
Abstract
This study assessed the effect of vegetative filter strip (VFS) in removal of suspended sediment (SS), total nitrogen, total phosphorus and Escherichia coli (E. coli) in overland flow to improve receiving water quality standards. Four and half kilograms of cowpat manure was applied to the model pasture 14 m beyond the edge of vegetated filter strip (VFS) comprising 10-m Napier grass draining into 20-m Kikuyu grass (VFS II), 10-m Kikuyu grass draining into 20-m Napier grass (VFS III) and native grass mixture of Couch–Buffel (VFS I-control). Overland flow water samples were collected from the sites at positions 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 and 30 m along the length of VFSs. E. coli removal by Napier grass VFS was on the order of log unit, which provided an important level of protection and reduced surface-flow concentrations of E. coli to below the 200 (CFU 100 mL−1) recommended water quality standards, but not for nutrients and SS. The Napier grass showed highest efficiency (99.6 %), thus outperforming both Kikuyu grass (85.8 %) and Couch–Buffel grasses VFS (67.9 ± 4.2 %) in removing E. coli from overland flow. The low-level efficiency of native Couch–Buffel grasses in reducing E. coli in overland flow was because of preferential flow. Composition and design of VFS was instrumental and could be applied with a high potential of contracting the uncertainty in improving water quality standards through mass reduction of SS, nutrients and E. coli load in watersheds.
Similar content being viewed by others
1 Introduction
Microbial faecal pathogens in manure-borne zoonotic infectious diseases draw attention because of increased morbidity and mortality in humans worldwide through both foodborne and waterborne avenues (Berry and Wells 2010). Potential routes of exposure for the public include contaminated milk or meat from livestock operations, crops from direct application of manure or manure-contaminated overland flow, and contamination into surface waters and drinking water (Bicudo and Goyal 2003). Increasing evidence illustrates a direct geographic correlation for increased risk for communities living in close proximity to food animal production areas or manure applied crop fields (Casey et al. 2015). A major challenge for both large-scale production operations and small-scale local farming is how to ameliorate impacts of livestock on surface water quality to protect human health, while maintaining production and meeting nutritional needs (Hubbard et al. 2004; Randolph et al. 2007; Gerber et al. 2010). As faecal contamination of surface water bodies by livestock feed operations can be multi-factorial in nature, various management strategies must be employed to adequately reduce public health risk, including limits on proximity to surface water sources, duration and rotation of grazing time and various mitigation strategies to reduce the contribution of overland flow of manure-borne pathogens into surface waters (Doran and Linn 1979; Edwards et al. 1997; Bicudo and Goyal 2003; Tyrell and Quinton 2003; Gerber et al. 2010; Casey et al. 2015). In recent years, vegetated filter strip (VFS) has emerged as a low-cost, locally implementable, best management practice option to minimize these health risk, particularly for small-scale operations or to reduce risk from open pasture-derived overland flow (Stout et al. 2005; Liu et al. 2008; McLaughlin et al. 2013; Yu et al. 2013). However, recent literature has emphasized the importance of determining local hydrological parameters (including soil, water quality and temperature) in the design and implementation of these VFSs (Krutz et al. 2004; Trask et al. 2004; Stout et al. 2005; Guber et al. 2009; Sabbagh et al. 2009; Islam et al. 2013) and focusing on locally implementable and sustainable solutions. As global climate change will result in increased periods of both flooding and drought in vulnerable areas, site-specific mitigation strategies to reduce public health risk of contamination of surface waters by overland flow must be explored, along with delineation of parameters influencing the efficacy of these mitigation strategies (Boxall et al. 2009; Chiang et al. 2012). VFSs are widely used and are increasingly viewed as a practical, low-cost management option for improving the quality of overland flow from non-point pollution source areas (Chaubey et al. 1995; Lim et al. 1998; Lewis et al. 2010). VFSs can remove pollutants by various mechanisms, which might include decomposition, vegetative consumption, volatilization, and infiltration, adsorption of pollutants, dilution, filtration and settling (Vanderholm and Dickey 1979; Koelsch et al. 2006; Koelsch and Ward 2009). The effectiveness of VFS for sediment removal in overland flow from strip mines, nutrients and solids removal from feedlot, pasture and cropland and treatment of wastewater was demonstrated more decades ago (Doran and Linn 1979; Gross et al. 1991; Coyne et al. 1995). However, the consistency in performance of VFS across all topography and pollutants has been questioned (Lim et al. 1998), although they have the potential of markedly improving the quality of incoming overland flow. Some studies have reported that VFS are usually not sufficient for reducing overland faecal coliform concentrations to primary contact standards (Walker et al. 1990; Coyne et al. 1995). In a study involving swine manure, it was reported that VFS did not remove significant nitrate nitrogen (NO3-N) or faecal coliforms (FC) (Chaubey et al. 1994). VFSs remove far less pollutant load for concentrated flow than for diffuse flow, indicating that the flow regime within the VFS is critically important to good performance (Dillaha et al. 1986; Gallagher et al. 2013). The exact form of improvement and design parameters for optimal performance are difficult to define, despite it being apparent that VFSs are generally capable of contributing to overland flow quality improvements (Lim et al. 1998); thus, it is very important to define local parameters and develop site-specific models. Environmental safety requires that manure should be deposited away from waters sources (Larsen et al. 1994; Edwards et al. 2000). An intervening land area or a zone separating manure and the water body of concern help in the improvement of water quality (Lim et al. 1998). As a result of uncertainty associated with predicting VFS performance, it is important to develop and test a design strategy locally (Lim et al. 1998). Martinez et al. (2013) proposed a design procedure in which VFS length depends on soil, incoming pollutant concentrations and goals with regard to pollutant reduction. Factors such as land slope have also been incorporated in the VFS length design; therefore, accurate methods for determining optimal placement, dimensions and orientation of buffer area must be developed for VFS to be most effective and economical (Edwards et al. 1996; Lim et al. 1998). The study herein on a management scale investigation advances previous soil box and overland field investigations (Lewis et al. 2005, 2009; Trask et al. 2004; Tate et al. 2006; Sullivan et al. 2007) and related investigations on Giardia duodenalis (Miller et al. 2007) and Cryptosporidium spp. (Miller et al. 2008). As mentioned, surface water sources may get contaminated from many small livestock operations located along rivers, streams, waterways and road ditches (Douglas-Mankin and Okoren 2011), with significant morbidity and mortality in human populations caused by the ingestion of water contaminated with microbial pathogens (Benham et al. 2006). Furthermore, other disease-causing bacteria are indicated by the presence of faecal coliform concentrations (Douglas-Mankin and Okoren 2011), but work has suggested limitations of coliforms as indicator organisms, particularly in tropical regions of the world (Hazen 1988; Rochelle-Newall et al. 2015). E coli indicator colony-forming units (CFUs) can provide a direct measure of potentially pathogenic bacteria, and water quality standards for E. coli bacteria in several places in the world have been established, which allow for bacterial limits in surface water depending upon its intended use (Douglas-Mankin and Okoren 2011). Currently, there are no guidelines for microbial limits in surface water for irrigation, fish farming, swimming and drinking besides the Environmental Management and Coordination Act (EMCA) of 1999 laws of Kenya (EMCA 1999). However, the EMCA law falls short of requiring limits on microbial levels for discharge into surface water for irrigation, swimming and drinking. However, World Health Organization (WHO) provides these guidelines, which include drinking water with the maximum contaminant level for E. coli set at 0 colony-forming units per 100 mL (0 CFU 100 mL−1), primary contact such as swimming (160–2358 CFU 100 mL−1), secondary contact such as boating and fishing (2358 CFU 100 mL−1) (World Health Organization 2011 ). Surface waters and subsequently human health can be impacted by a variety of contaminants through overland flow sources, including large amounts of nutrients, such as nitrogen and phosphorus, fertilizers, pesticides and other anthropogenic contaminants (Douglas-Mankin and Okoren 2011). Additionally, anthropogenic nitrogen and phosphorus can contribute to eutrophication and hypoxia, impacting freshwater ecosystems and source waters for recreation and drinking (Dale et al. 2007). Olilo et al. (2014) and Douglas-Mankin and Okoren (2011) reported that taste and odour problems in drinking water could be attributed to eutrophication and subsequent production of various metabolites in cyanobacteria blooms; these blooms can also produce acute and chronic gastrointestinal, liver, dermato- and neuro-toxins, in addition to skin and gastrointestinal allergens and irritants (Carmichael 2001). Excessive nitrogen concentrations (nitrate, NO3-N) in water taken by infants may contribute to blue baby syndrome or infantile methemoglobinaemia (Douglas-Mankin and Okoren 2011). As lagoons and vegetative filter strips are common management practices for controlling pollution from livestock feedlots (Delgado et al. 1995; Koelsch et al. 2006) into source waters, it is important to evaluate interactions between reductions in contributing factors to degrading surface water quality, particularly in vulnerable regions of the world where there is little to no treatment of drinking water. As global climate change results in increased periods of flooding and drought, particularly in vulnerable tropical areas, it is imperative to investigate both the efficacy of mitigation strategies such as VFS in a site-specific manner and understand the interrelationship between contributing factors to degrading surface water quality (Rochelle-Newall et al. 2015). The objectives of this study were several-fold: (1) to examine the impact of VFS design and vegetation on reduction of SS, nutrient concentrations and faecal indicator concentrations (FIC) temporally and spatially in a simulated heavily grazed pasture in the eastern escarpment of the Mau Forest; (2) to compare this with overland flow through a control of native grasses; (3) to determine the interrelationship with meteorological and hydrological parameters with reduction of these parameters; (4) to determine whether predictive models adequately matched observed VFS removal and the effect of VFS vegetation in improving water quality by mass reduction of E. coli in overland flow under these meteorological and hydrological parameters.
