
Abstract
The etiology of disease pathogenesis can be largely explained by genetic variations and several types of environmental factors. In genetically disease-susceptible individuals, subsequent environmental triggers may induce disease development. The human body is colonized by complex commensal microbes that have co-evolved with the host immune system. With the adaptation to modern lifestyles, its composition has changed depending on host genetics, changes in diet, overuse of antibiotics against infection and elimination of natural enemies through the strengthening of sanitation. In particular, commensal microbiota is necessary in the development, induction and function of T cells to maintain host immune homeostasis. Alterations in the compositional diversity and abundance levels of microbiota, known as dysbiosis, can trigger several types of autoimmune and inflammatory diseases through the imbalance of T-cell subpopulations, such as Th1, Th2, Th17 and Treg cells. Recently, emerging evidence has identified that dysbiosis is involved in the progression of rheumatoid arthritis, type 1 and 2 diabetic mellitus, and asthma, together with dysregulated T-cell subpopulations. In this review, we will focus on understanding the complicated microbiota-T-cell axis between homeostatic and pathogenic conditions and elucidate important insights for the development of novel targets for disease therapy.
Similar content being viewed by others


Overview of T-cell homeostasis
T cells are major immune cells for defending hosts as well as controlling the development of immune-mediated inflammatory diseases.1, 2 Naive T cells from the thymus migrate to secondary lymphoid tissues, in which they encounter antigens by interacting with antigen–MHC complexes and then become activated and differentiated into effector T cells. Effector T cells proliferate and produce numerous effector molecules, including pro-/anti-inflammatory cytokines and cytotoxic molecules, to protect the host from various pathogenic microorganisms. After the antigen is cleared, most effector T cells undergo programmed cell death, but some survive and differentiate into memory T cells. In this process, T-helper cells (Th cells) help B cells make antibodies and promote cytotoxic T-cell function. Effector Th cells can be classified into Th1, Th2, Th17 and regulatory T (Treg) subpopulations based on their unique cytokine properties.2 For example, whereas Th1 cells produce IFNγ to promote cellular immune responses against intracellular microorganism infection, Th2 cells produce IL-4, IL-5 and IL-13 to promote humoral immune responses against parasites and allergens.1, 2
Over the last two decades, Th17-cell subpopulation has been identified and studied by several research groups.3 Th17 cells produce the potent pro-inflammatory cytokine IL-17, which leads to tissue injury and is involved in the pathogenesis of inflammatory and autoimmune diseases.4 In contrast, Treg cells produce an anti-inflammatory cytokine, IL-10, to suppress excessive immune responses to protect the host.2 In other words, Treg cells play indispensable, central roles in maintaining immune tolerance to self-antigens and facilitating tissue repair. Several reports have proposed that Treg cells include thymus-derived Treg (tTreg, Helios+ and GATA3+) cells and peripherally derived Treg (pTreg, Helios− and RORγt+) cells, the latter of which are induced by the peripheral tissue microenvironment and function in both peripheral and mucosal homeostasis.5
In addition to classical Th-cell subpopulations, T-follicular helper (Tfh) cells and Th9 cells have recently been defined as new subpopulations that produce IL-21 and IL-9, respectively. Tfh cells help B cells differentiate into antibody-producing cells mainly in the germinal center and are involved in autoimmune diseases.6 Although the function and importance of Th9 cells is difficult to clarify in humans, IL-9 is known to promote mast cell and T-cell growth as well as class-switching to IgE in B cells.7 On the basis of these functions of IL-9, Th9 cells may be a unique subpopulation in specific murine models of disease, such as asthma, helminth infections and other autoimmune diseases.7 However, Tfh and Th9-cell subpopulations need to be clarified in greater detail as unique subpopulations upon specific pathological conditions. Overall, Th-cell polarization and subsequent functions may be determined by the cytokine environment under specific conditions. Well-balanced Th-cell differentiation is important in preserving a healthy condition. However, alterations in Th-cell differentiation occur in specific environmental conditions, and these alterations could disrupt the immune system balance, leading to the development of diseases. In particular, Treg cell deficiency enhances autoimmune and inflammation responses.
T-cell homeostasis and the microbiota
The etiology of disease pathogenesis can be largely explained by genetic variations and several types of environmental factors, such as diet, smoking and infection. In genetically disease-susceptible individuals, the subsequent environmental factors may induce the initiation of diseases. The human body is continuously exposed to environmental microorganisms and is colonized by a variety of microbes, known collectively as the microbiota, that comprise an ecological community of commensal, symbiotic and pathogenic microorganisms.8, 9 With the development of culture-independent modern techniques for detecting microorganisms, such as 16S rRNA–DNA sequencing and whole-genome shotgun sequencing, ~1200 different bacterial species have been commonly identified in the human gut microbiota, the majority of which is composed of five phyla: Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria and Verrucomicrobia.9, 10 Of interest, ~160 distinct bacterial species have been found to differ in each individual, and the relative abundance of common bacterial phyla is also different in each individual.10 The composition and abundance of the microbiota may be affected beginning early in life by such factors as genetics, prenatal environment, diet and antibiotic treatment.11
The host immune system has co-evolved with the microbiota populating the human body, and this microbiota provides many beneficial functions to its host, including synthesizing nutrients, protecting against invasion by pathogens and regulating immune responses to self-antigens.12, 13, 14 This suggests that the microbiota plays a fundamental role in the balance of the host immune system between activation and tolerance. In other words, disruption of this relationship could contribute to the development of diseases. Among the various immune cells, the microbiota has been shown to be associated with the development of Th1, Th2, Th17 and Treg cells.15, 16, 17 Relatedly, germ-free (GF) animal models that have never been exposed to any microorganisms have revealed the importance of the microbiota in the development of the host immune system.18 GF mice have defects in the spleen and mesenteric lymph nodes and show reduced Treg cell induction and the absence of Th17 cells as well as Th1/Th2 imbalance, which is biased toward the Th2 response.12, 19 In this experimental system, Bacteroides fragilis recovered the development of the Th1-associated immune response through a bacterial product, polysaccharide A-dependent pathway.17 In mice under specific pathogen-free conditions, polysaccharide A derived from Bacteroides fragilis inhibits Th17 development and induces Treg cell accumulation by engaging with Toll-like receptor 2 expressed by T cells, leading to the maintenance of immune homeostasis.20 In other studies, while segmented filamentous bacteria (SFB) were shown to induce a Th17 immune response, Clostridium sp. was shown to promote Treg cell induction.15, 19 In addition, in the case of co-infection with SFB and Listeria monocytogenes, SFB-specific T cells and Listeria-specific T cells were shown to differentiate into Th17 and Th1 cells, respectively.21 This directly suggests that cognate bacterial antigens could determine the differentiation of Th cells. Therefore, the development of Th cells can be controlled by the microbiota, and the disturbance of Th-cell development by an altered microbiota may contribute to the pathogenesis of diseases.
