
Abstract
Rice is the primary staple food for more than 50% of the world’s population. Narrowing the gap between the maximum potential food crop yield and actual yield is critical for improving the current yield, resource use efficiency, and global food security. Here, we examined the fertilizer use efficiency (FUE), radiation use efficiency (RUE), and temperature production efficiency (TPE) of four management treatments (super high yield [SH], high yield and high efficiency [HH], farmer level [FP], and basic production level [CK]). SH and HH treatments significantly reduced the yield gap by 22.4 and 9.5%, respectively. The large yield gap between HH and FP was mainly attributed to high specific leaf weight at the jointing stage (7.5–7.7 mg·cm−2), and the high leaf area maintained during tillering–jointing stages (35.4–37.6 m2·m−2·per day). Compared with FP, HH increased the specific leaf weight in the heading stage (8.2–8.4 mg·cm−2), relative crop growth rate, net assimilation rate (NAR), and mean leaf area index (> 2.6). Moreover, compared with FP, HH significantly increased partial factor productivity (PFP) of nitrogen, FUE, TPE, and RUE owing to greater yield and NAR after the full heading stage. Although the HH yield was 93.32% that of SH, HH increased PFP of fertilizer (12.5%), fertilizer nitrogen (9.07%), and fertilizer K2O (36.34%), and required 26% less fertilizer than SH. The findings of this study could facilitate high-efficiency rice production and bridging of yield gaps.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rice (Oryza sativa L.) is a staple food for more than 50% of the world’s population (Iizumi et al., 2017; Zhang et al., 2016) and is the dominant cereal crop in China (Khush, 2013). Over the past 20 years, the annual increase in rice yields has dropped below 1% worldwide; however, rice yields must continue to increase by 1.2–1.5% per year to meet the expected demand of the increasing population and economic development in the next decade (Alam et al., 2013). Narrowing the gap between the maximum potential food crop yield and actual yield is critical (Yuan et al., 2021), especially as crop yields must continue to increase at an annual rate of 2% to ensure national food security in China (Fan et al., 2012).
China is the source of 28% of the global rice supply (Deng et al., 2019) and possesses 36% of the global area under rice cultivation (Wang et al., 2020). As the largest production base for japonica rice, Northeast China contributes 53% of the national japonica rice supply, with Heilongjiang Province being the largest producer in China (National Bureau of Statistics of China, 2016). Therefore, bridging the gap in japonica rice yield in Northeast China could enhance food security in China as well as globally (Wang et al., 2018).
Extreme temperatures negatively influence rice growth, and in turn, rice yield in China (Jia et al., 2019, 2022; Wang et al., 2019). This is especially true in Northeast China, which has experienced frequent cold waves in the past (Jia et al., 2019; Shimono et al., 2007; Sun et al., 2017; Zhang et al., 2017). Numerous studies have demonstrated that Northeast China has a great potential to increase its rice yield (Chen et al., 2017; Wang et al., 2020; Zhang et al., 2014a, 2014b). Therefore, to enhance average farm yields and reduce the yield gap (Peng et al., 2008; Zhang et al., 2009), it is necessary to develop countrywide adaptation strategies based on spatial rice production patterns (Zhang et al., 2014a, 2014b, 2017).
Investigation of cold-region japonica rice production in Heilongjiang province has revealed large yield gaps and resource use efficiency. The average yield gap is used to estimate the gaps between current farm yields and the potential yield (Van Ittersumetal., 2013). To improve crop yields, the following aspects are critical: understanding the yield potentials and yield gaps of major cereal crops, reducing yield and resource use efficiency gaps, and improving average farm yields (Liu et al., 2016). Yield gaps are caused by limiting factors that can be roughly categorized into climatic, soil, and social factors, varieties and cultivation measures, and other aspects (Deng et al., 2019). Among these, light and temperature affect biomass production by modulating the environment for crop growth (Ramankutty et al., 2002). Temperature fluctuations, increases in temperature (≥ 10 °C), and solar radiations are key factors that limit potential yield (Wang et al., 2020); the contribution of meteorological factors to the yield gap is approximately 8% (Liu et al., 2016). Currently, the actual crop radiation use efficiency (RUE) is only 1‒2% worldwide, which is far lower than the maximum theoretical crop RUE of 5‒6% (Loomis & Williams, 1963).
Increasing biomass and improving crop RUE are potential key strategies of increasing crop yield (Zhang et al., 2009). Furthermore, to reduce gaps in both yield and efficiency, high-efficiency populations can be grown by breeding hardy varieties, determining reasonable densities, optimizing fertilization (Foley et al., 2011), and eliminating nutrient overuse (Mueller et al., 2013). There are considerable opportunities for reducing the environmental impact of agriculture by eliminating nutrient overuse while still allowing an approximate 30% increase in the production of major cereal crops such as rice. For instance, breeding new rice varieties can lead to a 12% increase in yields (Zhang et al., 2009). However, in practice, 45‒70% of the potential for increased production remains untapped (Mueller et al., 2013).
Few studies have identified the yield gaps and efficiency between different yield levels in Northeast China, and to date, systematic analysis has not been employed to estimate the rice yield and efficiency gaps across this region. Bridging of these gaps is necessary to improve yield and use efficiencies that are based on resources. It is difficult to draw concrete conclusions based on the quantification of yield gaps and efficiency between different yield levels of japonica rice in the cold regions of Heilongjiang Province; therefore, elucidating the relationship would facilitate the development of strategies to address the gaps.
To highlight the relationship between yield gap and resource use efficiency, we examined the resource use efficiency and production efficiency of four management treatments for 2 years. Using an integrative analysis of radiation-temperature production potential and crop yield performance, we explored factors affecting yield and efficiency. The aims of this study were to: (1) identify the key factors contributing to the high grain yield and yield gaps, (2) discover the mechanism underlying high-efficiency rice production, and (3) explore the potential options for bridging the yield gaps and increasing efficiency. The results of the present study could facilitate high-efficiency rice production and help bridge yield gaps.
