
Abstract
Major depressive disorder (MDD) and bipolar disorder (BD) are highly disabling illnesses defined by different psychopathological, neuroimaging, and cognitive profiles. In the last decades, immune dysregulation has received increasing attention as a central factor in the pathophysiology of these disorders. Several aspects of immune dysregulations have been investigated, including, low-grade inflammation cytokines, chemokines, cell populations, gene expression, and markers of both peripheral and central immune activation. Understanding the distinct immune profiles characterizing the two disorders is indeed of crucial importance for differential diagnosis and the implementation of personalized treatment strategies. In this paper, we reviewed the current literature on the dysregulation of the immune response system focusing our attention on studies using inflammatory markers to discriminate between MDD and BD. High heterogeneity characterized the available literature, reflecting the heterogeneity of the disorders. Common alterations in the immune response system include high pro-inflammatory cytokines such as IL-6 and TNF-α. On the contrary, a greater involvement of chemokines and markers associated with innate immunity has been reported in BD together with dynamic changes in T cells with differentiation defects during childhood which normalize in adulthood, whereas classic mediators of immune responses such as IL-4 and IL-10 are present in MDD together with signs of immune-senescence.
Similar content being viewed by others


Introduction
Mood disorders have been recognized by the World Health Organization as a major source of disability, morbidity, and mortality worldwide [1]. Among mood disorders, major depressive disorder (MDD) and bipolar disorders (BD) are the most frequent and disabling ones. The lifetime prevalence is about 12% for MDD and 2% for BD [2]. Despite the partially shared clinical presentation during depressive episodes, numerous elements distinguish the two disorders and point to a different pathophysiology. BD is characterized by the presence of manic or hypomanic symptomatology, is highly heritable (approximately 70–80%), tends to have an earlier age of onset and a higher recurrence risk [3]. Lithium, the mainstay of BD treatment, is considered to be a second-line add-on drug in MDD while antidepressants, the first-line treatment of MDD, have dubious clinical efficacy in BD and might have deleterious effects [4]. Differential diagnosis between MDD and BD is usually complicated by the fact that in BD a depressive episode is often its first clinical manifestation [5, 6] and nearly 40% of BD patients are initially misdiagnosed as MDD [7], thus leading to a first-line antidepressant monotherapy that could aggravate the outcome of BD [8, 9].
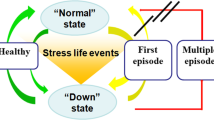
During the last years, increasing evidence for the involvement of immune dysregulations in mood disorders has been accumulating [10,11,12,13] with a focus on the inflammatory response system (IRS), suggesting that an activation of the IRS should be considered as one of the main pathological underpinnings of mood disorders [14]. Furthermore, activation of the IRS with an overproduction of inflammation-regulating cytokines has been shown to affect different mechanisms associated with mood, emotion, and cognition, including neurotransmission, microglial activation, HPA dysregulation, and brain plasticity [15,16,17] (For a summary see Fig. 1).

A HPA axis. In case of inflammation, in response to pro-inflammatory cytokines (especially IL-1, IL-6, TNF-α, and IFN-α), there is increased secretion of corticotrophin-releasing hormone, adrenocorticotropic hormone, and cortisol [157, 158]. Normally, glucocorticoids then act as negative feedback on the inflammatory response [159] to avoid the deleterious effects of excessive production of inflammatory mediators. However, in case of prolonged inflammation (i) chronic high levels of glucocorticoids cause resistance to glucocorticoid feedback on the HPA axis, thus allowing pro-inflammatory signaling pathways to avoid normal feedback inhibition [160]; (ii) the pro-inflammatory cytokines themselves decrease the expression, translocation and downstream effects of glucocorticoid receptors, thereby blunting the negative feedback loop of the HPA axis allowing for further elevation of cortisol levels [161]. Accordingly, increased cortisol levels that are resistant to regulatory feedback by the HPA axis are among the most consistently replicated markers of mood disorder [62]. B Microglia. Neuroinflammation can induce microglial activation. Under physiological conditions, microglia monitors the integrity of synapses [162], removes apoptotic and necrotic cells, and promotes the maintenance of synaptic homeostasis [163]. In turn, microglial activation amplifies the innate immune response by the secretion of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 [164], thus increasing the production of reactive oxygen and nitroxygen species [165]. Currently, it seems that the dualistic classification of microglia activation is not fully representative of the wide repertoire of microglial states and functions in development, plasticity, aging, and disease [166], and further research is then needed to clarify the role of microglia in mood disorders. Prolonged microglial activation induces pathological neuronal apoptotic mechanisms destroying functional neuronal pathways and inhibiting the construction of new pathways, which may manifest in reduced neural plasticity and brain connectivity [167], leading to suboptimal brain function and maladaptive behaviors [168, 169]. Of notice, microglial activation in mood disorders, despite associating with depressive psychopathology, could also play a protective role in counteracting the detrimental effects of yet undefined brain insults, as observed in some brain inflammatory conditions [170]. Increased microglial activation in association with high levels of IL-6 and TNF-α has been observed in mood episodes [171]. PET studies of 17-kDa translocator protein (TSPO) binding, confirmed the presence of a microglial activation both during acute illness episodes and in euthymia [93, 172, 173]. Moreover, microglia activation was found to be associated with cognitive dysfunctions [174], the severity of depression [175], and suicide [176], in agreement with post-mortem findings of higher density of activated microglia in patients with mood disorders who died of suicide [177]. Inhibiting microglial activation with the broadly anti-inflammatory minocycline leads to better neuroplasticity and normalization of the kynurenine pathway in animal models of depression [153, 154], whereas, in treatment-resistant patients with activated peripheral markers of inflammation [155] it has an antidepressant effect. C Plasticity. In the case of inflammation, synaptic plasticity, learning, and memory are inhibited [178]. Pro-inflammatory cytokines affect the availability of brain-derived neurotrophic factor (BDNF), which is the main responsible for structural and functional cellular support [179]. Changes in BDNF levels have been widely reported in depression and have been suggested to underlie the behavioral and mood changes observed in the disorder [180]. Another marker of neuroplasticity is S100B whose effects depend on its concentration. Nanomolar concentrations are associated with the activation of growth and differentiation of neurons and astrocytes whereas micromolar concentrations cause apoptosis of cells, can be neurotoxic, and stimulate the expression of pro-inflammatory cytokines [181]. High serum S100B levels have been reported in major depressive and manic episodes but not in current euthymic mood disorder suggesting glial involvement in the pathogenesis of mood disorders [182]. D Kynurenine Pathway. Inflammatory cytokines (more specifically Interleukin (IL)-2, tumor necrosis factor (TNF)-α, and interferon (IFN)-δ) increase the conversion of tryptophan to kynurenine by activating the indolamine 2,3-dioxygenase [183, 184]. This mechanism causes the depletion of tryptophan and a subsequent decrease in serotonin levels [185]. Moreover, kynurenine degradation leads to the formation of 3- hydroxykynurenine (3-HK) and quinolinic acid (QUIN) or kynurenic acid (KA) [185]. While KA shows a neuroprotective effect, competitively antagonizing NMDA glutamate receptors, 3-HK and QUIN seem to exert neurotoxic effects [186]. Furthermore, IL-6 and TNF-α have been shown to directly increase serotonin turnover by facilitating its release and conversion into 5-hydroxyindoleacetic acid [187, 188]. Of note, serotoninergic abnormalities are a well-established feature of mood disorders pathophysiology [189, 190], with a relative decrease in cortical 5-HT stimulation during depression [191], and excessive activation of the kynurenine pathway, which seems to be shifted towards its neurotoxic branch and contribute to lower the availability of 5-HT [192, 193]. Furthermore, TNF-α, IL-8, IFN-γ, and activation of the kynurenine pathway, have been associated with white matter microstructure alteration in mood disorders [127,128,129,130], a phenotype linking response to antidepressant treatment [101, 131, 132], cognitive impairment [133], exposure to childhood traumatic experiences [134, 135], and severe depressive psychopathology [136]. Effects of inflammatory markers on white matter could also contribute to the association of brain microstructure with depression, as observed in depressive syndromes secondary to medical illnesses involving increased systemic inflammation [194].
When directly looking for specific biomarkers of activation of the IRS in mood disorders, available literature seems to converge towards a framework characterized by alterations in both innate and adaptive immunity with increased inflammatory gene expression in blood cells and higher serum levels of both pro-inflammatory and anti-inflammatory cytokines. To date, few studies tried to discriminate MDD from BD based on differences in immune responses. However, the comprehension of distinct immune profiles characteristic of these two disorders holds paramount significance for facilitating accurate differential diagnoses and the development of personalized treatment strategies.
Thus, the aim of this narrative review is to critically review the current body of literature concerning immune and IRS abnormalities focusing on the most studied biomarkers, e.g. C-reactive protein (CRP), blood cell counts, pro- and anti-inflammatory cytokines, chemokines, monocyte gene expression and T cell subset determinations. We will briefly summarize consistent findings observed separately in MDD and BD patients to provide some background to subsequentially specifically focus on studies utilizing these markers to discriminate between MDD and BD.
Methods
We searched PubMed and ScienceDirect databases for articles up to December 2022. The search was restricted to papers written in English. We included original studies, reviews, and meta-analyses investigating the role of CRP, blood counts, cytokines and chemokines, gene expression, and immune cell populations in mood disorders, especially focusing on studies directly comparing MDD vs BD. We included only peer-reviewed articles; gray literature was excluded. Due to the multifaceted nature of the research question, the inclusion criteria were non-systematic, and papers were included from the query results or from the references cited in the selected articles. All figures were prepared using BioRender.
C-reactive protein
CRP is a pentameric acute-phase protein whose circulating concentrations rise in response to the activation of the innate humoral system due to a wide range of acute and chronic inflammatory conditions [18]. CRP is produced by hepatocytes following IL-6 secretion by macrophages and T cells, playing a primary role in innate immunity as an early defense system [19]. CRP usually does not freely cross the BBB, however, in case of inflammation, it can pass and interact with the CNS [20, 21]. Moreover, CRP itself seems to cause BBB disruption through the binding with some ligands (i.e., Fc gamma receptors, CD16, CD32) expressed in astrocytes and endothelial cells [22]. In the context of mood disorders, different meta-analyses have investigated CRP in MDD and BD.