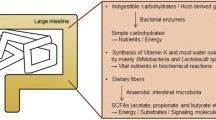
Recently, emerging reports have highlighted the role of the microbiota in the pathogenesis of autoimmune and inflammatory diseases, such as rheumatoid arthritis (RA), type 1 and 2 diabetes mellitus (DM), and asthma. Curiously, because these diseases exhibit organ-specific disease pathology, the microbiota, which is restricted to the gut, lung and skin, requires signals to communicate with the host immune system in distant organs. As signal mediators, microbes per se, circulating antibodies or immune cells, hormones and microbiota-derived metabolites including lipopolysaccharide, short-chain fatty acids (SCFAs, for example, butyrate, propionate and acetate), and bile acid may communicate with distant organs, physiologically enabling the development of distant organ-specific diseases (Figure 1).10 However, although communication with distal organs is a very critical point in demonstrating the direct relationship between the microbiota and disease development, it remains to be addressed.

Microbiota diversity is determined by environmental factors and signals to distal organs that contribute the development of diseases. The microbiota is established by other environmental factors, such as dietary fiber, saturated lipids, infection and antibiotics, and its colonization depends on the physiological condition of each tissue. Altered commensal microbiota in the gut or lung could influence the progression of various tissue-specific diseases through signal mediators, including microbes, microbial metabolites and circulating immune cells.
T-cell-associated inflammatory diseases affected by altered microbiota
Rheumatoid arthritis
RA is a systemic autoimmune disease that results in bone and cartilage destruction by joint inflammation.22 The hallmark of RA as an autoimmune disease is the production of autoantibodies such as rheumatoid factor and anti-citrullinated protein antibody.22 Together with increased autoantibody production, the production of pro-inflammatory cytokines is elevated in the joint synovium of patients with RA.23 The joints of patients with RA are complicated tissues where innate and adaptive immune cells as well as joint resident cells, like synoviocytes and chondrocytes, are involved in joint inflammation.23 Among several inflammatory cells, Th17 cells producing IL-17, IL-21 and IL-22 have been identified as the major pathological factor in the exacerbation of RA.24, 25 The frequency of Th17 cells and IL-17 levels are also increased in the peripheral blood and synovial fluid of patients with RA.26
Although genome-wide association studies and subsequent meta-analyses have found genetic risk factors like HLA-DR, the pathogenesis of RA cannot be fully interpreted by considering only genetic variations.22 Recently, the gut microbiota has been proposed as an indispensable environmental factor in the progression of RA.27 Surprisingly, RA patients have been shown to have alterations in the composition of the microbiota. For example, while the abundance of Bifidobacterium and Bacteroides is lower in patients with RA, that of Lactobacillus salivarius, Lactobacillus iners and Lactobacillus ruminis is higher in patients with early RA.28, 29 Likewise, Prevotella copri is positively correlated with new-onset untreated RA patients.30 Chen et al.31 have shown that the decreased gut microbial diversity of RA patients is associated with disease duration and that the expansion of rare microbial lineages characterizes the RA-associated gut microbiota.
Emerging evidence has highlighted that the alteration of T-cell differentiation by the gut microbiota and its metabolic products as one of the most abundant environmental factors encountered by the human body has implicated in autoimmune diseases.32 Such evidence indicates that the gut microbiota may control the host immune system, triggering T-cell differentiation. In parallel with this hypothesis, it has been reported that Th17 cells are abundant in the small intestinal lamina propria of the gut and do not accumulate in the absence of commensal microbiota, as in GF mice.33, 34 In particular, the colonization of SFB, a gut symbiont, induces and activates Th17 cells to produce the pro-inflammatory cytokine IL-17 in GF mice.19 Conversely, polysaccharide A produced from Bacteroides fragilis, which is also a gut symbiont, enhances the induction of Th1 and Foxp3+ Treg cells to attenuate the Th17 response.17, 35 Thus, the composition of the gut microbiota plays a pivotal role in the balance between inflammatory Th cells and suppressive Treg cells to maintain immune tolerance under healthy conditions.
The alteration of the gut microbiota, namely dysbiosis, may influence immune tolerance and lead to the development of autoimmune diseases such as RA in genetically susceptible models.27 In a murine experimental model, the colonization of SFB in the gut of spontaneous arthritis-prone K/B × N mice under GF conditions leads to the induction of functional Th17 cells and the reduction of Treg cells, driving the onset of autoimmune arthritis.36 In this process, SFB can promote the production of serum amyloid A in the ileum to induce Th17-cell differentiation.19 Regarding the plasticity of Th-cell differentiation, recent studies have described that RORγt precursor Th cells could differentiate into either Th17 or Treg cells depending on the local concentration of IL-6, a Th17-driving molecule.37, 38 On the basis of this hypothesis, SFB may govern the differentiation of pro-inflammatory Th17 cells and anti-inflammatory Treg cells through the regulation of each Th cell subset-driving cytokine milieu under specific microenvironmental conditions.16 Consistent with the above findings, Atarashi et al.33 showed that commensal bacteria-derived adenosine 5′-triphosphate could promote the expression of Th17-prone molecules, such as IL-6, IL-23p19 and transforming growth factor-β-activating integrins-αV and -β8 in lamina propria cells, and preferentially induce Th17-cell differentiation.