Materials and Methods
Plant Material and Growth Conditions
The study was conducted in the Acheng District in Harbin, Heilongjiang Province, China (longitude, 127°04′ E; latitude, 45°52′ N). The soils in the paddy fields were classified as Black soil. Basic soil fertility (0–20 cm) was analyzed before starting the experiment in 2017 and 2018 (Table1). The Oryza sativa L. subsp. japonica variety DN427 was used.
Experimental Design
Field experiments were carried out in a randomized complete block design in triplicates of four treatments, namely: (1) basic production management (control, CK), without NPK application; (2) farmer management (following the local farmers’ fertilizer practice, FP), with total N, P2O5, and K2O application rates of 150, 60, and 45 kg·ha−1, respectively; (3) high yield and high efficiency (HH), with total N, P2O5, and K2O application rates of 150, 60, and 45 kg·ha−1, respectively; and (4) super high yield (SH), with total N, P2O5, and K2O application rates of 200, 60, and 45, respectively. The HH treatment employed alternate irrigation, whereas all other treatments used submerged irrigation, which involves a continuous water layer of 1–3 cm from transplanting until approximately 10 days before rice harvest. Alternation irrigation consists of the following: maintaining a 2 cm water layer for 7–9 d after transplanting, with a soil water potential of − 10 kPa (measured by the WET-2Soil Moisture Meter; Cambridge, UK) before the booting stage; re-watering at the booting stage with a water layer of 1–3 cm; irrigating to achieve a saturated state; and drying naturally until a soil water potential of − 20 kPa from the heading stage to maturity stage was achieved (Sun et al., 2012).Approximately 35-day-old seedlings were transplanted at three seedlings per hill with a spacing of 13.3 cm (hill space) by 30 cm (row space)for each yield level. Each plot area measured 300 m2, with three replicates. Urea (containing 46% N) was used as nitrogen fertilizer, diammonium phosphate (containing 18% N and 46% P2O5) was used as phosphate fertilizer, and potassium (containing 50% K2O) sulfate was used as potassium fertilizer (Table 2).
Field Sampling and Lab Analyses
A field climate meter (RR-9100; Hangzhou, Zhejiang, China) was used to measure the field temperature and light radiation. The total radiation from sunshine hours was calculated using the Ǻngström-Prescott method (Angstrom, 1924).
For each replication, ten hills with the same crop growth were selected at tillering, jointing, full heading, and maturity stages, and three replications were performed. Then, the samples were divided into roots, stems, leaves, and panicles. After the samples had been fixed at 105 °C for 30 min, they were oven dried at 80 °C to a constant weight before the calculation of the accumulated biomass.
The leaf area was monitored from the tillering to the maturing stage. The standing leaf area of the stand was calculated using every leaf of selected plants. The specific leaf weight (Saengwilai et al., 2020), leaf area duration, crop growth rate (Bowsher et al., 2016), relative crop growth rate (Li et al., 2016), and net assimilation rate (NAR; Hirose, 1984) were used to evaluate the production capacity of photosynthetic products. They were calculated as follows:
where, PLA1 and PLA2 are the leaf areas at times t1 and t2, respectively, Wt1 and Wt2, represent the total plant dry matter accumulation at t1 and t2, respectively, and it is the number of days between two measurements.
For each yield level, three replicates in a 5 m2 area were randomly selected at the mature stage to determine the actual yield. Representative plants from 10 hills were selected in triplicate for each yield level to measure effective panicles (EP), grains per panicle, 1000-grain weight (GW), and seed setting rate. The yield gaps (GY) between CK and FP, FP and HH, and HH and SH were defined as GY1, GY2, and GY3, respectively. Light temperature yield potential productivity (YPT) was calculated using a method described by Lai et al. (2014), using the equation YPT = YP × f (T); where, YP is the photosynthetic production potential and f (T) is the temperature correction factor. The f (T) of thermophilic crops was calculated as follows:
where T is the daily average temperature. YP was calculated as follows:
where YP (kg·hm−2) is the photosynthetic potential productivity per unit area, C is the conversion factor with a value of 666.7, and ΣQi (MJ·m−2) is the total radiation in each month of the growing season. The other parameters are described in Table 3.
The partial factor productivity of fertilizer (PFP) was calculated as follows: PFP (kg·kg−1) = yield (kg·ha−1)/fertilization amount (kg·ha−1). Fertilizer use efficiency (FUE) was calculated as follows: FUE (kg·kg−1) = [yield in the fertilized area (kg·ha−1)-yield in non-fertilized area (kg·ha−1)]/fertilization amount (kg·ha−1). The PFP gaps between FP and HH and between HH and SH were defined as GPFP1 and GPFP2, respectively. The FUE gaps between FP and HH, and HH and SH were defined as GFUE1 and GFUE2, respectively.
RUE was calculated as follows: RUE (g·MJ−1) = aboveground biomass during the growth period (g·m−2)/total intercepted solar radiation per unit area (MJ·m−2). Temperature production efficiency (TPE) was calculated as follows: TPE (g·m−2·°C−1·d−1) = aboveground biomass during the growth period (g·m−2)/effective accumulated temperature during the growth period (°C). Gaps in RUE between CK and FP, FP and HH, and HH and SH were defined as GRUE1, GRUE2, and GRUE3, respectively; gaps in TPE between CK and FP, FP and HH, and HH and SH were defined as GTPE1, GTPE2, and GTPE3, respectively.
Three representative samples were obtained for each growth stage: tillering, jointing, full heading, and maturity. The yield performance parameters were calculated as previously described by Zhang et al. (2007):
where MLAI is the mean leaf area index, D is the total days in the growth period, MNAR is the mean NAR, HI is the harvest index, EP is the effective panicles, GN is the filled grain number per panicle, and GW is the 1000-grain weight.
