

Abstract
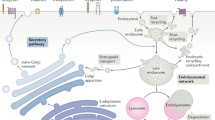
Recycling of endocytic receptors to the cell surface involves passage through a series of membrane-bound compartments by mechanisms that are poorly understood. In particular, it is unknown if endocytic recycling requires the function of multisubunit tethering complexes, as is the case for other intracellular trafficking pathways. Herein we describe a tethering complex named endosome-associated recycling protein (EARP) that is structurally related to the previously described Golgi-associated retrograde protein (GARP) complex. The two complexes share the Ang2, Vps52 and Vps53 subunits, but EARP contains an uncharacterized protein, syndetin, in place of the Vps54 subunit of GARP. This change determines differential localization of EARP to recycling endosomes and GARP to the Golgi complex. EARP interacts with the target SNARE syntaxin 6 and various cognate SNAREs. Depletion of syndetin or syntaxin 6 delays recycling of internalized transferrin to the cell surface. These findings implicate EARP in canonical membrane-fusion events in the process of endocytic recycling.








Similar content being viewed by others
References
Henne, W. M., Buchkovich, N. J. & Emr, S. D. The ESCRT pathway. Dev. Cell 21, 77–91 (2011).
Bonifacino, J. S. & Rojas, R. Retrograde transport from endosomes to the trans-Golgi network. Nat. Rev. Mol. Cell Biol. 7, 568–579 (2006).
Leto, D. & Saltiel, A. R. Regulation of glucose transport by insulin: traffic control of GLUT4. Nat. Rev. Mol. Cell Biol. 13, 383–396 (2012).
Raposo, G. & Marks, M. S. Melanosomes–dark organelles enlighten endosomal membrane transport. Nat. Rev. Mol. Cell Biol. 8, 786–797 (2007).
Maxfield, F. R. & McGraw, T. E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 5, 121–132 (2004).
Johannes, L. & Popoff, V. Tracing the retrograde route in protein trafficking. Cell 135, 1175–1187 (2008).
Cullen, P. J. & Korswagen, H. C. Sorting nexins provide diversity for retromer-dependent trafficking events. Nat. Cell Biol. 14, 29–37 (2012).
Seaman, M. N., McCaffery, J. M. & Emr, S. D. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J. Cell Biol. 142, 665–681 (1998).
Seaman, M. N. Cargo-selective endosomal sorting for retrieval to the Golgi requires retromer. J. Cell Biol. 165, 111–122 (2004).
Arighi, C. N., Hartnell, L. M., Aguilar, R. C., Haft, C. R. & Bonifacino, J. S. Role of the mammalian retromer in sorting of the cation-independent mannose 6-phosphate receptor. J. Cell Biol. 165, 123–133 (2004).
Mallard, F. et al. Early/recycling endosomes-to-TGN transport involves two SNARE complexes and a Rab6 isoform. J. Cell Biol. 156, 653–664 (2002).
Bonifacino, J. S. & Hierro, A. Transport according to GARP: receiving retrograde cargo at the trans-Golgi network. Trends Cell Biol. 21, 159–167 (2011).
Geuze, H. J., Slot, J. W., Strous, G. J., Lodish, H. F. & Schwartz, A. L. Intracellular site of asialoglycoprotein receptor-ligand uncoupling: double-label immunoelectron microscopy during receptor-mediated endocytosis. Cell 32, 277–287 (1983).
Grant, B. D. & Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 10, 597–608 (2009).
Van der Sluijs, P. et al. The small GTP-binding protein rab4 controls an early sorting event on the endocytic pathway. Cell 70, 729–740 (1992).
Sonnichsen, B., De Renzis, S., Nielsen, E., Rietdorf, J. & Zerial, M. Distinct membrane domains on endosomes in the recycling pathway visualized by multicolor imaging of Rab4, Rab5, and Rab11. J. Cell Biol. 149, 901–914 (2000).
Ullrich, O., Reinsch, S., Urbe, S., Zerial, M. & Parton, R. G. Rab11 regulates recycling through the pericentriolar recycling endosome. J. Cell Biol. 135, 913–924 (1996).
