
Abstract
In guilds, such as the large predators, species compete over multiple resources that are both consumable and non-consumable. The niche complementary hypothesis states that competitors can overlap in resource use if there is low overlap for at least one shared resource. In Africa, cheetah, Acinonyx jubatus, compete with other large carnivores across space, time and prey and are known to be subordinate; however, rarely has the entire guild been considered when assessing resource partitioning by cheetah. Using camera-trap data from four areas sampled over 4 years, we compared spatial and temporal overlap between cheetah and other large carnivores. Moreover, we estimated short-term avoidance of sites by cheetah after another large carnivore had visited. We show that cheetah align with the niche complementary hypothesis to avoid other large carnivores. Cheetah experienced high overlap with African wild dog, Lycaon pictus, activity patterns but avoided sites that were used frequently by them. The inverse was observed for lion, Panthera leo, and spotted hyena, Crocuta crocuta, with cheetah avoiding them across time but not space. Cheetah experienced moderate overlap with leopard, Panthera pardus, across both space and time, with only possible short-term avoidance occurring. Finally, cheetah exhibited high levels of spatial and temporal overlap with preferred prey species, suggesting a novel demonstration of the trade-off between resource acquisition and predator avoidance. Our results showcase the importance of taking multiple scales and resource axes into consideration when determining species abilities to co-exist and provides tools for managers working in highly managed systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Species that occur sympatrically, and are members of the same guild, typically compete for resources (Schoener 1983). Within the carnivore guild, competition or intraguild predation has been documented as important in defining the community structure and in some cases are believed to be the cause of the decline of subordinate species’ populations (Palomares and Caro 1999; Caro and Stoner 2003; Clements et al. 2014; Swanson et al. 2014). Competition occurs in one of two forms; exploitative competition, where one species reduces the availability of a resource for another, or interference competition, where one species behaviourally impacts another's use of a shared resource through direct interactions (i.e., harassment, kleptoparasitism, and predation) (Carothers and Jaksić 1984). Resource partitioning is a mechanism that promotes species co-existence through different use of food, space, or time (Harrington et al. 2009). In heterogeneous environments, resource partitioning is effective when species exhibit differences in their ability to exploit resources (Ziv et al. 1993; Wauters et al. 2002). Co-existing competitors must separate themselves by at least one resource axis, meaning that a high overlap in one niche dimension can be compensated for by low overlap in another, as stated in the niche-complementarity hypothesis (Vieira and Port 2007). In addition, several theories, including game or interference competition theory, suggest that dominant carnivores should be unrestricted with their resource use, matching that of their common prey, whereas resource partitioning by subordinate carnivores should be a trade-off between resource acquisition and risk avoidance (Vanak et al. 2013; Palomares et al. 2016).
Resource partitioning of non-consumable resources can involve spatial, temporal, and spatiotemporal partitioning. Spatial partitioning is when a species avoids habitats/sites that competitors use. Arboreal mammals (i.e., Macaca silenus, M. radiata, Semnopithecus johnii, and Ratufa indica) in the Western Ghats use vertical stratification to co-exist, occupying different heights within the forest canopy (Sushma and Singh 2006). Where they co-occur, ocelots, Leopardus pardalis, and bobcats, Lynx rufus, avoid each other through variation in microhabitat use (Horne et al. 2009), as do cougars, Puma concolor, and jaguars, Panthera once (Palomares et al. 2016). Temporal partitioning occurs when a species is active at different times to its competitors. Here, avoidance is used to describe a lack of overlap between species for a given resource. Mink, Neovison vison, become active during the day when otters, Lutra lutra, or polecats, Mustela putoris, (both of which are largely nocturnal) are present but remained nocturnal when these species are absent (Harrington et al. 2009). Spatiotemporal partitioning is the avoidance of habitats/sites shortly after a competitor was present (Niedballa et al. 2019). Studies have also looked at multiple axes simultaneously, including in Brazil where fox species (i.e., Cerdocyon thous and Psedalopex gymnocercus) had low overlap in activity patterns, moderate overlap across habitat use, but no differences in their prey use (Vieira and Port 2007). In South Africa, black-backed jackals, Canis mesomelas, dominated Cape foxes, Vulpes chama, and separate across multiple axes (space, time, and diet), but bat-eared foxes, Otocyon megalotis, which overlapped considerably with jackals in space and time, only separated by diet (Kamler et al. 2012). These examples highlight the importance of resource partitioning in the community structuring of carnivores.
Africa has a diverse large carnivore guild, consisting of lion, P. leo, leopard, P. pardus, spotted hyena, Crocuta crocuta, cheetah, Acinonyx jubatus, African wild dog, Lycaon pictus, brown hyena, Parahyaena brunnea, and striped hyena, Hyaena hyaena, with the first five being sympatric across most of their range (Vanak et al. 2013). All seven species’ global populations are declining, driven mainly by habitat loss and fragmentation, as well as human persecution (Ripple et al. 2014). Across their ranges, these carnivores are increasingly being relegated to smaller areas, often fenced game reserves, creating artificially high densities which in turn may cause an increase in competition (Darnell et al. 2014). Cheetah are a subordinate member of this guild due to their small build and largely solitary nature. Cheetah suffer high rates of cub and adult mortality (Laurenson 1994; Hunter 1998; Mills and Mills 2017) and frequently have their kills stolen by other large carnivores (Mills and Biggs 1993; Broekhuis et al. 2018). Given this subordinate status, cheetah have received considerable attention regarding possible resource partitioning to allow their co-existence with larger competitors (Durant 1998; Cozzi et al. 2012; Broekhuis et al. 2013; Vanak et al. 2013; Bissett et al. 2015; Swanson et al. 2016; Dröge et al. 2017; Cornhill and Kerley 2020a; Cornhill et al. 2021).