In addition to K/B × N mouse model, SKG mice and IL-1 receptor antagonist knockout (IL-1rn−/−) mice, other spontaneous T-cell-mediated arthritis models do not develop arthritis under GF conditions.39, 40 The colonization of Lactobacillus bifidus drives the development of arthritis in IL-1rn−/− mice.39 In another arthritis-prone DBA1 mouse model, the mice were divided into collagen-induced arthritis (CIA)-susceptible and CIA-resistant groups after CIA induction.41 After that, the microbiota derived from CIA-susceptible mice caused an increased incidence of arthritis when transferred to GF mice compared to the microbiota derived from CIA-resistant mice.41 At the same time, IL-17 in serum and Th17 cells in the spleen were upregulated in GF mice transferred with the microbiota derived from CIA-susceptible mice.41 Therefore, it has been demonstrated that in various genetically susceptible models, the gut microbiota could orchestrate the development of autoimmune arthritis through a Th cell-mediated pathway.
Beyond the gut microbiota, the association of the oral and lung microbiota with RA disease pathology has been described. Interestingly, the presence of Porphyromonas gingivalis, which is known as periodontitis-associated bacteria and to enable translational modifications resulting in citrullinated proteins, in RA patients could have a significant impact on the relationship of RA pathogenesis with distal tissue microenvironments.42, 43 Citrullination is generated by peptidyl-arginine-deiminases, resulting in a loss of tolerance and anti-citrullinated protein antibody responses related to RA development.22 Furthermore, P. gingivalis is known to promote Th17 responses through the production of Th17-polarizing cytokines even in chronic periodontitis.44 Accordingly, P. gingivalis as an oral microbe could be associated with the development of RA.45 Recent reports have shown that there are anti-citrullinated protein antibodies in induced sputum and lung tissues of patients with RA.46 Especially, Prevotella, known as a representative gut microbe of RA patients, is present in bronchoalveolar lavage fluid of RA patients and is positively correlated with systemic rheumatoid factor.46
Although numerous studies have suggested the possibility of deciphering the interplay between the microbiota and T-cell-mediated host immune responses in the progression of RA, the precise mechanism is still needed for the development of a therapeutic strategy. Table 1 summarizes the changes in the microbiota in RA patients and animal models of chronic arthritis.
DM and obesity
DM is one of the most common metabolic diseases, affecting the balance of blood glucose homeostasis and resulting from a failure of insulin production or insulin unresponsiveness.47 There are two major types of DM: type 1 (T1D) and type 2 diabetes (T2D). T1D is an organ-specific autoimmune disorder characterized by T-cell-mediated destruction of pancreatic β cells and insulin-dependent DM. The disease prevalence of T1D has risen steadily in developed countries, suggesting that numerous alterations to environmental factors such as diet, sanitation, infection and antibiotic use could cause disease development.48 Strikingly, the incidence of T1D in a mouse model under GF conditions was accelerated compared to that in a mouse model under a conventional environment.48, 49 This indicates that a commensal microbiota could optimize immune homeostasis to prevent the development of autoimmune diabetes.
In the disease progression of T1D, CD8 T cells are predominantly recruited to the islets, but their function to destroy β-cells requires the help of CD4 T cells.50 T1D is considered a Th1-cell-mediated disease that functions in an IFN-γ-dependent manner, which is associated with the CD8 T-cell response.51, 52 Insulin-dependent ectopic expression of IFN-γ in transgenic mice was demonstrated to sufficient cause the development of diabetes.53 Consistent with this result, increased IFN-γ was shown to be positively correlated with disease progression in non-obese diabetic (NOD) mice, which is a spontaneous autoimmune diabetes murine model used to study T1D.54 Considering infection by pathogens, when NOD mice were infected with attenuated Salmonella typhimurium, the incidence of T1D decreased via Th1 cell- and IFN-γ-dependent inhibition.52, 55, 56 In contrast, insulin-dependent ectopic expression of IL-4 ameliorated autoimmune diabetes in NOD mice.57 Helminth infections, such as Schistosoma mansoni, Trichinella spiralis and Heligmosomoides polygyrus, suppressed autoimmune diabetes in NOD mice via inducing the shift to a Th2 immune response.52, 58, 59 However, there is also controversial evidence for a Th1/Th2 paradigm in T1D pathogenesis. Neither recombinant IFN-γ nor deficiency of IL-4 exacerbated diabetes in NOD mice.60, 61
Th17 cells have been described as a critical regulator beyond the Th1/Th2 paradigm in the pathogenesis of T1D, but their role in disease development has been more controversial. While the suppression of Th17 cells significantly reduced the development of diabetes in NOD mice, the colonization of SFB as a robust inducer of Th17 cells promoted a Th17-mediated immune response to protect against autoimmune diabetes in NOD mice.62, 63 Besides, islet antigen-specific Th17 cells are able to be converted into IFN-γ-producing Th1-like cells, inducing diabetes in mice.64 Overall, the role of Th17 cells in T1D development may be determined by the disease stage or immune composition in disease-susceptible environments.