MDD
In MDD, a cumulative meta-analysis confirmed higher mean levels of CRP in patients compared to controls [23] with a moderate heterogeneity (Q = 51, p < 0.0001, I2 = 62%). In drug-naïve first-episode MDD patients [24] a meta-analysis of 5 studies comprising 179 patients and 210 HC, found that CRP in MDD drug-naive patients was higher (g = 0.53, 95% CI = 0.24–0.82, P < 0.001). Interestingly, a recent study positively correlated circulating CRP levels with cerebrospinal fluid (CSF) levels of CRP and cytokine receptors/antagonists and depressive symptom severity in patients with the highest levels [20].
BD
A meta-analysis investigating 23 original studies and including 1737 BD patients and 81932 HC, found that BD had higher CRP concentration [25]. When subgrouping for bipolar mood state, CRP concentrations were moderately increased in people with a depressive episode (g = 0.67, 95% CI = 0.23–1.11; p = 0.003; 11 studies, 408 patients vs 955 HC) and euthymia (g = 0.65, 95% CI = 0.40–0.90; p < 0.0001; 17 studies, 829 patients vs 79808 HC) and more substantially increased during mania (g = 0.87, 95% CI = 0.58–1.15; p < 0.0001; 14 studies, 500 patients vs 1169 HC). The extent of the increases in CRP concentrations was not related to symptom severity, while CRP concentrations were higher in patients who were not taking psychiatric medication. Finally, CRP concentration was found to decrease after the resolution of a depressive or manic episode. Another meta-analysis based on 37 studies and a total of 2215 BD patients and 3750 HC [26] reported elevated CRP levels in BD compared to HC (g = 0.70, 95% CI = 0.31–1.09). When considering euthymic, depressive, and manic episodes separately, CRP was elevated in both depressive and manic episodes but not in euthymia. Meta-regression showed that gender (r = −0.02, p = 0.021) and duration of illness (r = −0.116, p = 0.046) affected the CRP levels and a meta-analysis of 51 studies reported a positive association between CRP and depression (d = 0.22, 95% CI = 0.15, 0.28, p < .001) in the presence of considerable heterogeneity [27].
MDD versus BD
No meta-analysis has investigated CRP differences between MDD and BD, but a growing literature is now emerging. Bipolar depressed patients (n = 191) had an adjusted OR for inflammation (CRP > 3 mg/L) of 1.44 (95% CI = 0.6–3.4), while unipolar depressed patients (n = 332) had an adjusted OR for inflammation of 0.25 (95% CI = 0.1–0.6) [28]. During acute mood episodes, CRP levels in BD (n = 176) were significantly higher compared to MDD (n = 86). In the post-treatment remission phase, CRP levels were comparable among the diagnostic groups (p = 0.726). The predictive analysis to differentiate bipolar vs unipolar depressive episodes basing on CRP level during acute episodes achieved an AUC of 0.676 and sensitivity and specificity values of 0.636 and 0.737, respectively [29]. A predictive study found that CRP, together with age, direct bilirubin, lactic dehydrogenase, and free triiodothyronine was able to discriminate between adolescent MDD (n = 160) vs BD (n = 101) patients with a balanced accuracy of 73% and an AUC of 0.78 [30]. BD patients showed a higher rate of increased CRP than MDD (respectively 47.4% vs 36.5%) [31]. CRP levels of 6.21 mg/L have been suggested to discriminate between MDD and BD-II in both depressed and euthymic states [32].
CRP
These findings suggest a greater involvement of the IRS in BD as compared to MDD. However, the involvement is not stable but fluctuates together with the phases of the illnesses. Also, recent literature indicates that childhood trauma plays a role in activating the IRS [33].
Blood counts
Blood count ratios, nominally Neutrophil/Lymphocyte ratio (NLR), Platelet/Lymphocyte ratio (PLR), and Monocyte/Lymphocyte ratio (MLR), are low-cost and reproducible markers of IRS activation [34]. Blood count ratios were found to correlate with established markers of inflammation such as CRP and IL-6 [35] and to be less affected by confounding conditions than neutrophils, platelets, or lymphocytes separately [36]. Neutrophils are critical for the first line of immune defenses regulating the innate response through phagocytosis, apoptotic action, chemotaxis, oxidative stress, and secretion of inflammatory mediators [37]. Platelets are a non-specific first-line inflammatory marker that mediate the recruitment of neutrophils and macrophages and their effector functions. Platelet activation is mediated by serotonin, dopamine, glutamate, cytokines, and P-selectin, all of which play generally an important role in psychiatric disorders [38]. Peripheral monocyte count can be considered as a sign of chronic inflammation and might be an indirect marker of microglia activation in the CNS, monocytosis seems to be linked with high levels of activated brain microglia [39, 40]. Finally, lymphocytes are primarily involved in adaptive immunity. Antibody responses are mediated by B lymphocytes whereas cell-mediated immune responses are carried out by T cells which can show a regulatory (anti-inflammatory), an inflammatory or protective function [41]. These ratios are used as biomarkers of inflammation both in medical and neuropsychiatric disorders [42,43,44].
MDD
One meta-analysis focused on MDD showing that patients had higher NLR as compared with HC (SMD = 0.670; p = 0.028; I2 = 89.931%) [42].
BD
Bipolar patients had higher NLR as compared with HC (7 studies, 693 BD vs 641 HC, SMD = 0.672; p < 0.001; I2 = 82.4%) and higher PLR (4 studies, 431 BD vs 368 HC, SMD = 0.425; p = 0.048; I2 = 86.53%). Subgroup analysis evidenced an influence of the bipolar phase on the overall estimate with studies including subjects in manic and any bipolar phase showing a significantly higher NLR and PLR as compared with HC whereas the effect was not significant among studies including only euthymic bipolar subjects. The manic phase of BD is consistently associated with increased neutrophil levels sometimes in conjunction with reduced lymphocyte levels [45]. Less consistent findings have been reported in euthymic bipolar patients who seem to have lower NLR than manic patients.
MDD versus BD
Single studies compared blood cell count between BD and MDD. In a recent large-scale cross-sectional study including 16,174 Chinese affective disorder patients, BD, compared with MDD patients, had higher NLR and MLR and lower PLR, although, MLR was a risk factor for both MDD and BD [46, 47]. However, bipolar patients were younger than MDD and PLR was significantly associated with age. On the contrary, a cross-sectional study using electronic health records of 13,888 psychiatric patients, did not find NLR to differ between bipolar affective disorder and depression after correcting for covariates [48].
Although few studies directly compared MDD and BD the available data are in agreement with those on the CRP in showing a greater activation of the IRS in BD compared to MDD. The altered PLR in BD suggests a greater endothelial dysfunction which could affect the trafficking of peripheral inflammatory markers to the brain [49].
Blood counts
The findings derived from meta-analyses and single studies comparing blood count ratios also consistently confirm that mood disorder patients present with an activation of the IRS when compared with HC. Moreover, a growing literature suggests a stronger activation in BD patients than in MDD patients and, interestingly, an even stronger activation in BD manic patients than in BD-depressed patients.
The stronger abnormalities of the PLR in BD as compared to MDD suggest a greater involvement of endothelial stickiness and permeability in this disorder.
Cytokines and chemokines
Cytokines are small free and membrane-bound proteins that act as signaling molecules to regulate immune cellular activities and inflammation [50]. Cytokines are produced by both immune (e.g., macrophages, lymphocytes, mast cells) and non-immune (e.g., parenchymal cells, endothelial and epithelial cells, fibroblasts, adipocytes, and stromal cells) cells and the same cytokine can be produced by multiple cell types [51]. In the brain, cytokines are produced by microglia, astrocytes, endothelial cells, and neurons [52]. Further, cytokines from the periphery can affect inflammatory processes in the brain via several pathways including humoral (passing the BBB), neural (through the vagus nerve stimulation), and cellular (through the recruitment of leukocytes to the brain) [53]. Physiologically, cytokines are involved in brain development and support neuronal integrity, neurogenesis, and synaptic remodeling [54]. Cytokines can also promote behavioral alterations by influencing neurocircuitry and neurotransmitter systems [55].
Chemokines are a group of secreted proteins within the cytokine family involved in cell migration. By stimulating the migration of cells, they are crucial for immune surveillance as well as for inflammatory reactions. Their function can be either homeostatic, inflammatory, or with a dual function. Homeostatic chemokines are expressed at constitutive levels and are required for basal immune cell migration at a steady state, while inflammatory chemokines are expressed under the control of pro-inflammatory factors, attract immune cells to the site of inflammation [56, 57], induce the release of pro-inflammatory mediators, and aid in the control of T helper (Th)1/Th2/Th17 phenotypic polarization [58].
As indicated above, changes in the activation of the IRS parallel mood fluctuations in acute illness episodes. These episodes of IRS activation show increased levels of both inflammatory and anti-inflammatory cytokines. Antidepressant treatments may then enhance the activity of anti-inflammatory cytokines promoting the return to euthymia. However, immune alterations have also been reported in euthymic patients, suggesting a persistent activation of the IRS with some cytokine alterations becoming apparent during symptomatic relapses while others persist independently of the phase of the illness.
Although several studies have been performed investigating immune/inflammatory markers in depression and across mood states, it is difficult to draw a coherent picture because of contrasting findings in a plethora of tested cytokines/chemokines and their soluble receptors (see Table 1). The majority of the studies performed to investigate these markers in unipolar and bipolar depression focused only on a small number of specific cytokines/chemokines, and there is a great variety in the studies. Conflicting results have been obtained.
In this review, we will focus on just a few cytokines and chemokines, i.e. IL-6, TNF, IL-10, IL-4, CCL2, and IL-8, on which there is consistent literature. We will thereafter review the few studies that directly compared MDD versus BD in studies with larger panels of cytokines and chemokines.
The inflammatory cytokines IL-6 and TNF-α
MDD
A recent meta-analysis of 82 studies performed in 6010 participants (3212 participants with MDD and 2798 HCs) reported consistent increased levels of the pro-inflammatory cytokines IL-6 (g = 0.621; P < 0.001) and TNF-α (g = 0.638; P < 0.001) in MDD compared to HC [59]. Interestingly, different biomarker profiles could be associated with subtypes of MDD, with elevated levels of TNF in atypical depression (characterized, according to DSM-5, by a higher reactivity to environmental stressors) [60], and mixed findings in melancholic depression [59], less reactive to the environment, and with a higher HPA axis activation which might influence peripheral levels of inflammatory markers [61,62,63]. Interestingly, elevated levels of plasma TNF-α were found in patients with atypical depression [60], but not in patients with melancholic depression [59]. This finding is in line with the hypothesis that melancholic depression is associated with increased activity of the HPA axis which would lower inflammation [61, 62].