Traditionally, cheetah have been regarded as open savanna specialists that only hunt (and are mainly active) during the diurnal/crepuscular periods, largely to avoid the spatial and temporal activity peaks of dominant carnivores (Caro 1994; Laurenson 1994; Durant 1998; Hayward and Slotow 2009). Recently, work in other landscapes demonstrated that cheetah use a variety of open-to-closed woodlands effectively (Hunter 1998; Broomhall et al. 2003; Broekhuis et al. 2013; Rostro-García et al. 2015; Cornhill et al. 2021) and are active on moonlit nights. These studies, showcasing the variability in cheetah resource use, highlight the potential for greater spatial and temporal overlap between cheetah and other large carnivores than previously suggested. However, most attempts to resolve the effects of dominant competitors on cheetah behavior did not take the entire guild into account (Durant 1998; Vanak et al. 2013; Swanson et al. 2016; Dröge et al. 2017) or considered only the effect of temporal partitioning (Hayward and Slotow 2009). As carnivores’ ranges become increasingly restricted, understanding how the entire guild co-exists will become essential to managers and conservationist trying to conserve these species, especially those that are subordinate like the cheetah.
In this study, we aim to determine which factors, if any, cheetah use to separate themselves across space and time from other members of the large carnivore guild (lion, leopard, spotted hyena, and African wild dog) while still maintaining access to prey. Using camera-trap data, we analyze resource partitioning across two axes: spatial, and temporal on both the long- and short-term (spatiotemporal) scale. In addition, we explore how the effects of factors such as prey capture frequency and habitat characteristics (visibility) alter a cheetah’s resource use, as these may also influence cheetah behavior (Vanak et al. 2013). Overall, we hypothesize that cheetah should avoid other carnivores along at least one axis, thereby selecting for less risky areas and/or times. Additionally, we hypothesize that cheetah should align their spatial and temporal patterns with those of their preferred prey. Specifically, with respect to the spatiotemporal analysis, we predict that cheetah will avoid (camera-trap) sites shortly after other large carnivores have visited those sites (Broekhuis et al. 2013; Cornhill and Kerley 2020a). For the temporal and spatial analysis, we predict that cheetah should utilize times/sites least utilized by other carnivores (Durant 1998) and avoid extra-large-bodied prey (> 350 kg) that are too large for them to catch (Clements et al. 2014), but which are attractive to larger carnivores (Vanak et al. 2013).
Methods
Study area
This study was conducted in four fenced protected areas (two state-managed and two private reserves), in close proximity to one another in the KwaZulu-Natal province of South Africa (Fig. 1, Table 1). The state-managed reserves are Hluhluwe-iMfolozi Park (HiP) and uMkhuze Game Reserve (uMkhuze), and the privately owned reserves are Phinda Private Game Reserve (Phinda) and Manyoni Private Game Reserve (Manyoni). The region is characterized as hot, humid and subtropical with two distinct seasons: a dry winter from April to September, and a wet, hot summer from October to March (Balme et al. 2007). The reserves are located within the Maputaland Center of Endemism (Steenkamp et al 2004) and are dominated by mosaics of open grassland and various types of open-to-closed woodland (‘bushveld’) (Steenkamp et al. 2004). Currently, three out of the four reserves house an intact large carnivore guild (cheetah, lion, leopard, spotted hyena, and African wild dog). Resident African wild dogs do not occur in Phinda, but transients occasionally pass through and did during the 2016 survey. Brown hyena occur very rarely (two-to-three individuals in Manyoni) in the study area and were not considered further.

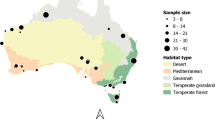
Map of the reserves (green) used for this study, and their location in KwaZulu-Natal, South Africa, with the black dots representing camera-trap sites: HiP (N = 46), Manyoni (N = 39), Phinda (N = 42), and uMkhuze (N = 40)
Data collection
From 2013 to 2016 (Table 1), cameras were set up in each of the four reserves for approximately 6 weeks, once each year or every alternate year (Miller et al. 2018). Each camera station consisted of two cameras (PantheraCam V-series), normally on opposite sides of the road/track, pointing toward each other to capture both sides of individual animals (Miller et al. 2018). Cameras were placed approximately 40 cm above the ground, triggered by movement with an 8 s delay and active 24 h a day. Camera stations were spaced about 1–3 km apart along roads, drainage lines, or game paths. For each image, the camera recorded the time and date, and experts identified the species photographed. We recorded when both cameras were inactive at a site (e.g., due to animal interference) to account for variable effort per site (Swanson et al. 2016).