The differentiation of Treg cells as a counterpart to inflammatory T cells can also be shaped by the commensal microbiota in the intestines. In particular, T-cell receptors of intestinal Treg cells are educated by intestinal antigens, including microbial components, to suppress the immune activation against commensal microorganisms and maintain immune tolerance.5 For example, Cluster IV and XIVa Clostridia induces the Treg cell population through the production of SCFA, which induces Foxp3 expression in CD4 T cells.15, 65, 66 Intestinal Foxp3+ Treg cells have several mechanisms to suppress mucosal immune activation through IL-10 and transforming growth factor-β expression.67, 68 As mentioned above, when Treg cells are reduced in GF mice, the expression of suppressive factors, including IL-10, CTLA4 and ICOS, are simultaneously decreased in GF mice compared to conventionally reared mice but not specific pathogen-free mice.15, 65, 69
T2D is regarded as an inflammatory disorder associated with obesity and accompanied by chronic, low-grade inflammation of adipose tissue.70 Unlike T1D, T2D does not properly respond to insulin treatment, and therefore, it is categorized into insulin-resistant DM. T2D can be induced by a high-fat diet (HFD) linked to obesity as a metabolic inflammatory condition, leading to the production of pro-inflammatory cytokines such as IL-1β, IL-6 and tumor necrosis factor-alpha.71 T cells, in general, are infiltrated and accumulated in obese adipose tissue with the accumulation of macrophages.71, 72 Among T-cell subpopulations, CD8 T cells are highly infiltrated in the adipose tissue of HFD mice, whereas the number of CD4 T cells, particularly Treg cells, is strikingly reduced, even though Treg cells in adipose tissue have unique T-cell receptor (TCR) repertoires.70, 73 Together with the reduction of Treg cells, the number of Th17 cells also markedly decreased in HFD mice.74 In parallel with this result, IL17/RORγt-deficient CD4 T cells promote the development of T2D and obesity.74
Two independent research groups have determined that the ratio of Firmicutes to Bacteroidetes phyla is increased in the gut microbiota in HFD-fed mice and obese humans, which is associated with increased nutrient production in the host.75, 76 This indicates that obesity as a metabolic inflammatory condition can contribute to the alteration of the gut microbiota. Similarly, GF mice transferred with the gut microbiota of HFD-fed mice showed a greater increase in total body fat than those transferred with normal microbiota.76 Importantly, it has been demonstrated that HFD-induced gut microbiota causes a reduction of intestinal Th17 cells through the impairment of the ability of intestinal antigen-presenting cells to induce functional Th17 cells.74, 77 Karlsson and Qin research groups have observed the changes in the gut microbiota in T2D and surprisingly found a decrease in an SCFA-producing strain, Clostridia, known as a potent inducer of Treg cells.13, 78, 79 Overall, recent reports suggest that the altered gut microbiota contributes to the reduction of Th17 and Treg cells in the progression of T2D and obesity. Table 1 summarizes the changes in the microbiota in diabetes patients and NOD mice.
Asthma
Asthma is one of the most common chronic airway diseases and is characterized by airway inflammation, airway hyper-responsiveness, reversible airway obstruction and airway remodeling.80 Immunologically, two types of asthma have been largely defined: allergic asthma and non-allergic asthma. Interestingly, distinct T-cell phenotypes exist in asthmatic patient subpopulations. Allergic asthma, as a Th2-cell-driven disorder, has been considered to be eosinophilic asthma.80 The presence of Th2 cells is increased in the airways of patients with allergic asthma.81 In parallel with the functions of the Th2-associated cytokines IL-4, IL-5 and IL-13, infiltration of eosinophils and increased immunoglobulin E (IgE) levels in serum and bronchoalveolar lavage fluid from patients with asthma are the hallmarks of Th2-associated inflammation.80, 81, 82 However, some patients suffer from asthma despite an absence of Th2 cytokines. This non-allergic asthma, as a non-Th2-driven disorder, has been categorized as neutrophilic asthma.80 Microbial components and their products stimulate the innate immune system, resulting in the production of IL-8, IL-1 and tumor necrosis factor-alpha.83 These pro-inflammatory cytokines drive the shift toward Th1 and Th17 immune responses, which promote the recruitment of neutrophils.83 IL-17A is increased in the lungs of patients with asthma, and the levels of IL-17 correlate with disease severity.84, 85
Recent rapid increases in the disease prevalence have occurred worldwide along with an urban living style. Asthma is caused by the interplay between genetic risk factors and subsequent exposure to environmental factors such as allergens, smoking and infection. Although genetic risk alleles have been reported in the literature related to the pathogenesis of asthma, the disease prevalence over the recent decades, particularly that of childhood asthma development in westernized countries, indicates that environmental factors play a crucial role in disease development.86 Moreover, emerging evidence suggests an association of the composition of the lung microbiota with the development and exacerbation of asthma. Anatomically, the airway microbiota consists of dispersed bacteria from the oral cavity and microbes obtained through the inhalation of air, containing 104–106 bacterial cells per m3.87 Although most inhaled microbes are passed through without colonization, some of the microbes appear to colonize in the bronchi of healthy lungs, including the genera Prevotella, Veillonella and Streptococcus.88
Commensal airway microbiota can induce subclinical activation of the Th17 immune response to protect against respiratory pathogens.89, 90 Interestingly, GF mice that lack any exposure to microbes and microbial colonization showed more severe immune responses of ovalbumin-induced allergic airway inflammation, suggesting that the commensal microbiota is essential for developing a normal immune system and protecting against airway inflammation.91 With the progress of recent serological approaches to identify microbe-specific antibodies and species-targeted PCR-based approaches, several studies have shown that the upper and lower airways of patients with asthma have a bacterial microbiota distinct from that of healthy people, which indicates a relationship between airway microbial diversity and susceptibility to asthma.