Statistical Analysis
Data analyses were performed using SPSS 21.0 software (IBM, Armonk, NY, USA). Analysis of variance (ANOVA) was used to evaluate the effects of the different yield levels on yield, yield components, yield performance parameters, RUE, and TPE. The statistical model included variation due to year, treatment, and year × treatment interactions. The results are reported as the mean values of triplicates. Standard deviation (SD) was calculated directly from the crude data from at least three different replicates in each experiment. Microsoft Excel 2016 was used for mapping and calculating the standard errors of the mean, which are shown in the graphs as error bars. Multiple stepwise regression was used to analyze the relationship between FUE and yield gap.
Results
Yield Gap and Yield Components of Cold-Region Japonica Rice Under Different Management Treatments
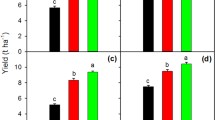
The YPT of rice in Heilongjiang Province was 16, 163.74 kg·ha−1 (16, 372.04 kg·ha−1 in 2017 and 15, 955.43 kg·ha−1in 2018). The SH, HH, FP, and CK management treatments achieved 65.7, 52.8, 43.3, and 28.1% of the YPT, respectively. The yield gap between CK and YPT was 11, 625.7 kg·ha−1 (71.9% of the YPT). Compared with CK treatment, HH and SH displayed an increase in yield of 15.7 and 43.9%, respectively. The GY1, GY2, and GY3 of cold-region japonica rice were 2462.0, 1534.0, and 2074.7 kg·ha−1, respectively (Fig. 1).

Grain yield under different yield levels of cold-region japonica rice. Error bars indicate the mean ± SD (n = 3); different lowercase letters indicate significant difference (P < 0.05). SH super high yield, HH high yield and high efficiency, FP farmer level, CK basic production level
Significant differences were observed in the number of EP under different management treatments, but there were no differences in the seed setting rate and GW. The values of the number of EP, grain number per panicle, and yield under the CK treatment were lower than those under the FP treatment. Compared with those under FP, the number of EP significantly increased under HH, and the number of EP and grain number per panicle significantly increased under SH (Table 4).
Yield Performance Parameters of Cold-Region Japonica Rice Under Different Management Treatments
As the yield level increased, the MLAI and EP significantly increased, the MNAR gradually increased, and the 1000-GW decreased or remained unchanged. MLAI, MNAR, and EP reached their maximum values under SH and were significantly higher than those under the other management treatments. The HI was significantly higher under HH than under FP and SH. The extent of changes in MLAI and GW reflected the inconsistencies between groups and individuals, and changes in HI and MNAR reflected source-sink coordination differences (Table 5). Compared with those under FP, the MLAI (13.9%, on average), HI (3.9%, on average), and EP (22.5%, on average) under HH were significantly higher, indicating that the population size of cold-region japonica rice is more manageable and that the source-sink coordination is better in the case of HH. Under SH, the MLAI (27.3%, on average), MNAR (12.9%, on average), and EP (38.3%, on average) were significantly higher (compared with FP), whereas HI (2.9%, on average) was significantly lower (P < 0.05), indicating that the source-sink relationship is not coordinated, and that the translocation efficiency of dry matter is reduced.
Fertilizer PFP and FUE of Cold-Region Japonica Rice Under Different Management Treatments
The PFPn and PFP of cold-region japonica rice were significantly different among different yield levels. Compared with FP, HH displayed a significant increase (8.13%, on average) in PFP, whereas SH displayed a significant decrease (3.61%, on average), but the change was not significant. Values for GPFP2 and GPFP3 were 2.6 and 3.8 kg·kg−1, respectively. FUE and PFPn were improved significantly under the HH treatment (53.13% higher than that under FP, on average) and SH treatment (18.09% higher than that under FP, on average). The FUE gaps between FP and HH and between HH and SH were defined as GFUE1 and GFUE2, respectively. GFUE1 and GFUE2 were 3.9 and 1.7 kg·kg−1, respectively, and GPFPn1 and GPFPn2 were 9.1 and 4.9 kg·kg−1 N, respectively (Table 6).
According to multiple stepwise regression analysis results, PFPn was the main factor influencing the production of GY2 and GY3 (Table 7). The amount of nitrogen fertilizer used was the same in both HH and FP treatments. However, the irrigation methods and N application strategies, which are conducive to the accumulation and transportation of photosynthetic compounds, differed and represented the key factors influencing the GY2 value. In addition, the incremental application of N fertilizer was responsible for the GY3 value (Table 7).
Population Quality of Cold-Region Japonica Rice Under Different Management Treatments
Growth Period
Compared with those under FP, the tillering stage of cold-region japonica rice occurred two days later, and the jointing stage to maturity stage occurred 2–3 days earlier under CK, whereas the booting stage to maturity stage occurred 2–4 days later under HH and SH (Table 8).
Biomass Accumulation and Transport
The biomass of stems and leaves at the full heading stage and the biomass of panicles were significantly higher under HH and SH than that under FP or CK. The translocation rate of the vegetative dry matter of cold-region japonica rice was significantly lower under CK than under other treatments, and was also significantly lower under HH and SH than under FP. Additionally, the translocation contribution rate was significantly higher under CK than under other management treatments and significantly lower under HH and SH than under FP management. The above results indicate that dry matter transport is correlated with yield level; the higher the yield level, the greater the mass that is assimilated in the vegetative organs and the higher the contribution rate of photosynthetic contraction to spike development after the full heading stage (Table 9).
Crop Growth Rate and Relative Crop Growth Rate
The crop growth rate of cold-region japonica rice at each growth stage was significantly lower under FP than under other management treatments. Compared with FP, HH was not associated with significant difference in growth rate from full heading stage to maturity stage, but this growth rate significantly increased under other management treatments. The relative growth rate (RGR) after the full heading stage was significantly higher under SH than that under the other management treatments. Compared with that under FP, the RGR of cold-region japonica rice increased by 16.9% under SH. The above results indicate that increasing the growth rate and RGR after the full heading stage is conducive to yield improvement (Fig. 2).

