
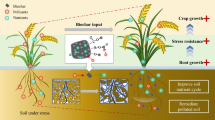
Abstract
Plants regulate root exudates to form the composition of rhizosphere microbial community and resist disease stress. Many studies advocate intervention with biochar (BC) and exogenous microbe to enhance this process and improve plant defenses. However, the mechanism by which BC mediates exogenous microorganisms to enhance root exudate-soil microbial defensive feedback remains unclear. Here, a BC-based Bacillus subtilis SL-44 inoculant (BC@SL) was prepared to investigate the defensive feedback mechanism for plants, which enhanced plant growth and defense more than BC or SL-44 alone. BC@SL not only strengthened the direct inhibition of Rhizoctonia solani Rs by solving the problem of reduced viability of a single SL-44 inoculant but also indirectly alleviated the Rs stress by strengthening plant defensive feedback, which was specifically manifested by the following: (1) increasing the root resistance enzyme activities (superoxide dismutase up to 3.5 FC); (2) increasing the abundance of beneficial microbe in soil (0.38–16.31% Bacillus); and (3) remodeling the composition of root exudates (palmitic acid 3.95–6.96%, stearic acid 3.56–5.93%, 2,4 tert-butylphenol 1.23–2.62%, increasing citric acid 0.94–1.81%, and benzoic acid 0.97–2.13%). The mechanism reveals that BC@SL can enhance the positive regulatory effect between root exudates and microorganisms by optimizing their composition. Overall, BC@SL is a stable and efficient new solid exogenous soil auxiliary, and this study lays the foundation for the generalization and application of green pesticides.
Graphical Abstract

Highlights
-
The problems of effective viable bacteria easy loss were solved by BC@SL.
-
Rs stress was relieved by BC@SL through direct inhibition and defensive feedback.
-
BC@SL optimized defensive feedback by reshaping root exudates and microbiota.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Crop losses caused by plant diseases have become a global problem, and the average yield losses of major food crops range from 17% to 30% (Andreo-Jimenez et al. 2021). For example, R. solani is a widespread soil-borne pathogen which can infect a wide range of plants such as Cruciferae, Gramineae, Solanaceae, etc. (Liu et al. 2020; Pedras and Okanga 1999). R. solani is one of the most common diseases of Brassica chinensis L., which causes root rot, leaf rot and even death of the plant, seriously jeopardizing Brassica chinensis productivity (Pedras and Okanga 1999). Those soil-borne pathogens could destroy soil microecology and force plant exudates and rhizosphere microorganisms to lose their self-regulation ability (Liu et al. 2020). In recent years, many studies have reported complex synergistic interactions between plant defense systems and soil microbial communities against pathogens, suggesting that beneficial microorganisms in plant roots could enhance plant stress tolerance (Liu et al. 2020; Wei et al. 2020). The beneficial microbe recruitment is pivotal to plants’ defense system (Shalev et al. 2022; Wen et al. 2023), among which beneficial microorganisms, such as Bacillus (Wen et al. 2023), Flavobacterium (Kwak et al. 2018) and Pseudomonas (Shalev et al. 2022), could protect plants from fungal pathogens aggregating in the rhizosphere and endosphere. In addition, many studies have shown that root exudates are the main measures for plants to remodel rhizosphere soil microflora, including sugars, amino acids, organic acids, fatty acids, and secondary metabolites (Wen et al. 2023; Zhalnina et al. 2018) and playing a role in the prevention and control of pathogenic microorganisms (Hu et al. 2018; Zhalnina et al. 2018). Thus, considering the large differences in root exudates and their microbial communities in different degrees, optimizing root exudates and microbial communities in plant disease resistance is complicated (Hu et al. 2018). Therefore, how to optimize the rhizosphere microbial community and regulate key root exudates for plant disease resistance is a key central issue addressed in this study.
Many scientists have recently explored the production of antimicrobial metabolites and the recruitment of beneficial microorganisms in rhizosphere exudates to further optimize the root exudate–soil microbial community defensive feedback (Dang et al. 2023; Sun et al. 2020). For example, Dang et al. (2023) explored the difference of root exudates and soil microbial communities under Ralstonia solanacearum stress and verified the properties of root exudates to recruitment beneficial microbes and inhibit R. solanacearum. However, many discrepancies have been found in some studies, where numerous antimicrobial metabolites have dual inhibitory effects, inhibiting the growth of beneficial microorganisms in addition to pathogens (Ayaz et al. 2008; Suarez-Fernandez et al. 2020). Phenolic compounds are known to have an inhibitory effect on Fusarium oxysporum (Suarez-Fernandez et al. 2020). Excessive secretion of phenolic compounds could also inhibit the growth of Bacillus (Ayaz et al. 2008). Unexpectedly, Yuan et al. (2018) reported that phenolic compounds could enhance the root colonization of Bacillus at appropriate concentrations. Based on this literature review, regulating root exudate metabolites to near-appropriate levels and inhibiting pathogenic bacteria without damaging beneficial microorganisms are extremely complex processes in different crop rhizospheric environments. Therefore, developing novel technologies for controlling the root exudate–soil microbial community defensive feedback is considerably necessary.
Although plants could regulate the root exudate–soil microbial community defensive feedback by themselves, numerous studies have demonstrated that abiotic factors (Ahmed et al. 2023; Wang et al. 2019; Wei et al. 2020) and exogenous inoculants (Kwak et al. 2018; Shalev et al. 2022) contribute to feedback optimization and improve crop disease prevention. Such biotic and abiotic exogenous addition to protect plants from pathogens stress caused widespread attention. Bacillus, a plant growth-promoting rhizobacteria (PGPR), has been used as a biotic exogenous addition. It could be recruited by organic acids, amino acids, and flavonoid compounds from root exudates (Sharma et al. 2020; Wen et al. 2023; Zhou et al. 2023a) and produce antifungal compounds, such as surfaceacins, iturin, and fengycin, to further enhance plant disease resistance (Huang et al. 2017; Li et al. 2021). Some researchers have shown that exogenous beneficial microbes could help plants enhance the defense system (Shalev et al. 2022; Wu et al. 2022). However, the application has some limitations, such as microorganisms’ susceptibility to environmental influences, weak adaptability, and slow proliferation, resulting in low survival ratio and thus failure to play an excellent role in disease resistance (Maddalwar et al. 2021; Wu et al. 2022). Biochar (BC) has become an excellent soil abiotic exogenous addition worldwide due to its low cost, good adsorption performance, and strong oxidation reduction capacity (An et al. 2022a, 2022b; Wu et al. 2022). Sun et al. (2020) and Kolton et al. (2016) reported that BC could stimulate the production of organic acids in root exudates and increase the abundance of microorganisms such as Actinobacteria and Bacteroidetes in the soil. Since BC and PGPR can modulate root exudates and microbial communities, BC and PGPR synergy has been hypothesized to potentially optimize root exudate-soil microbial community defensive feedback to achieve better results than applying PGPR and BC separately.
Nowadays, integrating BC and PGPR to improve soil microecology, enhance plant disease resistance, promote plant growth, and repair soil has become a research hotspot (Wei et al. 2020). Many studies have explored the soil microbial community and soil metabolomics under the synergistic effect of BC and PGPR (Wu et al. 2022). However, root exudates as a key factor in soil microbial regulation are still not included in BC and PGPR synergy studies. In view of this, the utility and interaction between key root exudates and key microorganisms in defensive feedback must be determined under the synergistic effects of BC and PGPR, which may be beneficial in optimizing root exudate–microbial community defensive feedback.
