
Abstract
The northeastern part of the Carpathian Lowland is one of the most species-rich parts of the region, despite the long-term and sometimes extreme changes in land use. The fragmented remains of formerly contiguous forests maintain a significant part of the wildlife living in different oak, mixed oak, and gallery forest, and even faunas of connected grassland habitats. To reveal how the parallel Carpathian and Pannonian effects form the local insect fauna, the Noctuoidea assemblages of eight forest patches were assessed and compared, based on the distribution data of 479 species. Three types of assemblages were designated, with similar diversity but different compositions. The most unique and valuable ones can be found in the colline area of Transcarpathia, where the high habitat diversity supported by the topography promotes the coexistence of the species of humid forest fauna and many steppic and southern elements. The protection and planned management of these remained forest patches and their network in international cooperation can maintain a significant part of the fauna of the affected regions.
Similar content being viewed by others


Introduction
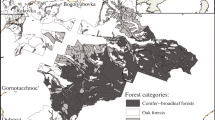
The northeastern part of the Carpathian Lowland has been formed mainly by the Tisza and Szamos Rivers and additionally by some small water streams. Formerly, this region was covered with large marshlands (Pécsi and Sárfalvi 1960), and it is the coldest (with 8.9 ºC annual mean temperature) and most humid (with an average of 609 mm annual precipitation) part of the lowland with significant continental effect on its climate and wildlife (Baranyi 2009). After the last ice age, a continuous deciduous forest has been grown where the regular floods and the nearness of the Carpathians formed the vegetation (Simon 1952). The region is located at the ternary border of Hungary, Romania and Ukraine. A significant part of the area is the Bereg Region belonging to the Chop-Mukachevo Basin, situated on the border of Hungary and Ukraine which is the border of the Pannonicum (Carpathian Lowland) and the Carpathicum (mountainous area) at the same time. The first reports on the flora of its forests were made in the 1940s; while, the investigation of the marshlands started 10 years later (Hargitai 1943; Simon 1960). The most valuable and unique part of the flora contains mountainous plants inhabiting lowland areas such as geophytes flowering in early spring (Crocus heuffelianus Herb, Gagea spathacea (Hayne) Salisb., Anemone nemorosa L., Isopyrum thalictroides L., Lathraea squamaria L.). Beyond that, in the grasslands and forest edges many forest-steppe species appear (Buglossoides purpureo-coeruleum (L.) Holub., Dianthus carthusiana L., Lathyrus niger (L.) Bernth., L. sylvestris L., Peucedanum alsaticum L., Rosa gallica L.); while, the species assemblages of hayfields and meadows are also especially species-rich (Lesku 2008). Referring to the strong Carpathian effect on the land snail (Mollusca) fauna, malacologists formerly mentioned the region as "Praecarpathicum" (Deli et al. 1997; Deli and Sümegi 1999), where the species richness and even the biodiversity reach a higher level than in the other parts of the lowland, considering such invertebrate taxa, as, e.g., land snails, orthopterans, carabid beetles and lepidopterans (Deli et al. 1997; Ködöböcz 1997; Magura et al. 1997; Deli and Sümegi 1999; Ködöböcz and Magura 1999; Gálik et al. 2001). Nevertheless, the lowlands are less studied than the connected hilly and mountainous areas. Orthopterological studies were started in the 1950s in the area, and more recent investigations were implemented since the early 2000s (Likovitch 1957, 1959; Kristin et al. 2011; Nagy et al. 2011, 2015; Szanyi et al. 2015a). Contrary, night active lepidopterans of the area have been studied only in the last decade (Lyashenko 2009; Geryak 2010, 2012, 2013; Kanarsky et al. 2011; Szanyi et al. 2012, 2015b, c, d, e, 2016, 2017; Nowacki et al. 2018).
Night active lepidopterans are a widely used indicator group of natural value and status of arboreal habitats, and a species-rich target taxa of biodiversity monitoring studies (Usher and Keiller 1998; Summerville et al. 2004; Rákosy and Schmitt 2011; Truxa and Fiedler 2012; Horváth 2013). For their quantitative sampling, a wide scale of different methods has been developed and used during the last decades (e.g., Baker 1963; Yela and Holyoak 1997; Beck and Linsenmair 2006; Nowinszky and Puskás 2011). Since several representatives of them are hostplant and habitat specialists, the ratio and relative frequency of their faunal components (life forms) can be used for evaluation and comparison of their assemblages in the same way as their species richness and species composition (Altermatt 2010; Merckx and Slade 2014; Végvári et al. 2014).
Here we compare the Noctuoidea fauna of the arboreal habitats of the Precarpathian hills and lowlands in order to evaluate how the two different faunas overlap and combine. The species composition of the local and regional fauna, and the composition of faunal types and faunal components were also evaluated, parallelly with the publishing of the check list of the regional Noctuoidea fauna. To support the conservation and management of the unique assemblages and habitats, the conservation value of the eight sampled habitats were also evaluated.
