3.0)64 were used for major data analysis and visualization. The following steps were performed in order: data normalization, log-transformation, highly variable gene selection, and PCA. The expression levels were calculated as counts per 10,000 (NC); briefly, the total mapped read counts of a gene within one cell were scaled by the number of total mapped reads of that cell and times multiplied by 10,000. The log2(NC + 1)-transformed values were used for further analysis unless indicated otherwise. The highly variable genes were selected based on the raw count data with flavor as “Seurat_v3”. The top 1000 were selected. PCA was run with the selected highly variable genes.
Data integration was performed on the top highly variable genes via Harmony (version 0.0.6)88 with the default parameters. UMAP was performed on the Harmony-corrected PCA embedding. The neighborhood graphs were calculated using scanpy.pp.neighbors, with 15 local neighborhoods and 50 Harmony-corrected PCAs using the mKNN graph. The connectivities were computed using the UMAP method with Euclidean distance. Then, UMAP embedding was performed using scanpy.tl.umap with a minimal effective distance of 0.5, a spread of 1.0, an initial learning rate of 1.0, a negative sample weighting of 1.0 and a negative edge sample rate of 5. Louvain clustering was performed using sc.tl.louvain on the neighborhood calculated in the previous steps, with a resolution of 1, with the vtraag package and with no weights from the mKNN graph. For each cluster, a logistic regression model was trained on the reference dataset and used to annotate organoid cell types. After cell type annotation, the proportions of various cells in each sample were calculated. Milopy (version 0.1.1) was used to calculate and visualize the changes in cell proportions between wild-type and CTCF-mutated cells in 1-month samples.
DEGs for each cell type population between wild-type and CTCF-mutated samples (see Supplementary Data 8) were selected by Seurat via FindMarkers with min. pct = 0.25 and an adjusted p-value < 0.05. GO enrichment analysis was performed on the sets of differentially expressed genes for each cell type using the enrichGO function in the clusterProfiler (version 3.18.1) package.
Statistical analysis
All statistical analyses were performed using GraphPad Prism software. Statistical parameters are reported as sample quantity (n represents the number of animals, cells or organoids per group), and the statistical details of experiments can be found in the figure legends. All data and graphs are presented as mean ± SD, unless stated otherwise. The n-values of statistical samples and the exact p-values are indicated in the figures. The statistical data are provided in the Source Data file.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The ChIP-seq, RNA-seq, BL-Hi-C, QHR-4C, snRNA-seq and scRNA-seq data reported in this paper have been deposited in the Gene Expression Omnibus (GEO) database under accession code GSE214692 and in the Genome Sequence Archive database89 in the National Genomics Data Center90 (GSA) under accession code CRA008223 and HRA003128. The dataset of this paper has been submitted to the figshare repository (https://doi.org/10.6084/m9.figshare.26010637). The referenced Allen fetal brain data is available from the European Genome Phenome Archive under accession code EGAS00001004107 (https://ega-archive.org/datasets/EGAD00001006049). The GWAS table source of autism spectrum disorder used in this study is available in the GWAS Catalog under accession code EFO_0003756 (https://www.ebi.ac.uk/gwas/efotraits/EFO_0003756). Source data are provided with this paper.
Code availability
All code used in this study was previously published and no customized code was used in this paper.
References
Lupianez, D. G. et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell 161, 1012–1025 (2015).
Hnisz, D. et al. Activation of proto-oncogenes by disruption of chromosome neighborhoods. Science 351, 1454–1458 (2016).
Franke, M. et al. Formation of new chromatin domains determines pathogenicity of genomic duplications. Nature 538, 265–269 (2016).
Rowley, M. J. & Corces, V. G. Organizational principles of 3D genome architecture. Nat. Rev. Genet 19, 789–800 (2018).
Zheng, H. & Xie, W. The role of 3D genome organization in development and cell differentiation. Nat. Rev. Mol. Cell Biol. 20, 535–550 (2019).
Phillips, J. E. & Corces, V. G. CTCF: master weaver of the genome. Cell 137, 1194–1211 (2009).
Nora, E. P. et al. Targeted degradation of CTCF decouples local insulation of chromosome domains from genomic compartmentalization. Cell 169, 930–944 e922 (2017).
Hu, G. et al. Systematic screening of CTCF binding partners identifies that BHLHE40 regulates CTCF genome-wide distribution and long-range chromatin interactions. Nucleic Acids Res. 48, 9606–9620 (2020).
Li, J. et al. An alternative CTCF isoform antagonizes canonical CTCF occupancy and changes chromatin architecture to promote apoptosis. Nat. Commun. 10, 1535 (2019).