Crop growth rate and relative crop growth rate of cold-region japonica rice under the four different yield levels. Error bars indicate the mean ± SD (n = 3); different lowercase letters indicate significant difference (P < 0.05). CK basic production level, FP farmer level, HH high yield and high efficiency, SH super high yield, TS tillering stage, JS jointing stage, BS booting stage, HS heading stage, FH full heading stage, MS maturity stage
Leaf Area Duration and NAR
The leaf area duration under SH treatment was the highest, and the leaf area duration gap between different management treatments increased gradually as growth progressed. With a progression in growth, the NAR of cold-region japonica rice showed a downward trend. During the tillering stage–jointing stage and full heading stage–maturity stage, the NAR of japonica rice was the highest under SH (Fig. 3). Furthermore, the specific leaf weight under CK showed an upward trend with the growth period but was significantly lower than under other management treatments. The specific leaf weight in both tillering and heading stages under HH and TS–HS under SH significantly increased compared with those under HH (Fig. 3).

Leaf area duration, net assimilation rate, and specific leaf weight of cold-region japonica rice under four different yield levels. Error bars indicate the mean ± SD (n = 3); different lowercase letters indicate significant difference (P < 0.05). CK basic production level, FP farmer level, HH high yield and high efficiency, SH super high yield, TS tillering stage, JS jointing stage, BS booting stage, HS, heading stage, FH full heading stage, MS maturity stage
Resource use Efficiency and the Efficiency Gap of Cold-Region Japonica Rice Under Different Management Treatments
The average potential RUE of cold-region japonica rice was 1.62 g·MJ−1. RUE significantly differed between treatments (SH > HH > FP > CK), with the SH, HH, FP, and CK treatments achieving 80.9, 68.8, 61.1, and 38.3% of the potential RUE, respectively. GRUE1 of tillering stage–jointing stage, jointing stage–full heading stage, full heading stage–maturity stage, and the full growth period of cold-region japonica rice were 0.62, 0.36, 0.29, and 0.37 g·MJ−1, respectively; in the cases of GRUE2 and GRUE3, the values were 0.22, 0.13, 0.12, and 0.13 g·MJ−1, respectively, and 0.24, 0.24, 0.37, and 0.20 g·MJ−1, respectively. The maximum RUE gaps for the tillering stage–jointing stage and jointing stage–full heading stage occurred in GRUE1, with values of 0.62 g·MJ−1 and 0.36 g·MJ−1, respectively. The full heading stage-maturity stage had the highest GRUE3 values (0.37 gMJ−1). The findings show that by using the HH treatment, RUE may be significantly enhanced from full heading stage to maturity stage (Table 10).
The average potential TPE of cold-region japonica rice was 2.61 g·m−2·°C−1·d−1, with the SH, HH, FP, and CK treatments achieving 77.6, 64.4, 56.1, and 36.8% of the potential TPE, respectively. The TPE also differed significantly between yield levels (SH > HH > FP > CK) and increased under HH (14.8%) and SH (38.4%) than under FP. The GTPE1, GTPE2, and GTPE3 values were 0.50, 0.22, and 0.35 g·m−2·°C−1·d−1, respectively (Fig. 4).

Production efficiency of temperature of cold-region japonica rice under four different yield levels. Error bars indicate the mean ± SD (n = 3); different lowercase letters indicate significant difference (P < 0.05). SH super high yield, HH high yield and high efficiency, FP farmer level, CK basic production level
Relationship Between RUE, TPE, and Population Quality of Cold-Region Japonica Rice Under Different Management Treatments
RUE of the whole growth period showed a significant positive correlation with the leaf area duration from the tillering stage–jointing stage (R2 = 0.96), while the leaf area duration of TS–JS showed a significant positive correlation with the specific leaf weight of the heading stage (R2 = 0.97) and the highest correlation coefficient (Fig. 5a). With an increase in leaf area duration in the tillering stage–jointing stage, the specific leaf weight of the heading stage under each yield treatment increased, and the RUE of the whole growth period increased (Fig. 5c). When the leaf area duration of the tillering stage–jointing stage was 35.4–37.6 m2·m−2·per day and the specific leaf weight under HS was in the range of 8.2–8.4 mg·cm−2, the RUE of the whole growth period (1.44–1.24 g·MJ−1) was higher compared to that observed under CK and FP (Fig. 5c). Specific leaf weight at the jointing stage showed a significant positive correlation with the leaf area duration of the tillering stage–jointing stage (R2 = 0.96; Fig. 5a) and was in the range of 7.5–7.7 mg·cm−2, while the leaf area duration of tillering stage–jointing stage was higher than observed under CK and FP (Fig. 5b). TPE and MLAI, as well as the MLAI and growth rate of the full heading stage, were significantly positively correlated and showed the highest correlation coefficient (Fig. 5a). When the MLAI value exceeded 2.6, TPE under HH and SH was higher than under the other yield treatments (Fig. 5d).

The relationship between RUE, TPE, and population quality.X1, X2, and X3,leaf area duration of tillering stage (TS)–jointing stage (JS), JS–full heading stage (FH), and FH–maturity stage (MS), respectively; X4, X5, X6, X7, and X8,specific leaf weight of TS,JS,heading stage (HS), FH, and MS, respectively; X9, X10, and X11, crop growth rate of TS–JS,JS–FH, and FH–MS, respectively; X12: RUE of full growth period;X13: TPE. CK basic production level, FP farmer level, HH high yield and high efficiency, SH super high yield, MLAI mean leaf area index.*P < 0.01
Relationship Between the Yield and Efficiency Gaps of Cold-Region japonica Rice
The relationships between the yield, TPE, and RUE gaps of cold-region japonica rice at different yield levels were significantly positive (R2 = 0.9297 or 0.9273). As the yield gap shrank, the TPE and RUE gaps were also reduced (Fig. 6).