Material and methods
Sampling sites
Data collected of eight sampling sites (Table 1) situated in the hilly areas of the Precarpathian region and the northeast part of the Carpathian Lowland were involved in the analysis. Half of the sites situated in West Ukraine; while, the other half of them were located in Northeast Hungary.
Beregovo (UA)
Precarpathian hilly area in the surroundings of Beregovo city. The sampled forest has a double-layer canopy. In the upper layer, Tilia cordata Mill. and Quercus petraea (Matt.) Liebl. are dominant while in the lower layer, Tilia species are abundant. In the shrub layer, Acer tataricum L. and Crataegus monogyna Jacq. are common. The diverse landscape of the surroundings also contains gallery forests, and wet- and meso-xerophyl grasslands (mainly different pastures and hayfields).
Czorna hora (UA)
Small isolated volcanic hill with gentle southern and southwestern slopes, providing extraordinary warm climatic conditions to this region with a 9.9 ºC annual average temperature and a high amount of precipitation (an average of 750 mm/year) (Gerenczuk 1981). Contrary, the eastern slopes are steep with a functioning stone mine. The hill is mainly covered by forests dominated by oak (Quercus spp.). In the warm southern slopes, a mosaic of low, xerophilous oakwood forests and forest-steppes can be found. In the upper canopy of those, Quercus petraea, while in the lower, Tilia tomentosa Mill.and Fraxinus ornus L. were dominant. Along the bank of the Tisza River, gallery forest has grown (Nowacki et al. 2018).
Kireshi (UA)
Diverse landscape with mixed oak forest, forest-steppe and steppe grasslands in the Chop-Mukachevo Basin, close to the city of Khust (Tsys 1968; Marinich 1982, 1985). The largest lowland population of Narcissus poeticus L. in Central Europe lives in the “Narcissi Valley”, which is also located near to the city. The area of the valley is 257 ha, that has belonged to the Carpathian Biosphere Reserve, since 1978 (Didukh and Shelyag-Sosonko 2003).
Velyka dobron' (UA)
The Velyka Dobron Game Reserve is a similarly large, mixed forest of the Ukrainian part of the Bereg Region, as the Hungarian Lónya Forest. Its dominant forest type is the oak-ash-elm gallery forest (Fraxino pannonicae-Ulmetum), in which Quercus robur L. is the most dominant species, but Populus canescens (Aiton) Sm. and Ulmus minor Mill. also appear with remarkably high relative frequencies. The bush layer is also diverse, with high frequencies of Crataegus monogyna, Acer tataricum, and A. campestre L. The other important forest association of the area is the especially dense and closed Querco robori-Carpinetum, with significantly less diverse bush and grass layers. Grassland patches of the area are mainly formed and maintained by different kinds of land use. These secondary biotopes of the area provide habitats for unique and highly diverse invertebrate assemblages with significant remains of the wildlife of the former peatlands of the Szernye Marsh (Szanyi et al. 2012, 2015b, d, e, 2016, 2017).
Vámosatya (HUN)
Samplings were made in the Bockerek Forest, which is one of the largest remains of the former continuous forests in the Bereg Lowland with its 700 ha area. It is situated among Gelénes, Tákos and Vámosatya villages. The dominant association of the area is the Querco robori-Carpinetum, but in lower parts of it, oak-ash-elm gallery forests (Fraxino pannonicae-Ulmetum) and occasionally, planted Picea abies (L.) H. Karst. patches also can be found. Both the grass and shrub-layers are dense and highly diverse. A significant part of the forest is protected and belongs to a Forest Reserve. The checklist of the area was composed based on the catchings of a Jermy-type light trap of the Forest Research Institute (ERTI), located on the northern edge of the forest, in Vámosatya village (Szanyi et al. 2015b, c, 2016).
Bátorliget (HUN)
The Fényi Forest is about 270 ha oak forest on a sandy area, which is one of the most interesting and valuable parts of the Nyírség region, located south from the Bereg region. The main forest types of the area are similar to the forests of the Bereg, with higher frequencies of Populus alba L., Tilia platyphyllos Scop., Ulmus laevis Pall., etc. The Lathraea squamaria L., Leucojum aestivum L., Listera ovata (L.) Bluff & Fingerh and Orchis militaris (L.) are all protected rarities of this strictly protected Forest Reserve. A Jermy-type light trap of the Department of Evolutionary Zoology and Human Biology of the University of Debrecen worked for 20 years in the margin of the forest (Szanyi et al. 2015b, 2016).
Lónya (HUN)
Now, the Lónya Forest is the largest forest refuge of the Bereg Lowland with a near-natural state. It covers a wide belt along the Csaronda channel east from the line formed by Lónya, Mátyus and Tiszakerecseny villages. The two main forest types of the area are the oak-hornbeam (Querco robori-Carpinetum) and the oak-ash-elm gallery forests (Fraxino pannonicae-Ulmetum), with diverse hygrophilous grass layers, which also contain species characteristic to mountainous beech and oak forests. Alder-ash forest patches also can be found in the protected forest, which is belonging to the Natura 2000 network. Data of the local fauna were assessed based on the samplings of Zoltán Varga (Szanyi et al. 2015b, 2016).