Ren, M. et al. Hydrolysis of GTP on rab11 is required for the direct delivery of transferrin from the pericentriolar recycling compartment to the cell surface but not from sorting endosomes. Proc. Natl Acad. Sci. USA 95, 6187–6192 (1998).
Yamashiro, D. J., Tycko, B., Fluss, S. R. & Maxfield, F. R. Segregation of transferrin to a mildly acidic (pH 6.5) para-Golgi compartment in the recycling pathway. Cell 37, 789–800 (1984).
Brandhorst, D. et al. Homotypic fusion of early endosomes: SNAREs do not determine fusion specificity. Proc. Natl Acad. Sci. USA 103, 2701–2706 (2006).
Tiwari, A. et al. Endothelial cell migration on fibronectin is regulated by syntaxin 6-mediated α5β1 integrin recycling. J. Biol. Chem. 286, 36749–36761 (2011).
Riggs, K. A. et al. Regulation of integrin endocytic recycling and chemotactic cell migration by syntaxin 6 and VAMP3 interaction. J. Cell Sci. 125, 3827–3839 (2012).
Watson, R. T., Hou, J. C. & Pessin, J. E. Recycling of IRAP from the plasma membrane back to the insulin-responsive compartment requires the Q-SNARE syntaxin 6 but not the GGA clathrin adaptors. J. Cell Sci. 121, 1243–1251 (2008).
Proctor, K. M., Miller, S. C., Bryant, N. J. & Gould, G. W. Syntaxin 16 controls the intracellular sequestration of GLUT4 in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 347, 433–438 (2006).
Gee, H. Y., Tang, B. L., Kim, K. H. & Lee, M. G. Syntaxin 16 binds to cystic fibrosis transmembrane conductance regulator and regulates its membrane trafficking in epithelial cells. J. Biol. Chem. 285, 35519–35527 (2010).
Bose, A. et al. The v-SNARE Vti1a regulates insulin-stimulated glucose transport and Acrp30 secretion in 3T3-L1 adipocytes. J. Biol. Chem. 280, 36946–36951 (2005).
Kreykenbohm, V., Wenzel, D., Antonin, W., Atlachkine, V. & von Mollard, G. F. The SNAREs vti1a and vti1b have distinct localization and SNARE complex partners. Eur. J. Cell Biol. 81, 273–280 (2002).
McMahon, H. T. et al. Cellubrevin is a ubiquitous tetanus-toxin substrate homologous to a putative synaptic vesicle fusion protein. Nature 364, 346–349 (1993).
Galli, T. et al. Tetanus toxin-mediated cleavage of cellubrevin impairs exocytosis of transferrin receptor-containing vesicles in CHO cells. J. Cell Biol. 125, 1015–1024 (1994).
Daro, E., van der Sluijs, P., Galli, T. & Mellman, I. Rab4 and cellubrevin define different early endosome populations on the pathway of transferrin receptor recycling. Proc. Natl Acad. Sci. USA 93, 9559–9564 (1996).
Prekeris, R., Klumperman, J., Chen, Y. A. & Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. J. Cell Biol. 143, 957–971 (1998).
Trischler, M., Stoorvogel, W. & Ullrich, O. Biochemical analysis of distinct Rab5- and Rab11-positive endosomes along the transferrin pathway. J. Cell Sci. 112, 4773–4783 (1999).
Lee, S. H., Valtschanoff, J. G., Kharazia, V. N., Weinberg, R. & Sheng, M. Biochemical and morphological characterization of an intracellular membrane compartment containing AMPA receptors. Neuropharmacology 41, 680–692 (2001).
Hoogenraad, C. C. et al. Neuron specific Rab4 effector GRASP-1 coordinates membrane specialization and maturation of recycling endosomes. PLoS Biol. 8, e1000283 (2010).
Soding, J., Biegert, A. & Lupas, A. N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 33, W244-8 (2005).
Yu, I. M. & Hughson, F. M. Tethering factors as organizers of intracellular vesicular traffic. Annu. Rev. Cell Dev. Biol. 26, 137–156 (2010).
Kelley, L. A. & Sternberg, M. J. Protein structure prediction on the Web: a case study using the Phyre server. Nat. Protoc. 4, 363–371 (2009).