BD
Studies directly comparing different mood states in BD showed similarly high TNF-α levels [64, 65]. Regarding IL-6, higher levels have been found in manic/hypomanic rather than in depressive and euthymic patients. However, the literature is not clear on relationships with BD phases [66], because also, both a significant decrease in IL-6 [64] levels in manic compared with depressed patients and negative findings of a difference between mood states have been reported [65, 67].
Different studies focused also on TNF receptors showing higher levels of sTNFR1 in both active phases and the euthymic state compared to the depressive state [68, 69]. Also, higher levels of sTNFR80 [70] have been found in manic/hypomanic rather than in depressive and euthymic patients. Finally, an association between these markers and both depressive and manic symptoms has been reported [66]. All in all, recent independent meta-analyses suggest a shared inflammatory profile between both depressive and manic stages characterized by high concentrations of IL-6 and TNF-α [26, 66, 71, 72].
Both higher and lower markers have been reported in euthymic patients. Findings from systematic reviews and meta-analyses showed a permanent increase of IL-6 [73], and unaltered levels of TNF-α [73, 74], whereas contrasting findings have been reported on sTNFR1 [72,73,74].
The anti-inflammatory/regulatory cytokines IL-10 and IL-4
MDD
Higher levels of IL-10 were consistently reported in MDD compared to HC [59]. Furthermore, IL-10 levels were not significantly altered in patients who were antidepressant-free but only in participants who were using antidepressants. This seems to suggest that an antidepressant regimen would boost IL-10-mediated immunosuppression and reduce inflammation.
BD
Increased levels of anti-inflammatory cytokines other than IL-4 are scarcely reported in BD. Regarding IL-4, the consistent finding is an increase in IL-4 during manic phases [71, 72, 75].
Studies directly comparing different mood states showed a significant increase in IL-4 levels [64] in manic compared with depressed state patients. And in euthymic compared to HC [76].
Finally, negative findings of a difference between mood states have been reported for IL-10 which resulted in higher compared to controls in both groups of patients [67].
The chemokines CCL2 and IL-8
MDD
There is also evidence that chemokine alterations occur in MDD. Increased peripheral levels of CCL2 have been observed in MDD [59].
Similarly, increased peripheral [77, 78] and central [79] levels of IL-8 have been reported although less consistently possibly due to age and medication effects as suggested by higher levels in females but not in males [80]. A meta-analysis also showed that peripheral IL-8 levels were significantly decreased in drug-naïve first-episode MDD patients compared with controls [24].
BD
The most consistent findings in BD are: increased CCL2 [68, 81] and IL-8 [82] levels in depressed BD compared to HC [83]; and lower [68] levels of IL-8 in manic patients compared to controls. Further, although no association was observed with values measured in the peripheral blood of euthymic patients, increased IL-8 levels have been reported in the CSF [84].
MDD versus BD
Few studies, implementing both conventional and machine learning methodologies, considered cytokines and chemokines as possible tools to differentiate BD from MDD. Using elastic net penalized logistic regression, we recently showed that BD was predicted by the levels of several cytokines and chemokines including TNF-α, CCL3, CCL4, CCL5, and CCL11; while MDD was predicted mainly by the levels of IL-4, IL-6, and IL-10 with a sensitivity of 79% and a specificity of 99% [83].
With a similar approach and with a sensitivity of 0.77% and specificity of 0.76% IL-6 and IL-10 were associated with BD in a discovery cohort, whereas in a replication cohort, IL-10 was confirmed and TNF-α, CCL3, and CCL11 individuated [85].
Also, higher IL-6, sTNFR2, and lower TNF-α have been identified as predictors of BD through a hierarchical logistic regression, reaching 98.1% accuracy [86].
Considering 10 initial analytes and integrated feature selection, performed through a multivariate wrapper technique, with a support vector machine with a linear kernel and leave-one-subject out cross-validation IL-4 and IL-10 were able to discriminate BD from MDD with a sensitivity of 62% and specificity of 66% [87].
Cytokines and chemokines
A great heterogeneity characterizes the study of cytokines and chemokines in mood disorder patients reflecting the technical heterogeneity of the used assays, the intrinsic heterogeneity of the two disorders with influences of body weight and childhood trauma on the levels of the cytokines, the episodic nature of these illnesses, and the variable effects of drug treatment. So, conclusions are very difficult to draw from the cytokine/chemokine studies, regarding differences between MDD and BD.
Both MDD and BD are characterized by increased pro- and anti-inflammatory cytokines. The studies directly comparing BD versus MDD show higher levels of certain chemokines (CCL3 and CCL11, in two studies) in BD compared to MDD; also, the chemokine IL-8 is prominent in BD. This suggests a greater involvement in the trafficking of cells in BD.
Monocyte gene expression
Monocytes/macrophages are important producers of pro-and anti-inflammatory cytokines and chemokines. The expression of genes regulating inflammatory processes has been extensively studied in monocytes of BD and MDD patients [88]. These studies were not limited to a rostrum of strictly inflammatory genes (such as IL-1, IL-6, CCL2, CCl20, TNF, etc.), but also extended to genes playing a role in the differentiation, apoptosis, mitochondrial function, and lipid metabolism of the cells (such as BAX, BCL10, EGR1, EGR2, NR1H3, ABCA1, ABCG1, MVK, and HMOX). All these genes had been found in whole genome analysis studies discriminating monocytes of MDD and BD patients from those of healthy controls.
MDD
In MDD patients monocytes show an abnormal expression of genes involved in pathways implicated in apoptosis, growth, lipid metabolism, and redox reactions, suggesting a mitochondrial dysfunction and an early aging of the cells [14].
However, in cases of a history of childhood trauma and/or in cases above 30 years of age, there is an additional overexpression of a variety of strictly inflammatory genes involved in the regulation of the IRS via the expression of a number of excitatory and inhibitory genes, amongst which e.g. IL-6, IL-1, CCL20, CXCL2 and DUSP2 and TNFAIP3 [14].
In younger mild MDD cases of less than 30 years with a first episode of depression without signs of anxiety, even a reduced expression of these strictly inflammatory genes was found [89].
BD
In the manic and depressive phases of BD, an increased number of circulating monocytes has been reported [90]. These monocytes show overexpression of a variety of the strictly inflammatory genes involved in the regulation of the IRS via the expression of the same excitatory and inhibitory genes as in MDD.
In euthymic phases of the disorder the overexpression of the inflammatory genes is absent in monocytes [91], and in fact in one report there even is a decrease in the expression of such genes as compared to healthy controls [92].
Also, in mania and bipolar depression PET studies [93] focusing on the activation of microglia report an activated status with an increased PET signal in several regions including the cingulate cortex and the hippocampus.
MDD vs BD
Although no study directly compared monocyte gene expression between MDD and BD, a study investigated the expression of TREM-1 and PU.1 in monocytes of melancholic MDD patients and BD patients with active disease compared to controls [94]. TREM-1 is an important membrane-bound molecule involved in inflammation regulation [95], while PU.1 is a transcription factor important for monocyte/macrophage development and differentiation [96].
TREM-1 gene expression was significantly increased in monocytes of BD patients, while in MDD patients TREM-1 expression only tended to be overexpressed. In contrast, PU.1 gene expression was increased in the monocytes of MDD patients, while it was not increased in the monocytes of BD patients.
Monocyte gene expression
Pieces of evidence show the presence of monocyte activation both in the periphery and the brain in both disorders. Monocytes gene expression suggest that in BD monocytes are more involved in inflammation regulation than the monocytes of MDD patients, while the monocytes of MDD patients are more involved in the regulation of growth and differentiation.
T cell populations
Extensive studies have been performed on subpopulations of T helper cell subsets involved in regulating the inflammatory response in mood disorders (Table 2). T cells are actors of adaptive immunity and together with NK cells contribute to and regulate the cell-mediated immune response. However, the role of monocytes/macrophages and T helper cells within the CNS are not limited to immune and inflammatory response regulation, but also includes the regulation of neurogenesis [97]; further they have been shown to play a role in the maintenance of cognition [98], learning and behavior [99]. Adaptive and innate immune support, ensured by cell trafficking across the BBB, is essential for brain maintenance and repair in healthy conditions, and it is disrupted in several brain disorders [100]. There is a crosstalk between neurons and leukocytes, influencing brain function and structure in mood disorders [101]. Although the field is in its infancy, the few available studies in mood disorders suggest that immune cells influence brain white matter microstructure [102,103,104], and are in turn influenced by pharmacological treatments also partially mediating their effects on the brain [103], thus supporting the study of circulating immune cell composition and activation status as a new promising biomarker. The post-mortem observed abnormal density of T lymphocytes in subgroups of mood disorder patients is consistent with this perspective [105]. Both post-mortem and brain imaging studies also suggest an activation of microglia in patients with mood disorders [39]. Finally, premature aging was observed both in BD (increased proportion of late‐differentiated T cells and reduced proportion of activated T cells after stimulation) which is associated with the number of episodes [106], and MDD, especially in those patients exposed to childhood adversity who show higher signs of inflammation [14], with the expansion of senescent memory T cells [107].
MDD
Th1/Th2
An increase in the Th1/Th2 ratio in the peripheral blood of MDD patients was reported [108], together with a decrease in Th2 cells [91]. However, a recent meta-analysis reported reduced percentages of both Th1 and Th2 [109]. In remission phases, the number of T cells seems to remain unchanged but the in vitro proliferative status of these cells is diminished. Further, T cells undergo apoptosis to prevent uncontrolled proliferation suggesting that T cells may adapt to MDD [110].
Th17 cells
High levels of Th17 cells have been reported in severe forms of MDD, characterized by high depression scores and high suicide risk [111]. However, in less severe forms Th17 counts are normal, and generally, MDD patients are characterized by impaired maturation of Th17 cells (and of Th2 cells and NK cells) and by decreased serum levels of the T cell differentiation factor IL-7 and the T cell activation marker sCD25 [112].
T Regulatory cells
Reports of reduced levels of regulatory cells involve different populations including natural T regulatory cells (Treg) [108, 113, 114] and NK regs [115]. These deficiencies are paralleled by an increased inflammatory state of monocytes as a sign of chronic low-grade inflammation [107]. Antidepressant therapy increased the number of T regulatory cells [107].
Cytotoxic cells
A recent meta-analysis showed no differences between patients and controls for CD8 cells. Further, a higher NK absolute cell count was reported but no difference was observed for relative percentages [109].
BD
TH1/Th2
Although discrepant findings have been reported on Th1 [116] and Th2 [92] cell populations, with both higher levels of Th1 and Th2 and no difference [90] compared to controls, a recent meta-analysis suggest that an imbalance between Th1 and Th2 is present in BD with a shift towards Th2 [117].