2 Materials and methods
2.1 Study site description
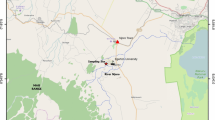
The study site was located at Tatton Agriculture Park (TAP), a livestock and crops research and demonstration unit or facility adjacent to and bordering Njoro River, Njoro campus, Egerton University, Kenya (Fig. 1). The farm itself is located 22 km from Lake Nakuru and 172 km west of Nairobi in the East African Rift Valley. TAP has a hilly topography. It is located down-slope from Mau Forest and slightly above Njoro River and accompanying underlying shrubby vegetation. It is located in the eastern escarpments of Mau Ranges that drains Lake Nakuru (1754 m above sea level), Lake Baringo (978 m above sea level) and Lake Victoria (1154 m above sea level). The study was undertaken from August 2013 to December 2014 at field 18 at TAP between 00°22′35″S and 35°55′15″E, at an elevation of 2315 m above sea level. On an annual basis, this region experiences a wet season from April to August, a short rainy season from September to December and a dry season from January to March. The experimental field at TAP and the surrounding riparian forest has a clay-loamy-sand soil type. The underlying material in these soil types is loamy-clay and Miocene age material of very low permeability.

(Source: CNES/Astrium and Google Inc, 2016)
Sampling site at Tatton Agriculture Park, eastern escarpment of the Mau Forest, Njoro River Watershed, Kenya
2.2 Vegetated filter strips
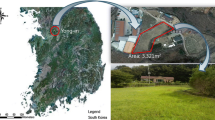
The predominant vegetation on the study site consists of mixed native African couch grass Cynodon dactylon (L.), Pers., wire grass Eleusine indica (L.) Gaertner, African bristle grass Setaria sphacelata (Schumach) Stapf & C. E. Hubb ex M. B. Moss, Buffel grass Cenchrus ciliaris L. (Marshall et al. 2012) and rescue grass Bromus catharticus Vahl. Native grasses were cleared during field preparation. Plot I (control) comprised of 30 m of mixed native Couch grass and Buffel grass, while plot II consisted of 10-m Napier grass followed by 20-m Kikuyu grass, and plot III consisted of 10-m Kikuyu grass followed by 20-m Napier grass, each of which was planted in triplicate into each of the blocks I, II, III (Fig. 2). During rainfall events from August 2013 to December 2014, field experimental trials were carried out at TAP farm, along the eastern escarpment of Mau Forest, Njoro River watershed, in the following randomized complete block design. There were nine fields divided into three blocks of three fields each and each field measured 30 m long by 4 m wide. Vegetated field plots were established 6 months prior to initiation of the experiment by ploughing fallow grassland field and planting grass vegetation at different proportions of Kikuyu grass, Napier grass and native grasses in three plot designs, each performed in triplicate. Two of the fields in each block were designed with different proportions of Kikuyu grass (Pennisetum clandestinum Hochst. ex Chiov.) and Napier grass (Pennisetum purpureum Schumach), while the third was planted with a mixture of native vegetation of Couch grass and Buffel grass (control). The fields were planted with Kikuyu grass and Napier grass because both of these grasses are important sources of livestock fodder, preferred by farmers in their livestock pastures locally in Kenya. These two grasses are widely available and used by farmers to benefit livestock. The two grasses produce high amounts of forage and resist both drought and high-intensity livestock grazing. Both locally and internationally, these grasses could be useful as buffer zone grasses to protect water sources from contamination by faecal coliforms and other contaminants. The relative density of Napier grass, Kikuyu grass and native Couch–Buffel grasses ranged from 45 to 97 % over the study period. By initiation of experiments, 100 % cover had been established on all experimental fields with mean heights of 2.0, 0.5 and 0.10–0.20 m for Napier grass, Kikuyu grass and Couch grass and Buffel grass plots, respectively. Each field was oriented with major axes of a 4 m × 44 m running up and down-slope. The fields were level across the minor axes and had a constant 15 % slope along the major axes. The upper 14 m of each field was used to represent pasture and was treated with dairy cattle manure, while the remainder of each field acted as VFS. Five-cm exposed soil borders were prepared in around the fields to isolate overland flow. Plastic gutters were installed along the length of each field, distributed at 10, 20 and 30 m from the top and separated the fields into three sections of equal area. Removable watertight covers were installed on the gutters so that the overland flow would simply cross the gutters and continue down-slope when covered. Designed covers could be easily removed to collect overland flow samples as necessary. Each gutter drained to the centre, such that overland samples could be collected through a short pipe into a bucket sampler. The gutter slope was sufficient to minimize deposition of solid material. A metallic quadrat (1 m2) was used to measure overlying leaf litter and stem residual dry matter, which included the dropped leaves and litter above the ground level in each of the Napier grass VFS, Kikuyu grass VFS and Couch–Buffel grasses VFS (control). The leaf litter and stems of these grasses from each of the field plots were then stored in cool boxes and transported to the soil science laboratory, EU, and then dried at 105 °C in an oven for 48 h to remove the moisture. The oven-dried leaf litter and stem were then weighed to the nearest 0.01 g.

Layout design of VFS in field plots II, III and I at TAP, eastern escarpment of Mau Forest, Njoro River Watershed. Block III: VFS I (Couch–Buffel grasses = control); VFS II (N = Napier grass, K = Kikuyu grass); VFS III (K, N); Block II: VFS I (Control); VFS III (K, N); VFS II (N, K); and Block I: VFS I (Control); VFS II (N, K); VFS III (K, N); CT = collecting troughs
2.3 Prediction of E. coli mass removal effectiveness by VFS
The uniform width of VFS (m) was selected because increased width helps reduce the spread of pathogens in VFSs as described by Lim et al. (1998). Along with preliminary concentration data, overland flow data from this VFS experiment were used to compute the predicted effectiveness of E. coli mass removal through a calculated flow-weighted mean overland concentrations, mass transport and mass removal effectiveness of analysis parameters. Mass removal effectiveness was calculated from (Lim et al. 1998):
where E(x) is mass removal effectiveness (%), M is mass transport (g), and x is VFS length (m). The term M(0) is mass entering VFS. The data were fitted to the following first-order exponential decay models to calculate rate coefficients that can facilitate comparison to earlier studies in cases where concentrations or mass transport was significantly affected by VFS length (Lim et al. 1998):
where C is concentration (mg L−1) of constituent I initially entering VFS, M is mass transport (mg) of constituent I transported past VFS length (m), x is VFS length, and C 0, M 0 and k are coefficients (k is the rate coefficient = 1/x). C 0 and M 0 were equal to the initial concentrations and mass transport, respectively.
As area pollutant source length appear to alter performance of VFS (Overcash et al. 1981); steady-state VFS performance in terms of reducing incoming concentrations of soluble pollutants was analysed according to Overcash et al. (1981) with slight modifications:
where P c reduction (as proportion of incoming value) in concentration of pollutant entering the VFS, P m reduction (as proportion of incoming value) in mass transport of pollutant entering the VFS, D = ratio of infiltration to total rainfall, K = ratio of VFS to pollutant source length.
2.4 Application of manure
Manure was applied to the fields at the rate of 5.8 kg ha−1 on all the fields at 14 m above the VFS in order to simulate grazed pasture, prior to every rainfall event. The tropical rainfall events were accurately predicted using the Kenya Meteorological Department weather focuses or predictions of the research area, rainfall patterns, cloud type and cover, air temperature pattern and humidity and the tropical wind patterns per season in the area. The prediction error of rainfall occurrence was ±3 days. The manure application rate was 40 kg N ha−1(gross application of 5.2 kg manure). The total experimental field area was 300 m2, so the gross application was 1200 g of manure. This rate of manure application was equivalent to the manure that would be produced from a stocking density of six 450 kg animal units/ha for a 7-day grazing duration, which would represent a heavily grazed condition. Natural rainfall was used to generate overland flow. Rainfall intensity was recorded per event at a mean of 54 mm. The cattle manure used originated from free range grazing dairy cattle within confines of Egerton University feeding on native grass, Kikuyu grass, Rhodes grass hay and Napier grass hay. The manure was collected by scooping it off a cattle pen mud floor and included no urine. The manure was collected 1 day before the experiment and refrigerated (1 °C) until applied to the fields.
2.5 Meteorological and hydrological parameters
Meteorological and hydrological data were recorded every time field sampling was carried out during natural rainfall events, throughout the sixteen months sampling period (Table 1). A YSI meter was used to measure soil temperature and soil pH during sample collection. Soil moisture content was also measured. Soil temperature (1.5 m soil depth) was also measured along the vegetated filter strips per month. Rainfall was measured from Egerton University meteorological station located within the campus at 2 km away from the experimental site using non-recording rain gauge. Irradiance was measured using radiometer at Nakuru meteorological station. Electrical conductivity (µS cm−1) was measured in the field during sample collection using a conductivity probe YSI (Yellow spring, OH, USA) multi-probe meter. Surface water samples were taken from each of the sampling buckets from the overland flow source and collected in sterile polypropylene sample containers with leak proof lids. They were placed in 125-mL Whirl-Park sample papers and put on ice in cool boxes before being transported to the laboratory for analysis. Samples for sediment and nutrient analyses were collected in acid-washed sample bottles and stored in ice.