Barabás (HUN)
The sampling site was located in the Kaszony Hill, a small hill with 197 m height, which is situated between Hungarian Barabás and Ukrainian Kason villages and divided by the state border. The habitats of the Hungarian side are parts of the Natura 2000 network. The higher part of the hill forms a plateau, with steep rocky southern and eastern slopes. The diverse landscape of the hill maintains especially high habitat diversity, with different types of forests and grassland associations. Beyond the widely distributed oak forest types of the region, patches of the mixed durmast oak—silver linden forest, and the xeric durmast oak forest should be highlighted. The rocky grasslands of the eastern slope are under conservation management. The fauna was assessed based on collections of Zoltán Varga (Szanyi et al. 2015b, 2016).
Sampling methods
Data collection was carried out with different types of light traps in each studied site. The type of traps used were depended on the features of the studied habitats, the phenology and the available resources (both material and human). One of the commonly used trap types was the fixed light trap (in some cases standardized Jermy-type ones) with 125 W or 250 W mercury vapor bulbs. Mobile traps with UV light source and white sheets lighted with 125 W, 250 W or 400 W mercury vapor bulbs were also generally used. Each of these methods is widely used and accepted in faunistic and community ecological studies and even in plant protection and forestry monitoring schemes.
Data analysis
The checklists of the eight studied area were revised, corrected and uploaded in a common table. In the case of scientific names, book of Varga (2011) was followed, the whole names with the author’s name and the date of description are listed in the Annex I. The faunas of different sites were compared based on the species composition, using principal coordinate (PCoA) and cluster analysis, as multidimensional statistical methods. Because of the differences in the study intensity and the used sampling methods, only binary data of species were used in the analysis. The calculations were made with SynTax 2000 (Podani 1997a,b) program using Jaccard similarity index in both types of the analysis.
To characterize assemblage types formed by multidimensional methods, the total and the mean species richness and the ratio of species belonging to different taxa (families and subfamilies), life forms, and faunal types were used. In the case of life forms and faunal types, the work of Varga et al. (2004) was followed. Since the different assemblage could be characterized only based on data of two or three sites; thus, the statistical comparison of the assemblages based on the means of studied variables was not possible. The means of the characteristic variables were calculated and presented with MS Excel in Office 365.
Results
In the eight sampling sites, totally 479 species of the Noctuoidea superfamily were collected (Supplement I). The most species-rich family was Noctuidae with 330 species. Among them, the ratio of species belonging to subfamilies of Xyleninae, Hadeninae and Noctuinae were the highest (Table 2). Most Hadeninae and Noctuinae species feed on herbaceous plants, and prefer open habitats and forest edges, and some of them, especially the bivoltine, and migratory ones, can be seen as dangerous pests in agriculture. Contrarily, many Xyleninae species have leaf-feeding larvae living in the canopy layer of arboreal habitats and cause economic damage in forestry.
During the studies, some especially important and unexpected distribution data were also collected on some less known species, such as Dioszeghyana schmidti, which has been thought to be rare and sporadically distributed in the mixed oak forests of the southern slopes of the Hungarian mountains. However, it appeared in the fauna of Beregovo. Dicycla oo, which is characteristic species of xeric oak forests and shrub forests, and Shargacucullia gozmanyi, a characteristic species of xeric grasslands of sandy lowland areas and rocky grasslands of low mountains also live in the same habitat. Apamea syriaca tallosi, which species was already known from some habitats of the Bereg region (Szanyi et al. 2015b, d, e, 2016) was caught in all sites, except Kireshi. This species refers to the humid character of the studied habitats since this Carpathian endemic subspecies prefers wet habitats in contrast with the known related Ponto-Mediterranean-Iranian subspecies (Zilli et al. 2009).
Numerous relatively rare oligophagous leaf-feeding species generally living in the canopy layer of gallery forests (e.g., Gluphisia crenata, Earias clorana, Ipimorpha retusa, etc.) were found in all sampling sites. Additionally, the number of monophagous oak specialist species was equally high in all studied habitats. Characteristic members of this group were Moma alpium, Drymonia dodonaea, D. querna, Egira conspicillaris, Spatalia argentina, Stauropus fagi, Minucia lunaris, Notodonta tritophus, etc. The appearance of the character species of dense meadows was also an interesting and important result. Although these species, such as Autographa jota, A. pulchrina, A. bractea, Diachrysia chryson, and Lamprotes c-aureum prefer hilly and low mountain areas in the Carpathian Basin, they were absent from samples of Precarpathian hilly sites (Czorna Hora, Beregovo and Kireshi).