TH17
Higher levels of activated CD3+ T cells have been reported more than a decade ago in both symptomatic and euthymic BD patients compared to healthy controls [118], these increases included higher levels of Th17 cells [119]. Patients with BD in a euthymic state indeed have increased levels of Th17 cells compared to healthy individuals [91, 92]. However, there is also a report not showing such increased levels of Th17 cells in euthymic BD patients [90].
To support a view of a high involvement of Th17 in BD a cytokine study has shown higher levels of IL-17 in the circulation of patients with BD in remission [120].
It is also of interest to mention that studies in offspring of a bipolar parent show that in teenage time and adulthood, Th17 cell levels follow a dynamic course as compared to Th17 levels in healthy controls with initial reduced levels in young adulthood. This suggests proneness of the T cell system of BD patients to influences of sex hormones fluctuating in teenage/young adulthood time [113].
T regulator cells
A very recent study shows that euthymic patients with BD show higher levels of Tregs compared to healthy controls. The literature on BD on this cell population is contradictory. An early study showed higher circulating Tregs in younger-than-40-year-old patients with BD in an active disease state [90], but decreased levels in relatively old euthymic BD patients (median 52 years, [92]). Other studies have also shown lower levels of Tregs in patients with euthymic BD [116, 121, 122].
Similar to Th17 cells dynamic changes have been observed in children of a bipolar parent who are at risk for bipolar disorder [113]. Particularly in teenage time reduced numbers of T regulatory cells were found. These reduced numbers of Tregs were associated with a high expression of pro-inflammatory genes in monocytes suggesting a high inflammatory state in teenagers with a bipolar parent; things changed in young adulthood when T cells producing pro-inflammatory cytokines and expression of pro-inflammatory genes in monocytes are reduced suggesting a transition to an anti-inflammatory state [112, 113].
Cytotoxic cells
A reduction of CD8 cells has been reported in BD [121] whereas no differences between BD and HC have been reported for NK cells [103].
MDD vs BD
Becking et al. directly compared the Th17 and Treg cell populations (as well as Th1 and Th2 populations) between MDD patients and BD patients of whom the vast majority was in euthymia. Compared to MDD patients, BD patients showed significantly increased levels of Th17, Th2, Th1, and T regulatory cells (all p < 0.02). Also, In BD patients, levels of Th17 and T regulatory cells were increased compared to HC (p = 0.03, p = 0.02, respectively), while MDD patients showed decreased levels of Th17 and Th2 compared to HC (p = 0.03, p = 0.01, respectively). Of the various medications only SSRI/SNRI usage could explain part of the Th2 decrease in MDD [119]. A study performed on patients during a depressive episode showed that BD patients had lower CD8+ T cell compared to MDD, while there was no difference in the CD4+ T cell [123]. Finally, Li et al. reported lower CD4+ T cell and NK cells but higher CD8+ T cell proportions in BD-depressed patients compared to MDD and a higher CD4/CD8 ratio (a marker of immune-senescence) in MDD compared to BD [124].
Thus collectively, results although not consistently suggest the presence of differences in T cell immune regulation between patients with BD and MDD.
Immune cell populations
A partial T cell defect starting early in adolescence, involving a reduction of naïve T cells and Treg cells and an expansion of memory and senescent T cells with a dynamic pattern of premature immunosenescence seems to characterize MDD patients, especially with childhood trauma [14, 102, 119, 125, 126]. More complicated is the picture in BD where substantial changes in the frequencies of cell populations can be observed throughout the lifespan and possibly also in different mood episodes. Differences between MDD and BD in this pattern are present according to age and phase of the illness and involve different cell populations including Th17 and Th2 which are able to differentiate the two disorders.
Discussion
Although the literature agrees on the presence of distinct profiles of immune biomarkers, the characterization of these profiles is less clear (Fig. 2). Focusing only on the most consistent findings, some pro-inflammatory cytokines such as IL-6 and TNF-α have been associated with both disorders in different studies. Directly comparing the two disorders MDD has been associated with increased levels of IL-4 and IL-10, whereas BD has been associated with NLR, CRP, and several chemokines including CCL3, CCL4, CCL5, and CCL11 and, on the whole, a greater involvement of the IRS.

A immune/inflammatory markers associated only with MDD. B immune markers associated with both disorders but showing different peripheral levels. The differences between the two disorders are represented by a triangle with its vertex indicating the lower plasma level. C immune/inflammatory markers associated only with BD.
It’s not by chance that IL-6 and TNF-α are involved in the main psychopathological mechanism associated with mood disorder (Fig. 1). IL-6 and TNF-α have been associated with white matter microstructure alteration in mood disorders [127,128,129,130], a phenotype linking response to antidepressant treatment [101, 131, 132], cognitive impairment [133], exposure to childhood trauma [134, 135], and severe depressive psychopathology [136].
Focusing on the differences between MDD and BD we observed that the markers increased in MDD, including gene expression of monocytes, are involved in macrophage activation and Th2, Treg differentiation. In BD, increased markers are involved in monocyte/macrophage responses and cellular activation, proliferation, and migration, especially of the Th2 pathway. These patterns could also explain the reduced gray matter volumes reported in BD compared to MDD and the greater number of episodes characteristics of these patients [137].
Whereas the majority of the cytokines are involved in a monocyte/macrophage response and in the recruitment of cells responsible for the adaptive immune response, cytokines, such as IL-4 and IL-10, may protect from an over-reactive immune system, promote repair by immune-regulatory mechanisms and induce a tolerance counteracting the pro-inflammatory system. These actions may be promoted also by IL-6 which exhibits context-dependent immune-regulatory activities [138]. In line with the finding of the role of IL-4 and IL-10 in differentiating BD from MDD, Th2 cell populations have been shown to play the same role, confirming the involvement of these cell populations in mood disorders. Higher levels of Th17 also seem to associate with BD diagnosis where, although counterintuitively, correlated with higher white matter integrity, this finding, together with a lack of association of BD with IL-17 levels may be explained by the high plasticity of Th17 cells which, when induced by IL-6/TGF-b are less inflammatory and have a higher expression of IL-10 [139].
The heterogeneity observed in the literature well reflects the one characteristic of these disorders. Several factors may determine the activation status of the IRS and the mediators involved: (1) the episodic nature of these disorders with different polarities and periods of well-being; (2) the high presence of childhood trauma consistently associated with inflammation; (3) the comorbidities with metabolic disorders such as obesity, type 2 diabetes, and metabolic syndrome which also are linked to inflammation; (4) pharmacologic treatments which on the contrary may reduce inflammation. All these factors make it rather difficult to draw a coherent picture.
From a clinical point of view, in physiological conditions, cytokines interact with 5-HT to shape sleep architecture [140], and perturbing microglial functions, disrupt sleep and the homeostatic processes associated with synaptic potentiation and dendritic spine density [141, 142]. Cytokines’ release follows a circadian pattern which is dysregulated in mood disorders [143]; winter depression associated with higher macrophage activity and lower lymphocyte proliferation [144]; and antidepressant treatments targeting the biological clock normalize both immune functions and depressive symptoms [144, 145]. Disruption of rhythms across the lifespan leads to the onset, recurrence, and worsening of mood disorders [146]: it can be surmised that an abnormal immune system activation could play a key role in this process [147]. Again, the abnormalities in circadian rhythms differ in MDD and BD [148] suggesting different underlying mechanisms.
Inflammatory status has been also associated with response to antidepressant treatments in MDD [149, 150] and BD [17]. Higher levels of circulating cytokines, especially IL-6 and TNF-α, hamper antidepressant response and contribute to treatment resistance [150, 151]. Immune/inflammatory mechanisms have therefore been proposed as possible targets for antidepressant psychopharmacology, and randomized, placebo-controlled trials confirmed the potential efficacy of anti-inflammatory and immune-modulatory agents in treating depression only in subgroups of patients with increased peripheral inflammation [149]; on the other hand, conventional antidepressants share immune-modulatory and anti-inflammatory properties, which could reduce inflammation during depression [152]. Also, inhibiting microglial activation with the broadly anti-inflammatory minocycline leads to better neuroplasticity and normalization of the kynurenine pathway in animal models of depression [153, 154], whereas, in treatment-resistant patients with CRP > 3 [155] it has an antidepressant effect. In agreement with a different immune profile in MDD and BD, a recent study from our group showed how MDD and BD patients respond differently to an immune-modulatory treatment with low-dose IL-2: higher effect of treatment in BD; increase of CD4+Naïve T cells and decrease in CD4+Central Memory cells only in MDD [156]. These effects are in agreement with the different involvement of the IRS in the two disorders and support the usefulness of this perspective in clinical practice.
As already noticed, the available literature is characterized by high heterogeneity indeed, some clinical features, known to affect IRS, should be taken into account when trying to use immune markers to differentiate MDD from BD: (i) age; (ii) BD type I or II; (iii) number of previous episodes; (iv) duration of the illness; (v) pharmacologic treatment; (vi) smoking and BMI; (vii) childhood trauma.
Conclusion
Altogether, available findings about the role of immune-related biomarkers suggest that they can provide new targets to differentiate patients subgroups among the wide heterogeneity of mood disorders, including in providing new endophenotypes for the early differentiation between bipolar and unipolar illnesses; and to address new treatments aimed at preventing the detrimental effect of the illness on brain structure and function, which appears to be related to the repeated insult caused by neuroinflammatory mechanism during the lifetime recurrence of mood episodes. Following this perspective, the differential activation of innate immunity and monocyte/macrophagic activity associated with BD could contribute to explain the worse brain integrity and cognitive deterioration observed in the illness.
The interest for further clinical and preclinical investigations is then warranted to elucidate the immune/inflammatory mechanisms as a possible target for antidepressant psychopharmacology, in order to translate the acquired knowledge into clinical practice aiming at personalized treatment regimens for depressed patients, and to further understand the most promising perspective in elucidating the pathophysiology of mood disorders.
References
Kessler RC, Akiskal HS, Ames M, Birnbaum H, Greenberg P, Hirschfeld RM, et al. Prevalence and effects of mood disorders on work performance in a nationally representative sample of U.S. workers. Am J Psychiatry. 2006;163:1561–8.
Steel Z, Marnane C, Iranpour C, Chey T, Jackson JW, Patel V, et al. The global prevalence of common mental disorders: a systematic review and meta-analysis 1980-2013. Int J Epidemiol. 2014;43:476–93.
McIntyre RS, Berk M, Brietzke E, Goldstein BI, Lopez-Jaramillo C, Kessing LV, et al. Bipolar disorders. Lancet. 2020;396:1841–56.
Taylor DM, Barnes TRE, Young AH. The Maudsley prescribing guidelines in psychiatry. New Jersey: John Wiley & Sons; 2021.
