
Abstract
The urban effects on macroinvertebrate trophic structure were evaluated in a stream located in central Brazilian Cerrado. Samplings were evaluated from May/2004 to March/2005 in the upper, middle and lower sections. Characterization of ecological conditions and habitat diversity indicated headwater as “natural” site, up and downstream the urban region were “altered” ones, and the sites located nearest to urban area were considered “impacted”. A well oxygenated and transparent water, acid pH, low electrical conductivity (<0.22 µS/cm), and low nutrients concentrations (<0.03 mg/L PO −24 , <0.05 mg/L N–NH4 and <3.0 mg/L N–NO −23 ) was registered for headwater area. Stretches located near the urban region had higher electrical conductivity (48–100 µS/cm), higher ammonium (7.16 mg/L), and poor oxygenated water. The trophic organization was different along the stream and related to environmental preservation degree. Shredders occurred only in the headwater (Phylloicus sp., Trichoptera). Predators were dominant in upstream sites of the urban area (>90 % of biomass). In the sites located near the urban area, an alternative predominance was observed between gathering collectors and predators, with the gathering collectors dominance in the area impacted by city sewage inflow (>90 %). There was a progressive participation of scrapers in stretches located downstream of the city. Biomass was dominated by predators (>50 % in six sites) encompassing larger organisms (>20 mm), and gathering collectors were density dominants (from 53 to 99 %). Predators were the most dominant functional feeding group (22 taxa), followed by scrapers (eight), gathering collectors (seven) and filtering collectors (five). The results state that the Sobradinho stream is under different degrees of environmental impacts with lower preservation levels of its natural conditions, even in areas close to its headwater.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Anthropogenic pressures in urban areas are usually multifactorial and derived by point and non-point sources, and urbanization has often infringed into natural areas, causing alterations in physical, chemical and biological characteristics of aquatic systems (Paul and Meyer 2001).
Streams in agricultural and urban landscapes have been affected by natural and anthropogenic disturbances. These disturbances may fundamentally alter trophic network structure, ecological processes and their integrity, as discussed by Müller et al. (2000).
In areas of dense human population in the urban environment, the watercourses have showed highly degraded water quality, receiving a wide variety of impacts such as domestic and industrial wastewater, sediments and trash. Thus, these rivers tend to lose their natural features and their original biological diversity (Dudgeon 1992).
Their effects can be evaluated and monitored by biotic community analyses. The aquatic macroinvertebrate community is widely used in environmental evaluations (e.g., Rosemberg and Resh 1993). This community represents an intermediary portion in the food chain stream. On the one hand, they are feeding sources for fish; on the other, they feed on algae and bacteria. Thus, due to their intermediary position, macroinvertebrates play an important role in the natural energy flow and recycling of nutrients, especially in small size watercourses that pass through urban regions (Karr 1981).
Also, because of their sessile habits, the ability for colonizing different types of substrates and their relatively long life term, this group of animals is widely used as water quality indicators (Kerans and Karr 1994; Couceiro et al. 2011). As bioindicators, they respond to environmental changes in a variety of ways. These changes may occur in the composition and relative abundance of groups and species, as well as in the trophic measures with emphasis in the feeding functional categories (Resh and Jackson 1993; Barbour et al. 1999).
Although an extensive literature exists on urban effects on macrobenthic organisms (e.g., Cosser 1998; Walsh et al. 2001; Fenoglio et al. 2002; Vasconcelos et al. 2008), little is known about the effects of anthropogenic inputs on the trophic structure of aquatic communities in Brazilian urban areas, especially in central region, where these impacts have yet been poorly documented (Marsalek et al. 2006; Fonseca et al. 2014).
Functional classifications based upon feeding can permit the elaboration of trophic dynamics in water courses shaped as guild or functional feeding groups (FFGs) (Grubbs et al. 1995; Cummins et al. 2005). In the Brazilian lotic ecosystems, this classification has been widely used for measuring resources availability and their use by aquatic macroinvertebrates (Couceiro et al. 2011).