Chlamydia pneumoniae and Mycoplasma pneumoniae infections have been found to be associated with the acute exacerbation of asthma.92 A significant increase in microbes of the Proteobacteria phylum, including Haemophilus spp., Pseudomonas spp. and Klebsiella spp., has been consistently detected in the airways of patients with asthma, unlike those of healthy people.93, 94, 95 In addition, the microbiome profile of stable asthmatic patients indicates a positive correlation between the abundance of specific airway microbiome members, such as Comamonadaceae, Sphingomonadaceae and Oxalobacteraceae, and the degree of bronchial hyper-responsiveness, a key feature of asthma.93 Therefore, it is envisaged that the heterogeneity in asthma phenotypes may be caused by the distinct microbial compositions in the airways. Using next-generation sequencing and phylogenetic microarrays, cohort research of asthmatic patients has demonstrated that there is a distinct microbial community between the airways of corticosteroid-sensitive and corticosteroid-resistant (CR) patients.94 Corticosteroid-sensitive patients were enriched in Bradyrhizobium and Fusobacterium members, which have a longer acyl chain lipopolysaccharide with relatively low endotoxicity, reducing the bacteria-induced innate immune response.94 Conversely, CR patients were enriched in Proteobacteria, including Neisseria and Haemophilus, which have short acyl chains with high endotoxicity, enhancing IL-8 production through Toll-like receptor 4 activation.86, 94
As mentioned above, in case of allergic asthma, environmental bacteria exposure in early life may affect the microbiota composition and modulate the host immune response. Of interest, it has been reported that altered symbiotic microbiota early in life could induce an exacerbated type 2 immune response and allergic inflammation.95 In the murine model of exposure to house dust mite allergen immediately after birth, the mice developed enhanced Th2-mediated inflammation and airway hyper-responsiveness. Under these conditions, with the compositional shift from a predominance of Proteobacteria and Firmicutes to Bacteroidetes phyla, CD4+ CD25+ Foxp3+ Helios− Treg cells expanded.96, 97 In another study, patients with severe asthma, especially those with neutrophilic airway inflammation, exhibited an abundance of Haemophilus, Streptococcus and Moraxella sp. in induced sputum, and this abundance was positively correlation with the neutrophil counts and IL-8 concentrations in patient samples.98 In addition, the airway commensal microbiota and their metabolically active products can induce subclinical activation of the mucosal Th17 immune response.97
Asthma is a very heterogeneous disease that is strongly influenced by environmental factors early in life. Although it remains to be addressed, we suggest that the altered microbiota profile provides clues to account for the heterogeneity of disease pathogenesis. Table 1 summarizes the changes in the microbiota in patients with asthma.
Summary and future perspectives
Immunologically, abundant evidence has indicated that the expansion of distinct T-cell subpopulations and their function may orchestrate specific autoimmune or inflammatory diseases through multifactorial pathways. Here we have raised significant questions regarding how and why a specific T-cell subpopulation is expanded and its unique features lead to the development of diseases. The etiology of diseases can be largely divided into genetic variations and environmental factors. In this review, among several hypotheses, we focused on the possibility that commensal microbes, which are the most abundant environmental factor in the body, could shape immune systems and potently trigger immune activation in genetically disease-susceptible individuals. In particular, we attempted to describe how the relationships between T cells of the adaptive immune system and commensal microbiota, symbionts or pathogens play crucial roles in healthy and disease conditions.
In homeostatic conditions, Bacteroides fragilis drives the induction of Th1 or Treg cells, protecting the host from infection caused by several pathogens or preventing immune activation by self-antigens.17, 20 In addition, SFB and Clostridia-produced SCFA induce the expansion of Th17 and Treg cells, respectively, to maintain immune homeostasis (Figure 2).15, 19 However, in genetically disease-susceptible models, these bacterial species and their products regulate disease processes. For example, the colonization of SFB activates Th17 cell responses to accelerate autoimmune arthritis in a spontaneous arthritis model, K/B × N mice, or to protect against autoimmune diabetes in diabetes-prone NOD mice.36, 63 The colonization of Lactobacillus bifidus also drives the development of arthritis in arthritis-prone IL-1rn−/− mice.39 The HFD-induced gut microbiota directs a decrease of Th17 and Treg cells to suppress T2D and obesity.77 At this time, we wonder whether each T-cell subpopulation has specific TCRs educated by commensal bacterial antigens. In other words, do bacterial antigens generate distinct T-cell subpopulations with bacterial antigen-specific TCRs? This question would suggest the possibility that mucosal tissue-specific T cells may differ from systemic T cells, as the generation of different TCRs depends upon the local microenvironments.

Microbiota mediates T-cell differentiation in homeostatic or pathogenic conditions. In mice under germ-free (GF) conditions, Bacteroides fragilis restores the development of the Th1-associated immune response through a bacterial product, polysaccharide A (PSA)-dependent pathway, while in mice under specific pathogen-free (SPF) conditions, PSA derived from B. fragilis induces Treg cell accumulation. Segmented filamentous bacteria (SFB) induces a Th17 immune response through adenosine 5′-triphosphate (ATP) production or serum amyloid A (SAA) produced by innate cells. Clostridium sp. promotes Treg cells through short-chain fatty acid (SCFA) production. Antigen-presenting cells (APCs) activated by cognate bacterial antigens could facilitate the generation of tissue-specific T cells derived from systemic T cells in a specific tissue environment.
Several human metagenomic studies using 16S rRNA sequencing and whole-genome shotgun have been performed and have identified differences in the taxonomic microbiologic community between healthy individuals and disease patients. Beyond recognizing compositional dysbiosis, researchers have attempted to translate the compositional clusters of microbiota into functional repertoires such as mucin-degrading bacteria, oxidative stress response-related bacteria and butyrate-producing bacteria. Consequently, such research would facilitate an understanding of the relationship between dysbiosis and disease pathogenesis. However, because investigations of the human microbiota are primarily focused on the correlation with disease pathological phenotypes, these are needed to address the issue of causality. To systematically overcome the limits of human microbiota research, various mouse model systems have been used for mechanistic investigations, due to the fact that they have strong similarities with human genetics and the human gut microbiota. In an attempt to understand the effects of the human microbiota, human microbiota-associated mice have been developed and used in human-like ecologic systems.
Although we present the interplay between the microbiota and T cells in healthy and disease conditions, the overall immune system is a complex network affected by genetic susceptibility and various environmental triggers that initiate and contribute to disease progression. Hence, to understand the role of the microbiota in the development of diseases, we need to elucidate the broad network that exists between the innate and adaptive immune system in different disease conditions and the effect of dysbiosis on this network. In addition, with the field of metagenomics continuing to grow, it will be necessary to discover the linkages between different microbiota profiles under specific pathogenic conditions and different pathologic mechanisms shown in diseases.
References
Luckheeram RV, Zhou R, Verma AD, Xia B . CD4+ T cells: differentiation and functions. Clin Dev Immunol 2012; 2012: 925135.
Zhu J, Yamane H, Paul WE . Differentiation of effector CD4 T cell populations. Annu Rev Immunol 2010; 28: 445–489.
Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol 2005; 6: 1123–1132.