During 2017, we measured vegetation openness or visibility at each camera site, to represent perceived predation risk (le Roux et al. 2018). We used a 1.6 × 0.3 m Nudds’ density board, that was marked into 20 cm sections (Nudds 1977). The observer positioned their eye level at approximately 80 cm height (an estimate of cheetah eye height), as visibility differs depending on the vantage point (Henley 2001). The visibility was measured in the four cardinal and the four primary inter-cardinal directions based on compass headings, at the center point between the two cameras. The distance at which approximately half of each 20 cm vertical section on the board was blocked by vegetation (Nudds 1977) was noted (to the closest meter) for all eight directions. We used the same observer for all sites to standardize bias. The final visibility value used was an averaged distance per site.
Panthera, a non-government organization, conducts regular camera-trap surveys within several protected areas in KwaZulu-Natal, primarily to obtain robust population estimates of leopard but also to track the relative abundance of other species. Images from these surveys were used for our analyses (Table S1). We recognize a possible bias in the data, as cameras were located to maximize the probability of capturing leopard (Miller et al. 2018). However, all carnivores regularly visited the sites and the surveys covered a full representation of the habitat types available to them. As this is a comparative study assessing point-specific spatial and temporal activity of the focal species (cheetah), as well as their competitors and prey, the data are deemed appropriate.
Statistical analysis
We conducted all statistical analysis in R (R Core Team 2021) and all mapping and spatial analysis was conducted in QGIS (QGIS Development Team).
Spatial analysis
For the spatial analysis, we analyzed the data by camera-trap site to estimate spatial avoidance or overlap among the cheetah and the other large carnivores. Due to low capture rates of cheetah, given the small populations in many of the small fenced reserves, data for all years were combined. When we had multiple consecutive captures of a species at a camera-trap site, we defined independence as 30 min between photos (Si et al. 2014). Additionally, we calculated prey abundance using a relative abundance index (RAI), defined as the number of independent prey captures per 100 trap days (Miller et al. 2018). RAIs are used for prey abundance, whereas predators are absolute counts given the quantity at which they are observed and the biases that can occur with RAIs and low densities of individuals (Sollmann et al. 2013). We grouped prey species into four categories based on their average adult female mass (Owen-Smith and Mills 2008): small (< 25 kg), medium (25–99 kg), large (100–350 kg), and extra-large (> 350 kg). Cheetah commonly predate small, medium, and large-bodied prey (the latter only typically available to cheetah male coalitions; Clements et al. 2014; Broekhuis et al. 2018). We assessed spatial avoidance as a hurdle model (package: glmm), in two steps: first to detect the presence of cheetah at a site (binomial response) and the second investigated cheetah abundance given that the site was used (truncated response) (Swanson et al. 2016).
We fit the data using a generalized linear mixed-effect model with a binomial response variable: cheetah presence/absence. The presence/absence of lion, leopard, spotted hyena, African wild dog and small, medium, large and extra-large-bodied prey were fixed effects, along with visibility [also a binomial variable based on quantiles into high (≥ 50th quantile) and low (< 50th quantile) visibility sites]. We included the trapping effort and camera-trap site ID nested in reserve as a random variable to control the different number of days sampled at sites/reserves and pseudoreplication.
To identify factors affecting the increased use of a site by cheetah, we used a truncated model, including only sites that had a minimum of one cheetah capture (Swanson et al. 2016). Once again, we ran a generalized linear mixed-effect model using the Poisson distribution with cheetah abundance (all captures summed per camera-trap site) as the response variable. Lion, leopard, spotted hyena, and African wild dog abundance were used as fixed effects, as was visibility and prey RAI per size class. Camera trap site ID nested in reserve and trap effort were used as random variables.
We determined the relative importance of carnivores on cheetah behavior by assessing the analysis of deviance for different models. The models included a null model (no fixed effects), a model with only visibility and prey size categories, models with visibility, prey and one carnivore species, and the full model containing visibility, prey, and all carnivores. The analysis of deviance was calculated using the model deviance and the formula: \(\frac{{{\text{Null model}} - {\text{model being tested}}}}{{{\text{Null model}} - {\text{full model}}}}\) (Swanson et al. 2016).
Temporal analysis
To test for temporal avoidance or overlap of cheetah with other carnivores and prey species, we calculated activity patterns for all species and measured the overlap between cheetah and all other species using the overlap package (Meredith and Ridout 2016). Using a kernel density plot, we calculated activity patterns for all carnivores and prey categories using all camera-trap images from all sites and reserves (Miller et al. 2018). We did not test for differences between reserves due to the small sample size of cheetah captures per reserve. We determined the overlap in activity patterns between cheetah and all other species using the non-parametric estimator for the coefficient of overlap (Miller et al. 2018). An overlap value of 0 indicates no overlap and 1 is an entire overlap of activity patterns by the two species (Ridout and Linkie 2009). We used a Kolmogorov–Smirnov test to determine overlap significance (Hayward and Slotow 2009).