Macroinvertebrate trophic interactions can have a strong effect on ecological processes by directly influencing flows of energy and materials (Wallace and Webster 1996). FFGs therefore allow inferences on the functioning and productivity of aquatic ecosystems, and can be used to investigate the effects of anthropogenic impacts on these processes.
Merritt et al. (2008) also suggested that proportions of the different FFGs can be used as indicators of stream ecosystem attributes, instead of measurements that require large investments in time and equipment and provide only a ‘snapshot’ view of ecosystem function. The ratios between FFGs can indicate the relative importance of stream ecosystem features, such as trophic state (autotrophy or heterotrophy), the relative amounts of coarse and fine particulate organic matter being transported and stored in the environment, and the stability of the channel, without the necessity of measuring these variables.
Biomass and secondary production have been used to address many ecological questions concerning the construction of quantitative food webs and to assess the effects of pollution and catchment land use change (Benke 2010).
The goal of the present study was to characterize aquatic macroinvertebrates using density and biomass spatial distribution of the feeding functional groups and correlating these aspects with the visual qualification of the ecological conditions preservation and with the habitat diversity. This study was conducted in a peri-urban stream including headwater, urban and agriculture areas, which allowed to compare the main impacts acting on the stream and, consequently, on the aquatic macroinvertebrates.
Materials and methods
Study area
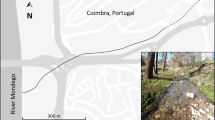
The Sobradinho stream is located at the central northeastern portion of the Federal District, between parallels 15°36′25″ and 15°45′35″ and the meridians 47°53′00″ and 47°3′08″. This stream belongs to São Bartolomeu River watershed, whose main river cuts the Federal District from north to southward (Fig. 1).

Localization of São Bartolomeu Basin and the Sobradinho stream sub-basin with the eight sites (Sobr1 to S8) along the Sobradinho stream, central Brazil. Schematic representation
The regional climate is Aw (rainy tropical, according to Köppen classification), characterized by two seasons: the cool-dry (May–September) and the warm-rainy (October–April). Sobradinho sub-basin is located at an altitude of 1000–1200 m, and air temperature ranges between 18 and 22 °C. The annual mean precipitation is around 1500 mm, ranging from 750 to 2000 mm (Silva et al. 2008).
The Sobradinho stream is 28 km long and its drainage basin comprises 153 km2. Localities close to the stream headwater are 0.50 m deep and 0.60 m wide, while near the mouth of the São Bartolomeu River, the stream reaches up to 2.0 m depth and about 9.0 m width.
The upper part of Sobradinho stream is under the influence of domestic effluents due to a huge urban area. Near its mouth in the São Bartolomeu River, it is under rural cover, even though agriculture is considered a secondary activity in the area.
Sampling and procedures
Eight sites were established (SOBR01, SOBR02, SOBR03, SOBR04, SOBR05, SOBR06, SOBR07 and SOBR08), distributed from the Sobradinho stream headwater until its mouth in the São Bartolomeu River (Fig. 1). The areas surrounding the headwater, pollution sources, nearby urban centers and rural areas were considered. One site (SOBR01) was located in a protected area, although surrounded by subsistence agriculture and Eucalyptus plantation. Three sites (S0BR2, SOBR7 and SOBR8) were situated within a land rural cover. The other four sites (SOBR3, SOBR4, SOBR5 and SOBR6) were situated near urban areas and received effluents from stabilization ponds. Abiotic parameters and aquatic macroinvertebrates sampling (three for each stretch) were conducted for 8 months, during rainy (December/2004–March/2005) and dry (May–August/2005) seasons.
The sampling stations were classified using an adaptation of the rapid assessment protocol proposed by Barbour et al. (1999). Several habitat attributes were considered (e.g., water and sediment characteristics, land use, embankments, riparian vegetation zone width, size and frequency of riffles and pools), which are assigned numerical scores along a gradient of optimal to poor based on visual inspection. The score was determined by adding partial values given to each individual parameter. The scores indicate the degree of preservation of the sampling stations as follows: 0–40 points, “impacted”; 41–60 points, “altered”; over 61, “natural”.