In this study, PGPR B. subtilis SL-44 and BC were selected to prepare the solid BC-based SL-44 inoculant (BC@SL) to control R. solani Rs invasion in Brassica chinensis L. First, the morphology, physicochemical structure, preservation, microbial inhibition, and plant growth-promoting properties of BC@SL were analyzed. Next, the rhizosphere microbial community and key root exudates were identified, and their key roles were further explored. Finally, the regulation of Rs and SL-44 by appropriate levels of root exudate metabolites was explored, revealing the mechanism and effect of BC@SL in optimizing root exudate–microbial community defensive feedback.
2 Materials and methods
2.1 Microbial strains and BC@SL preparation
The biocontrol-promoting strain B. subtilis SL-44 and the pathogen R. solani Rs were obtained from Shihezi University, Xinjiang, China. The BC was purchased from Henan Zhongbang Protection Technology Co., China and modified with amino. Additional file 1: Table S1 shows the elemental composition. The culture processes of SL-44 and Rs strains are provided in Additional file 1: Text S1. BC@SL was prepared as follows: the resuspended SL-44 was first mixed with BC in sterile condition at a ratio of 1:5 (w/v), and then the mixture was shaken and cultivated for 4 h at 28 °C. Finally, the mixture was spread out onto trays and subsequently dried in a vacuum oven for 12 h at 50 °C.
2.2 Inhibition performance and sample characterization
The effect of SL-44 and BC on Rs colony size was explored and observed on PDA. The cell dry weight method (Mills & Lee 1996) was selected to further quantify and observe the inhibition ability of BC and SL-44 on Rs, and its detailed determination method is described in Additional file 1: Text S2. The morphology of BC@SL was characterized using SEM (FlexSEM1000, Japan). The BC and BC@SL group information were determined by FTIR (Nicolet 5700, Japan) and XPS spectra (Thermo Escalab 250Xi, USA). TGA (SDTA851e, Switzerland) was used to quantify the weight of microbial biomass at a heating rate of 10 °C min–1 and a temperature range of 25–800 °C. Zeta potential was measured by a Zetasizer Nano ZS90 (Malvern Instruments, UK) at the ratio of 1:100 (w/v).
2.3 Pot experiment and root enzyme activity associated with plant resistance
Potting soil was obtained from the pine forest of Xian Polytechnic University, China, and classified as alkaline soil. The soil does not contain Rs. The physical and chemical properties are shown in Additional file 1: Table S2. Brassica chinensis L. seeds were sterilized using 70% alcohol. Five treatments were set up, with six replicates per treatment, and eight seeds were seeded in each pot with 700 g soil. The corresponding treatments were labeled as CK, RsT, BCRs, SLRs, and BC@SLRs. The five treatments of pot experiment included CK without BC, SL-44, and Rs; RsT with 30 mL Rs; BCRs with 7 g BC + 30 mL Rs; SLRs with 30 mL SL-44 + 30 mL Rs; and BC@SLRs with 7 g BC@SL (prepared with 6 g BC and 30 mL SL-44) + 30 mL Rs, in which 30 mL Rs fungal suspension was poured in soils of pots at the end of the fourth week. The growth indices, including height, root length, fresh weight, and dry weight, of plants were measured on the fifth weekend (Dixit et al. 2016). The disease index of 20 Brassica chinensis L. plants was recorded 1 week after Rs inoculation (fifth weekend) and the detailed description of the measure of disease index are shown in Additional file 1: Text S3.
Plant roots (0.2 g) were placed into a pre-chilled mortar with PBS buffer (pH 7.4) on the fifth weekend and grinded until they became homogeneous. Then, the grinding fluid was transferred into a 10 mL EP tube and centrifuged, and the supernatant of the crude enzyme solution was extracted and stored at 4 °C. The antioxidant enzyme activities of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) were measured using NBT photoreduction, guaiacol, and decline-in-H2O2-absorbance methods, respectively(Zhao et al. 2019). Finally, the defense enzyme activity of chitinases (Kadoo and Badere 2017) and β-1,3 glucanase (Ren et al. 2016) were estimated in accordance with the ratio of reducing sugar production, with laminarin and glucose as the substrate.
2.4 Quantity of SL-44, Rs, and microbial community analysis
The quantity of SL-44 in SLRs and BC@SLRs was determined at 0, 1, 2, 3, 4, 5 weeks, and the quantity of Rs in the RsT, SLRs, BCRs, and BC@SLRs was measured at 0 and 5 weeks by absolute quantification PCR (AQ-PCR). Primers were designed to detect SL-44 and Rs by the 16S rRNA genes of B. subtilis and the 18 s rRNA genes of R. solani, acquired from GenBank. The primers SF (5′- GCGTAGAGATGTGGAGGAA -3′) and SR (5′- TAGGATTGTCAGAGGATGTCA -3′) were used for detecting SL-44, and the primers RF (5′- AGGGTGTCCTCAGCGATAGA -3′) and RR (5′- TCCTTGGAGCATGCCTGTTT -3′) were used for detecting Rs. The melting point curves of SL-44 and Rs are single peaks in Additional file 1: Figure S3 a, b, indicating that the primers had good specificity.
Rhizospheric soil was collected from CK, RsT, SLRs, BCRs, BC@SLRs and bulk soil from BCK, BRsT and BBC@SLRs. These samples were subsequently analyzed using high-throughput sequencing of 16 s and 18 s rRNA genes. Total DNA was extracted using a DNA extraction kit for high-throughput sequencing of 16 s and 18 s rRNA genes. After the genomic DNA was extracted, DNA integrity, purity, and concentration were determined by ω = 1% agarose gel electrophoresis. Macroscopic genomic DNA was used as a template based on the sequencing of bacterial V3V4 and fungal V4 in the region. The primers (338F-806R) were used for PCR amplification. Following the standard flow of NEBNext Ultra DNA Library Prep Kit for Illumina (New England Biolabs, USA), PE 250 sequencing was performed on the Illumina Hiseq 2500 platform.
2.5 Collection and discrimination of root exudate metabolites
The Brassica chinensis L. seedlings were intactly removed from the CK, RsT, and BC@SLRs to collect root exudates. The Brassica chinensis L. root was submerged in a conical flask (three plants per bottle) containing 50 ml of 0.9% sodium chloride solution. The conical flasks were randomly placed in the growth chamber and photosynthesis was performed for 4 h and the collected root exudates were freeze-dried and concentrated to 10 ml. The microbial cells and root tissues were filtered with a polyether sulfone membrane (0.45μm) and stored in a refrigerator at −20 ℃.
Then, total organic carbon (TOC) was used to represent the root exudate content, and its concentration was quantified by Shimadzu TOC-L Analyzer (Muti N/C 2100). The contents of soluble sugars, phenolic acids, and proteins in the secretions were determined by spectrophotometers (UC 1600) at specific wavelengths, as described in Additional file 1: Text S4. The root exudates were extracted by ethyl acetate and then were discriminated by GC–MS (Agilent 8890-7000D). Details of the extraction and GC–MS are provided in Text S5. The NIST98 mass spectrometry database was used to analyze the mass spectra and determine the substance names of each component.
2.6 Response of SL-44 and Rs to root exudates
A 3 mm sterile filter paper was placed in the center of a water agar (0.5%, w/v) containing root exudates (20 µg ml–1) of CK, RsT, and BC@SLRs. 10 μl SL-44 bacterial suspension and Rs cells suspension were added to the center of the filter paper, then cultured in a biochemical incubator at 28 °C for 4 days and colony diameters were observed.
The effects of CK, RsT, BC@SL root secretion on the growth of SL-44 and Rs were studied in LB and PDB containing 20 µg ml–1 root exudates, respectively. Subsequently, SL-44 and Rs were inoculated with corresponding medium at 1% inoculum, incubated for 24 h (SL-44) and 48 h (Rs) at 28 °C, respectively. The Rs was quantified by cell dry weight method (Mills and Lee 1996) and SL-44 was determined by Bio-Rad protein assay (Bradford 1976) with the same procedure as the previous method. SL-44 colony count was calculated from standard curve: colony count (107 CFU ml–1) = 48.49 107 CFU ml–1 × OD595.