Walker, G., Dorrell, R. G., Schlacht, A. & Dacks, J. B. Eukaryotic systematics: a user’s guide for cell biologists and parasitologists. Parasitology 138, 1638–1663 (2011).
Perez-Victoria, F. J. et al. Ang2/Fat-free is a conserved subunit of the Golgi-associated Retrograde Protein (GARP) complex. Mol. Biol. Cell 21, 3386–3395 (2010).
Gorvel, J. P., Chavrier, P., Zerial, M. & Gruenberg, J. rab5 controls early endosome fusion in vitro. Cell 64, 915–925 (1991).
Bucci, C. et al. The small GTPase rab5 functions as a regulatory factor in the early endocytic pathway. Cell 70, 715–728 (1992).
Perez-Victoria, F. J., Mardones, G. A. & Bonifacino, J. S. Requirement of the human GARP complex for mannose 6-phosphate-receptor-dependent sorting of cathepsin D to lysosomes. Mol. Biol. Cell 19, 2350–2362 (2008).
Perez-Victoria, F. J. & Bonifacino, J. S. Dual roles of the mammalian GARP complex in tethering and SNARE complex assembly at the trans-golgi network. Mol. Cell. Biol. 29, 5251–5263 (2009).
Perez-Victoria, F. J. et al. Structural basis for the wobbler mouse neurodegenerative disorder caused by mutation in the Vps54 subunit of the GARP complex. Proc. Natl Acad. Sci. USA 107, 12860–12865 (2010).
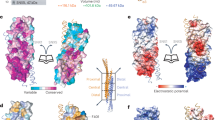
Abascal-Palacios, G., Schindler, C., Rojas, A. L., Bonifacino, J. S. & Hierro, A. Structural basis for the interaction of the Golgi-Associated Retrograde Protein Complex with the t-SNARE Syntaxin 6. Structure 21, 1698–1706 (2013).
Bock, J. B., Klumperman, J., Davanger, S. & Scheller, R. H. Syntaxin 6 functions in trans-Golgi network vesicle trafficking. Mol. Biol. Cell 8, 1261–1271 (1997).
Hutt, D. M., Baltz, J. M. & Ngsee, J. K. Synaptotagmin VI and VIII and syntaxin 2 are essential for the mouse sperm acrosome reaction. J. Biol. Chem. 280, 20197–20203 (2005).
Kennedy, M. J., Davison, I. G., Robinson, C. G. & Ehlers, M. D. Syntaxin-4 defines a domain for activity-dependent exocytosis in dendritic spines. Cell 141, 524–535 (2010).
Hong, W. & Lev, S. Tethering the assembly of SNARE complexes. Trends Cell Biol. 24, 35–43 (2014).
Wendler, F. & Tooze, S. Syntaxin 6: the promiscuous behaviour of a SNARE protein. Traffic 2, 606–611 (2001).
Moreau, D. et al. Genome-wide RNAi screens identify genes required for Ricin and PE intoxications. Dev. Cell 21, 231–244 (2011).
Bassik, M. C. et al. A systematic mammalian genetic interaction map reveals pathways underlying ricin susceptibility. Cell 152, 909–922 (2013).
Feinstein, M. et al. VPS53 mutations cause progressive cerebello-cerebral atrophy type 2 (PCCA2). J. Med. Genet. 51, 303–308 (2014).
Farias, G. G. et al. Signal-mediated, AP-1/clathrin-dependent sorting of transmembrane receptors to the somatodendritic domain of hippocampal neurons. Neuron 75, 810–823 (2012).
Prabhu, Y. et al. Adaptor protein 2-mediated endocytosis of the β-secretase BACE1 is dispensable for amyloid precursor protein processing. Mol. Biol. Cell 23, 2339–2351 (2012).
Chen, Y. et al. Rab10 and myosin-Va mediate insulin-stimulated GLUT4 storage vesicle translocation in adipocytes. J. Cell Biol. 198, 545–560 (2012).
Li, W. P., Liu, P., Pilcher, B. K. & Anderson, R. G. Cell-specific targeting of caveolin-1 to caveolae, secretory vesicles, cytoplasm or mitochondria. J. Cell Sci. 114, 1397–1408 (2001).
Kaech, S. & Banker, G. Culturing hippocampal neurons. Nat. Protoc. 1, 2406–2415 (2006).