Based on the principal coordinate and cluster analysis performed with binary data of species distributions three types of species assemblages could be distinguished (Figs. 1 and 2). The first well-defined group is formed by the sites of Precarpathian hilly region (Beregovo, Czorna Hora and Kireshi). The especially high number of differential species of this assemblage type also showed its unique character. The second and third groups containing only lowland sites showed larger similarity and separated from the first group along the 1st Axis of the PCoA ordination. In the second group, there were three sites (Velyka Dobron, Vámosatya and Bátorliget) with gallery forest, which is characteristic for floodplains, and wet habitats, such as marshlands and moors, which abound in mountainous species. The two sites (Lónya and Barabás) of the third group showed similar but less humid character. These two groups separated along the 2nd PCoA axis which represented mainly a humidity gradient; while, the 1st Axis showed a gradient of elevation and/or distance from the Carpathians and its huge arboreal habitats. Especially low number of differential species in the Group 3 referred the relatively low species richness and high number of common species in these habitats.

Result of principal coordinate analysis (PCoA) made with Jaccard index based on checklists of the studied habitats (inf. content 1st Axis: 24.03, 2nd Axis: 18.92)

Result of the cluster analysis made with Jaccard index based on the checklists of the studied habitats
The total species richness of the assemblage types was similar, but considering the mean number of species per site, the first group showed the highest species diversity, followed by the second and third groups, respectively (Table 3). Based on the values of the Whittaker’s indices, the most species-rich group showed medium in-group heterogeneity; while, the species-poor third group was the most uniform in species composition. Each value of in-group heterogeneity was lower than the between-group value, that supported the results of the multivariate analysis (Table 3).
Studied sites can be characterized by the similar compositions of different faunal types. The widely distributed (common) eurytopic Euro-Siberian species were dominant in each assemblage type and showed a slightly higher ratio in the third group. The ratio of Boreo- and South-Continental species were also relatively high, between 17.4 and 25.9%; while, other faunal types were represented with an especially low number of species (Table 4).
The diverse biogeographical character of the fauna living in the lowland habitats was shown mainly by the South- and Central-European species, such as Tyria jacobaeae, Lygephila lusoria, Tiliacea aurago, Eugnorisma depuncta, etc. These species do not tolerate cold and humid microclimates, contrarily, the mountainous Boreo-Continental elements (e.g., Photedes extrema, Gortyna flavago, Enargia paleacea, Mniotype adusta, Mythimna impura, Diarsia brunnea) prefer these conditions.
Faunal components refer to the habitat preference of the species. Considering them separately, the ratios of the generalist, silvicolous, oakwood and steppic species were the highest, in decreasing order (Table 5). If species of deciduous forests were assumed, their ratios were extremely high compared to all other types, with values between 40.3 and 40.5%. Among this group, the eurytopic, silvicolous Catocala promissa, Meganola albula, Nola aerugula, Thalpophila matura and Acronicta auricoma, the willow-poplar Catocala electa, Catocala fraxini and Earias vernana, the nemoral Brachionycha nubeculosa and Amphipyra perflua, the Lithophane consocia, Lithophane furcifera, Furcula bicuspis and Pheosia gnoma belonging to the birch-alder type were characteristic, that referred to the high natural values of the studied habitats.
The more xeric patches of the oak forest of Beregovo and Barabás could be characterized by the occurrence of Polyphaenis sericata and Nycteola revayana belonging to the Oakwood faunal components.
Although the ratio of steppic species (e.g., Deltote deceptoria, Antitype chi, Arctia villica, Cucullia absinthii, C. fraudatrix, and Shargacucullia verbasci) was relatively low but it was higher in the assemblages of Precarpathian hilly areas (Group 1: Beregovo, Czorna Hora), where on the slopes, the ratio of mesic and meso-xerophilous grasslands (steppic- and rocky grasslands) were higher. These habitats provided suitable conditions also for migratory species, such as Dysgonia algira, Aedia leucomelas, Heliothis viriplaca, Mythimna unipuncta, Spodoptera exigua, Trichoplusia ni, etc.
Ratios of species belonging to the mesophilous (e.g., Apamea crenata, Hadena bicruris, Phytometra viridaria, etc.), and altoherbosa (e.g., Hecatera dysodea, Orthosia opima, etc.) groups, which are characteristic for natural and semi-natural, mainly humid habitats showed medium values (8.1–9.8% and 6.6–7.4%).
The ratio of arundiphilous (e.g., Rhizedra lutosa, Sedina buettneri, Mythimna straminea and Nonagria typhae, etc.) species was relatively low.
Discussion
The remains of the formerly huge and continuous forests of the northeast part of the Carpathian Lowlands and neighboring hilly areas of the Precarpathian region maintain a high-level diversity both in the flora and fauna (Simon 1952, 1953, 1960; Varga 2003; Fekete & Varga 2006). Lowlands of the region geographically belong to the Pannonian Lowland (Alföld), but they have special character considering both climate and wildlife (Hargitai 1943; Simon 1952, 1960; Baranyi 2009). The relatively cold continental climate and wide distribution of meso-hygrophilous forests differ from the other part of the lowland. To explore how the different biogeographical effects reveal in the local fauna, and which kind of assemblages lives in the remains of the former contiguous forest, Noctuoidea fauna of eight mainly forested sampling sites were compared and characterized.