Weaver CT, Harrington LE, Mangan PR, Gavrieli M, Murphy KM . Th17: an effector CD4 T cell lineage with regulatory T cell ties. Immunity 2006; 24: 677–688.
Tanoue T, Atarashi K, Honda K . Development and maintenance of intestinal regulatory T cells. Nat Rev Immunol 2016; 16: 295–309.
King C, Tangye SG, Mackay CRT . Follicular helper (TFH) cells in normal and dysregulated immune responses. Annu Rev Immunol 2008; 26: 741–766.
Kaplan MH, Hufford MM, Olson MR . The development and in vivo function of T helper 9 cells. Nat Rev Immunol 2015; 15: 295–307.
NIH HMP Working Group NIH HMP Working Group Peterson J, NIH HMP Working Group Garges S, NIH HMP Working Group Giovanni M, NIH HMP Working Group McInnes P, NIH HMP Working Group Wang L et al. The NIH Human Microbiome Project. Genome Res 2009; 19: 2317–2323.
Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012; 486: 207–214.
Schroeder BO, Bäckhed F . Signals from the gut microbiota to distant organs in physiology and disease. Nat Med 2016; 22: 1079–1089.
Tamburini S, Shen N, Wu HC, Clemente JC . The microbiome in early life: implications for health outcomes. Nat Med 2016; 22: 713–722.
Wu HJ, Wu E . The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012; 3: 4–14.
Belkaid Y, Hand TW . Role of the microbiota in immunity and inflammation. Cell 2014; 157: 121–141.
Kim D, Yoo SA, Kim WU . Gut microbiota in autoimmunity: potential for clinical applications. Arch Pharm Res 2016; 39: 1565–1576.
Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011; 331: 337–341.
Gaboriau-Routhiau V, Rakotobe S, Lécuyer E, Mulder I, Lan A, Bridonneau C et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009; 31: 677–689.
Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL . An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005; 122: 107–118.
Smith K, McCoy KD, Macpherson AJ . Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin Immunol 2007; 19: 59–69.
Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009; 139: 485–498.
Round JL, Lee SM, Li J, Tran G, Jabri B, Chatila TA et al. The toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 2011; 332: 974–977.
Longman RS, Yang Y, Diehl GE, Kim SV, Littman DR, Microbiota . Host interactions in mucosal homeostasis and systemic autoimmunity. Cold Spring Harb Symp Quant Biol 2013; 78: 193–201.
McInnes IB, Schett G . The pathogenesis of rheumatoid arthritis. N Engl J Med 2011; 365: 2205–2219.
McInnes IB, Schett G . Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol 2007; 7: 429–442.
van den Berg WB, Miossec P . IL-17 as a future therapeutic target for rheumatoid arthritis. Nat Rev Rheumatol 2009; 5: 549–553.
Gaffen SL . The role of interleukin-17 in the pathogenesis of rheumatoid arthritis. Curr Rheumatol Rep 2009; 11: 365–370.
Leipe J, Grunke M, Dechant C, Reindl C, Kerzendorf U, Schulze-Koops H et al. Role of Th17 cells in human autoimmune arthritis. Arthritis Rheum 2010; 62: 2876–2885.
Wu X, He B, Liu J, Feng H, Ma Y, Li D et al. Molecular insight into gut microbiota and rheumatoid arthritis. Int J Mol Sci 2016; 17: 431.
Vaahtovuo J, Munukka E, Korkeamäki M, Luukkainen R, Toivanen P . Fecal microbiota in early rheumatoid arthritis. J Rheumatol 2008; 35: 1500–1505.
Liu X, Zou Q, Zeng B, Fang Y, Wei H . Analysis of fecal Lactobacillus community structure in patients with early rheumatoid arthritis. Curr Microbiol 2013; 67: 170–176.
Scher JU, Sczesnak A, Longman RS, Segata N, Ubeda C, Bielski C et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife 2013; 2: e01202.
Chen J, Wright K, Davis JM, Jeraldo P, Marietta EV, Murray J et al. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med 2016; 8: 43.
Duan J, Kasper DL . Regulation of T cells by gut commensal microbiota. Curr Opin Rheumatol 2011; 23: 372–376.
Atarashi K, Nishimura J, Shima T, Umesaki Y, Yamamoto M, Onoue M et al. ATP drives lamina propria T(H)17 cell differentiation. Nature 2008; 455: 808–812.
Ivanov II, Frutos Rde L, Manel N, Yoshinaga K, Rifkin DB, Sartor RB et al. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008; 4: 337–349.
Telesford KM, Yan W, Ochoa-Reparaz J, Pant A, Kircher C, Christy MA et al. A commensal symbiotic factor derived from Bacteroides fragilis promotes human CD39(+) Foxp3(+) T cells and Treg function. Gut Microbes 2015; 6: 234–242.
Wu HJ, Ivanov II, Darce J, Hattori K, Shima T, Umesaki Y et al. Gut-residing segmented filamentous bacteria drive autoimmune arthritis via T helper 17 cells. Immunity 2010; 32: 815–827.
Lochner M, Peduto L, Cherrier M, Sawa S, Langa F, Varona R et al. In vivo equilibrium of proinflammatory IL-17+ and regulatory IL-10+ Foxp3+ RORgamma t+ T cells. J Exp Med 2008; 205: 1381–1393.
Zhou L, Chong MM, Littman DR . Plasticity of CD4+ T cell lineage differentiation. Immunity 2009; 30: 646–655.
Abdollahi-Roodsaz S, Joosten LA, Koenders MI, Devesa I, Roelofs MF, Radstake TR et al. Stimulation of TLR2 and TLR4 differentially skews the balance of T cells in a mouse model of arthritis. J Clin Invest 2008; 118: 205–216.
Maeda Y, Kurakawa T, Umemoto E, Motooka D, Ito Y, Gotoh K et al. Dysbiosis contributes to arthritis development via activation of autoreactive T cells in the intestine. Arthritis Rheumatol 2016; 68: 2646–2661.