The main metabolites in the exudates affecting SL-44 and Rs were screened, including palmitic acid, stearic acid, benzoic acid, 2,4 tert-butylphenol, and citric acid. Additional file 1: Figure S1a illustrates the screening method. These substances were dissolved in acetone solution (100 mM) and then added into LB and PDB with different concentrations (2,4-tert-butylphenol and citric acid 0, 0.10, 0.25, 0.50, 0.75, 1.00 mM; palmitic acid, stearic acid, benzoic acid 0, 0.25, 0.50, 1.00, 2.00, 3.00 mM; general system 20 ml). The methods used to count the cells of SL-44 and Rs were Bio-Rad protein assays and cell dry weighting, respectively. Additional file 1: Figure S1b, c shows that the acetone solution does not affect the growth of SL-44 and Rs by determining the amount of growth.
2.7 Statistics
A one-way ANOVA was performed to assess the differences among the biological duplicates. In addition, the LSD-test was used to detect differences between treatments. On the basis of the results of high-throughput sequencing, QIIME2 (2019.4, version 1.30.1) was used for alpha diversity and Shannon index analyses and drawing the sparse curve. The taxonomic composition of species was analyzed using the classification-sklearn algorithm. The Bray–Curtis distance matrix was calculated using the leveled ASV/OTU table, and the distance matrix was analyzed by principal coordinate analysis (PCoA). Three replicate treatments were set up for each experiment.
3 Results and discussion
3.1 Preparation and performance measurement of BC@SL
The surface morphology of SL-44 is shown in Fig. 1a. SL-44 shows a short oval rod-like structure with a length of about 1.0 µm. The inhibitory effect of BC and BC@SL on Rs is verified by plate culture, and Rs grew very well in the medium (Fig. 1b1). Figure 1b2 shows the Rs is inhibited by BC, and Fig. 1b3 shows an inhibition circle of Rs is formed around SL-44 and BC@SL, but not BC. Huang et al. (2017) and Poveda et al. (2021) also reported that BC and Bacillus inhibit pathogens, respectively. Hence, BC coupled with SL-44 to prepare BC@SL is expected to achieve enhanced anti-pathogenic effect. To further explore the anti-pathogenic performance, the dry weights of Rs in PDB containing SL-44, BC, and BC@SL were measured, which were 0.15 ± 0.007b g, 0.12 ± 0.007c g, and 0.10 ± 0.011d g, respectively, significantly lower than that of individual culturing Rs at 0.18 ± 0.014a g, indicating that BC@SL enhances the ability to inhibit Rs. Additional file 1: Figure S2a-c shows that BC has rough surface and abundant pore structure, specific surface area of BC is 562 m2 g–1, and the average pore size is 3.28 nm, which could be considered as the carrier (Wu et al. 2022). As could be observed from Fig. 1c, SL-44 is loaded on the surface or internal pores of BC. The corresponding energy-dispersive spectroscopic mappings (Fig. 1d) demonstrate that C and O are evenly distributed over the BC@SL surface (Additional file 1: Figure S2d). Additional file 1: Figure S2e shows that BC@SL loses about 5% more weight than BC at 800 °C, and this weight loss comes from SL-44.

a SEM image of SL-44; b Growth of Rs on PDA (b1, b3) and BPDA (b2, PDA contained 1%BC); c SEM image of BC@SL; d EDS and corresponding elemental mapping of BC@SL; e FTIR spectra of BC and BC@SL; f XPS spectrum of C1s for BC@SL; g XPS spectrum of O1s for BC@SL; h Zeta potential of SL, BC and BC@SL; i The count and survival rate of bacteria inoculants SL-44 and BC@SL
The functional group of BC combined with SL-44 was investigated by FTIR and XPS (Fig. 1e–g, Additional file 1: Fig. S2f-i). The FTIR results showed that the characteristic peaks of the -OH (3446 cm−1), C=O (1642 cm−1), and P=O (1252 cm−1) functional groups of BC@SL remarkably shifted compared with those of BC and SL-44 (Adeyem and Arturo 2016; Liu et al. 2019). In the process of BC coupling with SL-44, the COOH on the BC surface could react with the -NH2 and P=O of SL-44 to form amide and phosphate complexes, respectively (Adeyem and Arturo 2016; Liu et al. 2019). The XPS spectra demonstrated that BC@SL had lower C and higher O than BC, with -OH decreasing from 17.57% to 24.49%, indicating that more oxygen-containing functional groups were generated during the fixation of SL-44 to BC. The peak sites of C=O and C–O were also changed from BC to BC@SL, which are important in the combination process of BC and SL-44 (Zhou et al. 2023b). Figure 1h shows SL-44 was loaded into BC to form BC@SL, with its zeta potential value in the middle. Analysis of FTIR, XPS, and zeta potentials showed that SL-44 was successfully and robustly loaded to BC. The stability of inoculants is shown in Fig. 1i. The viable bacterial number of BC@SL (9.62–9.33 log10 CFU·g−1) was significantly higher than that of SL-44 (8.83–7.80 log10 CFU·mL−1). The bacterial survival rate of BC@SL was 51.07%, whereas that of SL-44 was only 9.35%. These findings demonstrate that BC@SL was more stable than the liquid microbial inoculant SL-44, and such stability could be conducive to the preservation and transport of inoculants (Zhao et al. 2022).
3.2 Improvement in plant growth and defensive enzyme activity by BC@SL
The quantity of SL-44 and Rs was measured by AQ-PCR to investigate the inhibition and stability performance of BC@SL in soil under Rs treatment. As shown in Fig. 2a, the concentration of SL-44 was positive in BC@SLRs and negative in SLRs, indicating that BC@SL was more stable, which is consistent with the result in Fig. 1i. On the fourth weekend, when Rs was added into the soil, the amount of SL-44 decreases significantly (Additional file 1: Figure S3c). On the contrary, the quantity of SL-44 decreased slightly in BC@SL. A significant increase was observed in the number of viable bacteria in SL and BC@SL on the fifth weekend, which may be because (1) fungal mycelium secretes rich enzymes, metabolites, antibiotics, etc. to form a microenvironment for microbial growth around fungal mycelium, which is called mycelium, providing nutrients for bacterial growth (Zhang et al. 2022) and (2) BC could provide shelter to SL-44 and increase its number (Zhao et al. 2022) on the fifth weekend compared with the initial value.

a The changes of Rs and SL-44 measured between initial value and 5th weekend (Rs were added to soil at 4th weekend); b Plant biomass expressed as dry weight at the 5th weekend; c The fold change of SOD, POD and CAT and in RsT, BCRs, SLRs and BC@SLRs relative to CK; d The fold change of chitinase activity and β-1, 3 glucanase activity in RsT, BCRs, SLRs and BC@SLRs relative to CK. Stars indicate significant differences between treatments and CK (ANOVA, * P < 0.05; ** P < 0.01; *** P < 0.001, n = 3)
Figure 2a and Additional file 1: Figure S3d show that the number of Rs decreased in all treatments and conformed to the following order: RsT < BCRs < SLRs < BC@SLRs, which is relevant to the count of SL-44 in soil as it could inhibit the growth of Rs. In addition, in the initial stage of BC@SLRs, the count of Rs reached 13.99 × 104 copy g–1, but in the final stage, the count of Rs only reached 8.40 × 104 copy g–1, indicating that BC could increase the large amount of SL-44 available and reduce Rs. The Brassica chinensis L. biomass in Fig. 2b and Additional file 1: Figure S4a is RsT < BCRs < CK < SLRs < BC@SLRs. BC and B. subtilis have been reported to alleviate pathogen stress (Huang et al. 2017; Poveda et al. 2021), so BCRs and SLRs have higher biomass than RsT. Wei et al. (2020) has demonstrated that BC-inoculated Pseudomonas putida Rs-198 improved grape quality compared with BC and Rs-198 alone. In brief, BC@SL significantly increased the plant height, root length, fresh weight, and other indicators (Additional file 1: Table S3). The disease indices for plants in RsT, BCRs, SLRs and BC@SLRs were 63.75%, 47.50%, 45.00% and 32.50%, respectively, indicating that the addition of BC@SL significantly reduced the level of plant morbidity. These results reveal that BC@SL had not only anti-pathogenic performance but also excellent growth-promoting properties.