Song, Y. et al. CTCF functions as an insulator for somatic genes and a chromatin remodeler for pluripotency genes during reprogramming. Cell Rep. 39, 110626 (2022).
Bailey, C. G. et al. CTCF expression is essential for somatic cell viability and protection against cancer. Int J. Mol. Sci. 19, 3832 (2018).
Gong, S. et al. CTCF acetylation at lysine 20 is required for the early cardiac mesoderm differentiation of embryonic stem cells. Cell Regen. 11, 34 (2022).
Wan, L. B. et al. Maternal depletion of CTCF reveals multiple functions during oocyte and preimplantation embryo development. Development 135, 2729–2738 (2008).
Fedoriw, A. M., Stein, P., Svoboda, P., Schultz, R. M. & Bartolomei, M. S. Transgenic RNAi reveals essential function for CTCF in H19 gene imprinting. Science 303, 238–240 (2004).
Kwak, J. H. et al. Loss of the neuronal genome organizer and transcription factor CTCF induces neuronal death and reactive gliosis in the anterior cingulate cortex. Genes Brain Behav. 20, e12701 (2021).
Watson, L. A. et al. Dual effect of CTCF loss on neuroprogenitor differentiation and survival. J. Neurosci. 34, 2860–2870 (2014).
Gregor, A. et al. De novo mutations in the genome organizer CTCF cause intellectual disability. Am. J. Hum. Genet. 93, 124–131 (2013).
Bastaki, F. et al. Identification of a novel CTCF mutation responsible for syndromic intellectual disability - a case report. BMC Med. Genet. 18, 68 (2017).
Chen, F. et al. Three additional de novo CTCF mutations in Chinese patients help to define an emerging neurodevelopmental disorder. Am. J. Med. Genet. C. Semin. Med. Genet. 181, 218–225 (2019).
Konrad, E. D. H. et al. CTCF variants in 39 individuals with a variable neurodevelopmental disorder broaden the mutational and clinical spectrum. Genet. Med. 21, 2723–2733 (2019).
Wang, T. et al. Large-scale targeted sequencing identifies risk genes for neurodevelopmental disorders. Nat. Commun. 11, 4932 (2020).
Rosa-Garrido, M. et al. High-resolution mapping of chromatin conformation in cardiac myocytes reveals structural remodeling of the epigenome in heart failure. Circulation 136, 1613–1625 (2017).
Hirayama, T., Tarusawa, E., Yoshimura, Y., Galjart, N. & Yagi, T. CTCF is required for neural development and stochastic expression of clustered Pcdh genes in neurons. Cell Rep. 2, 345–357 (2012).
Sams, D. S. et al. Neuronal CTCF is necessary for basal and experience-dependent gene regulation, memory formation, and genomic structure of BDNF and Arc. Cell Rep. 17, 2418–2430 (2016).
McGill, B. E. et al. Abnormal microglia and enhanced inflammation-related gene transcription in mice with conditional deletion of Ctcf in Camk2a-Cre-expressing neurons. J. Neurosci. 38, 200–219 (2018).
Hiraide, T. et al. Genetic and phenotypic analysis of 101 patients with developmental delay or intellectual disability using whole-exome sequencing. Clin. Genet. 100, 40–50 (2021).
Saleh, S. et al. Spectrum of neuro-genetic disorders in the United Arab Emirates national population. Clin. Genet 100, 573–600 (2021).
Yin, M. et al. Molecular mechanism of directional CTCF recognition of a diverse range of genomic sites. Cell Res. 27, 1365–1377 (2017).
Chen, H. et al. CTCF variant begets to short stature by down-regulation of IGF1. J. Mol. Endocrinol. 70, e220193 (2023).
Alharbi, A. B. et al. Ctcf haploinsufficiency mediates intron retention in a tissue-specific manner. RNA Biol. 18, 93–103 (2021).
Tang, Z. et al. CTCF-mediated human 3D genome architecture reveals chromatin topology for transcription. Cell 163, 1611–1627 (2015).
Kubo, N. et al. Promoter-proximal CTCF binding promotes distal enhancer-dependent gene activation. Nat. Struct. Mol. Biol. 28, 152–161 (2021).
Merkenschlager, M. & Nora, E. P. CTCF and cohesin in genome folding and transcriptional gene regulation. Annu Rev. Genom. Hum. Genet 17, 17–43 (2016).
Thu, C. A. et al. Single-cell identity generated by combinatorial homophilic interactions between alpha, beta, and gamma protocadherins. Cell 158, 1045–1059 (2014).
Hasegawa, S. et al. Distinct and cooperative functions for the protocadherin-alpha, -beta and -gamma clusters in neuronal survival and axon targeting. Front Mol. Neurosci. 9, 155 (2016).