Spatiotemporal analysis
Following the linear regression methods described by Niedballa et al. (2019), we analyzed the data to determine any spatiotemporal avoidance of other carnivores by cheetah by determined the time interval between images of cheetah and other carnivores at a site, regardless of whether the cheetah was the first or second visitor. We considered only carnivore captures and calculated the time between subsequent photos at sites, as long as the camera had remained active. Cheetah were species A, and the other carnivores were species, B. Therefore, we ended up with time intervals for AB, the time between a photograph of A, followed by species B, and for BA, the time difference between a photograph of species B followed by A. We then fitted a generalized linear mixed-effect model using the lme4 package (Bates et al. 2015). The time interval was the response variable with the order of species (AB or BA) as the fixed effect. We also included site ID nested in reserve as a random variable to control for pseudoreplication across sites. The data failed normality assumptions as assessed by the Shapiro–Wilk test, so the response variable was log transformed. We ran the model separately with lion, leopard, spotted hyena, and African wild dog as species B. The linear model estimates the difference in the time interval between AB and BA, with a positive estimate, that does not overlap zero, showing avoidance and a negative estimate, not overlapping zero, showing attraction (Niedballa et al. 2019). P values indicate whether the time interval between AB and BA is different, i.e., does A (cheetah) avoid/follow species B (other carnivores) more than species B does to species A.
Results
In total, 167 camera-trap sites were surveyed for a total of 18,578 days. This yielded 214 photos of cheetah (128 independent captures at 56 sites, 34%), 1055 photos of lion (648 independent captures at 129 sites, 77%), 1547 photos of leopard (1337 independent captures at 146 sites, 87%), 2346 photos of spotted hyena (1923 independent captures at 139 sites, 83%), 659 photos of African wild dog (278 independent captures at 83 sites, 50%), 4440 photos of small-bodied prey (4102 independent captures at 161 sites, 96%), 42,226 photos of medium-bodied prey (20,758 independent captures at 167 sites, 100%), 14,037 photos of large-bodied prey (6772 independent captures at 165 sites, 99%), and 2017 photos of extra-large-bodied prey (1390 independent captures at 122 sites, 73%).
Spatial partitioning
The variation in cheetah presence at camera-trap sites was explained equally by prey availability/visibility and carnivore presence (Table 2). African wild dog presence alone explained a quarter of the additional deviance (Table 2) and was negatively related to cheetah presence (z = − 2.16, p = 0.031; Fig. 2F, Table S3). Lion, leopard, and spotted hyena presence explained the least additional deviance in cheetah presence (Table 2) and were not significantly related to cheetah presence (p > 0.16; Fig. 2G–I, Table S3).

Presence of cheetah at camera-trap sites depending on environmental factors including habitat visibility, prey visitation, and intraguild carnivores. The factors are visibility (A), small prey (B), medium prey (C), large prey (D), extra-large prey (E), African wild dog (F), lion (G), leopard (H), and spotted hyena (I). The y-axis is a scale of cheetah presence with zero indicating absence and one indicates presence. Medium-sized prey were photographed at all sites. Only African wild dog presence (F) was negatively associated with cheetah presence (p = 0.015). The black dot indicates the mean and the bars indicates the standard error of that mean
Slightly over 50% of the variance in cheetah abundance at camera-trap sites was explained by prey RAI and visibility (Table 2). Cheetah abundance was positively associated with the RAI of large-bodied prey (z = 3.208, p = 0.001; Fig. 3D, Table S4). Lion abundance explained an additional 21% of the variation, but cheetah abundance showed a non-significant relationship with lion abundance (p = 0.10; Fig. 3G, Table S4). Spotted hyena abundance explained 16.5% of the additional variation in cheetah abundance but displayed a non-significant relationship (p = 0.12; Fig. 3I, Table S4). Leopard and African wild dog abundances each explained relatively little additional variance in cheetah abundance (4.3%, Table 2) and were not significant factors in cheetah abundance (p > 0.2; Fig. 3H, F, Table S4).

Cheetah abundance at camera-trap sites in response to environmental factors, including habitat visibility, prey visitation, and other carnivores. The factors are visibility (A), small prey (B), medium prey (C), large prey (D), extra-large prey (E), African wild dog (F), lion (G), leopard (H), and spotted hyena (I). Visibility is a measure of percent, openness, and prey factors are a relative abundance index (RAI) as opposed to the absolute abundance rates. Only large-bodied prey abundance (D) was positively associated with cheetah abundance (p = 0.001) indicated by a solid blue regression line. Dashed blue lines indicated non-significant regression lines and black dots are the values for each individual camera-trap site
Temporal partitioning
All carnivore and prey species’ activity patterns indicated activity throughout the 24-h period, with peaks at different times (Fig. 4). Cheetah activity peaked at dawn with a smaller peak in activity at dusk (Fig. 4), as did African wild dog (Fig. 4E). Small-bodied prey activity peaked during the crepuscular periods (Fig. 4A), whereas medium- and large-bodied prey activity levels peaked during the day (Fig. 4B, C). Extra-large-bodied prey had the highest activity at dusk (Fig. 4D). Lion, leopard, and spotted hyena activity levels peaked at night (Fig. 4F, G, H).