As shown in Fig. 2c, the antioxidant enzymes were positive in all treatments after Rs invasion compared with CK. Pathogen stress causes an increase of reactive oxygen species (ROS) (Kong et al. 2022). Plants maintain in redox homeostasis by increasing SOD, POD, and CAT enzyme activities to eliminate excess ROS (Kong et al. 2022). These antioxidant enzymes that act synergistically in plants are the important members of the cellular defense enzyme system (Bogoutdinova et al. 2020; Zhao et al. 2019). Therefore, RsT, BCRs, SLRs, and BC@SLRs enhanced the SOD, POD, and CAT enzyme activities (Bogoutdinova et al. 2020). The specific values of the enzyme activities are demonstrated in Additional file 1: Figure S4b–d. Typically, the levels of antioxidant enzymes activities increase with enhance extrinsic stress. However, in Fig. 2c and Additional file 1: Figure S4b-d, SOD, POD, and CAT enzyme activities are RsT < BCRs < SLRs < BC@SLRs, and the SOD of BC@SLRs reaches 3.5-FC that of CK. This result is opposite to the Rs stress to each treatment (Fig. 2a). BC and PGPR have been reported to promote plant antioxidant enzyme activities (Kong et al. 2022; Wen et al. 2023).
The fold changes of the disease-resistant enzymes chitinase and β-1,3 glucanase are shown in Fig. 2d. The changes in each treatment after Rs invasion were more than 1.5 FC, which could be explained by the fact that plant root enzymes protect plants by destroying the mycelium and cell wall of fungi (Kadoo & Badere 2017; Kandoliya et al. 2017). In addition, the enzyme activities of BCRs, SLRs, and BC@SLRs were significantly decreased, indicating that the improvement of plant disease resistance by BC@SL does not depend on the improvement of disease resistance enzyme activity (Fig. 2d, Additional file 1: Figure S4e, and S4f). These results demonstrate that BC could provide shelter for SL-44 to better cope with external environmental stresses (Zhao et al. 2022), with a more stable population of anti-pathogenetic bacteria than SL-44 applied alone. In addition, BC@SL promoted plant roots to secrete more antioxidant enzymes in response to Rs stress.
3.3 Enhanced remodeling ability of microbial communities by BC@SL
Additional file 1: Table S4 shows that the alpha diversity index of bacteria was higher than that of fungi, with similar trends for bacteria and fungi in the BC, SL-44, and BC@SL treatments. However, the Chao1 and Shannon indices of RsT, BCRs, and SLRs decreased distinctly compared with those of CK, indicating that soil microbial diversity and abundance decreased (Han et al. 2018). The indices of BC@SLRs were larger than those of RsT, BCRs, and SLRs (Chao1 index of bacteria, 16.74–22.57%; Shannon index of bacteria, 2.22–4.53%), indicating that BC@SL could improve the abundance and diversity of soil microorganisms compared with BC and SL-44. Sun et al. (2020) and Shalev et al. (2022) reported that BC and exogenous inoculants could reshape microbial communities, respectively. In the present study, BC@SL enhanced the ability to reshape the microbial community structure and alleviated the destruction of soil microbiology by pathogenic bacteria.
Figure 3a, b and Additional file 1: Figure S5a-b show the phyla and genera in bacteria and fungi. After Rs was added into soil, the number of Actinobacteria, Chloroflexi, Acidobacteria in soil bacterial phylum decreased significantly (Fig. 3a). Plant responses to Rs invasion are mediated by inter-root regulation, such as the enrichment of beneficial microorganisms (Dang et al. 2023; Wu et al. 2022; Zhalnina et al. 2018). For example, Firmicutes is the phylum with the largest variation in soil abundance after the addition of Rs. The dominant genus of Firmicutes is Bacillus that inhibits Rs growth (Sun et al. 2020), accounting for 69.09% of the total Firmicutes in CK, whereas it accounts for about 84% of the total Firmicutes in the other treatments. Figure 3a, Additional file 1: Figure S5a shows the abundances of Bacillus in CK, RsT, BCRs, SLRs, BC@SLRs as 0.38%, 6.58%, 10.16%, 12.45%, and 16.31%, respectively. The count of Bacillus has great variety (from 0.38% to 16.31%) due to inter-root regulation of plants, so Bacillus has been inferred to be one of the main microorganisms controlling the invasion of Rs. Ascomycetes is a common phylum of terrestrial fungi (Mtibaà et al. 2018). Ascomycota is the most dominant phylum in soil, with a relative abundance ratio of > 90%. In addition, most saprophytic fungi, such as Chaetomium (Mtibaà et al. 2018), account for more than 35% of the abundance of Ascomycota in this sequence (Fig. 3b, Additional file 1: Figure S5b). The other two major phyla are the Basidiomycota and Mortierellomycota, whose abundances were increased by the inclusion of BC@SLRs. However, Basidiomycete showed a reduction in BC@SLRs for Rhizoctonia, which is consistent with Fig. 2a, suggesting that BC@SL may regulate the fungal community and protect the plant.

Relative abundance of dominant phyla levels; the top 10 dominant phyla accounting for more than 90% of the overall operational taxonomic units (OTUs): a Bacteria; b Fungi. Principal coordinates analysis (PCoA) based on microbial OUT levels;c Bacteria; d Fungi
The PCoA of bacterial and fungus communities showed three distinct clusters, in which the treatments were significantly different from CK (Fig. 3c, d). A difference could also be observed among different treatments under Rs stress, indicating that the microbial community after Rs invasion is different from that of healthy samples. The distance between BC@SLRs and RsT was larger than the distance between BCRs, SLRs and RsT, revealing that the addition of BC@SL reshaped the soil microbial structure. The results are consistent with the alpha diversity analysis and the relative abundance of species at the phylum and genus level.
The structure of the bulk soil microbial community was also determined to analyze differences in the rhizospheric microbial community. The alpha diversity indices demonstrated that microorganisms in the rhizosphere were more abundant and diversified than those in the bulk soil (Additional file 1: Table S4). Bacterial genera, including Pseudonocardia, Solirubrobacter, and Truepera, have thermophilic and aerobic habits, and fungal genus, such as Chaetomiumcan, is highly abundant in soil and could directly use cellulose as a carbon source (Hao et al. 2021; Zhalnina et al. 2018) (Additional file 1: Figure S5a-b). The lack of readily degradable sources of carbon produced by plant root exudates in bulk soil is one of the reasons for the difference (Hu et al. 2018). Figure 3c, d show a cluster of bulk soil alone, indicating significant differences in the rhizosphere, highlighting the importance of root exudates to the microbial community. This finding suggests that root exudates have a non-negligible effect on soil microbial community structure (Dang et al. 2023; Hu et al. 2018). Therefore, the effect of root exudates should also be considered, and the effect of exogenous auxiliaries (BC, SL-44, and BC@SL) on plant resistance to pathogens should be studied.