Gloeckner, C. J., Boldt, K. & Ueffing, M. Strep/FLAG tandem affinity purification (SF-TAP) to study protein interactions. Curr. Protoc. Protein Sci. 57, 19.20.1–19.20.19 (2009).
Ghosh, R. N., Gelman, D. L. & Maxfield, F. R. Quantification of low density lipoprotein and transferrin endocytic sorting HEp2 cells using confocal microscopy. J. Cell Sci. 107, 2177–2189 (1994).
Burgos, P. V. et al. Sorting of the Alzheimer’s disease amyloid precursor protein mediated by the AP-4 complex. Dev. Cell 18, 425–436 (2010).
Rojas, R. et al. Regulation of retromer recruitment to endosomes by sequential action of Rab5 and Rab7. J. Cell Biol. 183, 513–526 (2008).
Combet, C., Blanchet, C., Geourjon, C. & Deleage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 25, 147–150 (2000).
Acknowledgements
We thank X. Zhu and N. Tsai for technical assistance, J. Presley for discussions and R. Mattera and D. Gershlick for critical review of the manuscript. This work was funded by the Intramural Program of NICHD, NIH (ZIA HD001607).
Author information
Authors and Affiliations
Contributions
C.S., Y.C. and J.S.B. conceived the project. C.S. and Y.C. carried out most of the experiments. J.P. contributed to SNARE pulldowns and X.G. to subcellular fractionation experiments. C.S., Y.C., J.P., X.G. and J.S.B. analysed the data. J.S.B., C.S. and Y.C. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Phylogenetic analysis of Syndetin orthologs.
An initial BLASTP search revealed that the first 492 amino acids of human Syndetin comprised the largest conserved domain (DUF 2450). This sequence was used in two consecutive rounds of PSI-BLAST on genomes from different organisms. Orthologs were defined as (i) being large proteins (>500 amino acids in all super-groups, with the exception of some smaller proteins in the SAR/CCTH super-group), (ii) having a high α-helical content, (iii) having either DUF 2450 or DUF 2451 domains, and (iv) yielding human Syndetin and not Vps54 as one of three top hits in reverse PSI-BLAST searches. Listed are representative species, NCBI reference sequences and length of the Syndetin orthologs.
Supplementary Figure 2 Vps54 and Syndetin exhibit distinct organelle targeting and retrograde transport activities.
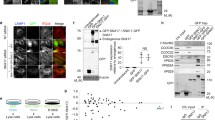
(a) Co-localization of Vps54-EGFP and Syndetin-EGFP with Ang2–13Myc in rat hippocampal neurons analyzed with antibodies to GFP, the myc epitope and MAP2 as described in the legend to Figure 3. Bars, 10 μm. (b) Syndetin KD does not affect retrograde transport from endosomes to the TGN. Control (mock), Ang2-, Vps54- and Syndetin-KD HeLa cells were treated with 0.5 μg/ml Cy3-conjugated B-subunit of Shiga toxin (STxB) at 37 °C for 15 min and chased for 1 h before fixation and immunostaining. Nocodazole treatment was applied at 10 μM for 2 h following the chase where indicated. Giantin (medial Golgi marker) and TGN46 (TGN marker) were visualized by immunostaining with rabbit antibody to Giantin and sheep antibody to human TGN46 followed by Alexa647 donkey anti-rabbit and Alexa488 donkey anti-sheep antibodies. Bars, 10 μm.
Supplementary Figure 3 Syndetin KD does not affect initial Tf uptake but increases Tf accumulation over time.
(a,b) HeLa cells were treated with the indicated siRNAs and incubated with Tf-Alexa568 at 37 °C for 3 min. After three washes with ice-cold PBS, cells were fixed. Images were acquired by confocal microscopy and quantified using ImageJ. Values are the mean ± SEM (n = 3 independent experiments). The numbers of cells quantified were: mock, 171; Vps54 KD, 167; Syndetin KD, 142. n.s., P = 0.86 (Mock vs. Vps54 KD) and P = 0.69 (Mock vs. Syndetin KD). (c) HeLa cells treated with the indicated siRNAs were incubated with Tf-Alexa647 at 37 °C for different times and then washed twice with ice-cold citrate buffer (pH 4.6) and three times with PBS to remove surface-bound Tf-Alexa647. Intracellular Tf-Alexa647 was quantified by FACS after detaching cells from the plates using 2 mM EDTA. At least 1x105 cells were analyzed for each group in each independent experiment. Values are the mean ± SEM (n = 3 independent experiments).