Temporal overlap of activity patterns for cheetah with small-sized prey (A), medium-sized prey (B), large-sized prey (C), extra-large-sized prey (D), African wild dog (E), lion (F), leopard (G), and spotted hyena (H) from camera-trap sites. The solid black line represents cheetah activity patterns, the dotted blue line is the prey or carnivore species, and the gray shaded area indicates the area of overlap
The activity patterns of cheetah were significantly different from all other carnivore and prey species (p ≤ 0.01, Table S5). Cheetah had the greatest temporal overlap with small-bodied prey (0.77), African wild dog (0.72), and large-bodied prey (0.70) (Fig. 4A, E, C). Cheetah had the least temporal overlap with spotted hyena (0.47) and lion (0.55) (Fig. 4F, H). Leopard (0.61), medium-bodied prey (0.63), and extra-large-bodied prey (0.65) had an intermediate level of temporal overlap with cheetah (Fig. 4G, B, D).
Spatiotemporal partitioning
There was a total of 43 captures of cheetah after lion (average of 5.2 days between), 61 after leopard (4.9 days), 64 after spotted hyena (5.7 days), and 8 after African wild dog (4.9 days). Additionally, there were 36 captures of lion after cheetah (5.8 days), 55 leopard (4.7 days), 53 spotted hyena (4.9 days), and 6 African wild dog (4.4 days). Due to the low number of consecutive captures between cheetah and other large carnivore, all sites were combined for this analysis. Cheetah showed spatiotemporal avoidance toward leopard but not more than leopard did to cheetah (p = 0.2). Cheetah showed an attraction toward sites visited by spotted hyena and a neutral reaction to those visited by lion and African wild dog, with all being non-significant (p > 0.5; Table S2).
Discussion
Our results demonstrate that cheetah space and time use follow the predictions of the niche-complementarity hypothesis, by avoiding other large carnivores on at least one of the non-dietary resource axes available to them, but they did not avoid any competitor on all axes (Table 3). Specifically, we show that cheetah avoided: African wild dog across space, spotted hyena and lion through time, and leopard through short-term avoidance (spatiotemporal). Moreover, we found support for cheetah aligning their habitat selection and activity patterns with that of their preferred prey and weak support for the avoidance of extra-large-bodied prey, as a way to avoid large carnivores that prefer large prey (Vanak et al. 2013). There is thus a trade-off in resource acquisition and risk avoidance in which cheetahs apparently prioritize finding suitable prey and are able to effectively reduce risky encounters with dangerous competitors without requiring constant avoidance behavior.
Cheetah showed the highest overlap in activity patterns with small- and large-bodied prey and their abundance at a camera-trap site was positively influenced by large-bodied prey. This supports our prediction that cheetah are attempting to align themselves with commonly utilized prey (Clements et al. 2014). The lack of support for medium-sized prey could have been biased by one of two things: the day-time activity of warthogs biasing that data diurnally (Hayward and Slotow 2009), and the high capture rate of this prey class at almost all sites due to the high abundance of species in this class in all reserves.
Our hypothesis for spatiotemporal avoidance by cheetah toward all other large carnivores was not supported. Previous studies have demonstrated a reactive response by cheetah toward lion and, in one case, leopard (Broekhuis et al. 2013; Swanson et al. 2016; Cornhill and Kerley 2020a), meaning that they avoided these species on a short spatiotemporal scale. Our data showed a spatiotemporal avoidance of sites after leopard had visited a camera-trap site. Spatiotemporal avoidance of other large carnivores could be occurring on a different scale than investigated here, given the cameras’ limited field of view.
We found mixed support for cheetah being active at times when other large carnivores are not. Our results showed cheetah to be active throughout the day and night. These results are aligned with the recently documented nocturnal behavior of cheetah (Cozzi et al. 2012; Broekhuis et al. 2014); 28.5% of cheetah captures in our study occurred at night. Cozzi et al. (2012) reported similar levels (25.6 ± 3.5%) of nighttime activity for cheetah and they suggested that moonlight facilitated nocturnal behavior, something we did not test. Cozzi et al. (2012), also, stated that the increased nocturnal activity of cheetah would lead to an increase in the overlap of activity patterns with other carnivores. We noted a greater overlap in activity patterns between cheetah and other large carnivores than that documented by Hayward and Slotow (2009); however, they collected their cheetah data from the literature using kill and movement data that would have been biased toward diurnal observation. This emphasizes the utility of using camera-trap data for assessing activity patterns. Moreover, future studies could investigate differences among seasons (i.e., wet/dry or summer/winter), something not done in this study due to a lack of data across all categories (Table 1).
Spatially, we again found mixed support for our hypothesis that cheetah would select sites with low-to-no dominant carnivore use. Cheetah presence was negatively related to African wild dog presence. This result supports our predictions, as cheetah had high temporal overlap with African wild dog, but avoided them across space, selecting for sites that African wild dog were not using. This may reflect exploitative competition between cheetah and African wild dog, as shown to occur at least one of our study sites (Cornhill and Kerley 2020b). However, it is also possible that this was not a product of cheetah avoiding African wild dog, but African wild dog avoiding sites where all other large carnivores were found (Vanak et al. 2013; Swanson et al. 2014; Dröge et al. 2017). In addition, lion abundance was positively associated with cheetah abundance, but otherwise large carnivores had no effect of cheetah habitat use. Dröge et al. (2017) found no spatial avoidance by cheetah toward other large carnivores, indicating that cheetah are most likely to select areas based on the prospects of obtaining prey.