3.4 BC@SL regulation of root exudates to resist pathogens
The exudate metabolites from CK, RsT, and BC@SLRs could be classified into seven categories, including organic acid, phenol, lipid, ketone, alcohol, amide, and alkane (Fig. 4a–c and Additional file 1: Table S5). Higher levels of organic acid and phenol were found in RsT than in CK and BC@SLRs (CK < BC@SLRs < RsT), as plants usually respond to pathogen invasion by increasing organic acids (Dang et al. 2023; Zhalnina et al. 2018). The secretion of organic acids and phenols in BC@SLRs was reduced, which was related to the decreased Rs (Fig. 2a) and the root secretion regulation by BC@SLRs. Additional file 1: Figure S6a shows the metabolite profiles of the PCoA based exogenous auxiliaries with sharp differences. The root exudates of the different treatments differed significantly, with BC@SLRs closer to CK than RsT. This phenomenon may be caused by reduced pathogen stress in BC@SLRs, indicating the exudate regulation capacity of BC@SL.

Composition analysis of root exudates. a-c The proportion of 49 root exudates with high content and matching quality higher than 70 in different treatments, a CK, b RsT and c BC@SLRs; The log2 (FC) of 27 significantly different root exudates metabolites in different treatments, d log2(RsT/CK), e log2(BC@SLRs/CK) and f log2(BC@SLRs/Rs). The metabolites in red box |log2 (FC)|> 0.58 (FC value 1.5 to 2 corresponds to log2(FC) value 0.58 to 1.3 indicates up-regulation of metabolites, similarly for down regulated metabolites); Stars indicate significant differences (ANOVA, * P < 0.05; ** P< 0.01; *** P< 0.001, n = 3)
Log2 (FC) in Fig. 4d–f and Additional file 1: Table S5 shows that the organic acids and phenols in RsT and BC@SLRs increased significantly compared with those in CK. Figure 4d shows seven metabolites (P < 0.001) and nine metabolites log2 (RsT/CK) reached 0.58, indicating that 2, 4 tert-butylphenol, citric acid and palmitic acid were upregulated. These metabolites have been proven to have certain antimicrobial properties, and they are often detected when plants are under disease stress (Ayaz et al. 2008; Suarez-Fernandez et al. 2020; Zhalnina et al. 2018). In Fig. 4e, although log2 (BC@SLRs/CK) has the same change trend as log2 (RsT/CK), the fold changes and significant differences among metabolites were decreased, and only five metabolites of log2 (BC@SLRs/CK) reached 0.58, and citric acid P < 0.001, which indicates the performance of BC@SL in regulating exudates. Meanwhile, Fig. 4f further illustrates that the metabolites were regulated by BC@SL. Phenol and organic acids, such as 2,4 tert-butylphenol, cinnamic acid, and palmitic acid, had negative increments in log2 (BC@SLRs/Rs). Although phenolic or organic acids, such as 2, 4-butyl phenol and palmitic acid, could inhibit the growth of pathogens (Ayaz et al. 2008; Suarez-Fernandez et al. 2020; Zhalnina et al. 2018), these metabolites are also toxic to plants and may induce some fungal diseases (Bao et al. 2022). In addition, cinnamic acid in root exudates has been reported to promote the growth of pathogens at low concentrations (Zhang et al. 2012). The negative effects of 2,4 tert-butylphenol, cinnamic acid, and palmitic acid on plant root microecology are a positive trend, and BC@SL had a positive effect on exudate regulation by inhibiting the secretion of these metabolites.
The total amount of root exudates was measured and expressed as the TOC content (Zhalnina et al. 2018). The addition of Rs reduced the total amount of root exudates, but in BC@SLRs, the situation was alleviated by reducing the stress of pathogens (Additional file 1: Table S6). As shown in Additional file 1: Figure S6b, the percentage of protein content was not high in BC@SLRs, but the variation was more obvious than in CK (2.19% to 5.19%), which may be because Brassica chinensis L. secreted more resistance enzymes to resist the invasion of pathogens. Brassica chinensis L. reduced the carbon source of Rs by decreasing the soluble sugar abundance of RsT, but this trend was attenuated after using BC@SL, where the trend for representative phenolic acids was similar to that of organic acids in Fig. 4a–c. These results prove that BC@SL could modify plant exudate metabolites, and this regulation could improve plant defense systems and thus significantly reduce plant morbidity.
3.5 Root exudate–soil microbial community defense feedback under BC@SL
The root exudate–soil microbial community defense feedback under BC@SL was investigated by constructing a path model on AMOS. Figure 5a shows that the root exudates and microbial community were directly influenced by BC@SL (path coefficient = 0.87 and 0.41), indicating that BC@SL had a greater direct effect on microbial community than root exudates while indirectly influencing root exudates by regulating microbial community (path coefficient = 0.57). Among them, BC@SL directly increased the number of Bacillus. The direct regulatory value of BC@SL on Bacillus reached 0.93. In addition, Rhizoctonia was negatively regulated by BC@SL, with a direct effect value of 0.32 and an indirect value of 0.62 through Bacillus. Thus, BC@SL had a direct regulatory effect on root exudates and microorganisms, and it indirectly facilitated the optimization of defensive feedback between the two.

a Path model outputs: numbers on arrows are the values of standardized path coefficients; Gray and red arrows indicate positive and negative effects, respectively; Microbial communities and exudates are represented by PCoA data; Bacillus and Rhizoctonia are represented by the corresponding genus-level abundance; b-d Variation of metabolism function profiles of 16 s high-throughput sequencing data analyzed by KEGG; b Relative percentages of primary metabolisms
Potential functional features were predicted by KEGG to explore the root exudate-soil microbial community defensive feedback mechanisms. In primary metabolisms, the abundances of biosynthesis, degradation/utilization/assimilation (DUA), reached more than 60%, and 16%, respectively (Fig. 5b). In Fig. 5b, c, biosynthesis and DUA were analyzed in detail for secondary potential metabolisms. After Rs invasion, the abundance of cofactors, prosthetic groups and biosynthesis increased, which can transmit cell signals, regulate metabolism and induce microbial aggregation (Fig. 5c). Cell structural biosynthesis improved in RsT and bulk soil, but nucleoside and nucleotide biosynthesis decreased. These results correspond to the positive performance of Firmicutes, which increased the cell wall synthesis (Figure S5a). In DUA, secondary metabolite degradation, fatty acid and lipid degradation, carboxylate degradation, and aromatic compound degradation had a high proportion in RsT and BC@SLRS (Fig. 5d), corresponding to the decrease in root secretions caused by the death of cabbage root cells after Rs invasion and the secretion of more aromatic organic acids to defend against pathogens (Dang et al. 2023; Zhalnina et al. 2018) (Fig. 4a–c, Additional file 1: Figure S6b). Although the broad gene trends in RsT and BC@SLRs were similar, the distinction, such as the decrease in fatty acid and lipid degradation and C1 compound utilization and assimilation, increased under BC@SL (Fig. 5d). This phenomenon is due to the alleviated death of plant roots and the roots reducing the secretion of organic acids and phenols under BC@SL (Dang et al. 2023; Zhalnina et al. 2018) (Fig. 4d–f). These results reveal that BC@SL and root exudates reshaped soil microbial community by shifting the carbon source available and survival environment to microorganisms in rhizosphere microecology (Liu et al. 2020; Zhalnina et al. 2018).