Mountoufaris, G. et al. Multicluster Pcdh diversity is required for mouse olfactory neural circuit assembly. Science 356, 411–414 (2017).
Ianevski, A., Giri, A. K. & Aittokallio, T. Fully-automated and ultra-fast cell-type identification using specific marker combinations from single-cell transcriptomic data. Nat. Commun. 13, 1246 (2022).
Ghirlando, R. & Felsenfeld, G. CTCF: making the right connections. Genes Dev. 30, 881–891 (2016).
Nakahashi, H. et al. A genome-wide map of CTCF multivalency redefines the CTCF code. Cell Rep. 3, 1678–1689 (2013).
Li, W. et al. Identification of critical base pairs required for CTCF binding in motif M1 and M2. Protein Cell 8, 544–549 (2017).
Hyle, J. et al. Auxin-inducible degron 2 system deciphers functions of CTCF domains in transcriptional regulation. Genome Biol. 24, 14 (2023).
Merkenschlager, M. & Odom, D. T. CTCF and cohesin: linking gene regulatory elements with their targets. Cell 152, 1285–1297 (2013).
Phillips-Cremins, J. E. et al. Architectural protein subclasses shape 3D organization of genomes during lineage commitment. Cell 153, 1281–1295 (2013).
Wu, Q. & Jia, Z. Wiring the brain by clustered protocadherin neural codes. Neurosci. Bull. 37, 117–131 (2021).
Li, Y. et al. The structural basis for cohesin-CTCF-anchored loops. Nature 578, 472–476 (2020).
Jia, Z. et al. Tandem CTCF sites function as insulators to balance spatial chromatin contacts and topological enhancer-promoter selection. Genome Biol. 21, 75 (2020).
Velasco, S. et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 570, 523–527 (2019).
Paulsen, B. et al. Autism genes converge on asynchronous development of shared neuron classes. Nature 602, 268–273 (2022).
Zuo, Z. et al. On the dependent recognition of some long zinc finger proteins. Nucleic Acids Res. 51, 5364–5376 (2023).
Huang, H. et al. CTCF mediates dosage- and sequence-context-dependent transcriptional insulation by forming local chromatin domains. Nat. Genet. 53, 1064–1074 (2021).
Bailey, C. G. et al. Structure-function relationships explain CTCF zinc finger mutation phenotypes in cancer. Cell Mol. Life Sci. 78, 7519–7536 (2021).
Hasegawa, S. et al. Clustered protocadherins are required for building fnctional neural circuits. Front Mol. Neurosci. 10, 114 (2017).
Jia, Z. & Wu, Q. Clustered protocadherins emerge as novel susceptibility loci for mental disorders. Front Neurosci. 14, 587819 (2020).
Yang, Y. et al. Opposite monosynaptic scaling of BLP-vCA1 inputs governs hopefulness- and helplessness-modulated spatial learning and memory. Nat. Commun. 7, 11935 (2016).
Xu, L. et al. Abnormal neocortex arealization and sotos-like syndrome-associated behavior in Setd2 mutant mice. Sci. Adv. 7, eaba1180 (2021).
Chang, A. N. et al. Neural blastocyst complementation enables mouse forebrain organogenesis. Nature 563, 126–130 (2018).
Katayama, Y. et al. CHD8 haploinsufficiency results in autistic-like phenotypes in mice. Nature 537, 675–679 (2016).
Liu, Y. et al. Functional dissection of PRC1 subunits RYBP and YAF2 during neural differentiation of embryonic stem cells. Nat. Commun. 14, 7164 (2023).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinforma. 12, 323 (2011).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Yu, G., Wang, L. G., Han, Y. & He, Q. Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Young, M. D. & Behjati, S. SoupX removes ambient RNA contamination from droplet-based single-cell RNA sequencing data. Gigascience 9, giaa151 (2020).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587 e3529 (2021).
McGinnis, C. S., Murrow, L. M. & Gartner, Z. J. DoubletFinder: doublet detection in single-cell RNA sequencing data using artificial nearest neighbors. Cell Syst. 8, 329–337 e324 (2019).
Hu, P. et al. Single-nucleus transcriptomic survey of cell diversity and functional maturation in postnatal mammalian hearts. Genes Dev. 32, 1344–1357 (2018).
Maere, S., Heymans, K. & Kuiper, M. BiNGO: a cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21, 3448–3449 (2005).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Dong, X. et al. YY1 safeguard multidimensional epigenetic landscape associated with extended pluripotency. Nucleic Acids Res. 50, 12019–12038 (2022).