2.6 Sample collection
Prior to manure application, approximately four soil samples (0–20 cm) were collected from each field and analysed. Soil samples were taken and analysed for total phosphorus (TP) and total nitrogen (TN). Samples were collected inside and outside of the VFS to compare if there was any difference in the nutrient content due to the continuous application of nutrient-rich run-on. Overland samples were collected manually at specified intervals (5, 10, 20, 30, 45, 60 min after the beginning of continuous overland flow) at VFS lengths of 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 and 30 m during overland flow. At a given sampling rainfall event time, an overland flow sample was collected first from the lower-most gutters draining into sterilized bucket sampler and then the next bucket sampler up the length of the experimental fields. A field assistant was stationed at each bucket sampler and began collecting the sample from the particular bucket sampler immediately following collection of the overland flow sample from the next down-slope bucket sampler during rainy events. Sample volumes were in the range of 0.1–2 L. The time required to collect each sample was recorded, so that overland rates could be computed. The rainfall water sample was also collected before reaching simulated pasture during the experiment and analysed to determine background quality of the rainfall water. All overland flow samples were weighed immediately after collection to determine sample masses and volumes. Measurements of pH were completed at the experiment site (Orion pH meter, model 290 A).
2.7 Laboratory analyses
All other laboratory analyses were performed prior to exceeding the holding times specified by Greenberg et al. (1992). Overland flow samples were refrigerated at 1 °C while awaiting analysis. The soil samples were further analysed for factors such as organic matter (particulate organic carbon and dissolved organic carbon) as described by Maciolek (1962). Total phosphorous, reactive soluble phosphorus and organic phosphorus were analysed by procedures described by Strickland and Parsons (1968, 1972). Nitrate nitrogen (NO3-N), nitrite nitrogen and organic nitrogen were analysed by the procedures described in American Public Health Association (APHA) (1992). Ammonia (NH3-N) was analysed using the standard methods described by Greenberg et al. (1992). Suspended sediment (filtration and drying method) and briefly suspended sediment concentration was analysed by filtering 50–100 mL of the sample through 0.45-μm filters using a vacuum. Zinc (Zn), total dissolved solids (TDS), were analysed as described in APHA (1992). Based on the weight difference in filter before and after filtration, suspended solids were calculated. Turbidity (nephelometric turbidity units) was measured in situ using turbidity meter. Total coliforms (TC), faecal coliforms (FC) and E. coli (CFU 100 mL−1) were analysed using standard membrane filtration method as described by Greenberg et al. (1992).
2.8 Data analyses
The observed VFS effectiveness in removal of E. coli mass from the overland flow was compared with the predicted VFS effectiveness using Nash–Sutcliffe method (1970). The significance of VFS effects was assessed using analysis of variance (ANOVA). Statistical analyses were performed as described by Zar (1996) and in PAST statistical package (Hammer et al. 2001). Analyses of variance were performed to determine the effects of VFS length on average concentration, average mass loss and average proportion of mass loss reduction for the cattle manure constituents studied. Least significant difference (LSD) testing was performed to determine significant differences in VFS length performance when analyses of variance indicated a significant overall VFS length treatment effects. All tests of significance were conducted at the p < 0.05 level.
3 Results
3.1 Predictive data model
In this study, SS approximately described the settling process. Most of that reduction (70–90 %) occurred within 10 m, which was a third of the entire length of each of the VFSs. Larger sediment particles such as sand settled more quickly, while finer particles such as clay and silt, and associated total phosphorus particles, remained suspended longer and reached outflows. Records were made of the observed and predictions of TN, TP, SS and E. coli mass transport collected from the overland flows at the exit of the vegetative length of Couch–Buffel grass VFS, Napier grass and Kikuyu grass (Fig. 3). The SS was significantly (p < 0.05) different in Napier grass, Couch–Buffel grass and Kikuyu grass, indicating that the predictive model returned higher values than the observed values. Similarly, the model also returned higher values for the total nitrogen and total phosphorus in all the VFSs. Predictive models adequately matched observed VFS removal (Fig. 3). The higher values were similarly returned for E. coli in all the VFS. The ratio of the study site length to pollutant source length in the watershed was 0.42.

Observations and predictions of SS, TN, TP and E. coli mass transport in overland flows at VFS length exit of Couch–Buffel, Napier and Kikuyu grasses from August 2013 to December 2014
3.2 Effect of VFS surface vegetation and root zone on meteorological and hydrological parameters in overland flows in the watershed
The soil at the site was clay–loam. The soil had a high water intake capacity; overland flows as a proportion of rainfall ranged from 1 to 23 % for a 73-mm h−1 natural rainfall event taken at the field’s outlet flow collecting point at the site. Type of grass vegetation substantially influenced hydrological parameters in overland flows (Table 2). Napier grass litters significantly (p < 0.05) reduced overland flows velocity more than in both Kikuyu grass and native Couch–Buffel grasses. The mean daily rainfall intensity was 54.4 mm in the eastern escarpment of Mau Forest, Njoro River Watershed. The mean overland flow rate in Napier grass was 2.5 cm−3 s−1. Soil moisture content in Napier grass was not significantly different from both Kikuyu grass and native Couch–Buffel grasses. Total dissolved solids were not significantly different between Napier grass, Kikuyu grass and Couch–Buffel grasses, respectively (Table 2). The mean total dissolved solid was 50 mg L−1 in Napier grass, which was not significantly different from Kikuyu grass VFS and Couch–Buffel grasses. The mean turbidity was 177 NTU in the Napier grass, which was significantly (p < 0.05) different from Kikuyu grass and Couch–Buffel grasses VFS.
Napier grass was effective in removing SS from the overland flow and was significantly (p < 0.05) different from Kikuyu grass VFS and Couch–Buffel grasses VFS. SS was significantly (p < 0.05) contributed from the background soil to the overland flow more than from manure, indicating that soil was a contributing factor to suspended solids. The SS ranged in speed from inflow rate of 1.16 × 103 to 4.51 × 105 g cm−1 s−1 and a mean speed of 2.83 × 104 g cm−1 s−1 in Napier grass VFS, while in both Kikuyu grass and Couch–Buffel grasses, the values were much lower and were significantly (p < 0.05) different from each other (Table 2). The mean temperature was 21 °C and did not fluctuate significantly in the overland flow along the length of Napier grass VFS, Kikuyu grass VFS and Couch–Buffel grasses VFS. The conductivity (EC) ranged from a minimum of 115 to a maximum of 220 µS cm−1 with a mean of 164 ± 5.6 µS cm−1 in the overland flow in Napier grass VFS, which was significantly different from Kikuyu grass VFS and Couch–Buffel grass VFS (Table 2). The pH ranged from a mean of 5.43 in clay loam to 5.84 in Couch–Buffel grass with a mean of 5.76 in the overland flow in Napier grass VFS, which was significantly (p < 0.05) different from Kikuyu grass VFS and Couch–Buffel grasses VFS. Although Napier grass effectively removed suspended solids as compared to Kikuyu grass and native Couch–Buffel grasses, total dissolved solids and EC were hardly removed by that VFS at the end of the filter strip length (similar to the situation in the other VFS).
3.3 Napier grass outperforms Kikuyu grass and native Couch–Buffel grasses in SS and nutrient removal in the overland flows in the watershed
Results of environmental factors and microorganisms in overland flow were computed and tabulated for the Njoro River watershed (Table 2). Analysis of variance (ANOVA) of the physico-chemical factors showed significant (p < 0.05) spatial variation of suspended sediment (SS), turbidity (NTU) and total dissolved solids (TDS), which were impacted significantly (p < 0.05) by Napier grass in the VFS. There was a significant (p < 0.05) effect of VFS length on suspended sediment concentration. The concentration of suspended solids decreased from the edge of the Napier grass along the length of VFS. Beyond 20 m VFS length, there were hardly any differences in the concentrations of suspended sediment concentration. The coefficient of variation of suspended sediment was 0.39. Beyond 10 m of Napier grass VFS, the overland flow started draining into a 20-m Kikuyu grass in the same VFS. The effective length for the removal of approximately 99 % of suspended sediment was 10 m, beyond which very insignificant quantities of suspended sediments were removed from the overland flow in Napier grass–Kikuyu grass VFS.
Nutrients also showed significant (p < 0.05) spatial variation of total nitrogen (TN), soluble reactive phosphorus (PO4-P) and total phosphorus (TP) in the overland flow and were also reduced by Napier grass in the VFS with mean PO4-P reduced to 21.26 μg L−1 and mean TP reduced to 53.25 μg L−1. Napier grass VFS was most effective at TP removal from overland flow, followed by Kikuyu grass. Napier grass removed 88 % of the total phosphorus and beyond 10 m from the edge of the field; no significant variation in the removal of the quantity of phosphorus was measured in the overland flow along the length of VFSs. Napier grass–Kikuyu grass VFS decreased mean NO3-N (0.25 mg L−1) and NH3-N (0.66 mg L−1) in the overland flow. Mean ammonium nitrogen was 13.32 μg L−1 in the overland flow of Napier grass–Kikuyu grass VFS, while TN was 53.25 μg L−1. Nitrate nitrogen (NO3-N) and ammonium nitrogen (NH3-N) also were significantly reduced by Napier grass–Kikuyu grass VFS, despite background contributions from soil into the runoff flow (with greater contribution than amount in initial manure). Mean dissolved organic carbon was 123.3 μg L−1 in overland flow of Napier grass–Kikuyu grass VFS, while mean Zinc concentration was 9.12 μg L−1 with no significant change for either nutrient along the length of this VFS.
While environmental factors measured in overland flow in Kikuyu grass–Napier grass VFS showed spatial variation (p < 0.05, Table 2), mostly decreasing from the upper part to the lower edge of the strips, these VFS were not as efficacious in rate of SS removal (Fig. 4), NO3-N removal (mean = 0.28 mg L−1), NH3-N (mean = 0.68 mg L−1), soluble reactive phosphorus (mean = 22.45 μg L−1) and total phosphorus (mean = 52.58 μg L−1) removal. In Couch grass–Buffel grass filter strip, the overland flow formed preferential flow along the length of VFS and nutrient removal was decreased relative to VFS II and III, although there was significant spatial variation of most environmental factors (Table 2); similarly, SS mass transport removal rate was slowest in the Couch–Buffel grass VFS (Fig. 4).