Judd LL, Akiskal HS, Schettler PJ, Coryell W, Endicott J, Maser JD, et al. A prospective investigation of the natural history of the long-term weekly symptomatic status of bipolar II disorder. Arch Gen Psychiatry. 2003;60:261–9.
McIntyre RS, Zimmerman M, Goldberg JF, First MB. Differential diagnosis of major depressive disorder versus bipolar disorder: current status and best clinical practices. J Clin Psychiatry. 2019;80:ot18043ah2.
Hirschfeld RM. Differential diagnosis of bipolar disorder and major depressive disorder. J Affect Disord. 2014;169:S12–16.
Pacchiarotti I, Bond DJ, Baldessarini RJ, Nolen WA, Grunze H, Licht RW, et al. The International Society for Bipolar Disorders (ISBD) Task Force report on antidepressant use in bipolar disorders. Am J Psychiatry. 2013;170:1249–62.
Vieta E. Antidepressants in bipolar I disorder: never as monotherapy. Am J Psychiatry. 2014;171:1023–6.
Maes M, Vandoolaeghe E, Ranjan R, Bosmans E, Bergmans R, Desnyder R. Increased serum interleukin-1-receptor-antagonist concentrations in major depression. J Affect Disord. 1995;36:29–36.
Maes M, Yirmyia R, Noraberg J, Brene S, Hibbeln J, Perini G, et al. The inflammatory & neurodegenerative (I&ND) hypothesis of depression: leads for future research and new drug developments in depression. Metab Brain Dis. 2009;24:27–53.
Blume J, Douglas SD, Evans DL. Immune suppression and immune activation in depression. Brain Behav Immun. 2011;25:221–9.
Wohleb ES, Franklin T, Iwata M, Duman RS. Integrating neuroimmune systems in the neurobiology of depression. Nat Rev Neurosci. 2016;17:497–511.
Simon MS, Schiweck C, Arteaga-Henríquez G, Poletti S, Haarman BC, Dik WA, et al. Monocyte mitochondrial dysfunction, inflammaging, and inflammatory pyroptosis in major depression. Prog Neuropsychopharmacol Biol Psychiatry. 2021;111:110391.
Dantzer R. Neuroimmune interactions: from the brain to the immune system and vice versa. Physiol Rev. 2018;98:477–504.
Najjar S, Pearlman DM, Alper K, Najjar A, Devinsky O. Neuroinflammation and psychiatric illness. J Neuroinflammation. 2013;10:43.
Benedetti F, Aggio V, Pratesi ML, Greco G, Furlan R. Neuroinflammation in bipolar depression. Front Psychiatry. 2020;11:71.
Sproston NR, Ashworth JJ. Role of C-reactive protein at sites of inflammation and infection. Front Immunol. 2018;9:754.
Black S, Kushner I, Samols D. C-reactive protein. J Biol Chem. 2004;279:48487–90.
Felger JC, Haroon E, Patel TA, Goldsmith DR, Wommack EC, Woolwine BJ, et al. What does plasma CRP tell us about peripheral and central inflammation in depression? Mol Psychiatry. 2020;25:1301–11.
Jeon MT, Kim KS, Kim ES, Lee S, Kim J, Hoe HS, et al. Emerging pathogenic role of peripheral blood factors following BBB disruption in neurodegenerative disease. Ageing Res Rev. 2021;68:101333.
Kuhlmann CR, Librizzi L, Closhen D, Pflanzner T, Lessmann V, Pietrzik CU, et al. Mechanisms of C-reactive protein-induced blood-brain barrier disruption. Stroke. 2009;40:1458–66.
Haapakoski R, Mathieu J, Ebmeier KP, Alenius H, Kivimaki M. Cumulative meta-analysis of interleukins 6 and 1beta, tumour necrosis factor alpha and C-reactive protein in patients with major depressive disorder. Brain Behav Immun. 2015;49:206–15.
Cakici N, Sutterland AL, Penninx B, Dalm VA, de Haan L, van Beveren NJM. Altered peripheral blood compounds in drug-naive first-episode patients with either schizophrenia or major depressive disorder: a meta-analysis. Brain Behav Immun. 2020;88:547–58.
Fernandes BS, Steiner J, Molendijk ML, Dodd S, Nardin P, Goncalves CA, et al. C-reactive protein concentrations across the mood spectrum in bipolar disorder: a systematic review and meta-analysis. Lancet Psychiatry. 2016;3:1147–56.
Solmi M, Suresh Sharma M, Osimo EF, Fornaro M, Bortolato B, Croatto G, et al. Peripheral levels of C-reactive protein, tumor necrosis factor-alpha, interleukin-6, and interleukin-1beta across the mood spectrum in bipolar disorder: a meta-analysis of mean differences and variability. Brain Behav Immun. 2021;97:193–203.
Howren MB, Lamkin DM, Suls J. Associations of depression with C-reactive protein, IL-1, and IL-6: a meta-analysis. Psychosom Med. 2009;71:171–86.
Osimo EF, Cardinal RN, Jones PB, Khandaker GM. Prevalence and correlates of low-grade systemic inflammation in adult psychiatric inpatients: an electronic health record-based study. Psychoneuroendocrinology. 2018;91:226–34.
Zhang X, Li X, Tong J, Li N, Zhang F. C-reactive protein in bipolar disorder and unipolar depression. J Nerv Ment Dis. 2022;210:510–4.
Wu X, Niu Z, Zhu Y, Shi Y, Qiu H, Gu W, et al. Peripheral biomarkers to predict the diagnosis of bipolar disorder from major depressive disorder in adolescents. Eur Arch Psychiatry Clin Neurosci. 2022;272:817–26.
Caldirola D, Dacco S, Cuniberti F, Grassi M, Lorusso S, Diaferia G, et al. Elevated C-reactive protein levels across diagnoses: the first comparison among inpatients with major depressive disorder, bipolar disorder, or obsessive-compulsive disorder. J Psychosom Res. 2021;150:110604.
Chang HH, Wang TY, Lee IH, Lee SY, Chen KC, Huang SY, et al. C-reactive protein: a differential biomarker for major depressive disorder and bipolar II disorder. World J Biol Psychiatry. 2017;18:63–70.
Brown M, Worrell C, Pariante CM. Inflammation and early life stress: an updated review of childhood trauma and inflammatory markers in adulthood. Pharmacol Biochem Behav. 2021;211:173291.
Balta S, Demirkol S, Unlu M, Arslan Z, Celik T. Neutrophil to lymphocyte ratio may be predict of mortality in all conditions. Br J Cancer. 2013;109:3125–6.
Guo J, Chen S, Chen Y, Li S, Xu D. Combination of CRP and NLR: a better predictor of postoperative survival in patients with gastric cancer. Cancer Manag Res. 2018;10:315–21.
Gibson PH, Croal BL, Cuthbertson BH, Small GR, Ifezulike AI, Gibson G, et al. Preoperative neutrophil-lymphocyte ratio and outcome from coronary artery bypass grafting. Am Heart J. 2007;154:995–1002.
Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol. 2006;6:173–82.
Dietrich-Muszalska A, Wachowicz B. Platelet haemostatic function in psychiatric disorders: effects of antidepressants and antipsychotic drugs. World J Biol Psychiatry. 2017;18:564–74.
Beumer W, Gibney SM, Drexhage RC, Pont-Lezica L, Doorduin J, Klein HC, et al. The immune theory of psychiatric diseases: a key role for activated microglia and circulating monocytes. J Leukoc Biol. 2012;92:959–75.
Muller N, Wagner JK, Krause D, Weidinger E, Wildenauer A, Obermeier M, et al. Impaired monocyte activation in schizophrenia. Psychiatry Res. 2012;198:341–6.
Alberts B. Molecular biology of the cell. 5th edn. New York: Garland Science; 2008.
Mazza MG, Lucchi S, Tringali AGM, Rossetti A, Botti ER, Clerici M. Neutrophil/lymphocyte ratio and platelet/lymphocyte ratio in mood disorders: a meta-analysis. Prog Neuropsychopharmacol Biol Psychiatry. 2018;84:229–36.
Zhou L, Ma X, Wang W. Inflammation and coronary heart disease risk in patients with depression in China Mainland: a cross-sectional study. Neuropsychiatr Dis Treat. 2020;16:81–86.
Wang J, Zhou D, Dai Z, Li X. Association between systemic immune-inflammation index and diabetic depression. Clin Interv Aging. 2021;16:97–105.
Bhikram T, Sandor P. Neutrophil-lymphocyte ratios as inflammatory biomarkers in psychiatric patients. Brain Behav Immun. 2022;105:237–46.
Wei Y, Feng J, Ma J, Chen D, Chen J. Neutrophil/lymphocyte, platelet/lymphocyte and monocyte/lymphocyte ratios in patients with affective disorders. J Affect Disord. 2022;309:221–8.
Bulut NS, Yorguner N, Carkaxhiu Bulut G. The severity of inflammation in major neuropsychiatric disorders: comparison of neutrophil-lymphocyte and platelet-lymphocyte ratios between schizophrenia, bipolar mania, bipolar depression, major depressive disorder, and obsessive compulsive disorder. Nord J Psychiatry. 2021;75:624–32.
Brinn A, Stone J. Neutrophil-lymphocyte ratio across psychiatric diagnoses: a cross-sectional study using electronic health records. BMJ Open. 2020;10:e036859.
Rybakowski JK, Wykretowicz A, Heymann-Szlachcinska A, Wysocki H. Impairment of endothelial function in unipolar and bipolar depression. Biol Psychiatry. 2006;60:889–91.
Ramesh G, MacLean AG, Philipp MT. Cytokines and chemokines at the crossroads of neuroinflammation, neurodegeneration, and neuropathic pain. Mediators Inflamm. 2013;2013:480739.
Arango Duque G, Descoteaux A. Macrophage cytokines: involvement in immunity and infectious diseases. Front Immunol. 2014;5:491.
Galic MA, Riazi K, Pittman QJ. Cytokines and brain excitability. Front Neuroendocrinol. 2012;33:116–25.
Capuron L, Miller AH. Immune system to brain signaling: neuropsychopharmacological implications. Pharmacol Ther. 2011;130:226–38.
Yirmiya R, Goshen I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav Immun. 2011;25:181–213.
Haroon E, Raison CL, Miller AH. Psychoneuroimmunology meets neuropsychopharmacology: translational implications of the impact of inflammation on behavior. Neuropsychopharmacology. 2012;37:137–62.
Rossi D, Zlotnik A. The biology of chemokines and their receptors. Annu Rev Immunol. 2000;18:217–42.