The interaction between the abundance of root exudates metabolites and the rhizosphere microorganisms was analyzed using spearman correlation (Fig. 6). The relevance of the different metabolites to microorganisms varied, with palmitic acid, stearic acid, and 2,4 tert-butyl-phenol more consistently correlated, and citric acid, while benzoic acid, and phenyl propionic acid were more consistently correlated. These results further indicate the interactions between exudates and microorganisms (Dang et al. 2023). Meanwhile, the results show that Rhizoctonia was positively correlated with palmitic acid and stearic acid but negatively correlated with benzoic acid, 2,4 tert-butylphenol, and citric acid (Fig. 6a). Bacillus was negatively correlated with palmitic acid, stearic acid, 2,4 tert-butylphenol, cyclohexanone, and 2-hydroxyoctanoic and positively correlated with citric acid and benzoic acid (Fig. 6b). Palmitic acid, stearic acid, and benzoic acid are important components of root exudates that regulate soil microbial communities, and pathogen Rs and beneficial bacteria SL-44 are affected.

Bacillus and Rhizoctonia are represented by the corresponding genus-level abundance. Heat map of spearman correction analysis between rhizosphere microorganisms of genus level and root exudates; a bacteria; b fungi
3.6 Microbe response to root exudates
The colony diameter of SL-44 was about d = 1.24 cm on solid medium with BC@SLRs exudates, and that of Rs was d = 3.42 cm on solid medium containing CK exudate (Additional file 1: Figure S7). Figure 7a shows that the count order of SL-44 growing in liquid medium was BC@SLRs > RsT > CK, and the order of Rs cell dry weight in liquid medium was CK > BCSLRs > RsT. The results of liquid culture medium are consistent with those of solid culture medium, thus verifying the differences in the metabolites of CK, RsT, and BC@SLRs exudates and the selectivity of plant root exudates to microorganisms (Liu et al. 2020).

a SL-44 and Rs grown in the medium contain the crude root exudates of CK, RsT and BC@SLRs; b-f SL-44 and Rs grown in the medium contain: b 2,4-ditert-butyl-phenol; c citric acid; d benzoic acid; e palmitic acid; and f stearic acid, respectively. The red upward arrows represent the metabolites promoting SL-44 or Rs growth in pot experiments, the blue downward arrow means the opposite; The red box marks the lower concentration of the measurement system; The different letters in each column display significant differences (LSD-test, P < 0.05)
Based on the experimental results (Figs. 4, 6), five root exudate metabolites were selected as representative metabolites to specifically analyze the effects of root exudates on SL-44 and Rs in Fig. 7. The results showed that the decline trend of SL-44 was greater than that of Rs with the increase in citric acid concentration, but its maximum growth concentration at 0.25 mM was higher than that of Rs at 0.10 mM (Fig. 7b). As shown in Fig. 7c, the quantity of SL-44 showed a trend of increasing and then decreasing with increasing benzoic acid concentration, while Rs was significantly suppressed by benzoic acid. Organic acid affects microorganisms by concentration (Hu et al. 2018). The presence of a suitable concentration (citric acid 0.25 mM and benzoic acid 1.00 mM) was obviously positive to SL-44 but negative to Rs. Numerous antimicrobial metabolites inhibit the growth of beneficial microbes in addition to pathogens (Ayaz et al. 2008; Suarez-Fernandez et al. 2020). However, different microbes have different sensitivity to metabolites (Hu et al. 2018; Liu et al. 2020). Figure 7a–d shows that SL-44 and Rs were more sensitive to citric acid and 2,4 tert-butylphenol than to other metabolites, with lower measurement system concentration of (0–1 mM; others 0–3 mM). In particular, SL-44 and Rs did not vary significantly with the concentration changes in palmitic acid and stearic acid (Fig. 7e–f). Additional file 1: Table S5 shows that palmitic acid and stearic acid were the two most abundant metabolites (3.56–6.96%), whereas citric acid and 2,4 tert-butylphenol were far less abundant (0.71%–2.62%). In addition, SL-44 was more sensitive to 2,4 tert-butylphenol than Rs, and they were inhibited at 0.25 and 0.50 mM, respectively (Fig. 7d). These results reveal that plants reconstruct the structure of soil microbial communities by the differential sensitivity of microorganisms to different exudate metabolites (Hu et al. 2018; Liu et al. 2020). Normally, plants secrete more benzoic acid, 2,4 tert-butylphenol, and other metabolites to resist pathogen invasion (Ayaz et al. 2008; Suarez-Fernandez et al. 2020). However, the concentrations of metabolites, such as 2, 4 tert-butyl phenol, palmitic acid, and stearic acid, could inhibit Rs after the addition of BC@SL (Fig. 4), thus aggregating more microorganisms that are beneficial to the root system, such as SL-44. A notable detail is that concentrations of citric acid and benzoic acid, two metabolites that promote SL-44 growth but inhibit Rs, continued to increase after BC@SL was used.
BC and PGPR regulate the microbial community by altering the soil environment (Wei et al. 2020; Wu et al. 2022). Figure 5a and Fig. 7 demonstrate BC@SL could optimize the composition of plant exudates to make them more suitable for beneficial microorganism. BC@SL altered the soil environment (Additional file 1: Table S2), reshaping the microbial community (Fig. 3) to alleviate the stress of pathogens. In addition, BC@SL optimized the composition of plant root exudates (Fig. 4) to enhance the recruiting of beneficial microorganism (Fig. 6). The aggregated beneficial microorganisms will improve the rhizosphere microecology including root exudates, microbial community (Zhalnina et al. 2018). BC and some exogenous inoculants regulate the root microecology of a wide range of plants (Ren et al. 2016; Sun et al. 2020). For example, BC could improve organic acids in root exudates of maize and increase the abundance of microbes, such as Actinobacteria and Bacteroidetes (Sun et al. 2020). Huang et al. (2017) and Shalev et al. (2022) reported that exogenous inoculants such as Bacillus and Pseudomonas could reshape microbial community of Capsicum annuum and Arabidopsis thaliana, respectively. In addition, Wu et al. (2022) explored the synergistic effect of BC and exogenous inoculants relive the pathogens stress by regulating the soil microbial community of Radix pseudostellariae. Therefore, the regulation of plant root microecology by BC@SL could have broad applicability, not only to Brassica chinensis L.
4 Conclusions
In this study, a novel exogenous soil solid exogenous auxiliary BC@SL based on BC and SL-44 was successfully prepared. The superiority and mechanism of BC@SL in helping plants resist pathogens were investigated. BC@SL alleviated Rs stress through direct inhibition and indirect feedback. Those can be specifically described as (1) BC@SL greatly enhanced the direct inhibition of Rs since BC@SL solved the problem of SL-44 inoculant in soil and synergizes the Rs inhibition by BC and SL-44; (2) BC@SL increased plant resistance enzyme activity, reshaped root microbial community, and regulated root exudate. (3) BC@SL enhanced the positive regulatory effect between root exudates-soil microbial defensive feedback. Overall, as a novel and efficient soil additive, BC@SL could regulate plants and soil microorganisms bidirectionally, change root microecology, promote plant growth, and increase plant resistance.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its Additional files.