The high species richness of the studied assemblages (totally 479 Noctuoidea species) proved the high natural value of their habitats, as it was shown also in the case of other invertebrate taxa, such as land snails, orthopterans, carabid beetles (Deli et al. 1997; Ködöböcz 1997; Magura et al. 1997; Deli and Sümegi 1999; Ködöböcz and Magura 1999; Gálik et al. 2001; Nagy et al. 2011, 2015; Szanyi et al. 2015a). Among studied locations, the surroundings of Beregovo showed the highest species number (362 species) (Kanarsky et al. 2011); while, the sampling site near Barabás (Kaszony Hill) proved to be the least species-rich area (Szanyi et al. 2015b, 2016). During former light trap samplings (between 1961 and 1970), the highest species richness in the Carpathian basin was recorded in the low Hungarian mountains (Varga and Gyulai 1978), but the local species richness of the sites was mainly lower than in our studied sites. This, and other unpublished data of the authors suggested that considering the Noctuoidea fauna, the northeastern part of the Lowland is one of the most species-rich parts of the Carpathian Basin.
Although the highest value of the studied fauna is the extraordinary species diversity, 42 species were found, which are protected in Hungary (Luperina zollikoferi, Mormo maura, Phalera bucephaloides, Shargacucullia gozmanyi, Tyria jacobaeae, Xestia sexstrigata etc.), and among them, 4 are also listed in Annexes of European Union Habitat Directives (Euplagia quadripunctaria, Arytrura musculus, Gortyna borelii lunata, Dioszeghyana schmidtii). Most of them were locally distributed and rare.
In the regional fauna, the ratio of species characteristic for meso-hygrophilous forests was especially higher than those, which prefer xeric forests and different steppe-like habitats. This showed that the northeast part of the Lowland and neighboring Precarpathian region belong to the continental forest climate zone rather, than to the forest-steppe climate zone (Kormány 1976), which is characteristic for the neighboring Nyírség Region.
The characteristic species of the region mainly belonged to different faunal components of deciduous forests (birch-alder, forest edge, silvicolous, willow-poplar and oakwood) preferring meso-hygrophilous habitats (marshes, meadows, gallery forests, etc.). Most of them are widely distributed Euro-Siberian, so-called "pseudomontana" species (Varga 2003), which are distributed rather in hilly areas and mountains of the Carpathian Basin (in some cases especially in areas connected to the Carpathicum), than in its forest-steppe and steppe-like habitats. These kinds of species formerly were known in the Mollusca (Deli et al. 1997), Carabidae (Coleoptera) (Magura et al. 1997; Ködöböcz and Magura 1999), and Orthoptera (Nagy et al. 2011, 2015; Szanyi et al. 2015a) fauna of the studied region.
Although the ratio of Mediterranean-Subtropic species, such as Aedia leucomelas, is still low, their appearance (regularly appeared in Beregovo, Czorna Hora, Velyka Dobron’ and Bockerek) shows the effect of climate change and the parallel spread of southern elements (Baranyi 2009), even in this continental area of Central Europe (Hickling et al. 2006; Parmesan 2006; Chen et al. 2011; Devictor et al. 2012).
During multivariate comparison of the quantitative composition of the studied sites, three types of assemblages could be differentiated. The first group contained three assemblages of the Precarpathian hilly areas, in which, the lower ratio of eurytopic generalist Euro-Siberian species, the higher ratio of the characteristic Boreo- and South-Continental steppic species, and even the especially high number of differential species (occurred only in this assemblages) referred to their unique character. These features showed that the Pannonian xerotherm steppe and forest-steppe fauna have reached the slopes of the isolated Precarpathian hills (as it also can be seen in the case of the xerotherm Quercus pubescens Willd. and Tilia tomentosa); while, remained patches of former huge forests have maintained significant part of the meso-hygrophilous Boreo-mountain element at the same time. High species diversity of the local faunas has been promoted by the overlap of the Carpathian- and Pannonian biogeographical regions, in which habitat diversity was formed and maintained by edaphic factors, long-term extensive forestry and agricultural land use.
The second assemblage type was characteristic for more humid, hygrophilous sites, among which the Bátorliget site showed the most specific character. This site is isolated as it is located in the sandy Nyírség Region, southern to the other sites. It has a special character since it is a marsh that can be considered as an ice age refugium. Thanks to these, species characteristic for marshes (e.g., Hypenodes pannonica and Arytrura musculus) and sandy areas (e.g., Cucullia balsamitae, Staurophora celsia) occurred together there.