Zlotnik A, Yoshie O. Chemokines: a new classification system and their role in immunity. Immunity. 2000;12:121–7.
Cyster JG. Chemokines and cell migration in secondary lymphoid organs. Science. 1999;286:2098–102.
Kohler CA, Freitas TH, Maes M, de Andrade NQ, Liu CS, Fernandes BS, et al. Peripheral cytokine and chemokine alterations in depression: a meta-analysis of 82 studies. Acta Psychiatr Scand. 2017;135:373–87.
Spoelma MJ, Serafimovska A, Parker G. Differentiating melancholic and non-melancholic depression via biological markers: a review. World J Biol Psychiatry. 2023;24:761–810.
Gold PW. The organization of the stress system and its dysregulation in depressive illness. Mol Psychiatry. 2015;20:32–47.
Stetler C, Miller GE. Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosom Med. 2011;73:114–26.
Baumeister D, Akhtar R, Ciufolini S, Pariante CM, Mondelli V. Childhood trauma and adulthood inflammation: a meta-analysis of peripheral C-reactive protein, interleukin-6 and tumour necrosis factor-alpha. Mol Psychiatry. 2016;21:642–9.
Ortiz-Dominguez A, Hernandez ME, Berlanga C, Gutierrez-Mora D, Moreno J, Heinze G, et al. Immune variations in bipolar disorder: phasic differences. Bipolar Disord. 2007;9:596–602.
Jacoby AS, Munkholm K, Vinberg M, Pedersen BK, Kessing LV. Cytokines, brain-derived neurotrophic factor and C-reactive protein in bipolar I disorder - results from a prospective study. J Affect Disord. 2016;197:167–74.
Vega-Nunez A, Gomez-Sanchez-Lafuente C, Mayoral-Cleries F, Bordallo A, Rodriguez de Fonseca F, Suarez J, et al. Clinical value of inflammatory and neurotrophic biomarkers in bipolar disorder: a systematic review and meta-analysis. Biomedicines. 2022;10:1368
Mora E, Portella MJ, Pinol-Ripoll G, Lopez R, Cuadras D, Forcada I, et al. High BDNF serum levels are associated to good cognitive functioning in bipolar disorder. Eur Psychiatry. 2019;60:97–107.
Barbosa IG, Rocha NP, Bauer ME, de Miranda AS, Huguet RB, Reis HJ, et al. Chemokines in bipolar disorder: trait or state? Eur Arch Psychiatry Clin Neurosci. 2013;263:159–65.
Barbosa IG, Machado-Vieira R, Soares JC, Teixeira AL. The immunology of bipolar disorder. Neuroimmunomodulation. 2014;21:117–22.
Siwek M, Sowa-Kucma M, Styczen K, Misztak P, Nowak RJ, Szewczyk B, et al. Associations of serum cytokine receptor levels with melancholia, staging of illness, depressive and manic phases, and severity of depression in bipolar disorder. Mol Neurobiol. 2017;54:5883–93.
Anderson G, Maes M. Bipolar disorder: role of immune-inflammatory cytokines, oxidative and nitrosative stress and tryptophan catabolites. Curr Psychiatry Rep. 2015;17:8.
Rosenblat JD, McIntyre RS. Bipolar disorder and inflammation. Psychiatr Clin North Am. 2016;39:125–37.
Rowland T, Perry BI, Upthegrove R, Barnes N, Chatterjee J, Gallacher D, et al. Neurotrophins, cytokines, oxidative stress mediators and mood state in bipolar disorder: systematic review and meta-analyses. Br J Psychiatry. 2018;213:514–25.
Munkholm K, Brauner JV, Kessing LV, Vinberg M. Cytokines in bipolar disorder vs. healthy control subjects: a systematic review and meta-analysis. J Psychiatr Res. 2013;47:1119–33.
Malhi GS, Irwin L, Hamilton A, Morris G, Boyce P, Mulder R, et al. Modelling mood disorders: an ACE solution? Bipolar Disord. 2018;20:4–16.
Brietzke E, Stertz L, Fernandes BS, Kauer-Sant’anna M, Mascarenhas M, Escosteguy Vargas A, et al. Comparison of cytokine levels in depressed, manic and euthymic patients with bipolar disorder. J Affect Disord. 2009;116:214–7.
Eyre HA, Air T, Pradhan A, Johnston J, Lavretsky H, Stuart MJ, et al. A meta-analysis of chemokines in major depression. Prog Neuropsychopharmacol Biol Psychiatry. 2016;68:1–8.
Leighton SP, Nerurkar L, Krishnadas R, Johnman C, Graham GJ, Cavanagh J. Chemokines in depression in health and in inflammatory illness: a systematic review and meta-analysis. Mol Psychiatry. 2018;23:48–58.
Wang AK, Miller BJ. Meta-analysis of cerebrospinal fluid cytokine and tryptophan catabolite alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder, and depression. Schizophr Bull. 2018;44:75–83.
Birur B, Amrock EM, Shelton RC, Li L. Sex differences in the peripheral immune system in patients with depression. Front Psychiatry. 2017;8:108.
Brietzke E, Kauer-Sant’Anna M, Teixeira AL, Kapczinski F. Abnormalities in serum chemokine levels in euthymic patients with bipolar disorder. Brain Behav Immun. 2009;23:1079–82.
Misiak B, Bartoli F, Carra G, Malecka M, Samochowiec J, Jarosz K, et al. Chemokine alterations in bipolar disorder: a systematic review and meta-analysis. Brain Behav Immun. 2020;88:870–7.
Poletti S, Vai B, Mazza MG, Zanardi R, Lorenzi C, Calesella F, et al. A peripheral inflammatory signature discriminates bipolar from unipolar depression: a machine learning approach. Prog Neuropsychopharmacol Biol Psychiatry. 2020;105:110136.
Isgren A, Jakobsson J, Palsson E, Ekman CJ, Johansson AG, Sellgren C, et al. Increased cerebrospinal fluid interleukin-8 in bipolar disorder patients associated with lithium and antipsychotic treatment. Brain Behav Immun. 2015;43:198–204.
Martinuzzi E, Barbosa S, Courtet P, Olie E, Guillaume S, Ibrahim EC, et al. Blood cytokines differentiate bipolar disorder and major depressive disorder during a major depressive episode: initial discovery and independent sample replication. Brain Behav Immun Health. 2021;13:100232.
Brunoni AR, Supasitthumrong T, Teixeira AL, Vieira EL, Gattaz WF, Bensenor IM, et al. Differences in the immune-inflammatory profiles of unipolar and bipolar depression. J Affect Disord. 2020;262:8–15.
Wollenhaupt-Aguiar B, Librenza-Garcia D, Bristot G, Przybylski L, Stertz L, Kubiachi Burque R, et al. Differential biomarker signatures in unipolar and bipolar depression: A machine learning approach. Aust N Z J Psychiatry. 2019;54:393–401.
Gibney S, Drexhage H. EPA-1405 – role of monocyte mRNA in mood disorders. Eur Psychiat. 2014;29:1.
Grosse L, Carvalho LA, Wijkhuijs AJ, Bellingrath S, Ruland T, Ambree O, et al. Clinical characteristics of inflammation-associated depression: monocyte gene expression is age-related in major depressive disorder. Brain Behav Immun. 2015;44:48–56.
Drexhage RC, Hoogenboezem TH, Versnel MA, Berghout A, Nolen WA, Drexhage HA. The activation of monocyte and T cell networks in patients with bipolar disorder. Brain Behav Immun. 2011;25:1206–13.
Becking K, Haarman BC, van der Lek RF, Grosse L, Nolen WA, Claes S, et al. Inflammatory monocyte gene expression: trait or state marker in bipolar disorder? Int J Bipolar Disord. 2015;3:20.
Vogels RJ, Koenders MA, van Rossum EF, Spijker AT, Drexhage HA. T cell deficits and overexpression of hepatocyte growth factor in anti-inflammatory circulating monocytes of middle-aged patients with bipolar disorder characterized by a high prevalence of the metabolic syndrome. Front Psychiatry. 2017;8:34.
Eggerstorfer B, Kim JH, Cumming P, Lanzenberger R, Gryglewski G. Meta-analysis of molecular imaging of translocator protein in major depression. Front Mol Neurosci. 2022;15:981442.
Weigelt K, Carvalho LA, Drexhage RC, Wijkhuijs A, de Wit H, et al. TREM-1 and DAP12 expression in monocytes of patients with severe psychiatric disorders. EGR3, ATF3 and PU.1 as important transcription factors. Brain Behav Immun. 2011;25:1162–9.
Bouchon A, Dietrich J, Colonna M. Cutting edge: inflammatory responses can be triggered by TREM-1, a novel receptor expressed on neutrophils and monocytes. J Immunol. 2000;164:4991–5.
Li G, Hao W, Hu W. Transcription factor PU.1 and immune cell differentiation (review). Int J Mol Med. 2020;46:1943–50.
Ziv Y, Schwartz M. Immune-based regulation of adult neurogenesis: implications for learning and memory. Brain Behav Immun. 2008;22:167–76.
Schwartz M, Kipnis J. A conceptual revolution in the relationships between the brain and immunity. Brain Behav Immun. 2011;25:817–9.
Kipnis J, Cohen H, Cardon M, Ziv Y, Schwartz M. T cell deficiency leads to cognitive dysfunction: implications for therapeutic vaccination for schizophrenia and other psychiatric conditions. Proc Natl Acad Sci USA. 2004;101:8180–5.
Croese T, Castellani G, Schwartz M. Immune cell compartmentalization for brain surveillance and protection. Nat Immunol. 2021;22:1083–92.
Benedetti F, Poletti S, Hoogenboezem TA, Locatelli C, Ambree O, de Wit H, et al. Stem cell factor (SCF) is a putative biomarker of antidepressant response. J Neuroimmune Pharmacol. 2016;11:248–58.
Poletti S, de Wit H, Mazza E, Wijkhuijs AJ, Locatelli C, Aggio V, et al. Th17 cells correlate positively to the structural and functional integrity of the brain in bipolar depression and healthy controls. Brain Behav Immun. 2017;61:317–25.
Furlan R, Melloni E, Finardi A, Vai B, Di Toro S, Aggio V, et al. Natural killer cells protect white matter integrity in bipolar disorder. Brain Behav Immun. 2019;81:410–21.
Magioncalda P, Martino M, Tardito S, Sterlini B, Conio B, Marozzi V, et al. White matter microstructure alterations correlate with terminally differentiated CD8+ effector T cell depletion in the peripheral blood in mania: combined DTI and immunological investigation in the different phases of bipolar disorder. Brain Behav Immun. 2018;73:192–204.