References
Adeyem AS, Arturo KA (2016) Interactions between algal extracellular polymeric substances and commercial TiO2 nanoparticles in aqueous media. Environ Sci Technol 50(22):12258–12265. https://doi.org/10.1021/acs.est.6b03684
Ahmed A, He P, He P, Wu Y, He Y, Munir S (2023) Environmental effect of agriculture-related manufactured nano-objects on soil microbial communities. Environ Int. https://doi.org/10.1016/j.envint.2023.107819
An X, Chen Y, Ao M, Jin Y, Zhan L, Yu B, Wu Z, Jiang P (2022a) Sequential photocatalytic degradation of organophosphorus pesticides and recovery of orthophosphate by biochar/α-Fe2O3/MgO composite: a new enhanced strategy for reducing the impacts of organophosphorus from wastewater. Chem Eng J. https://doi.org/10.1016/j.cej.2022.135087
An X, Wang H, Dong C, Jiang P, Wu Z, Yu B (2022b) Core-shell P-laden biochar/ZnO/g-C<sub>3</sub>N<sub>4</sub> composite for enhanced photocatalytic degradation of atrazine and improved P slow-release performance. J Colloid Interface Sci 608:2539–2548. https://doi.org/10.1016/j.jcis.2021.10.166
Andreo-Jimenez B, Schilder MT, Nijhuis EH, Te Beest DE, Bloem J, Visser JHM, van Os G, Brolsma K, de Boer W, Postma J (2021) Chitin- and keratin-rich soil amendments suppress Rhizoctonia solani disease via changes to the soil microbial community. Appl Environ Microbiol. https://doi.org/10.1128/aem.00318-21
Ayaz FA, Hayırlıoglu-Ayaz S, Alpay-Karaoglu S, Grúz J, Valentová K, Ulrichová J, Strnad M (2008) Phenolic acid contents of kale (Brassica oleraceae L. var. acephala DC.) extracts and their antioxidant and antibacterial activities. Food Chem 107(1):19–25. https://doi.org/10.1016/j.foodchem.2007.07.003
Bao L, Liu Y, Ding Y, Shang J, Wei Y, Tan Y, Zi F (2022) interactions between phenolic acids and microorganisms in rhizospheric soil from continuous cropping of panax notoginseng. Front Microbiol. https://doi.org/10.3389/fmicb.2022.791603
Bogoutdinova LR, Lazareva EM, Chaban IA, Kononenko NV, Dilovarova T, Khaliluev MR, Kurenina LV, Gulevich AA, Smirnova EA, Baranova EN (2020) Salt stress-induced structural changes are mitigated in transgenic tomato plants over-expressing superoxide dismutase. Biology. https://doi.org/10.3390/biology9090297
Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1006/abio.1976.9999
Dang K, Hou J, Liu H, Peng J, Sun Y, Li J, Dong Y (2023) Root exudates of Ginger induced by Ralstonia solanacearum infection could inhibit bacterial wilt. J Agric Food Chem 71(4):1957–1969. https://doi.org/10.1021/acs.jafc.2c06708
Dixit R, Agrawal L, Gupta S, Kumar M, Yadav S, Chauhan PS, Nautiyal CS (2016) Southern blight disease of tomato control by 1-aminocyclopropane-1-carboxylate (ACC) deaminase producing Paenibacillus lentimorbus B-30488. Plant Signal Behav. https://doi.org/10.1080/15592324.2015.1113363
Han S, Li X, Luo X, Wen S, Chen W, Huang Q (2018) Nitrite-oxidizing bacteria community composition and diversity are influenced by fertilizer regimes, but are independent of the soil aggregate in acidic subtropical red soil. Front Microbiol. https://doi.org/10.3389/fmicb.2018.00885
Hao J, Chai YN, Lopes LD, Ordóñez RA, Wright EE, Archontoulis S, Schachtman DP (2021) The effects of soil depth on the structure of microbial communities in agricultural soils in Iowa (United States). Appl Environ Microbiol. https://doi.org/10.1128/aem.02673-20
Hu L, Christelle AMR, Selma C, Zhang X, Ye M, Li B, Daniele M, Noemie C, Thomas S, Marcel GAVDH, Klaus S, Matthias E (2018) Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat Commun. https://doi.org/10.1038/s41467-018-05122-7
Huang Y, Wu Z, He Y, Ye B-C, Li C (2017) Rhizospheric Bacillus subtilis exhibits biocontrol effect against Rhizoctonia solani in pepper (Capsicum annuum). Biomed Res Int. https://doi.org/10.1155/2017/9397619
Kadoo MR, Badere RS (2017) Modulation of the activity of chitinases and β-1,3 glucanase in seedlings of cucumber and chilli by the aqueous extract of Cleistanthus collinus. Physiol Mol Plant Pathol 98:46–53. https://doi.org/10.1016/j.pmpp.2017.03.004
Kandoliya UK, Marviya GV, Rathod PJ, Vakharia DN, Golakiya BA (2017) Pathogenesis related hydrolytic enzymes induction in response to PGPR Seed Priming Against Wilt Pathogen (Fusarium oxysporum fsp ciceri) in Chickpea. Ind J Agric Biochem. https://doi.org/10.5958/0974-4479.2017.00030.2
Kolton M, Graber ER, Tsehansky L, Elad Y, Cytryn E (2016) Biochar-stimulated plant performance is strongly linked to microbial diversity and metabolic potential in the rhizosphere. New Phytol 213(3):1393–1404. https://doi.org/10.1111/nph.14253
Kong M, Liang J, White JC, Elmer WH, Wang Y, Xu H, He W, Shen Y, Gao X (2022) Biochar nanoparticle-induced plant immunity and its application with the elicitor methoxyindole in Nicotiana benthamiana. Environmental Science-Nano 9(9):3514–3524. https://doi.org/10.1039/d1en00953b
Kwak M-J, Kong HG, Choi K, Kwon S-K, Song JY, Lee J, Lee PA, Choi SY, Seo M, Lee HJ, Jung EJ, Park H, Roy N, Kim H, Lee MM, Rubin EM, Lee S-W, Kim JF (2018) Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat Biotechnol 36(11):1117–1117. https://doi.org/10.1038/nbt.4232
Li Y, He Y, Wang W, Li X, Xu X, Liu X, Li C, Wu Z (2021) Plant-beneficial functions and interactions of Bacillus subtilis SL-44 and Enterobacter cloacae Rs-2 in co-culture by transcriptomics analysis. Environ Sci Pollut Res 28(40):56333–56344. https://doi.org/10.1007/s11356-021-14578-y
Liu G, Sheng H, Fu Y, Song Y, Redmile-Gordon M, Qiao Y, Gu C, Xiang L, Wang F (2019) Extracellular polymeric substances (EPS) modulate adsorption isotherms between biochar and 2,2 ’,4,4 ’-tetrabromodiphenyl ether. Chemosphere 214:176–183. https://doi.org/10.1016/j.chemosphere.2018.09.081
Liu H, Brettell LE, Qiu Z, Singh BK (2020) Microbiome-mediated stress resistance in plants. Trends Plant Sci 25(8):733–743. https://doi.org/10.1016/j.tplants.2020.03.014
Maddalwar S, Kumar Nayak K, Kumar M, Singh L (2021) Plant microbial fuel cell: opportunities, challenges, and prospects. Biores Technol. https://doi.org/10.1016/j.biortech.2021.125772
Mills DR, Lee JM (1996) A simple, accurate method for determining wet and dry weight concentrations of plant cell suspension cultures using microcentrifuge tubes. Plant Cell Rep 15(8):634–636. https://doi.org/10.1007/bf00232467
Mtibaà R, Olicón-Hernández DR, Pozo C, Nasri M, Mechichi T, González J, Aranda E (2018) Degradation of bisphenol A and acute toxicity reduction by different thermo-tolerant ascomycete strains isolated from arid soils. Ecotoxicol Environ Saf 156:87–96. https://doi.org/10.1016/j.ecoenv.2018.02.077
Pedras MS, Okanga FI (1999) Strategies of cruciferous pathogenic fungi: detoxification of the phytoalexin cyclobrassinin by mimicry. J Agric Food Chem. https://doi.org/10.1021/jf980854r
Poveda J, Martínez-Gómez Á, Fenoll C, Escobar C (2021) The Use of biochar for plant pathogen control. Phytopathology 111:1490–1499. https://doi.org/10.1094/phyto-06-20-0248-rvw