The third assemblage type was less species-rich and had a less number of differential species. The appearance of species characteristic for hilly and low mountainous (colline) habitats (e.g., Hypena obesalis, H. crassalis, Apamea illyria, Mniotype adusta) also showed the fluctuation-zone character of the areas it was formerly detected in case of other invertebrate taxa (Deli et al. 1997; Magura et al. 1997; Ködöböcz and Magura 1999).
The studied regions have mainly lost their original character, and nowadays, only remaining patches of the former huge forests can be found. However, these patches still maintain a significant part of the fauna, with species either exclusively living there or being rare in other parts of the Pannonian Lowland.
Conclusions for future biology
The studied relatively small area has one of the most diverse Noctuoidea fauna in the Carpathian Basin, due to its unique transitional geographic location, diverse, changing land use, and climate change. To protect and further develop the natural value of the region, active and well-planned management of the remaining forest patches is essential. Their area should be maintained and even enlarged, and connections among the patches should be improved. Moreover, the two parts of the area separated by the country border require synchronized management, necessitating international cooperation involving at least the Hungarian and Ukrainian authorities, researchers (academies, universities, institutes), and even non-governmental organizations.
References
Altermatt F (2010) Climatic warming increases voltinism in European butterflies and moths. Proc R Soc B 277:1281–1287. https://doi.org/10.1098/rspb.2009.1910
Baker HG (1963) Evolutionary mechanisms in pollination biology. Science 139:877–883
Baranyi B (2009) Kárpátalja. Dialóg Campus Kiadó, Pécs–Budapest [in Hungarian]
Beck J, Linsenmair KE (2006) Feasibility of light-trapping in community research on moths: attraction radius of light, completeness of samples, nightly flight times and seasonality of Southeast-Asian hawkmoths (Lepidoptera: Sphingidae). J Res Lepid 39:18–37. https://doi.org/10.5962/p.266537
Chen IC, Hill JK, Ohlemüller R, Roy DB, Thomas CD (2011) Rapid range shifts of species associated with high levels of climate warming. Science 333:1024–1026. https://doi.org/10.1126/science.1206432
Deli T, Sümegi P (1999) Biogeographical characterisation of Szatmár-Bereg plain based on the Mollusc fauna. In: Hamar J, Sárkány-Kiss E (eds) The upper Tisza valley. Tiscia monograph series, Szeged, pp 471–477
Deli T, Sümegi P, Kiss J (1997) Biogeographical characterisation of the Mollusc fauna on Szatmár–Bereg Plain. In: Tóth E, Horváth R (eds) Proceedings of the research conservation, management” conference. ANP Füzetek, Vol I, Aggtelek, pp. 123–129
Devictor V, van Swaay C, Brereton T et al (2012) Differences in the climatic debts of birds and butterflies at a continental scale. Nat Clim Chang 2:121–124. https://doi.org/10.1038/nclimate1347
Didukh YP, Shelyag-Sosonko YR (2003) Geobotanical zoning of Ukraine and adjacent territories. Ukr Botanical J Sci J Nat Acad Sci Ukr 60(1):6–17
Gálik K, Deli T, Sólymos P (2001) Comparative malacological investigations on the Kaszonyi Hill (NE Hungary). Malakológiai Tájékoztató 19:81–88
Gerenczuk KI (1981) Priroda Zakarpatskoji oblasti. Vushcha Shkola, Lviv. (in Ukrainian)
Geryak Y (2010) The Noctuoidea (Insecta, Lepidoptera) of the Transcarpathian region. Sci Bull Uzhgorod Univ Ser Biol 29:126–139
Geryak Y (2012) New and little known species of the Noctuoidea (Lepidoptera, Insecta) for Ukrainian carpathians. Sci Bull Uzhgorod Univ Ser Biol 33:105–119
Geryak Y (2013) To the fauna of Noctuoidea (Lepidoptera, Insecta) of the Ukrainian carpathians. Sci Bull Uzhgorod Univ Ser Biol 35:73–83
Fekete G, Varga Z (eds.) (2006): Magyarország tájainak növényzete és állatvilága. MTA Társadalomkutató Központ, Budapest 460 pp.