Effect of VFS length on mass transport of SS (mg ha−1) in overland flow in Njoro River Watershed from August 2013 to December 2014
Zinc was detected in the soil and is an essential micronutrient for plant growth. Zinc was correlated to FC and E. coli in Couch–Buffel grasses (r = 0.33), Napier grass–Kikuyu grass VFS (r = 0.43) and Kikuyu grass–Napier grass VFS (r = 0.33) (Table 3). Zinc showed a close positive association with conductivity, indicating that it contributed to the conductivity of the overland flow in relation to the Napier grass, Kikuyu grass and native Couch–Buffel grasses (Table 3). Zinc was also closely associated with the overland flow water temperature, organic carbon as well as total nitrogen and total phosphorus in all the VFSs.
3.4 Napier grass surface vegetation and root zone more effectively protected against total coliform, faecal coliform and E. coli concentration in the overland flows in the watershed
Performance of Napier grass, Kikuyu grass and the native Couch–Buffel grasses VFS was tested using root mean square error (RMSE) for cumulative runoff in VFS, Pearson product moment correlation coefficients (r) and the Nash–Sutcliffe efficiency for E. coli concentrations in the overland flow (Fig. 5) in the watershed. There was a substantially different time for runoff to travel from the 0 m at the edge to the full length of the plot with 50 s for Couch grass–Buffel grass VFS, 75 s for Kikuyu grass in VFS 100 s for Napier grass in VFS. This was predictive of efficacy of removal of E. coli (30, 15 and 1.5 % respectively), indicating the role of surface vegetation and root zone in removing E. coli. During the study period, runoff was 75 % of the natural rainfall depending upon how heavy the rains were. Overall, Napier grass outperformed Kikuyu grass and native grasses with concentrations of FC and EC significantly reduced (Table 2). There was a significant (p < 0.05) transport of E. coli through the soil surface with the infiltrating water within the Napier grass–Kikuyu grass VFS. FC concentration in the overland flow ranged from 1.41 × 100 to 65 × 101 CFU 100 mL−1 with a mean of 4565 × 101 CFU 100 mL−1 for all VFS. E. coli concentration ranged from 1.2 × 100 to 86 × 101 CFU 100 mL−1 with a mean of 56 × 101 CFU 100 mL−1 (Table 2). Faecal coliform concentrations of approximately 4 × 107 CFU 100 mL−1 were recorded in the incoming runoff flow entering Napier grass from the catchment of VFS. This was similar for other VFS. Only 1.5 % of E. coli was detected at 30 m exit of Napier grass VFS, indicating that over 98.5 % of E. coli either infiltrated into the root zone system or was filtered by the vegetative leaf litters within the VFS. Kikuyu grass outperformed the native grasses but did not simply just perform as well as Napier grass in E. coli removal though there was a net removal of the organism (Fig. 5). August 2013 values decreased for all values for all three VFS. There was not much temporal variation except that point. There was substantially less rainfall in that month which contributed to that difference and the trend observed in that month of August 2013. Due to the high infiltration within VFS, 15 % f E. coli was detected at 30-m exit of Kikuyu grass VFS. Due to the high infiltration within VFS, 30 % of E. coli was also detected at 30-m exit of Couch grass–Buffel grass VFS. Total nitrogen (TN) and total phosphorus (TP) were negatively correlated with faecal coliforms, yet positively correlated with E coli, indicating TN and TP removal adequately predicted E. coli removal by VFS, while coliforms would not necessarily adequately approximate performance for E. coli removal (Table 3).

Temporal variation of VFS model parameters in overland flows in Njoro River Watershed from August 2013 to December 2014. Note Pearson product moment correlation coefficients (r) and root mean square error (RMSE) for cumulative overland flow in VFS; r, RMSE and the Nash–Sutcliffe are given for E. coli concentrations in the overland flow from August 2013 to December 2014
Increased leaf and stem cover was observed for Napier Grass (422 kg ha−1) relative to Kikuyu grass VFS (245 kg ha−1) or native Couch–Buffel grasses VFS (198 kg ha−1); furthermore, the observed leaf and stem cover included the dropped leaves and litter above the ground level. Residual dry matter reduced the transport capacity of E. coli more significantly (p < 0.05) in Napier grass better than in both Kikuyu grass and Couch–Buffel grasses in VFS, indicating that there was reductions in E. coli transport in the overlying surface vegetation improving infiltration capacity of overland flow.
3.5 Effect of VFS surface vegetation and root zone on mass losses of SS and nutrient in overland flows collected over time in the watershed
Napier grass VFS removed significant (p < 0.05) proportions of incoming SS. This was indicated through computations of mass transport of SS (Fig. 4). No significant effect on mass transport of the SS was observed beyond 10 m length of Napier grass from the upper edge of the VFS. Mass transport of SS ranged from 10 to 140,000 mg ha−1 with a mean of 46,666.6 mg ha−1 for all VFS. Kikuyu grass–Napier grass VFS also removed significant (p < 0.05) proportions of incoming SS and nutrient pollutants. The Spearman’s correlation coefficient indicated that the incoming pollutants in the overland flow had significant interactions with these VFS (Table 3). Although Couch grass–Buffel grass VFS also removed significant (p < 0.5) proportions of incoming SS and nutrient pollutants, the most daunting challenge in this set of VFS was the frequent formation of preferential flow. The preferential flow was the limiting factor in this set of VFS, which jeopardized the removal of suspended solid mass considerably. Mass transport of nutrients such as ammonium nitrogen ranged from 10 to 4500 mg ha−1 with a mean of 2250 mg ha−1 in the overland flow through all VFS. Total nitrogen mass ranged from 12 to 11,800 mg ha−1 with a mean of 737 mg ha−1 (Fig. 6). TN was impacted upon further along the VFS up to and including the final outlet, 30 m down the edge of the VFS in the watershed. A significant (p < 0.05) decrease in mass transport of total phosphorus was recorded as VFS length increased from 0 to 10 m, but there was no significant difference in the mass after 10 m VFS length (Fig. 7). VFS could hardly retain nutrients, and nutrient losses from grazed pasture were not significant from agronomic point of view. Analysed data indicate that after 30 m of Napier grass–Kikuyu grass VFS and Kikuyu grass–Napier grass VFS length, a net of 86 % of runoff inflow water had been removed, possibly by infiltration and approximately 86 % of sediment had been removed, probably due to sedimentation or mass advection into soil with infiltrated water.

Effect of VFS lengths on mass transport of TN (mg ha−1) in overland flow in Njoro River Watershed from August 2013 to December 2014

Effect of VFS lengths on mass transport of TP (mg ha−1) in overland flow in Njoro River Watershed from August 2013 to December 2014
3.6 Effect of surface vegetation and root zone on mass losses of total coliform, faecal coliform and E. coli in overland flow over the length of the VFS
All VFS shared two significant findings, despite differences in overall performances of mass losses of TC, FC and E. coli over their lengths. E. coli population concentration in the overland flows were affected by VFS length showing a decrease from 0 to 10 m; thereafter, very little decrease was observed (Fig. 8). In addition, no VFS had more outlet than inlet mass of microorganisms and trapping efficiency was directly related to the existing grass density (Fig. 9). The optimal stocking density of TAP was ten 460 kg dairy cow units/ha for 7-day grazing duration. This density was proportional to the overgrazed pastureland, which produces manure load and E. coli load that could potentially pollute the Njoro River, based on the figures recorded during this study.

Effect of VFS lengths on mass transport of E. coli (CFU 100 mL−1 ha−1) in overland flow in Njoro River Watershed from August 2013 to December 2014

Effect of VFS relative density on mass reduction efficiency (%) of E. coli by different grasses in Njoro River Watershed from August 2013 to December 2014
SS removal by Napier grass (p < 0.05) was associated with E. coli removal (Table 4; df = 35, r = 1, p < 0.05) in all VFS. This study shows that Napier grass VFS significantly (p < 0.05) trapped E. coli better than either Couch–Buffel grasses or Kikuyu grass. There was a continuous and sudden release of microorganisms in flowing water trapped in the Napier grass VFS and presence of a large group (clump) of bacteria in the samples. SS removal by Kikuyu grass was not significant (p < 0.05). SS removal by Couch–Buffel grass was not significantly different. The mean mass of FC and E. coli bacterial population removals at 30 m ranged from 39.6 to 99.6 CFU 100 mL−1 with a mean of 69.9 CFU 100 mL−1 for all VFS. The mean mass of E. coli bacterial population removals at 30 m ranged from 39.8 to 100.0 CFU 100 mL−1 with a mean of 69.9 CFU 100 mL−1. These removals translated into log reductions of 0.65–1.0. The mean percentage root zone retention of E. coli by VFS from August 2013 to December 2014 indicated that Napier grass showed the highest retention efficiency even beyond the 10 m via this mechanism (Fig. 10).

Mean percentage root zone retention of E. coli by VFS from August 2013 to December 2014
While all three vegetation types significantly reduced all indicators (p < 0.05) with Kikuyu achieving close to the same degree of E. coli mass flow removal after 10 m as Napier grass (Fig. 8), Couch grass–Buffel grass VFS did not perform as well as the Napier and Kikuyu grass VFS. The limiting factor in the Couch grass–Buffel grass was the formation of the preferential flow or channels within the VFS. The preferential flow could be the reason why most of these indigenous plants performed poorly in removing the microorganisms and hence less mass reduction from the overland flow in the extrapolated entire Njoro River Watershed. There were significant (p < 0.05) variations shown between the same VFS in different blocks I, II and III from August 2013 to December 2014 in the Njoro River Watershed (Table 4).