Ren L, Huo H, Zhang F, Hao W, Xiao L, Dong C, Xu G (2016) The components of rice and watermelon root exudates and their effects on pathogenic fungus and watermelon defense. Plant Signal Behav. https://doi.org/10.1080/15592324.2016.1187357
Shalev O, Karasov TL, Lundberg DS, Ashkenazy H, Pramoj Na Ayutthaya P, Weigel D (2022) Commensal Pseudomonas strains facilitate protective response against pathogens in the host plant. Nature Ecol Evolut 6(4):383. https://doi.org/10.1038/s41559-022-01673-7
Sharma M, Saleh D, Charron J-B, Jabaji S (2020) A crosstalk between brachypodium root exudates, organic acids, andbacillus velezensisB26, a growth promoting bacterium. Front Microbiol. https://doi.org/10.3389/fmicb.2020.575578
Suarez-Fernandez M, Marhuenda-Egea FC, Lopez-Moya F, Arnao MB, Cabrera-Escribano F, Nueda MJ, Gunse B, Lopez-Llorca LV (2020) Chitosan Induces plant hormones and defenses in tomato root exudates. Front Plant Sci. https://doi.org/10.3389/fpls.2020.572087
Sun C, Wang D, Shen X, Li C, Liu J, Lan T, Wang W, Xie H, Zhang Y (2020) Effects of biochar, compost and straw input on root exudation of maize (Zea mays L.): From function to morphology. Agric Ecosyst Environ. https://doi.org/10.1016/j.agee.2020.106952
Wang Y, Ma Z, Wang X, Sun Q, Dong H, Wang G, Chen X, Yin C, Han Z, Mao Z (2019) Effects of biochar on the growth of apple seedlings, soil enzyme activities and fungal communities in replant disease soil. Sci Horticult. https://doi.org/10.1016/j.scienta.2019.108641
Wei M, Liu X, He Y, Xu X, Wu Z, Yu K, Zheng X (2020) Biochar inoculated with Pseudomonas putida improves grape (Vitis vinifera L.) fruit quality and alters bacterial diversity. Rhizosphere 16:100261. https://doi.org/10.1016/j.rhisph.2020.100261
Wen T, Ding Z, Thomashow LS, Hale L, Yang S, Xie P, Liu X, Wang H, Shen Q, Yuan J (2023) Deciphering the mechanism of fungal pathogen-induced disease-suppressive soil. New Phytol 238(6):2634–2650. https://doi.org/10.1111/nph.18886
Wu H, Wu H, Jiao Y, Zhang Z, Rensing C, Lin W (2022) The combination of biochar and PGPBs stimulates the differentiation in rhizosphere soil microbiome and metabolites to suppress soil-borne pathogens under consecutive monoculture regimes. Global Change Biol Bioenerg 14(1):84–103. https://doi.org/10.1111/gcbb.12906
Yuan J, Wu Y, Zhao M, Wen T, Huang Q, Shen Q (2018) Effect of phenolic acids from banana root exudates on root colonization and pathogen suppressive properties of Bacillus amyloliquefaciens NJN-6. Biol Control 125:131–137. https://doi.org/10.1016/j.biocontrol.2018.05.016
Zhalnina K, Louie KB, Hao Z, Mansoori N, Rocha UND, Shi S, Cho H, Karaoz U, Loqué D, Bowen BP, Firestone MK, Northen TR, Brodie EL (2018) Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat Microbiol 3(4):470–480. https://doi.org/10.1038/s41564-018-0129-3
Zhang G, Raza W, Wang X, Ran W, Shen Q (2012) Systemic modification of cotton root exudates induced by arbuscular mycorrhizal fungi and Bacillus vallismortis HJ-5 and their effects on Verticillium wilt disease. Appl Soil Ecol 31:85–91. https://doi.org/10.1016/j.apsoil.2012.02.003
Zhang L, Zhou J, George TS, Limpens E, Feng G (2022) Arbuscular mycorrhizal fungi conducting the hyphosphere bacterial orchestra. Trends Plant Sci 27(4):402–411. https://doi.org/10.1016/j.tplants.2021.10.008
Zhao C, Zhang L, Zhang X, Xu Y, Wei Z, Sun B, Liang M, Li H, Hu F, Xu L (2019) Regulation of endogenous phytohormones alters the fluoranthene content in Arabidopsis thaliana. Sci Total Environ 688:935–943. https://doi.org/10.1016/j.scitotenv.2019.06.384
Zhao G, Tian Y, Yu H, Li J, Mao D, Faisal RM, Huang X (2022) Development of solid agents of the diphenyl ether herbicide degrading bacterium Bacillus sp. Za based on a mixed organic fertilizer carrier. Front Microbiol. https://doi.org/10.3389/fmicb.2022.1075930
Zhou X, Zhang J, Rahman MKU, Gao D, Wei Z, Wu F, Dini-Andreote F (2023) Interspecific plant interaction via root exudates structures the disease suppressiveness of rhizosphere microbiomes. Mol Plant 16(5):849–864. https://doi.org/10.1016/j.molp.2023.03.009
Zhou Y, Zhao X, Jiang Y, Ding C, Liu J, Zhu C (2023) Synergistic remediation of lead pollution by biochar combined with phosphate solubilizing bacteria. Sci Total Environ. https://doi.org/10.1016/j.scitotenv.2022.160649
Acknowledgements
Not applicable.
Funding
National Natural Science Foundation of China (22278325, 32060026), Qin Chuangyuan’s Scientists + Engineers Team in Shaanxi Province (2022KXJ-137), Youth Innovation Team of Shaanxi Universities (2022TD071), Xi’an Key Laboratory Performance Assessment Award Subsidy Project (2021JH-201-0004), Agricultural Technology R&D Project of Xi’an Science and Technology Bureau (22NYYF0037), and Shaanxi Provincial Key Research and Development Program (2022NY-053).
Author information
Authors and Affiliations
Contributions
Professor ZS Wu was conceived and supervised the study as corresponding author; XF An revised the manuscript and supervised the study as corresponding author; ZH Deng designed and performed this experiment, and he wrote the manuscript; JW Wang assisted on solving experimental problems in the study; YH He and F Tian revised the important academic content of the article. Z Tu and HJ Li performing this experiment together; F Tian assisted for performing experiments; Professor All authors approved the final article. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Handling editor: Lukáš Trakal
Supplementary Information
Additional file 1: Text S1.
Bacterial strain and culture medium. Text S2. Qualitative and quantitative analysis of Rs inhibition. Text S3. Determination of the growth indicators and disease index of pakchoi. Text S4. Extraction of root exudates with ethyl acetate. Figure S1. A flow chart of the metabolite’s selection criteria used for the study and grown of microbe in the medium contain the acetone. Figure S2. Characterization of biochar and SL-44. Figure S3. Gene copy of SL-44 and Rs used to express their content, determined by AQ-PCR. Figure S4. Plant growth image and enzyme activities. Figure S5. Relative abundance of dominant genus level for Bacteria and Fungi. Figure S6. Principal coordinate analysis of root exudates in different treatments and relative percentages of substances in the root exudates. Figure S7. Growth of SL-44 and Rs on plates treated with root exudates from CK, Rs, BC@SLRs. Table S1. Biochar elemental composition. Table S2. Soil character. Table S3. Plant growth index. Table S4. Alpha diversity index of bacteria and fungi in soil. Table S5. Discriminant metabolites in pakchoi root exudates through GC-MS analysis. Table S6 Composition and content of root exudates.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Deng, Z., Wang, J., He, Y. et al. Biochar-based Bacillus subtilis inoculant for enhancing plants disease prevention: microbiota response and root exudates regulation. Biochar 5, 81 (2023). https://doi.org/10.1007/s42773-023-00284-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42773-023-00284-0