Relationship between the yield and efficiency gaps of cold-region japonica rice in the first accumulated temperature zone of Heilongjiang. **P < 0.01
Coupling Effect of Irrigation, N and K Application on Rice Grain Yield
Irrigation, topdressing N and K application were independent variables. Grain yield was response variables. Based on the least square method, and binary quadratic regression equations were established to calculate the amounts of irrigation, topdressing N and K needed to maximize the above parameters (Table 11). The results showed that the influence of irrigation, N and K input on the dependent variables reached significant level (P < 0.01). The amounts of irrigation and topdressing N and K fertilizer corresponding to the maximum value were shown in Table 11. The maximum grain yield of was obtained when 10,187.7–10,562.9 m3 ha−1 of irrigation water, 76.047 kg ha−1 of topdressing N fertilizer and 26.733 kg ha−1 of topdressing K fertilizer were applied (Fig. 7).

Relationships among irrigation, N and K application and grain yield of japonica rice in the first accumulated temperature zone of Heilongjiang
Therefore, it is necessary to further analyze the water and topdressing fertilizer input to determine the best coupling treatments. The coupling effects of irrigation and topdressing of N and K application on the rice grain yield in the two-year experiment exhibited a downward convex shape. The 95, 90, 85, and 80% grain yields were analyzed. The grain yield in the 90% acceptable regions could be achieved at the acceptable range of 90% simultaneously, and the ranges of the three indicators were similar (Fig. 7). Considering all factors of the two years comprehensively, the grain yield could achieve the optimal value at the irrigation water of 10,187.7–10,562.9 m3 ha−1, and topdressing fertilizer of N and K of 50–80 kg ha−1 and 20–40 kg ha−1, respectively.
Discussion
Achieving High Yields of Cold-Region Japonica Rice in the First Accumulated Temperature Zone of Heilongjiang
Studies have found that a reasonable population structure is the key to ensuring yield, whereas simultaneously ensuring a balance in source-sink-translocation and efficient light use by the population is the key to increasing yield (Cao and Moss, 1989; Cook & Evans, 1983; Zelitch, 1982). Nevertheless, few studies have focused on group quality to identify the gaps in the yield between different yield treatments in Northeast China. According to the results of the present study, two factors could explain the yield gap between FP and HH.
First, alternating irrigation and an appropriate basal ratio to tillering and panicle fertilizer can achieve high yields (Sun et al., 2012). Although HH and FP utilized comparable amounts of N fertilizer, the irrigation methods and N fertilizer management differed. Under HH, alternating irrigation and a basal to tillering to panicle fertilizer ratio of 6:3:1 enhanced the accumulation and transportation of photosynthetic compounds, supporting HH as the best management treatment. The large GY2 yield gap was mainly attributed to the high specific leaf weight in the jointing stage, maintaining the high leaf area duration of the tillering stage–jointing stage under HH, resulting in increased specific leaf weight in the heading stage that promoted greater MLAI, RGR, and NAR values when compared with those under FP. Secondly, compared with FP, HH significantly increased partial factor productivity (PFP) of nitrogen, FUE, TPE, and RUE owing to greater yield and NAR after the full heading stage. This indicates that under HH, the population size of cold-region japonica rice is more reasonable, and the source-sink coordination is better than that under FP (Hirotsu et al., 2005).
RGR is a key variable in influential treatments of plant ecology, while the NAR is largely independent of RGR (Shipley, 2006). The grain yield depends mainly on photosynthates produced after full heading, which depends on NAR (Li et al., 2008). In the present study, the GY3 of cold-region japonica rice was 2074.7 kg·ha−1. The NAR during tillering stage–jointing stage and full heading stage–maturity stage growth and the RGR after full heading stage growth of cold-region japonica rice were significantly higher under SH than under other treatments. This indicates that a reasonable tillering population structure ensured that the population maintained a high photosynthetic potential and NAR after full heading stage as well as a relatively high population growth rate; this is the key explanation for the GY3 value. However, compared with those under FP, the MLAI, MNAR, and EP values increased significantly, and HI decreased significantly under SH. This indicates that the source-sink relationship is uncoordinated, and that the biological yield conversion efficiency decreased under SH.
Achieving High Efficiency of Cold-Region Japonica Rice in the First Accumulated Temperature Zone of Heilongjiang
The key strategies for increasing crop yield – when increasing the crop leaf area and harvest indices is not possible – include increasing the biomass and improving the crop RUE (Zhang et al., 2009). Compared to FP, HH was found to be effective at simultaneously increasing the yield and efficiency, as it reduced the yield gap by 9.5% and increased PFPn, FUE, PFP, TPE, and RUE.
Leaf photosynthetic rate also affects RUE (Sinclair & Horie, 1989), and a high RUE may increase the grain yield of rice (Zhang et al., 2009). The large RUE gap between HH and FP was mainly attributed to the high specific leaf weight of the jointing stage and leaf area duration of tillering stage–jointing stage under HH that increased the specific leaf weight of the heading stage, promoting greater biomass accumulation than that observed under FP. The increased specific leaf weight of the jointing stage and leaf area duration of tillering stage–jointing stage due to increased MLAI were also major contributors to the improved TPE under HH. These are the key factors involved in the bridging of the gaps in RUE and TPE by HH. Although the HH yield was 93.32% of the SH yield, HH increased PFP and PFPn and required less fertilizer application compared to SH. Less fertilizer application may reduce the pressure on the environment and decrease production costs. Thus, HH is an effective production method where the yield and fertilizer use efficiency are improved synergistically (Sun et al., 2018).