Liu X, Zeng B, Zhang J, Li W, Mou F, Wang H et al. Role of the gut microbiom in modulating arthritis progression in mice. Sci Rep 2016; 6: 30594.
Scher JU, Abramson SB . Periodontal disease, Porphyromonas gingivalis, and rheumatoid arthritis: what triggers autoimmunity and clinical disease? Arthritis Res Ther 2013; 15: 122.
Mikuls TR, Payne JB, Yu F, Thiele GM, Reynolds RJ, Cannon GW et al. Periodontitis and Porphyromonas gingivalis in patients with rheumatoid arthritis. Arthritis Rheumatol 2014; 66: 1090–1100.
Moutsopoulos NM, Kling HM, Angelov N, Jin W, Palmer RJ, Nares S et al. Porphyromonas gingivalis promotes Th17 inducing pathways in chronic periodontitis. J Autoimmun 2012; 39: 294–303.
Rosenstein ED, Greenwald RA, Kushner LJ, Weissmann G . Hypothesis: the humoral immune response to oral bacteria provides a stimulus for the development of rheumatoid arthritis. Inflammation 2004; 28: 311–318.
Scher JU, Joshua V, Artacho A, Abdollahi-Roodsaz S, Öckinger J, Kullberg S et al. The lung microbiota in early rheumatoid arthritis and autoimmunity. Microbiome 2016; 4: 60.
American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2009; 32: S62–S67.
Todd JA . Etiology of type 1 diabetes. Immunity 2010; 32: 457–467.
Alam C, Bittoun E, Bhagwat D, Valkonen S, Saari A, Jaakkola U et al. Effects of a germ-free environment on gut immune regulation and diabetes progression in non-obese diabetic (NOD) mice. Diabetologia 2011; 54: 1398–1406.
Bevan MJ . Helping the CD8+ T-cell response. Nat Rev Immunol 2004; 4: 595–602.
Savinov AY, Wong FS, Chervonsky AV . IFN-gamma affects homing of diabetogenic T cells. J Immunol 2001; 167: 6637–6643.
Lehuen A, Diana J, Zaccone P, Cooke A . Immune cell crosstalk in type 1 diabetes. Nat Rev Immunol 2010; 10: 501–513.
Sarvetnick N, Liggitt D, Pitts SL, Hansen SE, Stewart TA . Insulin-dependent diabetes mellitus induced in transgenic mice by ectopic expression of class II MHC and interferon-gamma. Cell 1988; 52: 773–782.
Rabinovitch A . Immunoregulatory and cytokine imbalances in the pathogenesis of IDDM. Therapeutic intervention by immunostimulation? Diabetes 1994; 43: 613–621.
Zaccone P, Raine T, Sidobre S, Kronenberg M, Mastroeni P, Cooke A . Salmonella typhimurium infection halts development of type 1 diabetes in NOD mice. Eur J Immunol 2004; 34: 3246–3256.
Raine T, Zaccone P, Mastroeni P, Cooke A . Salmonella typhimurium infection in nonobese diabetic mice generates immunomodulatory dendritic cells able to prevent type 1 diabetes. J Immunol 2006; 177: 2224–2233.
Mueller R, Krahl T, Sarvetnick N . Pancreatic expression of interleukin-4 abrogates insulitis and autoimmune diabetes in nonobese diabetic (NOD) mice. J Exp Med 1996; 184: 1093–1099.
Cooke A, Tonks P, Jones FM, O’Shea H, Hutchings P, Fulford AJ et al. Infection with Schistosoma mansoni prevents insulin dependent diabetes mellitus in non-obese diabetic mice. Parasite Immunol 1999; 21: 169–176.
Saunders KA, Raine T, Cooke A, Lawrence CE . Inhibition of autoimmune type 1 diabetes by gastrointestinal helminth infection. Infect Immun 2007; 75: 397–407.
Wang B, Gonzalez A, Hoglund P, Katz JD, Benoist C, Mathis D . Interleukin-4 deficiency does not exacerbate disease in NOD mice. Diabetes 1998; 47: 1207–1211.
Satoh J, Seino H, Abo T, Tanaka S, Shintani S, Ohta S et al. Recombinant human tumor necrosis factor alpha suppresses autoimmune diabetes in nonobese diabetic mice. J Clin Invest 1989; 84: 1345–1348.
Emamaullee JA, Davis J, Merani S, Toso C, Elliott JF, Thiesen A et al. Inhibition of Th17 cells regulates autoimmune diabetes in NOD mice. Diabetes 2009; 58: 1302–1311.
Kriegel MA, Sefik E, Hill JA, Wu HJ, Benoist C, Mathis D . Naturally transmitted segmented filamentous bacteria segregate with diabetes protection in nonobese diabetic mice. Proc Natl Acad Sci USA 2011; 108: 11548–11553.
Martin-Orozco N, Chung Y, Chang SH, Wang YH, Dong C . Th17 cells promote pancreatic inflammation but only induce diabetes efficiently in lymphopenic hosts after conversion into Th1 cells. Eur J Immunol 2009; 39: 216–224.
Atarashi K, Tanoue T, Oshima K, Suda W, Nagano Y, Nishikawa H et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013; 500: 232–236.
Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013; 504: 451–455.
Rubtsov YP, Rasmussen JP, Chi EY, Fontenot J, Castelli L, Ye X et al. Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity 2008; 28: 546–558.
Li MO, Wan YY, Flavell RA . T cell-produced transforming growth factor-β1 controls T cell tolerance and regulates Th1- and Th17-cell differentiation. Immunity 2007; 26: 579–591.
Geuking MB, Cahenzli J, Lawson MA, Ng DC, Slack E, Hapfelmeier S et al. Intestinal bacterial colonization induces mutualistic regulatory T cell responses. Immunity 2011; 34: 794–806.
Feuerer M, Herrero L, Cipolletta D, Naaz A, Wong J, Nayer A et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat Med 2009; 15: 930–939.
Donath MY, Shoelson SE . Type 2 diabetes as an inflammatory disease. Nat Rev Immunol 2011; 11: 98–107.
Lumeng CN, Bodzin JL, Saltiel AR . Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest 2007; 117: 75–184.