Schlaaff K, Dobrowolny H, Frodl T, Mawrin C, Gos T, Steiner J, et al. Increased densities of T and B lymphocytes indicate neuroinflammation in subgroups of schizophrenia and mood disorder patients. Brain Behav immun. 2020;88:497–506.
Rizzo LB, Swardfager W, Maurya PK, Graiff MZ, Pedrini M, Asevedo E, et al. An immunological age index in bipolar disorder: a confirmatory factor analysis of putative immunosenescence markers and associations with clinical characteristics. Int J Methods Psychiatr Res. 2018;27:e1614.
Grosse L, Carvalho LA, Birkenhager TK, Hoogendijk WJ, Kushner SA, Drexhage HA, et al. Circulating cytotoxic T cells and natural killer cells as potential predictors for antidepressant response in melancholic depression. Restoration of T regulatory cell populations after antidepressant therapy. Psychopharmacology. 2016;233:1679–88.
Li Y, Xiao B, Qiu W, Yang L, Hu B, Tian X, et al. Altered expression of CD4(+)CD25(+) regulatory T cells and its 5-HT(1a) receptor in patients with major depression disorder. J Affect Disord. 2010;124:68–75.
Foley EM, Parkinson JT, Mitchell RE, Turner L, Khandaker GM. Peripheral blood cellular immunophenotype in depression: a systematic review and meta-analysis. Mol Psychiatry. 2023;28:1004–19.
Toben C, Baune BT. An act of balance between adaptive and maladaptive immunity in depression: a role for T lymphocytes. J Neuroimmune Pharmacol. 2015;10:595–609.
Schiweck C, Valles-Colomer M, Arolt V, Muller N, Raes J, Wijkhuijs A, et al. Depression and suicidality: a link to premature T helper cell aging and increased Th17 cells. Brain Behav Immun. 2020;87:603–9.
Grosse L, Hoogenboezem T, Ambree O, Bellingrath S, Jorgens S, de Wit HJ, et al. Deficiencies of the T and natural killer cell system in major depressive disorder: T regulatory cell defects are associated with inflammatory monocyte activation. Brain Behav Immun. 2016;54:38–44.
Snijders G, Schiweck C, Mesman E, Grosse L, De Wit H, Nolen WA, et al. A dynamic course of T cell defects in individuals at risk for mood disorders. Brain Behav Immun. 2016;58:11–7.
Chen Y, Jiang T, Chen P, Ouyang J, Xu G, Zeng Z, et al. Emerging tendency towards autoimmune process in major depressive patients: a novel insight from Th17 cells. Psychiatry Res. 2011;188:224–30.
Suzuki H, Savitz J, Kent Teague T, Gandhapudi SK, Tan C, Misaki M, et al. Altered populations of natural killer cells, cytotoxic T lymphocytes, and regulatory T cells in major depressive disorder: association with sleep disturbance. Brain Behav Immun. 2017;66:193–200.
do Prado CH, Rizzo LB, Wieck A, Lopes RP, Teixeira AL, Grassi-Oliveira R, et al. Reduced regulatory T cells are associated with higher levels of Th1/TH17 cytokines and activated MAPK in type 1 bipolar disorder. Psychoneuroendocrinology. 2013;38:667–76.
Chen Z, Huang Y, Wang B, Peng H, Wang X, Wu H, et al. T cells: an emerging cast of roles in bipolar disorder. Transl Psychiatry. 2023;13:153.
Breunis MN, Kupka RW, Nolen WA, Suppes T, Denicoff KD, Leverich GS, et al. High numbers of circulating activated T cells and raised levels of serum IL-2 receptor in bipolar disorder. Biol Psychiatry. 2003;53:157–65.
Becking K, Haarman BCM, Grosse L, Nolen WA, Claes S, Arolt V, et al. The circulating levels of CD4+ t helper cells are higher in bipolar disorder as compared to major depressive disorder. J Neuroimmunol. 2018;319:28–36.
Keshri N, Nandeesha H, Kattimani S. Elevated interleukin-17 and reduced testosterone in bipolar disorder. Relation with suicidal behaviour. Asian J Psychiatr. 2018;36:66–68.
Barbosa IG, Rocha NP, Assis F, Vieira EL, Soares JC, Bauer ME, et al. Monocyte and lymphocyte activation in bipolar disorder: a new piece in the puzzle of immune dysfunction in mood disorders. Int J Neuropsychopharmacol. 2014;18:pyu021
Wieck A, Grassi-Oliveira R, do Prado CH, Rizzo LB, de Oliveira AS, Kommers-Molina J, et al. Differential neuroendocrine and immune responses to acute psychosocial stress in women with type 1 bipolar disorder. Brain Behav Immun. 2013;34:47–55.
Wu W, Zheng YL, Tian LP, Lai JB, Hu CC, Zhang P, et al. Circulating T lymphocyte subsets, cytokines, and immune checkpoint inhibitors in patients with bipolar II or major depression: a preliminary study. Sci Rep. 2017;7:40530.
Li S, Lv D, Qian C, Jiang J, Zhang P, Xi C, et al. Circulating T-cell subsets discrepancy between bipolar disorder and major depressive disorder during mood episodes: a naturalistic, retrospective study of 1015 cases. CNS Neurosci Ther. 2024;30:e14361.
Swallow VM, Nightingale R, Williams J, Lambert H, Webb NJ, Smith T, et al. Multidisciplinary teams, and parents, negotiating common ground in shared-care of children with long-term conditions: a mixed methods study. BMC Health Serv Res. 2013;13:264.
Bauer ME, Wieck A, Petersen LE, Baptista TS. Neuroendocrine and viral correlates of premature immunosenescence. Ann N Y Acad Sci. 2015;1351:11–21.
Benedetti F, Poletti S, Hoogenboezem TA, Mazza E, Ambrée O, de Wit H, et al. Inflammatory cytokines influence measures of white matter integrity in bipolar disorder. J Affect Disord. 2016;202:1–9.
Comai S, Melloni E, Lorenzi C, Bollettini I, Vai B, Zanardi R, et al. Selective association of cytokine levels and kynurenine/tryptophan ratio with alterations in white matter microstructure in bipolar but not in unipolar depression. Eur Neuropsychopharmacol. 2022;55:96–109.
Poletti S, Myint AM, Schuetze G, Bollettini I, Mazza E, Grillitsch D, et al. Kynurenine pathway and white matter microstructure in bipolar disorder. Eur Arch Psychiatry Clin Neurosci. 2018;268:157–168.
Poletti S, Melloni E, Aggio V, Colombo C, Valtorta F, Benedetti F, et al. Grey and white matter structure associates with the activation of the tryptophan to kynurenine pathway in bipolar disorder. J Affect Disord. 2019;259:404–12.
Bollettini I, Poletti S, Locatelli C, Vai B, Smeraldi E, Colombo C, et al. Disruption of white matter integrity marks poor antidepressant response in bipolar disorder. J Affect Disord. 2015;174:233–40.
Melloni EM, Poletti S, Dallaspezia S, Bollettini I, Vai B, Barbini B, et al. Changes of white matter microstructure after successful treatment of bipolar depression. J Affect Disord. 2020;274:1049–56.
Poletti S, Bollettini I, Mazza E, Locatelli C, Radaelli D, Vai B, et al. Cognitive performances associate with measures of white matter integrity in bipolar disorder. J Affect Disord. 2015;174:342–52.
Poletti S, Aggio V, Brioschi S, Bollettini I, Falini A, Colombo C, et al. Impact of early and recent stress on white matter microstructure in major depressive disorder. J Affect Disord. 2018;225:289–97.
Benedetti F, Bollettini I, Radaelli D, Poletti S, Locatelli C, Falini A, et al. Adverse childhood experiences influence white matter microstructure in patients with bipolar disorder. Psychol Med. 2014;44:3069–82.
Benedetti F, Bollettini I. Recent findings on the role of white matter pathology in bipolar disorder. Harv Rev Psychiatry. 2014;22:338–41.
Vai B, Parenti L, Bollettini I, Cara C, Verga C, Melloni E, et al. Predicting differential diagnosis between bipolar and unipolar depression with multiple kernel learning on multimodal structural neuroimaging. Eur Neuropsychopharmacol. 2020;34:28–38.
Arnett HA, Mason J, Marino M, Suzuki K, Matsushima GK, Ting JP. TNF alpha promotes proliferation of oligodendrocyte progenitors and remyelination. Nat Neurosci. 2001;4:1116–22.
McGeachy MJ, Bak-Jensen KS, Chen Y, Tato CM, Blumenschein W, McClanahan T, et al. TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat Immunol. 2007;8:1390–7.
Imeri L, Opp MR. How (and why) the immune system makes us sleep. Nat Rev Neurosci. 2009;10:199–210.
Wisor JP, Schmidt MA, Clegern WC. Evidence for neuroinflammatory and microglial changes in the cerebral response to sleep loss. Sleep. 2011;34:261–72.
Muzio L, Brambilla V, Calcaterra L, D’Adamo P, Martino G, Benedetti F. Increased neuroplasticity and hippocampal microglia activation in a mice model of rapid antidepressant treatment. Behav Brain Res. 2016;311:392–402.
Alesci S, Martinez PE, Kelkar S, Ilias I, Ronsaville DS, Listwak SJ, et al. Major depression is associated with significant diurnal elevations in plasma interleukin-6 levels, a shift of its circadian rhythm, and loss of physiological complexity in its secretion: clinical implications. J Clin Endocrinol Metab. 2005;90:2522–30.
Bonkat G, Wirz D, Rieken M, Gasser TC, Bachmann A, Braissant O. [Areas of application of isothermal microcalorimetry in urology: an overview]. Urol A. 2013;52:1092–6.
Melloni EM, Bravi B, Poletti S, Dallaspezia S, Barbini B, Zanardi R, et al. Antidepressant chronotherapeutics normalizes prefrontal 1H-MRS glutamate in bipolar depression. Prog Neuropsychopharmacol Biol Psychiatry. 2022;119:110606.
McClung CA. How might circadian rhythms control mood? Let me count the ways. Biol Psychiatry. 2013;74:242–9.
Wirz-Justice A, Benedetti F. Perspectives in affective disorders: Clocks and sleep. Eur J Neurosci. 2020;51:346–365.
Walker WH 2nd, Walton JC, DeVries AC, Nelson RJ. Circadian rhythm disruption and mental health. Transl Psychiatry. 2020;10:28.