The present study also demonstrated that the SH yield only reached 65.7% of the YPT, much lower than the reported threshold value of 80% (Lobell et al., 2009). Moreover, the attainable yield was found accounted 80.9%‒94.8% of the yield potential in Northeast China (Wang et al., 2018), which is 15.7%‒29.1% higher than that reported in our study. There are two possible explanations for this discrepancy; first, the low production efficiency in utilizing radiation and temperature by crops has become an important limiting factor in reducing the yield gap (Chen et al., 2017; Deng et al., 2019; Liu et al., 2016; Zhang et al., 2016). Indeed, the relationships between yield gaps and RUE and TPE gaps of cold-region japonica rice with different yield levels were significantly positive, consistent with the findings of previous studies (Peng et al., 2004; Strock et al., 2018). The RUE and TPE of SH achieved 80.9% and 77.6% of the potential RUE and TPE, respectively, indicating that it is difficult to eliminate the yield gaps between the current yields reported by farmers and the simulated maximum yield in the short term. However, it is feasible to eliminate the yield gaps between reported yields and regional trials (Deng et al., 2019). Second, FUE requires improvement; although high-level fertilizer input increases yield production, it reduces resource use efficiency and increases environmental pressure (Sun et al., 2018). Compared with those under HH, the PFP and PFPn values did not significantly increase with the increased amount of fertilization used under SH, which also had an extremely low FUE. Moreover, PFP was higher under FP than under SH. Although the “one-time fertilization” method can effectively reduce labor costs in the case of FP, it increases the loss of fertilizer resources and lowers the yield and FUE. Overall, achieving high yields at low costs should be recognized as advantageous in rice production (Trachsel et al., 2013).
Challenges and Prospects for Bridging the Rice Yield Gap in Heilongjiang Province
Small holdings are the main agricultural treatments in China (Zhang et al., 2016), and the current agronomic yield levels of farmers is affected by culture and income level (Shen et al., 2013). In the present study, the FP yield was 7000.0 kg·ha−1 on average in Heilongjiang, accounting for 43.3% of the YPT, which is far lower than the threshold value of 80% (Lobell et al., 2009). These results indicate a great opportunity for yield improvement in Heilongjiang Province in the future (Wang et al., 2020), as providing farmers with information on proper crop management strategies and breeding high-yield crop varieties may significantly improve the current yields (Stuart et al., 2016). Implementing nationwide macro-control and promoting agricultural technologies to improve the current crop management strategies could reduce GRUE2 (0.13 g·MJ−1), GTPE2 (0.22 m−2·°C−1·d−1), and GY2 (1534.00 kg·ha−1). Furthermore, other aspects of rice production under HH, such as its economic benefits and environmental sustainability, should be examined.
Conclusion
To bridge yield gaps and boost efficiency in japonica rice production in Northeast China, the current study set out to discover the principles underpinning high-efficiency rice production and investigate viable solutions. The yield disparity was dramatically narrowed by SH and HH treatments by 22.4 and 9.5%, respectively. HH was an effective method to simultaneously increase yield, PFPn, TPE, and RUE. The large yield gap between HH and FP was mainly attributed to high specific leaf weight at the jointing stage (7.5–7.7 mg·cm−2), and high leaf area duration maintained during the tillering–jointing stages (35.4–37.6 m2·d·m−2). Compared with FP, HH increased the specific leaf weight of the heading stage (8.2–8.4 mg·cm−2), promoting a high mean leaf area index (> 2.6), relative crop growth rate, and NAR. Moreover, owing to the greater yield and NAR after the full heading stage, HH significantly increased PFPn, FUE, TPE, and RUE (compared with FP). Although HH yield was 93.32% of SH, HH increased the PFP of fertilizer (12.5%), fertilizer nitrogen (9.07%), and required less fertilizer application than SH. Reliance on imports for even a small fraction of China's rice supply is cause for concern, especially when Heilongjiang’s cold stress causes a shortage of the grain. Compared to the FP management treatment, the HH management treatment is highly flexible and might make it easier to maintain yields and resource use efficiency. Indeed, it may be possible to achieve and maintain rice self-sufficiency in China, especially if the rate of increase in yield can be accelerated in Heilongjiang Province.
Data Availability
The data shown in this article are public, but data files cannot be uploaded. The research data are confidential.
References
Alam, M. M., Karim, M. R., & Ladha, J. K. (2013). Integrating best management practices for rice with farmers’ crop management techniques: A potential option for minimizing rice yield gap. Field Crops Research, 144, 62–68.
Angstrom, A. (1924). Solar and terrestrial radiation. Report to the international commission for solar research on actinometric investigations of solar and atmospheric radiation. Journal of the Royal Meteorological Society, 50, 121–126.
Bowsher, A., Mason, C., Goolsby, E., & Donovan, L. (2016). Fine root tradeoffs between nitrogen concentration and xylem vessel traits preclude unified whole-plant resource strategies in Helianthus. Ecology and Evolution, 6, 1016–1031.
Cao, W., & Moss, & D.N. (1989). Temperature effect on leaf emergence and phyllochron in wheat and barley. Crop Science, 29, 1018–1021.
Chen, Y., Wang, P., Zhang, Z., Tao, F., & Wei, X. (2017). Rice yield development and the shrinking yield gaps in China, 1981–2008. Regional Environmental Change, 17, 2397–2408.
Cook, M., & Evans, L. (1983). The roles of sink size and location in the partitioning of assimilates in wheat ears. Functional Plant Biology, 10, 313–327.
Deng, N., Grassini, P., Yang, H., Huang, J., Cassman, K. G., & Peng, S. (2019). Closing yield gaps for rice self-sufficiency in China. Nature Communications, 10, 1725.
Fan, M., Shen, J., Yuan, L., Jiang, R., Chen, X., Davies, W. J., & Zhang, F. (2012). Improving crop productivity and resource use efficiency to ensure food security and environmental quality in China. Journal of Experimental Botany, 63, 13–24.