4 Discussion
4.1 Predictive model
The predictive models adequately matched observed VFS removal. However, mass transport values of the parameters predicted were higher than the values observed using the model, though they explained the relationship between the concentration and load of SS, total nitrogen, total phosphorus and E. coli in the watershed. The model revealed that die-off was the second important source of microbial loss on the vegetation surface of all VFS. Losses in the overlying litter and surface vegetation resulted from sunlight radiation and surface temperature and high soil moisture saturation. This study revealed that quantitative evidence for each of these processes could help explain patterns of performance among different contaminants and their relationships to VFSs length and vegetation composition of Napier grass, Kikuyu grass and the native Couch–Buffel grasses. This indicates that it could be valid to utilize these model relationships to design VFS for managing contaminants in affected watersheds. This model could be used both for strategic management and tactical management of the water quality in the agricultural watershed. Further understanding of Napier grass, Kikuyu grass and the native Couch–Buffel grasses performance mechanisms of optimal vegetation lengths and densities could help guide field-based agricultural overland flow management. Sustainability (ecological, economic and social) of VFS at the land–animal–human interface would benefit greatly from further agricultural water quality studies investigating the field performance of combined VFS over multiple seasons and years and variable land areas and hydrological properties.
4.2 Effect of different grass species, surface vegetation and root zone on mass losses of suspended sediment and load in overland flows in the watershed
The overlying surface litter layer of Napier grass vegetation improved the storage of suspended sediment in surface layer through the removal process. It is hypothesized that surface vegetation-related effects within the vegetation types may have caused the slowing down of the overland flow velocity. The runoff flow conditions were reduced by VFS width and the surface overlying litter layer of the cow pat wastewater. Suspended sediment removal often occurred in the first 10 m of Napier grass VFS enhanced by the overlying surface litter layer from leaf drops and also in all other VFSs, consistent with the previous literature (Liu et al. 2008). Settling caused by decreased runoff flow velocity was the initial removal mechanism for suspended sediment. Similarly, Douglas-Mankin and Okoren (2011) reported that when wastewater was pumped from lagoons or storage structures the distributed soils were removed evenly in along the length of VFS.
4.2.1 Napier grass performance on suspended sediment settling
Sediment settling was greatest in Napier grass portion of the VFS. It is likely the Napier grass root zone soil system helped in enhanced SS settling via adsorption of particulate organic carbon, which helps in retention, degradation of chemicals through plants roots, bacteria, soil micro- and macro-fauna. The root zone of Napier grass was important in initiating a mechanism in slowing down observed pulses of microorganisms, indicating why Napier was more effective than Kikuyu grass in net reductions. We hypothesize that infiltration into subsoil zone, and backwater effect at the leading edge of the VFS, could have resulted in the decrease in the runoff flow velocity by Napier grass than Kikuyu grass and native Couch–Buffel grasses. Ling et al. (2005, 2006a, b) had suggested a decrease in the runoff flow velocity at the edge of grassed vegetation. Napier grass VFS in field plot II removed a significant (p < 0.05) amount of suspended sediment ranging from 30 to 600 mg with a mean of 370 mg, turbidity ranging from 80 to 500 NTU, with a mean of 289 NTU, but insignificant total nitrogen, conductivity and ammonium nitrogen from incoming runoff, supporting this hypothesis. Up to 88.7 % of suspended sediment was reduced by Napier grass between the inlet and outlet of the filter strip. More than three quarters of the suspended sediment was measured in the root zone system of Napier grass. This finding is consistent with the 70–90 % range reported in most studies found in the literature (Koelsch and Ward 2009). The suspended sediment concentration variation within this range was attributed to the site-specific conditions contributed by Napier grass’ large adventitious root zone system as the clay loam soil was uniformly distributed in the entire VFSs field plots. Lowrance et al. (1998) and Schmitt et al. (1999) made similar reports while working on filter strip performance and processes for different vegetation, widths and contaminants in surface overland flow from agricultural fields. Similarly, Lee et al. (2000) reported that switch grass buffer are effective in trapping coarse and sediment-bound nutrients, while switch grass–woody plants combination are effective in trapping clay and soluble nutrients from agricultural runoff flow. Some reports have also indicated that infiltration is higher under long-term forest vegetation than under long-term grass (Mankin et al. 2006), suggesting that grass–grass design may ultimately outperform grass alone. The Napier grass drained into a 20-m Kikuyu grass; future investigations could look more closely into potential mixed grass–grass design scenarios to optimize removal via different mechanisms.
4.2.2 Kikuyu grass performance on suspended sediment settling
The overland flow event swept nearly 87 % of the suspended sediment in Kikuyu grass VFS, in nearly all the 16 overland flow events following cowpat litter application to the Kikuyu grass. The cowpat from the source area increased hydraulic and contaminant loadings to Kikuyu grass as well as altering performance of Kikuyu grass VFS. The performance of Kikuyu grass VFS was influenced by five factors namely: (1) the length of VFS from the cowpat application area along the width of the filter strip; (2) soil hydraulics; (3) cowpat contaminant loadings; (4) type of vegetation leaf litter, stem and root zone system of Kikuyu grass; and (5) incoming overland flow rate and volume, suggesting that a mixed Napier–Kikuyu could have ameliorated some of this for example if there was 5 m or even 2 m of Napier followed by 5 or 8 m Kikuyu, or mixed vegetation plot throughout. Examination of Kikuyu grass surface vegetation indicated this was the primary mechanism of sorption of contaminants to, or remobilization and contribution to outflow of VFSs. Dilution of runoff water by natural rainfall during this study showed reduced concentrations of suspended solids, nutrients and E. coli in Napier grass more than Kikuyu grass and native Couch–Buffel grasses.
The root zone system of Napier grass, Kikuyu grass and Couch–Buffel grasses VFS filtered the entering overland flow contaminants by three possible pathways, namely: firstly, infiltration of nutrients and micro- and macro-fauna; secondly, sediment and sediment-bound particles; and thirdly, adsorption onto soil macro-pores particles and crevices and plant roots and overlying leaf surfaces including stomata and stem bark. Limited settling was observed in the initial 2-m length of the Kikuyu grass until after 3 m of the entire VFS measuring 20 m, which trapped a significant (p < 0.05) amount of suspended sediment through settling process. A possible explanation could be that the overland flow rate capacity was not stopped suddenly by the VFS and the flow volume was not resisted by the VFS. Sediment concentration was reduced by nearly 70–90 % in Kikuyu grass VFS.
4.2.3 Couch–Buffel indigenous grasses performance on suspended sediment settling
Surface vegetation of Couch–Buffel native grass species had the entire filter strip mixed with an equal proportion of both grasses. Suspended sediments to be trapped at the first 3 m, but the indigenous grass developed preferential flow within the VFS, jeopardizing the overall mass reduction of suspended sediment. Couch–Buffel grasses overlying surface vegetation and root zone systems filtered entering overland flow water by removing mass of the constituents past the sampling sites. The Couch–Buffel grasses effectiveness in removing the cowpat constituent mass transport was limited by preferential flow caused by different parts of the indigenous VFS grasses. Root zone of Couch–Buffel grass was not efficiently utilized because of the development of channel or preferential flows within the VFS.
Temperature was one of the most important environmental factors and did not show significant spatial variation along the length of Napier grass VFS, nor Kikuyu grass and Couch–Buffel grasses in the eastern escarpment of Mau Forest, Njoro River Watershed. Temperature variation was minimal since the vegetation height was always kept low and uniform among each plant species. Secondly, the tropical sun provided sufficiently high sunlight radiation that raised the soil temperatures during day followed by temperature inversion at the root zone during the night (Olilo et al. 2016).
4.3 Effect of different grass species, surface vegetation and root zone on mass losses of nutrients concentration and load in overland flows in the watershed
Napier grass overlying surface vegetation (10 m) in the first stretch of VFS removed significant (p < 0.05) concentrations and load of soluble reactive phosphorus (PO4-P), total phosphorus (TP) and total nitrogen (NO3-N). The measured nitrogen content included total nitrogen, ammonium and ammonia and nitrate. NO3-N removal by Napier grass VFS was much lower, and effluent concentration often exceeded that of influent, which indicated that nitrification was occurring in the VFS. Even VFS that exhibited increasing NO3 concentrations such as Napier grass in the watershed typically resulted in overall N reductions. Ammonia volatilization was a process occurring in VFS that contributed to N reduction and odour-related by-products. During a shallow, uniform and slow velocity runoff flow, a small proportion of less than 30 % ammonia was lost to the atmosphere resulting in slightly higher reductions of total N and ammonia in the surface water outflow.
Total phosphorus removal reflected the effectiveness of the Napier grass for solids removal. Phosphorus readily sorbed to soil particles such as clays that were carried by the runoff flow. Similar to solids removals, reductions percentages for total phosphorus ranged between 70 and 99 %, while ortho-phosphorus or soluble reactive phosphorus was soluble, and thus removal efficiency was consistently lower than that of total phosphorus in VFS. The results also corroborate previous studies that vegetated filter strips are capable of retaining water and pollutants associated with agricultural and livestock feedlot overland flows resulting from natural rainfall (Schmitt et al. 1999; Koelsch et al. 2006; Douglas-Mankin and Okoren (2011).