If cheetah focused exclusively on risk avoidance, they would be at risk of starvation, for they could not capture prey. This is similar to the trade-off (food vs risk) response of the Serengeti herbivore community, with Sinclair (1985) showing that as grazing became limiting in the dry season, grazers were more willing to accept the risk of predation, than when grazing was abundant. We showed that cheetah were willing to utilize resources that would be deemed risky (e.g., night time and closed habitat) under the landscape of fear hypotheses (Laundré et al. 2001) as the risk associated with these resources must have not been high enough to warrant outright avoidance, a result supported by Swanson et al. (2016). Rather, cheetah are driven by their need to acquire prey and thus use other mechanisms to avoid risk (the niche complementary hypothesis).
Cheetah are not the first species found to follow the predictions of the niche-complementarity hypothesis, this also occurs among other carnivore species (e.g., Vulpes spp. Jones and Barmuta 1998; Vieira & Port 2007; Kamler et al. 2012). The niche complementary hypothesis explains how species can co-exist even when they are outcompeted on one resource axis, as they separate themselves along another. Moreover, it explains that taking into account the different scales of the resource axes can further separate out the species and impact their ability to co-exist. Our results expand on the current literature and can provide scientists evaluating other species in other systems, a set of dimensions to evaluate their species upon.
Our study is the first to evaluate cheetah resource use taking into account the entire large carnivore guild and across multiple axes. It expands on previous work (Durant 1998; Cozzi et al. 2012; Broekhuis et al. 2013; Vanak et al. 2013; Bissett et al. 2015; Swanson et al. 2016; Dröge et al. 2017), giving managers more refined tools to achieve their objectives. This is especially useful to managers struggling with the challenges of isolated populations and reliance on tourism revenue, such as at our study sites. Pressure from tourists for large carnivore sightings often leads to reserves stocking animals at higher than appropriate densities, with management actions required to ameliorate possible problems (Slotow and Hunter 2009; Clements et al. 2016). A greater understanding of how all sympatric carnivores as a guild influence cheetah behavior may contribute to improved management of the species. Our results show that even with potentially artificially high carnivore densities and with fences restricting movements, cheetahs are able to co-exist with intact large carnivore guilds given adequate prey densities.
Our study shows that cheetah uses resource partitioning to avoid the potential costs of competition, but that this also represents a trade-off with the need to acquire prey. Resource partitioning does not need to occur on all axes for all competing large carnivores, nor was the response by cheetah symmetrical across all competing large carnivores. As we assessed resource use on multiple axes, we were able to show that cheetah avoided different carnivores on different axes. These results showcase the importance of looking at the intact predator guild and their use of several resources to obtain a full understanding of how competition shapes resource use.
Data availability
The data will be made available upon reasonable request to the corresponding author.
References
Balme G, Hunter L, Slotow R (2007) Feeding habitat selection by hunting leopards Panthera pardus in a woodland savanna: prey catchability versus abundance. Anim Behav 74:589–598. https://doi.org/10.1016/j.anbehav.2006.12.014
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Bissett C, Parker DM, Bernard RTF, Perry TW (2015) Management-induced niche shift? The activity of cheetahs in the presence of lions. Afr J Wildl Res 45:197–203
Broekhuis F, Cozzi G, Valeix M, McNutt JW, Macdonald DW (2013) Risk avoidance in sympatric large carnivores: reactive or predictive? J Anim Ecol 82:1098–1105. https://doi.org/10.1111/1365-2656.12077
Broekhuis F, Grünewälder S, McNutt JW, Macdonald DW (2014) Optimal hunting conditions drive circalunar behavior of a diurnal carnivore. Behav Ecol 25:1268–1275. https://doi.org/10.1093/beheco/aru122
Broekhuis F, Thuo D, Hayward MW (2018) Feeding ecology of cheetahs in the Maasai Mara, Kenya and the potential for intra- and interspecific competition. J Zool Lond 304:65–72. https://doi.org/10.1111/jzo.12499
Broomhall LS, Mills MGL, du Toit JT (2003) Home range and habitat use by cheetahs (Acinonyx jubatus) in the Kruger National Park. J Zool Lond 261:119–128
Caro T (1994) Cheetah of the Serengeti Plains: group living in an asocial species. University of Chicago Press, Chicago
Caro TM, Stoner CJ (2003) The potential for interspecific competition among African carnivores. Biol Conserv 110:67–75
Carothers JH, Jaksić FM (1984) Time as a niche difference: the role of interference competition. Oikos 42:403–406