Foley, J. A., Ramankutty, N., Brauman, K. A., Cassidy, E. S., Gerber, J. S., Johnston, M., Mueller, N. D., O’Connell, C., Ray, D. K., West, P. C. J. N., Balzer, C., Bennett, E. M., Carpenter, S. R., Hill, J., Monfreda, C., Polasky, S., Rockström, J., Sheehan, J., Siebert, S., … Zaks, D. P. (2011). Solutions for a cultivated planet. Nature, 478, 337–342.
Hirose, T. (1984). Nitrogen use efficiency in growth of Polygonum cuspidatum Sieb. et Zucc. Annals of Botany, 54, 695–704.
Hirotsu, N., Makino, A., Yokota, S., & Mae, T. (2005). The photosynthetic properties of rice leaves treated with low temperature and high irradiance. Plant and Cell Physiology, 46, 1377–1383.
Iizumi, T., Furuya, J., Shen, Z., Kim, W., Okada, M., Fujimori, S., Hasegawa, T., & Nishimori, M. (2017). Responses of crop yield growth to global temperature and socioeconomic changes. Scientific Reports, 7, 7800.
Jia, Y., Liu, H., Wang, H., Zou, D., Qu, Z., Wang, J., Zheng, H., Wang, J., Yang, L., & Mei, Y. (2022). Effects of root characteristics on panicle formation in japonica rice under low temperature water stress at the reproductive stage. Field Crops Research, 277, 108395.
Jia, Y., Wang, J., Qu, Z., Zou, D., Sha, H., Liu, H., Sun, J., Zheng, H., Wang, J., Yang, L., & Zhao, H. (2019). Effects of low water temperature during reproductive growth on photosynthetic production and nitrogen accumulation in rice. Field Crops Research, 242, 107587.
Khush, G. S. (2013). Strategies for increasing the yield potential of cereals: Case of rice as an example. Plant Breeding, 132, 433–436.
Lai, H., Yu, H., & Huang, J. (2014). Review on the calculation models of crop climate productive potential. Jiangsu Agricultural Science, 42, 11–14.
Li, H., Wang, X., Rengel, Z., Ma, Q., Zhang, F., & Shen, J. (2016). Root over-production in heterogeneous nutrient environment has no negative effects on Zea mays shoot growth in the field. Plant and Soil, 409, 405–417.
Li, Y. L., Fan, X. R., & Shen, Q. R. (2008). The relationship between rhizosphere nitrification and nitrogen-use efficiency in rice plants. Plant, Cell & Environment, 31, 73–85.
Liu, Z., Yang, X., Lin, X., Hubbard, K. G., Lv, S., & Wang, J. (2016). Maize yield gaps caused by non-controllable, agronomic, and socioeconomic factors in a changing climate of Northeast China. Science of the Total Environment, 541, 756–764.
Lobell, D. B., Cassman, K. G., & Field, C. B. (2009). Crop yield gaps: Their importance, magnitudes, and causes. Annual Review of Environment and Resources, 34, 179–204.
Loomis, R. S., & Williams, W. A. (1963). Maximum crop productivity: An extimate 1. Crop Science, 3, 67–72.
Mueller, N. D., Gerber, J. S., Johnston, M., Ray, D., Ramankutty, N., & Foley, J. A. (2013). Correction: Corrigendum: Closing yield gaps through nutrient and water management. Nature, 494, 380.
NBSC (2016). National Bureau of Statistics of China. http://www.stats.gov.cn/english/Statisticaldata/AnnualData/. Accessed 30 June 2016
Peng, S., Huang, J., Sheehy, J. E., Laza, R. C., Visperas, R. M., Zhong, X., Centeno, G. S., Khush, G. S., & Cassman, K. G. (2004). Rice yields decline with higher night temperature from global warming. Proceedings of the National Academy of Science USA, 101, 9971–9975.
Peng, S., Khush, G. S., Virk, P., Tang, Q., & Zou, Y. (2008). Progress in ideotype breeding to increase rice yield potential. Field Crops Research, 108, 32–38.
Ramankutty, N., Foley, J. A., Norman, J., & McSweeney, K. (2002). The global distribution of cultivable lands: Current patterns and sensitivity to possible climate change. Global Ecology and Biogeography, 11, 377–392.
Ramasamy, S., Ten Berge, H. F. M., & Purushothaman, S. (1997). Yield formation in rice in response to drainage and nitrogen application. Field Crops Research, 51, 65–82.
Saengwilai, P., Meeinkuirt, W., Phusantisampan, T., & Pichtel, J. (2020). Immobilization of cadmium in contaminated soil using organic amendments and its effects on rice growth performance. Exposure and Health, 12, 295–306.
Shen, J., Cui, Z., Miao, Y., Mi, G., Zhang, H., Fan, M., Zhang, C., Jiang, R., Zhang, W., Li, H., Chen, X., Li, X., & Zhang, F. (2013). Transforming agriculture in China: From solely high yield to both high yield and high resource use efficiency. Global Food Security, 2, 1–8.
Shimono, H., Okada, M., Kanda, E., & Arakawa, I. (2007). Low temperature-induced sterility in rice: Evidence for the effects of temperature before panicle initiation. Field Crops Research, 101, 221–231.
Shipley, B. (2006). Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A Meta-Analysis. Functional Ecology, 20(4), 565–574. https://doi.org/10.1111/j.1365-2435.2006.01135.x
Sinclair, T. R., & Horie, T. (1989). Leaf nitrogen, photosynthesis, and crop radiation use efficiency: A Review. Crop Science, 29, 90–98.
Strock, C. F., de la Riva, L. M., & Lynch, J. P. (2018). Reduction in root secondary growth as a strategy for phosphorus acquisition. Plant Physiology, 176, 691–703. https://doi.org/10.1104/pp.17.01583
Stuart, A. M., Pame, A. R. P., Silva, J. V., Dikitanan, R. C., Rutsaert, P., Malabayabas, A. J. B., Lampayan, R. M., Radanielson, A. M., & Singleton, G. R. (2016). Yield gaps in rice-based farming systems: Insights from local studies and prospects for future analysis. Field Crops Research, 194, 43–56.