Ping, W. et al. RBBP4 is an epigenetic barrier for the induced transition of pluripotent stem cells into totipotent 2C-like cells. Nucleic Acids Res. 51, 5414–5431 (2023).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Hu, B. et al. Biological chromodynamics: a general method for measuring protein occupancy across the genome by calibrating ChIP-seq. Nucleic Acids Res. 43, e132 (2015).
Ramirez, F. et al. deepTools2: a next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 44, W160–W165 (2016).
Grant, C. E., Bailey, T. L. & Noble, W. S. FIMO: scanning for occurrences of a given motif. Bioinformatics 27, 1017–1018 (2011).
Zhou, L. et al. ggmsa: a visual exploration tool for multiple sequence alignment and associated data. Brief. Bioinform. 23, bbac222 (2022).
Wagih, O. ggseqlogo: a versatile R package for drawing sequence logos. Bioinformatics 33, 3645–3647 (2017).
Liang, Z. et al. BL-Hi-C is an efficient and sensitive approach for capturing structural and regulatory chromatin interactions. Nat. Commun. 8, 1622 (2017).
Li, G., Chen, Y., Snyder, M. P. & Zhang, M. Q. ChIA-PET2: a versatile and flexible pipeline for ChIA-PET data analysis. Nucleic Acids Res. 45, e4 (2017).
Servant, N. et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biol. 16, 259 (2015).
Yang, T. et al. HiCRep: assessing the reproducibility of Hi-C data using a stratum-adjusted correlation coefficient. Genome Res. 27, 1939–1949 (2017).
Durand, N. C. et al. Juicer provides a one-lickc system for analyzing loop-resolution Hi-C experiments. Cell Syst. 3, 95–98 (2016).
Kruse, K., Hug, C. B. & Vaquerizas, J. M. FAN-C: a feature-rich framework for the analysis and visualisation of chromosome conformation capture data. Genome Biol. 21, 303 (2020).
Kechin, A., Boyarskikh, U., Kel, A. & Filipenko, M. cutPrimers: A new tool for accurate cutting of primers from reads of targeted next generation sequencing. J. Comput Biol. 24, 1138–1143 (2017).
Thongjuea, S., Stadhouders, R., Grosveld, F. G., Soler, E. & Lenhard, B. r3Cseq: an R/bioconductor package for the discovery of long-range genomic interactions from chromosome conformation capture and next-generation sequencing data. Nucleic Acids Res. 41, e132 (2013).
Wolock, S. L., Lopez, R. & Klein, A. M. Scrublet: computational identification of cell doublets in single-cell transcriptomic data. Cell Syst. 8, 281–291.e289 (2019).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with harmony. Nat. Methods 16, 1289–1296 (2019).
Chen, T. et al. The genome sequence archive family: toward explosive data growth and diverse data types. Genom. Proteom. Bioinforma. 19, 578–583 (2021).
Members, C.-N. & Partners Database resources of the national genomics data center, china national center for bioinformation in 2022. Nucleic Acids Res. 50, D27–D38 (2021).
Schrödinger, L. L. C. The PyMOL molecular graphics system, version 1.8. (2015).
Acknowledgements
We thank Professor Qiang Wu for sharing the protocol of the QHR-4C experiment. The authors also thank the Guangzhou Branch of the Supercomputing Center of the Chinese Academy of Sciences for support. This work was supported by the National Natural Science Foundation of China (31925009), National Key R&D Program of China (2021YFA1100300), the National Natural Science Foundation of China (U21A20195, 32300477), Independent Fund of the State Key Laboratory of Respiratory Diseases (SKLRD-Z-202102), Major Project of Guangzhou National Laboratory (GZNL2023A02010, GZNL2023A02008), and Guangzhou Key R&D Program (2023B03J1230).
Author information
Authors and Affiliations
Contributions
H.Y. initiated the study and designed the experiments. J.Z., Y.L., and H.R. conducted most of the experiments. G.H. performed the bioinformatic analysis for RNA-seq, ChIP-seq and Hi-C experiments. Y.H. and Z.M. helped with scRNA-seq analysis. W.Z. helped with Golgi and Ca2+ imaging. D.W. and Y.Q. helped with histology. N.M. helped with MEA measurements. Y.W., B.J., Haidong.W., H.L., L.Z., Hua W., and G.P. contributed to the work. H.Y., J.Z., G.H., Y.L., and H.R. wrote the manuscript. H.Y. conceived and supervised the entire study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Charles Bailey, Jacob Hanna and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, J., Hu, G., Lu, Y. et al. CTCF mutation at R567 causes developmental disorders via 3D genome rearrangement and abnormal neurodevelopment. Nat Commun 15, 5524 (2024). https://doi.org/10.1038/s41467-024-49684-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-49684-1
- Springer Nature Limited