4.4 Effect of different grass species and surface vegetation on mass losses of total coliform, faecal coliform and E. coli in overland flows in the watershed
4.4.1 Overlying vegetation
Although reductions of one order of magnitude, which was approximately about 1 log10, provided some protection of water resources, these reductions may still allow a large number of bacteria to exit the VFS. These VFSs added a level of protection that would allow outflow to meet current limits for faecal coliform bacteria in streams, but not drinking water and contact limits, which are 0 and 200 CFU 100 mL−1, respectively, according to the World Health Organizations (WHO)-recommended standard. The average reductions from Napier grass VFS in field plot II reduced the E. coli average outflow at 30 m by 99.6 % to 115 CFU 100 mL−1 (or <200 CFU 100 mL−1) and faecal coliform average outflow at 30 m by 100 % to 0 CFU 100 mL−1. In this type of VFS design, there is both environmental advantage and agricultural production consideration. The increasing VFS length does not translate into corresponding increases in effectiveness, suggesting a limit to the reductions possible from Napier grass VFS in field plot II. This could be possibly because of the existence of background level of these microorganisms and other constituents such as dissolved and particulate organic carbon that could not be surpassed in this field experiment. Surface overlying vegetation of Napier grass VFS removed up to 40.0 % significant (p < 0.05) concentration and load of E. coli, faecal coliform and total coliform from incoming overland flow. Although significant variation over time and space in total coliforms, faecal coliforms and E. coli reductions in comparison with solids and nutrients, Napier grass with a ground cover of 97 % is capable of turning good management of VFS into a reliable means of controlling environmental contaminants such as microorganisms to substantially reduce public health risks from overland flow into critical drinking, irrigation and swimming surface water supplies. Interestingly, this study showed only a 0.33 Spearman’s correlation between faecal coliforms and E. coli, indicating that coliforms may not sufficiently predict removal of pathogenic organisms.
E. coli in overlying surface vegetation of Napier grass could not be detected 7 days after natural rainfall events. Four reasons could explain microbial population drop in this watershed—losses resulting from infiltration, surface runoff, pulses and decay. Yet this study demonstrates that overland flow losses contribute to a small percentage of microbial organisms released from the cowpat as trapped microorganisms would be re-leased in pulses in these systems, because of either the accumulation of microorganisms or biofilm formation and later due to increased loading contributions of microorganisms to the water supply or overland flow. Three reasons could explain observed results of better performance of Napier grass than kikuyu grass at least at the exit: (1) the overlying surface litters of Napier grass and the root zones provide larger surface area to concentrate nutrients, suspended solids as well as faecal coliforms and E. coli; (2) the root zone of Napier grass through the mechanism of interflow of the overland flow ensured that faecal coliform and E. coli penetrated into the subsoil horizon through soil macro-pores; and (3) adsorption of E. coli onto the soil particles of the subsoil horizon. Napier grass leaf litter and stem had accumulated into residual dry matter and helped moderate E. coli infiltration through temperature and moisture fluctuations, which provided a cool, moist and nitrogen and phosphorus to promote the survival and multiplication of E. coli in VFS.
Similarly, Kikuyu grass in field plot III provided the suitable condition, which was favourable for E. coli survival, growth and multiplication, better than Couch–Buffel grass VFS. Surface overlying vegetation of Kikuyu grass with up to 99.7 % sorption the highest values of 99.6 % in Napier grass VFS compared to about 95.5 % soil cover of the Kikuyu grass VFS, while surface overlying vegetation of Couch–Buffel grass VFS in field plot I had the lowest mean percentage removals of faecal coliform and E. coli, respectively, at 46.3 and 39.8 %, although it had equivalent soil cover of 95.5 %. It was hypothesized the size of the leaves and the low quantity of litter and stem dry matter that did not help reduce the runoff flow in the Couch–Buffel grasses VFS (control).
4.4.2 Root zone retention of E. coli by Napier grass
Infiltration also contributed a significant proportion of the microbial population loss not only on overlying surface vegetation, but also into the root zone of the Napier grass VFS. The root zone of Napier grass showed that contact of bacteria with clay loam soil particles during infiltration and percolation showed higher bacterial detention and removal process in the field plots in the watershed. Infiltration of the overland flow resulted into more E. coli leaching into the root zone of Napier grass, implying that root zone was the most important interaction area of the E. coli and plant material in the Napier grass VFS. Large quantities of infiltrated water leached with over 50 % of the E. coli into the root zone system of Napier grass through soil macro-pores into the subsoil horizon. Similar findings were recently suggested in complementary study in Njoro River Watersheds in Kenya (Olilo et al. 2016). However, reductions in E. coli populations could also be attributed to the inability of E. coli to lower its metabolic rate to survive in the presence of low amounts of utilizable dissolved organic carbon and to compete with other micro-biota population (Sweeney et al. 1996) in the root zone system of Napier grass in the watershed (Olilo et al. 2016). Colonies of other micro-biota could be competing with the invading E. coli for the available nutrients including limited nitrate nitrogen and dissolved organic carbon in the dry, moist and wet clay loam root zone system of Napier grass. With the shortage of the nutrients in the root zone of Napier grass, high soil temperatures could stress E. coli populations during day and temperature inversions at night, acidic soil pH and high ultraviolet radiation from the tropical sunlight (Olilo et al. 2016).
Two processes occurred in the root zone soil system of Napier grass, namely retention of E. coli cells onto the soil macro-pores due to infiltration and filtration of overland flows and adsorption of E. coli cells onto the root zone soil particles. These processes improved interaction between E. coli and root zone system soil particles in the Napier grass VFS. The mass balance resulted into equilibrium in the root zone soil–water system of Napier grass VFS. Three stages were visualized in the processes of E. coli infiltration into root zone system of Napier grass movement into soil matrix, namely individual cells, attached cells to soil particles and adsorbed cells both individually and clumped together along the soil macro-pores column. Studies have shown that vegetated buffer strips can reduce faecal coliform bacteria by more than 99 % (Pachepsky et al. 2006; Tate et al. 2006; Sullivan et al. 2007; Cardoso et al. 2012). Grassland VFS is an effective method for reducing animal pasture inputs of overland flow E. coli into surface water systems (Tate et al. 2006; Duchemin and Hogue 2008; Davis et al. 2009, 2012). While values of faecal coliform and E. coli reductions in this study indicated lower values compared to figures available for simulated rainfall provided in literature (Douglas-Mankin and Okoren 2011), the result corroborates Mankin et al. (2006, 2007), who reported that when bacterial solution was passed through disturbed soil columns, there was a strong sorption of E. coli to clay loam soil.
The most effective removal of E. coli was attributed to Napier grass, which formed over 97.5 % cover of the soil. In this study, the effect of vegetated filter strips length to waste source length ratio was significant. The VFS length to waste source area ratio was 2.5. The ratio of VFS study site length to pollution source length was 0.42. Area ratio was significantly related to E. coli population (mass) reduction. This concurs with related studies, which pointed out that treatment rates increase as the VFS to waste area ratio increases (Dillaha et al. 1988; Dickey and Vanderholm 1981; Chaubey et al. 1994; Lewis et al. 2010; Martinez et al. 2013; Mohanty et al. 2013; Allaire et al. 2015; Rippy 2015). Furthermore, this study agrees with that of Mankin et al. (2006) who reported that average E. coli concentrations reduction to range from 67.5 to 96.0 % on four VFSs in Kansa USA. Mankin et al. (2006) also reported that the area ratio was significantly related to E. coli population (mass) reduction, with better treatment for VFS with area ratios >0.5, but these estimates were made at VFS inflow and outflow volumes, which limited accuracy of the nutrient mass and bacterial population reduction analyses (Mankin et al. 2006; Douglas-Mankin and Okoren 2011). Studies gave values ranging from 6 to 99 %, and a reduction of sediment delivery from 15 to 99 % (Buckhouse and Gifford 1976; Chaubey et al. 1994; Larsen et al. 1994; Coyne et al. 1998; Schmitt et al. 1999; Guber et al. 2011). Napier grass vegetation could enhance the management of watersheds, which face threats from livestock, agricultural, wildlife and human sewer non-point source pollution through the three main factors influencing watershed pollution: the efficiency of hydrologic transport system that enables contaminants in the overland flow to enter watershed aquatic systems, mechanisms that supply watershed with microbial contaminants and mechanisms that inhibit or attenuate microbial contaminants load (Tate et al. 2006; Ling et al. 2009).
5 Conclusions
In the heavily grazed simulation, VFSs design significantly impacted improvement of water quality to meet performance criteria with superior mass reduction of E. coli, nutrients and sediment concentration and load in overland flow by Napier grass as compared to Kikuyu grass and a mixture of indigenous Couch grass–Buffel grass in the watershed. From modelling and comparison of predicted versus observed mass reductions, SS settling, infiltration and dilution are important processes controlling the performance of VFSs. The VFS proportions of Kikuyu grass and Napier grass established were effective in sequestering E. coli load in the overland flows for improved surface water quality standards for domestic, recreational, irrigational, fisheries, aquaculture and wildlife uses. E. coli removal by VFS was on the order of log unit, which provided an important level of protection and reduced surface-flow concentrations of E. coli to below the 200 CFU 100 ml−1 WHO water quality guidelines. As waters that receive overland flow from livestock pastoral grounds may contain unacceptably high concentrations of pathogens and indicator organisms, along with excessive inputs of nitrogen (N) and phosphorus (P) that could accelerate eutrophication of water bodies by promoting growth of aquatic weeds such as algae, it is important to critically evaluate VFS design in a synergistic fashion. While the VFS design herein has the potential to improve overland flow and meet water quality standards for the Mau Forest region of the Njoro River Watershed, future work must critically evaluate temporal and spatial variation. Long-duration rainfall with high-volume overland flow created more E. coli extraction and fast release (desorption) from the cowpat resulting into high entrainment at the overlying vegetation and soil surface with the consequence of infiltration into root zone and interflow and subsoil horizon. It is critical to look at further mixed grass–grass designs to determine whether efficacy of reduction of SS, nutrient and faecal indicators organisms in the watershed can be improved. Future studies based on improving the management of preferential flow among the indigenous grass VFS could be important in managing agricultural, livestock and human sewer systems forming non-point sources of pollution in these agricultural and rangeland watersheds.