Sun, B. R., Gao, Y. Z., & Lynch, J. P. (2018). Large crown root number improves topsoil foraging and phosphorus acquisition. Plant Physiology, 177, 90–104. https://doi.org/10.1104/pp.18.00234
Sun, Y. J., Ma, J., Sun, Y. Y., Xu, H., Yang, Z. Y., Liu, S. J., Jia, X. W., & Zheng, H. Z. (2012). The effects of different water and nitrogen managements on yield and nitrogen use efficiency in hybrid rice of China. Field Crops Research, 127, 85–98.
Sun, Z., Wang, X., Yamamoto, H., Zhang, J., Tani, H., Zhong, G., & Yin, S. (2017). Extraction of rice-planting area and identification of chilling damage by remote sensing technology: A case study of the emerging rice production region in high latitude. Paddy Water Environment, 15, 181–191.
Trachsel, S., Kaeppler, S. M., Brown, K. M., & Lynch, J. P. (2013). Maize root growth angles become steeper under low N conditions. Field Crops Research, 140, 18–31.
Van Ittersum, M. K., Cassman, K. G., Grassini, P., Wolf, J., Tittonell, P., & Hochman, Z. (2013). Yield gap analysis with local to global relevance—a review. Field Crops Research, 143, 4–17.
Wang, J., Zhang, J., Bai, Y., Zhang, S., Yang, S., & Yao, F. (2020). Integrating remote sensing-based process model with environmental zonation scheme to estimate rice yield gap in Northeast China. Field Crops Research, 246, 107682.
Wang, P., Hu, T., Kong, F., Xu, J., & Zhang, D. (2019). Rice exposure to cold stress in China: How has its spatial pattern changed under climate change? European Journal of Agronomy, 103, 73–79.
Wang, X., Li, T., Yang, X., Zhang, T., Liu, Z., Guo, E., Liu, Z., Qu, H., Chen, X., & Wang, L. (2018). Rice yield potential, gaps and constraints during the past three decades in a climate-changing Northeast China. Agricultural and Forest Meteorology, 259, 173–183.
Yuan, S., Linquist, B. A., Wilson, L. T., Cassman, K. G., Stuart, A. M., Pede, V., Miro, B., Saito, K., Agustiani, N., & Aristya, V. E. (2021). Sustainable intensification for a larger global rice bowl. Nature Communications, 12, 1–11.
Zelitch, I. (1982). The close relationship between net photosynthesis and crop yield. BioScience, 32, 796–802.
Zhang, B., Zhao, M., Dong, Z., Chen, C., & Sun, R. (2007). “Three combination structure” quantitative expression and high yield analysis in crops. Acta Agronomica Sinica, 33, 1674–1681.
Zhang, T., Yang, X., Wang, H., Li, Y., & Ye, Q. (2014a). Climatic and technological ceilings for Chinese rice stagnation based on yield gaps and yield trend pattern analysis. Global Change Biology, 20, 1289–1298.
Zhang, W., Cao, G., Li, X., Zhang, H., Wang, C., Liu, Q., Chen, X., Cui, Z., Shen, J., Jiang, R. J. N., Mi, G., Miao, Y., Zhang, F., & Dou, Z. (2016). Closing yield gaps in China by empowering smallholder farmers. Nature, 537, 671–674.
Zhang, Y., Tang, Q., Zou, Y., Li, D., Qin, J., Yang, S., Chen, L., Xia, B., & Peng, S. (2009). Yield potential and radiation use efficiency of “super” hybrid rice grown under subtropical conditions. Field Crops Research, 114, 91–98.
Zhang, Z., Chen, Y., Wang, C., Wang, P., & Tao, F. (2017). Future extreme temperature and its impact on rice yield in China. International Journal of Climatology, 37, 4814–4827.
Zhang, Z., Wang, P., Chen, Y., Song, X., Wei, X., & Shi, P. F. (2014b). Global warming over 1960–2009 did increase heat stress and reduce cold stress in the major rice-planting areas across China. European Journal of Agronomy, 59, 49–56.
Acknowledgements
This work was supported by the Heilongjiang Province Applied Technology Research and Development Plan Project (No. GA20B101), the Heilongjiang Province Natural Science Foundation Project (LH2020C005), the Postdoctoral Fund to Research Start-up of Heilongjiang Province (No.LBH-Q21077), National Key Research and Development of China (No. 2016YFD0300104), the Agricultural Ecological Resources and Environmental Protection Service Project of Ministry of Agriculture and Rural Affairs (No. 13220061).\
Funding
This work was supported by the Heilongjiang Province Applied Technology Research and Development Plan Project (No. GA20B101), the Heilongjiang Province Natural Science Foundation Project (LH2020C005), the Postdoctoral Fund to Research Start-up of Heilongjiang Province (No.LBH-Q21077), National Key Research and Development of China (No. 2016YFD0300104), the Agricultural Ecological Resources and Environmental Protection Service Project of Ministry of Agriculture and Rural Affairs (No. 13220061).
Author information
Authors and Affiliations
Contributions
HZ and DZ designed the experiments. YJ, HL, and YM wrote the manuscript. HZ, JW, and HZ conducted the metabolite analyses and analyzed the data. YJ carried out the experiments. HW and JW prepared the experimental materials. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no competing interests to declare that are relevant to the content of this article.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jia, Y., Liu, H., Mei, Y. et al. Analysis of Gaps Yield and Resource use Efficiency of Cold-Region Japonica Rice. Int. J. Plant Prod. 17, 17–33 (2023). https://doi.org/10.1007/s42106-022-00225-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42106-022-00225-0