Gy K (1976) Szabolcs-Szatmár megye éghajlata. Szabolcs-Szatmári Szemle 1:32–40 (in Hungarian)
Hargitai Z (1943) Adatok a beregi sík erdeinek ismeretéhez. Debreceni Szemle 17:64–67 (in Hungarian)
Hickling R, Roy DB, Hill JK, Fox R, Thomas CD (2006) The distributions of a wide range of taxonomic groups are expanding polewards. Glob Change Biol 12:450–455. https://doi.org/10.1111/j.1365-2486.2006.01116.x
Horváth B (2013) Diversity comparison of nocturnal macrolepidoptera communities (Lepidoptera: Macroheterocera) in different forest stands. Natura Somogyiensis 23:229–238
Kanarskyi Y, Geryak Y, Lyashenko E (2011) Ecogeographic structure of the Moth fauna in upper Tisa river basin. Transylv Rev Syst Ecol Res 11:143–168
Ködöböcz V, Magura T (1999) Biogeographical connections of the carabid fauna (Coleoptera) of the Beregi–síkság to the carpathians. Folia Entomologica Hungarica 60:195–203
Ködöböcz V (1997) Az Északkeleti-Alföld futóbogár-faunája (Coleoptera, Carabidae) és állatföldrajzi kapcsolatai. PhD thesis. University of Debrecen in Hungarian
Kristín A, Balla M, Fabriciusová V, Hrúz V, Kanuch P (2011) Orthoptera and Mantodea in fragments of seminatural habitats in lowlands of SE Slovakia and SW Transcarpathian Ukraine. Articulata 26(2):109–121
Lesku B (2008) Beregi-sík. In: Király G, Molnár Zs, Bölöni J, Csiky J, Vojtkó A (eds) Magyarország földrajzi kistájainak növényzete. MTA ÖBKI, Vácrátót
Likovitch IM (1957) Nekotorye dannye o faune prjamokrylych (Orthoptera) Zakarpatja. [some data on orthoptera fauna of Ukrainian carpathians]. Doklady i Soobscenia Uzhgorodskogo Universiteta, Seria Biologia 1:61–64 (in Russian)
Likovitch IM (1959) K voprosu o vertikalnom raspredelenii prjamokrylych (orthoptera) v Zakarpate [on vertical distribution of orthoptera in Ukrainian Carpathians]. Uzhgorodskiy Gosudarstvennyj Universitet Nauchnye Zapisky 40:227–238 (in Russian)
Lyashenko EK (2009) Species composition, abundance and hawkmoths (Lepidoptera, Sphingidae) habitat distribution of Transcarpathian region. Sci Bull Uzhgorod Univ Ser Biol 25:167–170
Magura T, Ködöböcz V, Tóthmérész B, Molnár T, Elek Z, Szilágyi G, Hegyessy G (1997) Carabid fauna of the Beregi–síkság and its biogeographical relations (Coleoptera Carabidae). Folia Entomologica Hungarica 58:73–82
Marinich AM (1982) Ukrainian carpathians. In: Marinich AM, Lanko AI, Shishchenko PG, Shcherban MIF (eds) Physical geography of the Ukrainian RSR. Vishcha shk, pp.168–176 (in Russian)
Marinich AM (1985) Ukrainian carpathians. In: Marinich AM, Pashchenko VM, Shishchenko PG (eds) Nature of the Ukrainian SSR. Landscapes and physical-geographical zoning. Nauk dumka, Kyiv, pp. 180–188 (in Russian)
Merckx T, Slade EM (2014) Macro–moth families differ in their attraction to light: implications for light–trap monitoring programmes. Insect Conserv Divers 7(5):453–461. https://doi.org/10.1111/icad.12068
Nagy A, Sz S, Molnár A, Rácz IA (2011) Preliminary data on the orthoptera fauna of the Velyka Dobron wildlife reserve (Western Ukraine). Articulata 26(2):123–130
Nagy A, Batiz Z, Sz S (2015) Orthoptera fauna of the Hungarian part of the Bereg Plain (Northeast Hungary). Bul Inf Soc Lepid Rom 26:64–80
Nowacki J, Wasala R, Zydlik P (2018) Noctuid moths of xerothermic habitats in the Chorna Hora botanical reserve in transcarpathia (Ukraine) (Lepidoptera: Nolidae, Erebidae, Noctuidae). SHILAP Revista De Lepidopterología 46(184):593–614
Nowinszky L, Puskás J (2011) Efficiency of light–traps in relation the number of caught species. Int J Sci Nat 2:161–167
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Syst 37:637–669. https://doi.org/10.1146/annurev.ecolsys.37.091305.110100
Pécsi M, Sárfalvi B (1960) Magyarország földrajza. Akadémiai kiadó. Budapest in Hungarian
Podani J (1997b) SYNTAX 5.1.: a new version of PC and Macinthos computers. Coenoses 12:149–152
Podani J (1997a) Bevezetés a többváltozós biológiai adatfeltárás rejtelmeibe. Scientia, Budapest in Hungarian
Rákosy L, Schmitt T (2011) Are butterflies and moths suitable ecological indicator systems for restoration measures of semi–natural calcareous grassland habitats? Ecol Ind 11:1040–1045. https://doi.org/10.1016/j.ecolind.2010.10.010
Simon T (1952) Montán elemek az Észak-Alföld flórájában és növénytakarójában. Ann Biol Univ Debreceniensis 1:146–174 (in Hungarian)