Surface water samples were taken from the middle of the stream channel using polyethylene bottles in a cooled container to move to the laboratory for analyses of pH (pHmeter, Hack-sension 156), water electrical conductivity (conductivimeter, µS/cm, Hack-sension 156), turbidity (FTU, Hack-sension 156), dissolved oxygen (mg/L, Winkler method modified by Golterman et al. 1978) and total dissolved solids (TDS, mg/L, Hack-sension 156). Samples were taken for inorganic nutrients analyses of orthophosphate (PO −24 , mg/L), ammonium (N–NH4, mg/L) and nitrate (N–NO −23 , mg/L) according to standard methods recommended by APHA (2005).
For collecting aquatic macroinvertebrates, a ‘D’-net sampler (250 μm mesh, 30 cm width) was used. An area of 0.465 m2 (1.50 m × 0.31 m) was sampled in triplicate on the bottom of the stream for a distance of 1 m in the direction of the stream bank, until it reached the edge of the water at the bank (see Couceiro et al. 2011). Individual samples were preserved in labeled plastic bags containing 90 % alcohol until analysis. In the laboratory, macroinvertebrates were separated, identified and counted under a stereo-microscope. The specimens were identified using taxonomic keys (e.g., Pérez 1988; Pennak 1989; Merritt et al. 2008).
Macroinvertebrates were classified into five FFGs: gathering collectors, filtering collectors, shredders, predators and scrapers (Table 2), following the classification used by Cummins et al. (2005) and Merritt et al. (2008).
For each category, individuals were separated according to class sizes: class 1 (smaller than 10 mm); class 2 (between 10 and 15 mm); class 3 (between 15 and 20 mm); and class 4 (bigger than 20 mm).
For dry biomass determination, the invertebrates were placed in preweighed aluminum foils according to their functional group and size, overdried at 60 °C for 48 h, cooled in a desiccator and weighed again (precision 0.01 mg) in a Mettler H54 balance.
Descriptive analysis was applied for water variables using two different approaches. Mean and standard deviation were used as measures of central tendency and variation, respectively (Table 3). After classification based on the rapid habitat assessment protocol (Table 1), data from each category were considered within three different groups (“impacted”, “altered” and “natural”). The variables were described by their median, mean, minimum and maximum values (Table 3).
To verify differences among the sampling sites, Kruskal–Wallis test, with the multiple comparison Dunn test a posteriori, was used to verify whether or not data from the three groups resulting from the protocol application were significantly different for each water variable. Results were considered significant when p < 0.05.
To evaluate aquatic macroinvertebrate community structure, the density (individuals/m2) and biomass (g/m2) of the feeding functional groups were estimated. Results obtained from the analysis of these community features were submitted to Kruskal–Wallis test, with the multiple comparison Dunn test a posteriori, to verify the differences for each biological variable, according to the three groups resulting from the protocol application. Statistical analyses were performed using the program SPSS 12.0 for Windows (SPSS Inc., Illinois, USA).
Results
According to classification based on the rapid habitat assessment, differences in the preservation levels of ecological conditions along the Sobradinho stream were observed. Sites were grouped into three types: 1, natural; 2, altered; and 3, impacted. The main parameters of the rapid protocol that affected the results were the presence/absence of gallery forest, the land use in the surroundings and the diversity of habitats and microhabitats (Table 1).
SOBR01 site is located close to the stream headwater and was considered natural according to the protocol. Stretches near to the urban center were classified as impacted sites (SOBR03, SOBR04, SOBR05 and SOBR06). Altered stretches were obtained for the up (SOBR02) and downstream (SOBR07 and SOBR08) the urban region.