Nishimura S, Manabe I, Nagasaki M, Eto K, Yamashita H, Ohsugi M et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat Med 2009; 15: 914–920.
Garidou L, Pomié C, Klopp P, Waget A, Charpentier J, Aloulou M et al. The gut microbiota regulates intestinal CD4 T cells expressing RORγt and controls metabolic disease. Cell Metab 2015; 22: 100–112.
Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008; 57: 1470–1481.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006; 444: 1027–1031.
Céline P, Lucile G, Rémy B . Intestinal RORgt-generated Th17 cells control type 2 diabetes: a first antidiabetic target identified from the host to microbiota crosstalk. Inflamm Cell Signal 2016; 3: e1074.
Karlsson FH, Tremaroli V, Nookaew I, Bergström G, Behre CJ, Fagerberg B et al. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013; 498: 99–103.
Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012; 490: 55–60.
Lambrecht BN, Hammad H . The immunology of asthma. Nat Immunol 2015; 16: 45–56.
Robinson DS, Hamid Q, Ying S, Tsicopoulos A, Barkans J, Bentley AM et al. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. N Engl J Med 1992; 326: 298–304.
Cohn L, Elias JA, Chupp GL . Asthma: mechanisms of disease persistence and progression. Annu Rev Immunol 2004; 22: 789–815.
Earl CS, An SQ, Ryan RP . The changing face of asthma and its relation with microbes. Trends Microbiol 2015; 23: 408–418.
Molet S, Hamid Q, Davoine F, Nutku E, Taha R, Pagé N et al. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J Allergy Clin Immunol 2001; 108: 430–438.
Wang YH, Wills-Karp M . The potential role of interleukin-17 in severe asthma. Curr Allergy Asthma Rep 2011; 11: 388–394.
Fujimura KE, Lynch SV . Microbiota in allergy and asthma and the emerging relationship with the gut microbiome. Cell Host Microbe 2015; 17: 592–602.
Lighthart B . Mini-review of the concentration variations found in the alfresco atmospheric bacterial populations. Aerobiologia 2000; 16: 7–16.
Dickson RP, Erb-Downward JR, Huffnagle GB . Homeostasis and its disruption in the lung microbiome. Am J Physiol Lung Cell Mol Physiol 2015; 309: L1047–L1055.
Segal LN, Clemente JC, Tsay JC, Koralov SB, Keller BC, Wu BG et al. Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a Th17 phenotype. Nat Microbiol 2016; 1: 16031.
Wu W, Huang J, Duan B, Traficante DC, Hong H, Risech M et al. Th17-stimulating protein vaccines confer protection against Pseudomonas aeruginosa pneumonia. Am J Respir Crit Care Med 2012; 186: 420–427.
Herbst T, Sichelstiel A, Schär C, Yadava K, Bürki K, Cahenzli J et al. Dysregulation of allergic airway inflammation in the absence of microbial colonization. Am J Respir Crit Care Med 2011; 184: 198–205.
Sutherland ER, Martin RJ . Asthma and atypical bacterial infection. Chest 2007; 132: 1962–1966.
Huang YJ, Nelson CE, Brodie EL, Desantis TZ, Baek MS, Liu J et al. Airway microbiota and bronchial hyperresponsiveness in patients with suboptimally controlled asthma. J Allergy Clin Immunol 2011; 127: 372–381, e1–3.
Goleva E, Jackson LP, Harris JK, Robertson CE, Sutherland ER, Hall CF et al. The effects of airway microbiome on corticosteroid responsiveness in asthma. Am J Respir Crit Care Med 2013; 188: 1193–1201.
Marri PR, Stern DA, Wright AL, Billheimer D, Martinez FD . Asthma-associated differences in microbial composition of induced sputum. J Allergy Clin Immunol 2013; 131: 346–352.
Gollwitzer ES, Saglani S, Trompette A, Yadava K, Sherburn R, McCoy KD et al. Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat Med 2014; 20: 642–647.
Durack J, Boushey HA, Lynch SV . Airway microbiota and the implications of dysbiosis in asthma. Curr Allergy Asthma Rep 2016; 16: 52.
Green BJ, Wiriyachaiporn S, Grainge C, Rogers GB, Kehagia V, Lau L et al. Potentially pathogenic airway bacteria and neutrophilic inflammation in treatment resistant severe asthma. PLoS ONE 2014; 9: e100645.
Mejía-León ME, Barca AM . Diet, microbiota and immune system in type 1 diabetes development and evolution. Nutrients 2015; 7: 9171–9984.
Simpson JL, Daly J, Baines KJ, Yang IA, Upham JW, Reynolds PN et al. Airway dysbiosis: Haemophilus influenzae and Tropheryma in poorly controlled asthma. Eur Respir J 2016; 47: 792–800.
Acknowledgements
We thank all members of the Center for Integrative Rheumatoid Transcriptomics and Dynamics for their help and advice. This work was supported by grants from the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2015R1A3A2032927 and 2015R1C1A2A01055547).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Lee, N., Kim, WU. Microbiota in T-cell homeostasis and inflammatory diseases. Exp Mol Med 49, e340 (2017). https://doi.org/10.1038/emm.2017.36
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/emm.2017.36
- Springer Nature Limited
This article is cited by
-
Advancements in understanding the role of intestinal dysbacteriosis mediated mucosal immunity in IgA nephropathy
BMC Nephrology (2024)
-
Persistent dysbiosis of duodenal microbiota in patients with controlled pediatric Crohn’s disease after resolution of inflammation
Scientific Reports (2024)
-
An M cell-targeting recombinant L. lactis vaccine against four H. pylori adhesins
Applied Microbiology and Biotechnology (2024)
-
Dysbiosis of the Gut Microbiota in Patients with Psoriatic Arthritis is Closely Related to Lymphocyte Subsets and Cytokines
Inflammation (2024)
-
FK506 and Lactobacillus acidophilus ameliorate acute graft-versus-host disease by modulating the T helper 17/regulatory T-cell balance
Journal of Translational Medicine (2022)