Simon T (1960) Die Vegetation der Moore in den Naturschutzgebieten des Nördlichen Alföld. Acta Bot Hungar 6:107–137
Simon T (1953) Az Északi-Alföld erdői. Akadémiai Kiadó, Budapest
Summerville KS, Ritter LM, Crist TO (2004) Forest moth taxa as indicators of lepidopteran richness and habitat disturbance: a preliminary assessment. Biol Cons 116:9–18. https://doi.org/10.1016/S0006-3207(03)00168-X
Sz S (2012) A Nagydobronyi Vadvédelmi Rezervátum és környéke nagylepkefaunája (Macrolepidoptera). Állattani Közlemények 97(2):171–180 (in Hungarian)
Sz S (2015b) The night-active Macroheterocera of a Transcarpathian forest reserve. Bull De Inform Soc Lepidopterol Române 26:82–91
Sz S, Katona K, Rácz IA, Varga Z, Nagy A (2015a) Orthoptera fauna of the Ukrainian part of the Bereg plain (Transcarpathia, Western Ukraine). Articulata 30(1):91–104
Sz S, Szőcs L, Gy C, Varga Z (2015b) A Beregi–sík Noctuoidea (Lepidoptera: Macroheterocera) faunájának állatföldrajzi és ökológiai jellemzése. Állattani Közlemények 100(1–2):89–100 (in Hungarian)
Sz S, Szőcs L, Varga Z (2015c) A Bockerek–erdő Macroheterocera faunájának állatföldrajzi és ökológiai jellemzése. Erdészettudományi Közlemények 5(1):119–128 (in Hungarian)
Sz S, Nagy A, Varga Z (2016) Comparison of Macroheterocera assemblages of four forests in the Bereg plain (Hungary, Ukraine). Biologia 71(11):1281–1291. https://doi.org/10.1515/biolog-2016-0154
Sz S, Nagy A, Molnár A, Katona K, Tóth M, Varga Z (2017) Night–active Macroheterocera species in traps with synthetic attractants in the Velyka Dobron’ Game Reserve (Ukraine, Transcarpathia). Acta Zool Acad Sci Hungaricae 63(1):97–114
Sz S (2015a) Egy kárpátaljai erdőrezervátum jellemzése az éjjeli nagylepkefauna alapján. E-Acta Nat Pannonica 8:91–110 (in Hungarian)
Truxa C, Fiedler K (2012) Attraction to light—from how far do moths (Lepidoptera) return to weak artificial sources of light? Eur J Entomol 109:77–84
Tsys PN (1968) Region of the Transcarpathian Plain. Physico-geographical zoning. Ukrainian SSR, Kyiv Publishing House, Kyiv University, pp. 634–636 (in Russian)
Usher MB, Keiller SWJ (1998) The Macrolepidoptera of farm Woodlands: determinants of diversity and community structure. Biodivers Conserv 7:725–748. https://doi.org/10.1023/A:1008836302193
Varga Z (ed) (2011) Magyarország nagylepkéi—macrolepidoptera of hungary. Heterocera Press, Budapest (in Hungarian)
Varga Z, Gyulai I (1978) Die faunenelemente-einteilung der noctuiden ungarns und die verteilung der faunenelemente in den lokalfaunen. Acta Biologica Debrecina 15:257–295
Varga Z, Ronkay L, Bálint Zs, Gyula LM, Peregovits L (2004) Checklist of the fauna of Hungary. Volume 3. Macrolepidoptera. Hungarian Natural History Museum, Budapest
Varga Z (2003) A Kárpát–medence állatföldrajza. In: Láng I, Bedő Z, Csete L (eds) Növény, állat, élőhely. Magyar Tudománytár III, pp 89–119
Végvári Zs, Juhász E, Tóth JP, Barta Z, Boldogh S, Szabó S, Varga Z (2014) Life–history traits and climatic responsiveness in noctuid moths. Oikos 124(2):235–242. https://doi.org/10.1111/oik.01655
Yela JL, Holyoak M (1997) Effects of moonlight and meteorological factors on light and bait trap catches of noctuid moths (Lepidoptera: Noctuidae). Environ Entomol 26:1283–1290. https://doi.org/10.1093/ee/26.6.1283
Zilli A, Varga Z, Ronkay G, Ronkay L (2009) The witt catalogue—a taxonomic atlas of the Eurasian and North African Noctuoidea, Vol. 3, Apameini. Heterocera press, Budapest
Zs V, Juhász E, Tóth JP, Barta Z, Boldogh S, Szabó S, Varga Z (2014) Life–history traits and climatic responsiveness in noctuid moths. Oikos 124(2):235–242. https://doi.org/10.1111/oik.01655
Acknowledgements
Authors thank Attila Molnár, Krisztián Katona for their help in the field works, and Prof. Dr. Zoltán Varga for inspiration and useful assistance. Szabolcs Szanyi’s research was financed by the National Research Development and Innovation Office (NKFIH, grant PD 138329).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
No ethical approval is required.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Szanyi, S., Szanyi, K. & Nagy, A. Changes of the composition of the Noctuoidea fauna along a geographical gradient in the northeastern part of the Carpathian Lowland. BIOLOGIA FUTURA 74, 433–443 (2023). https://doi.org/10.1007/s42977-024-00210-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42977-024-00210-w