References
Allaire SE, Sylvain C, Lange SF, Thériault G, Lafrance P (2015) Potential efficiency of riparian vegetated buffer strips in intercepting soluble compounds in the presence of subsurface preferential flows. PLoS ONE 10(7):e0131840. doi:10.1371/journal.pone.0131840
American Public Health Association (1992) Standard methods for the examination of water and wastewater, 18th edn. American Public health Association, New York
Benham BL, Baffaut C, Zeckoski RW, Mankin KR, Pachepsky YA, Sadeghi AA, Brannan KM, Soupir ML, Habersack MJ (2006) Modeling bacteria fate and transport in watersheds to support TMDLs. Trans ASAE 49(4):987–1002
Berry ED, Wells JE (2010) Escherichia coli O157:H7: recent advances in research on occurrence, transmission, and control in cattle and the production environment. Adv Food Nutr Res 60:67–117
Bicudo JR, Goyal SM (2003) Pathogens and manure management systems: a review. Environ Technol 24(1):115–130
Boxall AB, Hardy A, Beulke S, Boucard T, Burgin L, Falloon PD, Haygarth PM, Hutchinson T, Kovats RS, Leonardi G, Levy LS, Nichols G, Parsons SA, Potts L, Stone D, Topp E, Turley DB, Walsh K, Wellington EM, Williams RJ (2009) Impacts of climate change on indirect human exposure to pathogens and chemicals from agriculture. Environ Health Perspect 117(4):508–514
Buckhouse JC, Gifford GF (1976) Water quality implications of cattle grazing on a semiarid watershed in southeastern Utah. J Range Manag 29:109–113
Cardoso F, Shelton D, Sadeghi A, Shirmohammadi A, Pachepsky Y, Dulaney W (2012) Effectiveness of vegetated filter strips in retention of Escherichia coli and Salmonella from swine manure slurry. J Environ Manag 110:1–7. doi:10.1016/j.jenvman.2012.05.012
Carmichael WW (2001) Health effects of toxin–producing cyanobacteria: “The CyanoHABs”. Hum Ecol Risk Assess 7(5):1393–1407
Casey JA, Kim BF, Larsen J, Price LB, Nachman KE (2015) Industrial food animal production and community health. Curr Environ Health Rep 2(3):259–271
Chaubey I, Edwards DR, Daniel TC, Moore PA Jr, Nichols DJ (1994) Effectiveness of vegetative filter strips in retaining surface-applied swine manure constituents. Trans ASAE 37:845–850
Chaubey I, Edwards DR, Daniel TC, Moore PA Jr, Nichols DJ (1995) Effectiveness of vegetative filter strips in controlling losses of surface-applied poultry litter constituents. Trans ASAE 38:1687–1692
Chiang LC, Chaubey I, Hong NM, Lin YP, Huang T (2012) Implementation of BMP strategies for adaptation to climate change and land use change in a pasture-dominated watershed. Int J Environ Res Public Health 9(10):3654–3684
Coyne MS, Gilfillen RA, Rhodes R, Blevins RL (1995) Soil and fecal coliform trappings by grass filter strips during simulated rain. J Soil Water Conserv 50:405–408
Coyne MS, Gilfillen RA, Villalba A, Zhang Z, Rhodes R, Dunn L, Blevins RL (1998) Fecal bacteria trapping by grass filter strips during simulated rain. J Soil Water Conserv 53:140–145
Dale V, Bianchi T, Blumberg A, Boyton W, Conley DJ, Crumpton W et al. (2007) Hypoxia in the Northern Gulf of Mexico. An Update by the EPA Science Advisory Board EPA-SAB-08-003. EPA Science Advisory Board: Washington DC, USA. pp 333
Davis A, Hunt W, Traver R, Clar M (2009) Bioretention technology: overview of current practice and future needs. J Environ Eng 135(3):109–117. doi:10.1061/(ASCE)0733-9372(2009)135:3(109)
Davis A, Traver R, Hunt W, Lee R, Brown R, Olszewski J (2012) Hydrologic performance of bioretention storm-water control measures. J Hydrol Eng 17:604–614. doi:10.1061/(ASCE)HE.1943-5584.0000467
Delgado AN, Periago EL, Viqueira FD (1995) Vegetated filter strips for wastewater purification: a review. Bioresour Technol 51(1):13–22
Dickey EC, Vanderholm DH (1981) Vegetative filter treatment of livestock feedlot runoff. J Environ Qual 10:279–284
Dillaha TA, Sherrard JH, Lee D (1986) Long-term effectiveness and maintenance of vegetated filter strips. VPI-VWRRC-Bulletin 153, Virginia Polytechnic Institute and State 7 University, Blacksburg, USA
Dillaha TA, Sherrard JH, Lee D, Mostaghimi S, Shanholtz VO (1988) Evaluation of vegetative filter strips as a best management practice for feed lots. J Water Poll Control Fed 60(7):1231–1238
Doran JWDM, Linn TT (1979) Bacteriological quality of overland water from pastureland. Appl Environ Microbiol 37:985–991
Douglas-Mankin KR, Okoren CG (2011) Field assessment of bacteria and nutrient removal by vegetative filter strips. Intl J Agric Biol Eng 4(2): 1–7. Open Access at http://www.ijabe.org
Duchemin M, Hogue R (2008) Reduction in agricultural non-point source pollution in the first year following establishment of an integrated grass/tree filter strip system in southern Quebec (Canada). doi:10.1016/j.agee.2008.10.005
Edwards DR, Daniel TC, Moore PA Jr (1996) Vegetative filter strip design for grassed areas treated with animal manures. Appl Eng Agric 12(1):31–38
Edwards DR, Coyne MS, Vendrell PE, Daniel TC, Moore PA Jr, Murdoch JF (1997) Fecal coliform and Streptococcus concentrations in runoff from grazed pastures in Northwest Arkansas. J Am Water Resour Assoc 33(2):413–422
Edwards DR, Larson BT, Linn TT (2000) Overland nutrient and fecal coliform content from cattle manure application to fescue fields. J Am Water Resour Assess 36:711–724
Environmental Management and Coordination Act (1999) Act No 8. of 1999 of the Laws of Kenya, Nairobi, Kenya. 79 pp
Gallagher DL, Lago K, Hagedorn C, Dietrich A (2013) Effects of strain type and water quality on soil-associated Escherichia coli. Int J Environ Sci Dev 4(1):25–31
Gerber PJ, Vellinga TV, Steinfeld H (2010) Issues and options in addressing the environmental consequences of livestock sector’s growth. Meat Sci 84(2):244–247
Greenberg AF, Clesceri LS, Eaton AD (1992) Standard methods for examination of water and waste water, 18th edn. American Public Health Association, Washington, DC
Gross CM, Angle JS, Hill RL, Weterlen MS (1991) Runoff sediment losses from tall fescue under simulated rainfall. J Environ Qual 20(1):604–607
Guber AK, Yakirevich AM, Sadeghi AM, Pachepsky YA, Shelton DR (2009) Uncertainty evaluation of coliform bacteria removal from vegetated filter strip under overland flow condition. J Environ Qual 38(4):1636–1644
Guber AK, Pachepsky YA, Yakirevich AM, Shelton DR, Sadeghi AM, Goodrich DC, Unkrich CL (2011) Uncertainty in modelling of fecal coliform overland transport associated with manure application in Maryland. Hydrol Process 25:2393–2404. doi:10.1002/hyp.8003
Hammer ØD, Harper DAT, Ryan PD (2001) Paleontological Statistics (PAST): paleontological statistics software package for education and data analysis. Palaeontol Electron 1(34):4–9
Hazen TC (1988) Fecal coliforms as indicators in tropical waters: a review. Tox Assess 3(5):461–477
Hubbard RK, Newton GL, Hill GM (2004) Water quality and the grazing animal. J Anim Sci 82E(Suppl):E255–E263
Islam N, Sadiq R, Rodriguez MJ, Francisque A (2013) Evaluation of source water protection strategies: a fuzzy-based model. J Environ Manag 121:191–201
Koelsch R, Ward J (2009) Best Management Practices (BMPs) to prevent manure pathogen movement to water resources. http://www.extension.org/pages/Best_Management_Practices_(BMPs)_to_Prevent_Manure_Pathogen_Movement_to_Water_Resources
Koelsch RK, Lorimor JC, Mankin KR (2006) Vegetative treatment systems for management of open lot runoff: review of literature. Appl Eng Agric 22(1):141–146
Krutz LJ, Senseman SA, McInnes KJ, Hoffman DW, Tierney DP (2004) Adsorption and desorption of metolachlor and metolachlor metabolites in vegetated filter strip and cultivated soil. J Environ Qual 33(3):939–945
Larsen RE, Miner JR, Buckhouse JC, Moore JA (1994) Water-quality benefits of having cattle manure deposited away from streams. Bioresour Technol 48:113–118
Lee KH, Isehart TM, Schultz RC, Michelson SK (2000) Multispecies riparian buffers trap sediment and nutrients during rainfall simulations. J Environ Qual 29:1200–1205
Lewis DJ, Atwill ER, Lennox MS, Hous L, Karle B, Tate KW (2005) Linking on farm dairy management practices to storm-flow fecal coliform loading for California coastal watersheds. J Environ Monit Assess 107:407–425
Lewis DJ, Atwill ER, Lennox MS, Pereira MD, Miller WA, Conrad PA, Tate KW (2009) Reducing microbial contamination in storm runoff from high use areas on California coastal dairies. Water Sci Technol 60(7):1731–1743. doi:10.2166/wst.2009.561
Lewis DJ, Atwill ER, Lennox MS, Pereira MD, Miller WA, Conrad PA, Tate KW (2010) Management of microbial contamination in storm runoff from california coastal dairy pastures. J Environ Qual 39:1782–1789. doi:10.2134/jea2009.0464