Branchi I, Poggini S, Capuron L, Benedetti F, Poletti S, Tamouza R, et al. Brain-immune crosstalk in the treatment of major depressive disorder. Eur Neuropsychopharmacol. 2021;45:89–107.
Arteaga-Henriquez G, Simon MS, Burger B, Weidinger E, Wijkhuijs A, Arolt V, et al. Low-grade inflammation as a predictor of antidepressant and anti-inflammatory therapy response in MDD patients: a systematic review of the literature in combination with an analysis of experimental data collected in the EU-MOODINFLAME Consortium. Front Psychiatry. 2019;10:458.
Benedetti F, Poletti S, Vai B, Mazza MG, Lorenzi C, Brioschi S, et al. Higher baseline interleukin-1β and TNF-α hamper antidepressant response in major depressive disorder. Eur Neuropsychopharmacol. 2021;42:35–44.
Benedetti F, Zanardi R, Mazza MG. Antidepressant psychopharmacology: is inflammation a future target? Int Clin Psychopharmacol. 2022;37:79–81.
Poggini S, Lopez MB, Albanese NC, Golia MT, Ibáñez FG, Limatola C, et al. Minocycline treatment improves cognitive and functional plasticity in a preclinical mouse model of major depressive disorder. Behav Brain Res. 2023;441:114295
Cheng D, Qin Z-S, Zheng Y, Xie J-Y, Liang S-S, Zhang J-L, et al. Minocycline, a classic antibiotic, exerts psychotropic effects by normalizing microglial neuroinflammation-evoked tryptophan-kynurenine pathway dysregulation in chronically stressed male mice. Brain Behav Immun. 2023;107:305–18.
Nettis MA, Lombardo G, Hastings C, Zajkowska Z, Mariani N, Nikkheslat N, et al. Augmentation therapy with minocycline in treatment-resistant depression patients with low-grade peripheral inflammation: results from a double-blind randomised clinical trial. Neuropsychopharmacology. 2021;46:939–48.
Poletti S, Zanardi R, Mandelli A, Aggio V, Finardi A, Lorenzi C, et al. Low-dose interleukin 2 antidepressant potentiation in unipolar and bipolar depression: safety, efficacy, and immunological biomarkers. Brain Behav Immun. 2024;118:52–68.
Beishuizen A, Thijs LG. Endotoxin and the hypothalamo-pituitary-adrenal (HPA) axis. J Endotoxin Res. 2003;9:3–24.
Silverman MN, Miller AH, Biron CA, Pearce BD. Characterization of an interleukin-6- and adrenocorticotropin-dependent, immune-to-adrenal pathway during viral infection. Endocrinology. 2004;145:3580–9.
Turrin NP, Rivest S. Unraveling the molecular details involved in the intimate link between the immune and neuroendocrine systems. Exp Biol Med. 2004;229:996–1006.
Pariante CM. Why are depressed patients inflamed? A reflection on 20 years of research on depression, glucocorticoid resistance and inflammation. Eur Neuropsychopharmacol. 2017;27:554–9.
Pace TW, Miller AH. Cytokines and glucocorticoid receptor signaling. Relevance to major depression. Ann N Y Acad Sci. 2009;1179:86–105.
Wake H, Moorhouse AJ, Jinno S, Kohsaka S, Nabekura J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J Neurosci. 2009;29:3974–80.
Eyo UB, Wu LJ. Bidirectional microglia-neuron communication in the healthy brain. Neural Plast. 2013;2013:456857.
Harry GJ, Kraft AD. Microglia in the developing brain: a potential target with lifetime effects. Neurotoxicology. 2012;33:191–206.
Lindenau J, Noack H, Asayama K, Wolf G. Enhanced cellular glutathione peroxidase immunoreactivity in activated astrocytes and in microglia during excitotoxin induced neurodegeneration. Glia. 1998;24:252–6.
Paolicelli RC, Sierra A, Stevens B, Tremblay M-E, Aguzzi A, Ajami B, et al. Microglia states and nomenclature: a field at its crossroads. Neuron. 2022;110:3458–83.
Machado-Vieira R, Soeiro-De-Souza MG, Richards EM, Teixeira AL, Zarate CA Jr. Multiple levels of impaired neural plasticity and cellular resilience in bipolar disorder: developing treatments using an integrated translational approach. World J Biol Psychiatry. 2014;15:84–95.
Paradise MB, Naismith SL, Norrie LM, Graeber MB, Hickie IB. The role of glia in late-life depression. Int Psychogeriatr. 2012;24:1878–90.
Park KM, Bowers WJ. Tumor necrosis factor-alpha mediated signaling in neuronal homeostasis and dysfunction. Cell Signal. 2010;22:977–83.
Scott G, Zetterberg H, Jolly A, Cole JH, De Simoni S, Jenkins PO, et al. Minocycline reduces chronic microglial activation after brain trauma but increases neurodegeneration. Brain. 2018;141:459–71.
Enache D, Pariante CM, Mondelli V. Markers of central inflammation in major depressive disorder: a systematic review and meta-analysis of studies examining cerebrospinal fluid, positron emission tomography and post-mortem brain tissue. Brain Behav Immun. 2019;81:24–40.
Stertz L, Magalhaes PV, Kapczinski F. Is bipolar disorder an inflammatory condition? The relevance of microglial activation. Curr Opin Psychiatry. 2013;26:19–26.
Haarman BCB, Riemersma-Van der Lek RF, de Groot JC, Ruhé HGE, Klein HC, Zandstra TE, et al. Neuroinflammation in bipolar disorder–A [11C]-(R)-PK11195 positron emission tomography study. Brain Behav Immun. 2014;40:219–25.
Li H, Sagar AP, Kéri S. Microglial markers in the frontal cortex are related to cognitive dysfunctions in major depressive disorder. J Affect Disord. 2018;241:305–10.
Setiawan E, Wilson AA, Mizrahi R, Rusjan PM, Miler L, Rajkowska G, et al. Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes. JAMA Psychiatry. 2015;72:268–75.
Holmes SE, Hinz R, Conen S, Gregory CJ, Matthews JC, Anton-Rodriguez JM, et al. Elevated translocator protein in anterior cingulate in major depression and a role for inflammation in suicidal thinking: a positron emission tomography study. Biol Psychiatry. 2018;83:61–9.
Steiner J, Bielau H, Brisch R, Danos P, Ullrich O, Mawrin C, et al. Immunological aspects in the neurobiology of suicide: elevated microglial density in schizophrenia and depression is associated with suicide. J Psychiatr Res. 2008;42:151–7.
Bourgognon JM, Cavanagh J. The role of cytokines in modulating learning and memory and brain plasticity. Brain Neurosci Adv. 2020;4:2398212820979802.
Zhang JC, Yao W, Hashimoto K. Brain-derived neurotrophic factor (BDNF)-TrkB signaling in inflammation-related depression and potential therapeutic targets. Curr Neuropharmacol. 2016;14:721–31.
Felger JC, Lotrich FE. Inflammatory cytokines in depression: neurobiological mechanisms and therapeutic implications. Neuroscience. 2013;246:199–229.
Schäfer BW, Heizmann CW. The S100 family of EF-hand calcium-binding proteins: functions and pathology. Trends in biochemical sciences. 1996;21:134–40.
Schroeter ML, Sacher J, Steiner J, Schoenknecht P, Mueller K. Serum S100B represents a new biomarker for mood disorders. Curr Drug Targets. 2013;14:1237–48.
Capuron L, Neurauter G, Musselman DL, Lawson DH, Nemeroff CB, Fuchs D, et al. Interferon-alpha-induced changes in tryptophan metabolism. relationship to depression and paroxetine treatment. Biol Psychiatry. 2003;54:906–14.
Capuron L, Ravaud A, Gualde N, Bosmans E, Dantzer R, Maes M, et al. Association between immune activation and early depressive symptoms in cancer patients treated with interleukin-2-based therapy. Psychoneuroendocrinology. 2001;26:797–808.
Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9:46–56.
Davis I, Liu A. What is the tryptophan kynurenine pathway and why is it important to neurotherapeutics? Expert Rev Neurother. 2015;15:719–21.
Wang J, Dunn AJ. Mouse interleukin-6 stimulates the HPA axis and increases brain tryptophan and serotonin metabolism. Neurochem Int. 1998;33:143–54.
Zhang J, Terreni L, De Simoni MG, Dunn AJ. Peripheral interleukin-6 administration increases extracellular concentrations of serotonin and the evoked release of serotonin in the rat striatum. Neurochem Int. 2001;38:303–8.
Arango V, Underwood MD, Mann JJ. Serotonin brain circuits involved in major depression and suicide. Prog Brain Res. 2002;136:443–53.
Barton DA, Esler MD, Dawood T, Lambert EA, Haikerwal D, Brenchley C, et al. Elevated brain serotonin turnover in patients with depression: effect of genotype and therapy. Arch Gen Psychiatry. 2008;65:38–46.
Pandey GN, Dwivedi Y, Rizavi HS, Ren X, Pandey SC, Pesold C, et al. Higher expression of serotonin 5-HT2A receptors in the postmortem brains of teenage suicide victims. Am J Psychiatry. 2002;159:419–29.
Erhardt S, Schwieler L, Imbeault S, Engberg G. The kynurenine pathway in schizophrenia and bipolar disorder. Neuropharmacology. 2017;112:297–306.
Myint AM, Kim YK, Verkerk R, Scharpe S, Steinbusch H, Leonard B. Kynurenine pathway in major depression: evidence of impaired neuroprotection. J Affect Disord. 2007;98:143–51.
Benedetti F, Palladini M, Paolini M, Melloni E, Vai B, De Lorenzo R, et al. Brain correlates of depression, post-traumatic distress, and inflammatory biomarkers in COVID-19 survivors: a multimodal magnetic resonance imaging study. Brain Behav Immun Health. 2021;18:100387.
Funding
European Union H2020 Grant 754740 “MOODSTRATIFICATION: Immune Signatures for Therapy Stratification in Major Mood Disorders”; Italian Ministry of Health, RF-2018-12367249; Ministry of Education, University and Research 201779W93T_005.
Author information
Authors and Affiliations
Contributions
SP and MGM led the review process, conducting literature searches, synthesizing findings, and producing a draft version of the manuscript. FB contributed to the review process by providing additional insights and contributing to manuscript revisions. SP conceived the idea and structure for the article, directed the overall process, and provided guidance throughout. All authors participated in editing and reviewing, and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Poletti, S., Mazza, M.G. & Benedetti, F. Inflammatory mediators in major depression and bipolar disorder. Transl Psychiatry 14, 247 (2024). https://doi.org/10.1038/s41398-024-02921-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-024-02921-z
- Springer Nature Limited