Supplementary Figure 4 Effects of Syndetin KD on the distribution of internalized Tf-biotin in subcellular fractions and on LC3-II levels.
(a) Equal numbers of HeLa cells treated with control (mock) or Syndetin siRNAs were allowed to internalize Tf-biotin for 30 min. After stripping surface-bound Tf-biotin with citrate buffer (pH 4.6), cells were chased for 30 min. Cells were lysed and fractionated by ultracentrifugation on 5–20% iodixanol gradients. Fractions were collected and analyzed by SDS-PAGE gels and immunoblotting for the indicated proteins. Tf-biotin was detected with Streptavidin-HRP polymer according to the manufacturer’s instructions. Molecular mass markers (in kDa) are indicated at left. (b) HeLa cells were treated with siRNAs targeting the proteins indicated on top and analyzed by SDS-PAGE and blotting for the proteins indicated at right. Bafilomycin A1 was applied at 250 nM for 4 h. Molecular mass markers (in kDa) are indicated at left. Uncropped images of the blots are shown in Supplementary Fig. 5c.
Supplementary Figure 5 Uncropped images of key panels in main figures.
Red boxes indicate the cropped portion of each immunoblot presented in the corresponding main figures. Ladder of molecular markers is shown on the left of each panel.
Supplementary information
Supplementary Information
Supplementary Information (PDF 1056 kb)
Differential localization of Vps54 and Syndetin in live H4 cells.
H4 cells stably expressing Vps54-EGFP were transfected with a plasmid encoding Syndetin-mCherry and imaged by live-cell confocal microscopy after 48 h. (MOV 35467 kb)
Co-localization of Syndetin with Rab4A observed by TIRF microscopy of live rat hippocampal neurons.
Rat hippocampal neurons were co-transfected with plasmids encoding Syndetin-EGFP and TagRFP-Rab4A on DIV-3 and examined by TIRF microscopy on DIV-7. Inset shows magnifications of the areas boxed with solid lines. (MOV 37196 kb)
Co-localization of Syndetin with Rab4A observed by confocal microscopy of live HeLa cells.
Syndetin-KD HeLa cells were transiently transfected with plasmids encoding mouse Syndetin-EGFP and TagRFP-Rab4A and imaged at 37 °C after 48 h. (MOV 35828 kb)
Low co-localization of Syndetin with Rab5A observed by confocal microscopy of live HeLa cells.
Syndetin-KD HeLa cells were transiently transfected with plasmids encoding mouse Syndetin-EGFP and TagRFP-Rab5A and imaged at 37 °C after 48 h. (MOV 36558 kb)
Rapid access of internalized Tf into the Syndetin compartment.
Syndetin-KD HeLa cells were transiently transfected with plasmids encoding Syndetin-4xEGFP and, after 48 h, imaged at after addition of Tf-Alexa568 to the medium. (MOV 1136 kb)
Rights and permissions
About this article

Cite this article
Schindler, C., Chen, Y., Pu, J. et al. EARP is a multisubunit tethering complex involved in endocytic recycling. Nat Cell Biol 17, 639–650 (2015). https://doi.org/10.1038/ncb3129
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3129
- Springer Nature Limited
This article is cited by
-
EMP3 sustains oncogenic EGFR/CDK2 signaling by restricting receptor degradation in glioblastoma
Acta Neuropathologica Communications (2023)
-
Involvement of a BH3-only apoptosis sensitizer gene Blm-s in hippocampus-mediated mood control
Translational Psychiatry (2022)
-
The rare mutation in the endosome-associated recycling protein gene VPS50 is associated with human neural tube defects
Molecular Cytogenetics (2019)
-
SNAREs define targeting specificity of trafficking vesicles by combinatorial interaction with tethering factors
Nature Communications (2019)
-
Amino acids stimulate the endosome-to-Golgi trafficking through Ragulator and small GTPase Arl5
Nature Communications (2018)