Clements HS, Cumming GS, Kerley GIH (2016) Predators on private land: broad-scale socioeconomic interactions influence large predator management. Ecol Soc 21:45–53. https://doi.org/10.5751/ES-08607-210245
Clements HS, Tambling CJ, Hayward MW, Kerley GIH (2014) An objective approach to determining the weight ranges of prey preferred by and accessible to the five large African carnivores. PLoS One 9:e101054. https://doi.org/10.1371/journal.pone.0101054
Cornhill K, Kerley GIH (2020a) Cheetah communication at scent-marking sites can be inhibited or delayed by predators. Behav Ecol Sociobiol 74:1–10. https://doi.org/10.1007/s00265-020-2802-9
Cornhill KL, Kerley GIH (2020b) Does competition shape cheetah prey use following African wild dog reintroductions? Afr J Wildl Res 50:75–85
Cornhill KL, Kelly C, Kerley GIH (2021) Lion reintroduction demonstrates that resident cheetah have a spatially reactive response to lion. Afr J Ecol 60:1–12. https://doi.org/10.1111/aje.12926
Cozzi G, Broekhuis F, McNutt JW, Turnbull LA, Macdonald DW, Schmid B (2012) Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecol 93:2590–2599
Darnell AM, Graf JA, Somers MJ, Slotow R, Gunther MS (2014) Space use of African wild dogs in relation to other large carnivores. PLoS ONE 9:1–9. https://doi.org/10.1371/journal.pone.0098846
Dröge E, Creel S, Becker MS, Jassiel M (2017) Spatial and temporal avoidance of risk within a large carnivore guild. Ecol Evol 7:189–199. https://doi.org/10.1002/ece3.2616
Durant SM (1998) Comparative refuges and coexsistance: an example from Serengeti carnivores. J Anim Ecol 67:370–386. https://doi.org/10.1046/j.1365-2656.1998.00202.x
Harrington LA, Harrington AL, Yamaguchi N, Thom MD, Ferreras P, Windham TR, Macdonald DW (2009) The impact of native competitors on an alien invasive: temporal niche shifts to avoid interspecific aggression? Ecology 90:1207–1216
Hayward MW, Slotow R (2009) Temporal partitioning of activity in large African carnivores: tests of multiple hypotheses. S Afr J Wildl Res 39:109–125
Henley S (2001) Habitat suitability and modelling for ungulates in the Thicket Biome. University of Port Elizabeth, Eastern Cape
Horne JS, Haines AM, Tewes ME, Laack LL (2009) Habitat partitioning by sympatric ocelots and bobcats: implications for recovery of ocelots in Southern Texas. Southwest Nat 54:119–126
Hunter LTB (1998) The behavioural ecology of reintroduced lions and cheetahs in the Phinda Resource Reserve, KwaZulu-Natal. University of Pretoria, Pretoria
Jones ME, Barmuta LA (1998) Diet overlap and relative abundance of sympatric dasyurid carnivores: a hypothesis of competition. J Anim Ecol 67:410–421
Kamler JF, Stenkewitz U, Klare U, Jacobsen NF, Macdonald DW (2012) Resource partitioning among Cape foxes, bat-eared foxes, and black-backed jackals in South Africa. J Wildl Manag 76:1241–1253. https://doi.org/10.1002/jwmg.354
Laundré JW, Hernández L, Altendorf KB (2001) Wolves, elk, and bison: reestablishing the “landscape of fear” in Yellowstone National Park, U.S.A. Can J Zool 79:1401–1409. https://doi.org/10.1139/cjz-79-8-1401
Laurenson MK (1994) High juvenile mortality in cheetahs (Acinonyx jubatus) and its consequences for maternal care. J Zool Lond 234:387–408
le Roux E, Kerley GIH, Cromsigt JPGM (2018) Megaherbivores modify trophic cascades triggered by fear of predation in an African Savanna ecosystem. Curr Biol 28:1–7. https://doi.org/10.1016/j.cub.2018.05.088
Meredith M, Ridout M (2016) Overlap: estimates of coefficient of overlapping for animal activity patterns. https://CRAN.Rproject.org/package=overlap
Miller JRB, Pitmann RT, Mann GK, Fuller AK, Balme GA (2018) Lions and leopards coexist without spatial, temporal or demographic effect of interspecific competition. J Anim Ecol 87:1709–1726. https://doi.org/10.1111/1365-2656.12883
Mills MGL, Biggs H (1993) Prey apportionment and related ecological relationships between large carnivores in Kruger National Park. Symp Zool Soc Lond 65:253–268
Mills MGL, Mills MEJ (2017) Kalahari Cheetah. Oxford University Press, Oxford. https://doi.org/10.1093/oso/9780198712145.001.0001
Niedballa J, Wilting A, Sollmann R, Hofer H, Courtiol A (2019) Assessing analytical methods for detecting spatiotemporal interactions between species from camera trapping data. Remote Sens Ecol Conserv 5:272–285. https://doi.org/10.1002/rse2.107
Nudds TD (1977) Quantifying the vegetation structure of wildlife cover. Wild Soc Bull 5:113–117
Owen-Smith N, Mills MGL (2008) Predator-prey size relationships in an African large-mammal food web. J Anim Ecol 77:173–183. https://doi.org/10.1111/j.1365-2656.2007.01314.x
Palomares F, Caro TM (1999) Interspecific killing among mammalian carnivores. Am Nat 153:492–508
Palomares F, Fernández N, Roques S, Chávez C, Silveira L, Keller C, Adrados B (2016) Fine-scale habitat segregation between two ecologically similar top predators. PLoS ONE 11:1–16. https://doi.org/10.1371/journal.pone.0155626
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed Feb 2021