Lim TT, Edwards DR, Workman SR, Larson BT, Dunn L (1998) Vegetated filter strip removal of cattle manure constituents in overland. Trans ASAE 41(5):1375–1381
Ling TY, Jong HJ, Apun K (2005) Die-off rate of Escherichia coli as a function of pH and temperature. J Phys Sci 16(2):53–63
Ling TY, Bengston RL, Drapcho CM, Achberger EC, Sabbagh GJ, Jackson J (2006a) Modeling of Escherichia coli density on land and concentration in surface runoff. Malays J Soil Sci 10:25–34
Ling TY, Layang HW, Then YP, Apun K (2006b) Impacts of pig farming on the water quality of Serin River. Sarawak Sains Malaysiana 35(1):45–50
Ling TY, Jong HJ, Apun K, Wan Sulaiman WH (2009) Quantifying Escherichia coli release from soil under high-intensity rainfall. Trans ASABE 52(3):785–792
Liu X, Zhang X, Zhang M (2008) Major factors influencing the efficacy of vegetated buffers on sediment trapping: a review and analysis. J Environ Qual 37(5):1667–1674
Lowrance R, Vellidis G, Wauchope RD, Gay P, Bosch DD (1998) Herbicide transport in a managed riparian forest buffer system. Trans ASAE 40:1047–1057
Maciolek JA (1962) Limnological organic analyses by quantitative dichromate oxidation Res. Rept, US Fish Wildl. Service 60. 61 pp
Mankin KR, Barnes PL, Harner JP, Kalita PK, Boyer JE (2006) Field evaluation of vegetative filter effectiveness and runoff quality from unstocked feedlots. J Soil Water Conserv 61(4):209–217
Mankin KR, Wang L, Hutchinson SL, Marchin GL (2007) Escherichia coli sorption to sand and silt loam soil. Trans ASABE 50(4):1159–1165
Marshall VM, Lewis MM, Ostendorf B (2012) Buffel grass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: a review. J Arid Environ 78:1–2. doi:10.1016/J.Jaridenv.2011.11.005
Martinez G, Pachepsky YA, Whelan G, Yakirevich AM, Guber A, Gish TJ (2013) Rainfall-induced fecal indicator organisms transport from manured fields: Model sensitivity analysis. 11/2013; 63C:121–129. doi:10.1016/j.envint.2013.11.003
McLaughlin SJ, Kalita PK, Kuhlenschmidt MS (2013) Fate of Cryptosporidium parvum oocysts within soil, water, and plant environment. J Environ Manag 131:121–128
Miller WA, Lewis DJ, Lennox M, Pereira MDG, Tate KW, Conrad PA, Atwill ER (2007) Climate and on-farm risk factors associated with Giardia duodenalis cysts in storm runoff from California coastal dairies. Appl Environ Microbiol 73(21):6972–6979
Miller WA, Lewis DJ, Pereira MDG, Lennox M, Conrad PA, Tate KW, Atwill ER (2008) Farm factors associated with reducing Cryptosporidium loading in storm unoff from dairies. J Environ Qual 37:1875–1882
Mohanty S, Torkelson A, Dodd H, Nelson K, Boehm A (2013) Engineering solutions to improve the removal of fecal indicator bacteria by bioinfiltration systems during intermittent flow of stormwater. Environ Sci Technol 47(19):10791–10798. doi:10.1021/es305136b
Olilo CO, Shivoga WA, Kitetu JJ, Agembe S (2014) Effects of environmental factors on cyanobacteria dynamics in Lake Baringo, Kenya. Lakes Reserv Res Manag 19(4):240–254
Olilo CO, Onyando JO, Moturi WN, Muia AW, Ombui P, Shivoga WA, Roegner AF (2016) Effects of vegetated filter strips on transport and deposition rates of Escherichia coli in overland flow in eastern escarpments of the Mau Forest, Njoro River Watershed, Kenya. Energy Ecol Environ 2016:1–26. doi:10.1007/s40974-0006-y
Overcash MR, Bingham SC, Westerman PW (1981) Predicting runoff pollutant reduction in buffer zones adjacent to land treatment sites. Trans ASAE 24(2):430–435
Pachepsky YA, Sadeghi AM, Bradford SA, Shelton DR, Guber AK, Dao T (2006) Transport and fate of manure-borne pathogens: modeling perspective. Agric Water Manag 86(1–2):81–92
Randolph TF, Schelling E, Grace D, Nicholson CF, Leroy JL, Cole DC, Demment MW, Omore A, Zinsstag J, Ruel M (2007) Invited review: role of livestock in human nutrition and health for poverty reduction in developing countries. J Anim Sci 85(11):2788–2800
Rippy M (2015) Meeting the criteria: linking biofilter design to fecal indicator bacteria removal. Wiley Interdiscip Rev Water 2(5):577–592. doi:10.1002/wat2.1096
Rochelle-Newall E, Nguyen TM, Le TP, Sengtaheuanghoung O, Ribolzi O (2015) A short review of fecal indicator bacteria in tropical aquatic ecosystems: knowledge gaps and future directions. Front Microbiol 6:308. doi:10.3389/fmicb.2015.00308
Sabbagh GJ, Fox GA, Kamanzi A, Roepke B, Tang JZ (2009) Effectiveness of vegetative filter strips in reducing pesticide loading: quantifying pesticide trapping efficiency. J Environ Qual 38(2):762–771
Schmitt TJ, Dosskey MG, Hoagland KD (1999) Filter strip performance and processes for different vegetation, widths, and contaminants. J Environ Qual 28:1479–1489
Stout WL, Pachepsky YA, Shelton DR, Sadeghi AM, Saporito LS, Sharpley AN (2005) Runoff transport of fecal coliforms and phosphorus released from manure in grass buffer conditions. Lett Appl Microbiol 41(3):230–234
Strickland JDH, Parsons TR (1968) A practical handbook of sea water analysis. Bull Fish Res Bio Can 167:310
Strickland JDH, Parsons TR (1972) A practical handbook of sea water analysis, 2nd edn. Bull Fish Res Bio Can 167:310
Sullivan T, Moore J, Thomas DR (2007) Efficacy of vegetated buffers in preventing transport of fecal coliform bacteria from pasturelands. Environ Manag 40(6):958–965
Sweeney NJ, Klemm P, McCormick BA, Moller-Nielsen E, Utley M, Schembri MA, Laux DC, Cohen PS (1996) The Escherichia coli K-12 gntP gene allows E. coli F-18 to occupy a distinct nutritional niche in the streptomycin-treated mouse large intestine. Infect Immun 64(9):3497–3503
Tate KW, Atwill ER, Bartolome JW, Nader G (2006) Significant Escherichia coli Attenuation by vegetated Buffers on Annual Grasslands. J Environ Qual 35:795–805
Trask JR, Kalita PK, Kuhlenschmidt MS, Smith RD, Funk TL (2004) Overland and near surface transport of Crysptosporidium parvum from vegetated and non-vegetated surfaces. J Environ Qual 33:984–993
Tyrell SF, Quinton JN (2003) Overland flow transport of pathogens from agricultural land receiving faecal wastes. J Appl Microbiol 94:87S–93S
Vanderholm DH, Dickey EC (1979) Design of vegetative filters for feedlot runoff treatment in humid areas. Trans ASAE 23(681–684):687
Walker SE, Mostaghimi S, Dillaha TA, Woeste FE (1990) Modelling animal waste management practices: impacts on bacteria levels in overland from agricultural lands. Trans ASAE 33:807–817
World Health Organization (2011) Guidelines for Drinking Water, 4th edn. World Health Organization, Geneva
Yu J, Yu H, Xu L (2013) Performance evaluation of various storm-water best management practices. Environ Sci Pollut Res Int 20(9):6160–6171
Zar JH (1996) Biostatistical analysis. Prentice Hall, New York
Acknowledgments
Professor Japheth O. Onyando (Chairman, Department of Agricultural Engineering and Technology), Dr. Wilkister N. Moturi (Chairperson, Department of Environmental Science) and Dr. Anastasia W. Muia (Department of Biological Sciences) of Egerton University (EU) designed this study. We want to thank the Dean of Faculty of Agriculture, EU, for making it possible to work in field 18 of Tatton Agriculture Park (TAP). The microbiology laboratory at Kenya Marine and Fisheries Research Institute (KMFRI) made it possible to complete total coliforms and Escherichia coli microbiological analyses. We thank Mr A. Ndegwa of Kenyatta University Herbarium Department for help in identifying local grass species. Mr Godfrey Maina of the Department of Environmental Science, EU, and Mr Chris Nyamweya of KMFRI did the cartographic work. We thank the Department of Water and Civil Engineering of EU for providing the meteorological data from weather station. EU Agricultural Electronic Library (TAEL) staff was instrumental to accessing scientific literature. We thank the Director of KMFRI, Dr. Renison Ruwa, for granting Ph.D. study under the EU-KMFRI Memorandum of Understanding (MOU) Study Programme. The Kenya National Commission of Science and Technology Science, Technology and Innovation PhD research grant funded this study under Grant No. NCST/ST & I/RCD/4th Call PhD/181.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Olilo, C.O., Onyando, J.O., Moturi, W.N. et al. Composition and design of vegetative filter strips instrumental in improving water quality by mass reduction of suspended sediment, nutrients and Escherichia coli in overland flows in eastern escarpment of Mau Forest, Njoro River Watershed, Kenya. Energ. Ecol. Environ. 1, 386–407 (2016). https://doi.org/10.1007/s40974-016-0032-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40974-016-0032-9