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337. https://doi.org/10.1007/s11182-018-1503-6
Ripple WJ, Estes JA, Beschta RL, Wilmers CC, Ritchie EG, Hebblewhite M, Berger J, Elmhagen B, Letnic M, Nelson MP, Schmitz OJ, Smith DW, Wallach AD, Wirsing AJ (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1–11. https://doi.org/10.1126/science.1241484
Rostro-García S, Kamler JF, Hunter LTB (2015) To kill, stay or flee: the effects of lions and landscape factors on habitat and kill site selection of cheetahs in South Africa. PLoS ONE 10:1–20. https://doi.org/10.1371/journal.pone.0117743
Schoener TW (1983) Field experiments on interspecific competition. Am Nat 122:240–285
Si X, Kays R, Ding P (2014) How long is enough to detect terrestrial animals? Estimating the minimum trapping effort on camera traps. PeerJ 2:e374. https://doi.org/10.7717/peerj.374
Sinclair ARE (1985) Does interspecific competition or predation shape the African ungulate community? J Anim Ecol 54:899–918
Slotow R, Hunter LTB (2009) Reintroduction decisions taken at the incorrect social scale devalue their conservation contribution: the African lion in South Africa. In: Hayward MW, Somers MJ (eds) Reintroduction of top-order predators. Wiley-Blackwell Publishing, New York, p 457
Sollmann R, Mohamed A, Samejima H, Wilting A (2013) Risky business or simple solution—relative abundance indices from. Biol Conserv 159:405–412. https://doi.org/10.1016/j.biocon.2012.12.025
Steenkamp Y, Van Wyk B, Victor J, Hoare D, Smith G, Dold T, Cowling R (2004) Maputaland-Pondaland-Albany. In: Mittermeier RA, Robles-Gil P, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, Da Fonesca GAB (eds) Hotspots revisited: earth’s biologically richest and most endangered ecoregions. Cemex, Mexico City, pp 219–228
Sushma HS, Singh M (2006) Resource partitioning and interspecific interactions among sympatric rain forest arboreal mammals of the Western Ghats, India. Behav Ecol 17:479–490. https://doi.org/10.1093/beheco/arj058
Swanson A, Arnold T, Kosmala M, Forester J, Packer C (2016) In the absence of a “landscape of fear”: how lions, hyenas, and cheetahs coexist. Ecol Evol 6:8534–8545. https://doi.org/10.1002/ece3.2569
Swanson A, Caro T, Davies-Mostert H, Mills MGL, Macdonald W, Borner M, Masenga E, Packer C, Macdonald DW, Borner M, Masenga E, Packer C (2014) Cheetahs and wild dogs show contrasting patterns of suppression by lions. J Anim Ecol 83:1418–1427. https://doi.org/10.1111/1365-2656.12231
Vanak AT, Fortin D, Thaker M, Ogden M, Owen C, Greatwood S, Slotow R (2013) Moving to stay in place: behavioral mechanisms for coexistence of African large carnivores. Ecology 94:2619–2631
Vieira EM, Port D (2007) Niche overlap and resource partitioning between two sympatric fox species in southern Brazil. J Zool 272:57–63. https://doi.org/10.1111/j.1469-7998.2006.00237.x
Wauters LA, Gurnell J, Martinoli A, Tosi G (2002) Interspecific competition between native Eurasian red squirrels and alien grey squirrels: does resource partitioning occur? Behav Ecol Sociobiol 52:332–341. https://doi.org/10.1007/s00265-002-0516-9
Ziv Y, Abramsky Z, Kotler BP, Subach A (1993) Interference competition and temporal and habitat partitioning in two gerbil species. Oikos 66:237–246
Acknowledgements
The authors thank Ezemvelo KZN Wildlife, Manyoni Private Game Reserve, and Phinda Private Game Reserve for allowing us access to your reserves and data. The authors thank Ezemvelo KZN Wildlife, Phinda Habitat Research Team, and Wildlife ACT for their help with data collection. Finally, the authors thank Nelson Mandela University for bursary support for KLC.
Funding
Open access funding provided by Nelson Mandela University. Funding was provided by Nelson Mandela University, Panthera, and Wildlife ACT.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and designed either through the design of the broader camera-trap study or the finer concept of this manuscript. Material preparation, data collection, and analysis were performed by Kristina Cornhill. The first draft of the manuscript was written by Kristina Cornhill, and all authors reviewed and revised this manuscript.
Corresponding author
Ethics declarations
Conflict of interest
None to declare.
Ethical approval
All data were collected through non-invasive methods. There was no harm done to the animals and they were not handled or manipulated in any way, only observed through camera traps.
Informed consent
All invested parties are aware of this manuscript and approve its submission for publication.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling Editor: Michael Somers.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cornhill, K.L., Balme, G.A., Hunter, L.T.B. et al. Cheetah spatiotemporal overlap with other large carnivores and prey at camera-trap sites: do they fit the niche-complementarity hypothesis?. Mamm Biol 103, 505–517 (2023). https://doi.org/10.1007/s42991-023-00368-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-023-00368-1