A well-oxygenated and transparent water with acidic pH, low electrical conductivity (>0.22 uS/cm) and low nutrient concentrations (<0.03 mg/L PO −24 , <0.05 mg/L N–NH4 and <3.0 mg/L N–NO −23 ) and alkalinity (11.95 mg/L CaCO3) was registered for the headwater area (classified as natural site by the protocol). Also, in this area significantly low values were obtained for turbidity, TDS and water electrical conductivity (Kruskal–Wallis/Dunn test; p < 0.05).
On the contrary, stretches located near urban regions and considered impacted had higher values in SOBR05 for orthophosphate (0.22 mg/L), water electrical conductivity (100 µS/cm) and TDS (48.6 mg/L). Significantly high values in impacted sites were obtained for ammonium and alkalinity, and also these sites had poorly oxygenated water (Kruskal–Wallis/Dunn test; p < 0.05). Nitrate was at a significantly higher value in the altered sites (Kruskal–Wallis/Dunn test; p < 0.05).
Box plots were constructed for some abiotic variables to show their variation among natural, altered and impacted sites, according to protocol (Fig. 2).

Box plots of values of dissolved oxygen (a), water electrical conductivity (b), alkalinity (c), nitrate (d), TDS (e), turbidity (f) and ammonium (g) representing the variation according to protocol, as natural (1), altered (2) and impacted (3) stretches. Central, superior and inferior lines show, respectively, median, 75 and 25 %, and vertical lines show max and min values
We recorded a total of 29,637 individuals distributed into 44 taxa in the 24 collections carried out over the study period. From these, 36 taxa belonged to Insecta, four to Mollusca, two to Annelida and only one taxon to Platyhelminthes and to Arachnida.
The fauna structure followed the general organization pattern of macroinvertebrate assemblages, with the occurrence of many rare taxa and a few abundant taxa. The headwater station obtained the highest species richness (34 taxa) and SOBR5 (the highest impact station) obtained only six taxa during this study. The richness of species was significantly higher (Kruskal–Wallis/Dunn test; p < 0.05) in the natural site, and sites located in the urban cover registered lower significant richness (Kruskal–Wallis/Dunn test; p < 0.05).
Predators were the most dominant feeding functional groups (FFG; 22 taxa), followed by scrapers (eight taxa), gathering collectors (seven taxa) and filtering collectors (five taxa); there was only one taxon for the shredders (Phylloicus sp., Trichoptera) (Table 2).
In the stretch most impacted (SOBR05), aquatic macroinvertebrates reached the highest density and biomass, mostly belonging to Chironomidae (Figs. 3, 4). This site was located downstream of the sewage treatment of the city. The value obtained by the protocol (18 points) was the lowest within the impacted stretches (30, 32 and 39 points) (Table 1).

Relative percentage (%) for a density and b biomass of the aquatic macroinvertebrate feeding functional groups at the sites in the Sobradinho stream, central Brazil. Rainy season (Dec/2004–Mar/2005) and dry season (May–Aug/2005)

Mean and SD for a density and b biomass of aquatic macroinvertebrates at the sites in the Sobradinho stream, central Brazil, with rainy season Dec/2004–Mar/2005 and dry season May–Aug/2005
Insects were predominant in all classes of functional groups analyzed in this study. At the SOBR01, considered a natural site, there were numerous gathering collectors and predators (Fig. 5a). On the other hand, predators were dominant in terms of biomass (Fig. 5b). Only at this site, there were organisms found belonging to shredders, but with low density. In the altered and impacted sites, gathering collectors were more numerous (>50 %) along the study period (Fig. 4a).

Biomass of aquatic macroinvertebrate feeding functional groups for each size class at the sites of the Sobradinho stream. (Class 1: <10 mm, Class 2: from 10 to 15 mm, Class 3: from 15 to 20 mm and class 4: >20 mm). (Pr predators, gath-col gathering collector, filt-col filtering collector, Scrap scrapers, shred shredders). Rainy season is Dec/2004–Mar/2005 and dry season May–Aug/2005)
Biomass measurements of feeding functional groups were significantly different among the three categories according to the protocol (Kruskal–Wallis/Dunn test; p < 0.05). Nevertheless, predators were dominant in most of the sites (SOBR01, 02, 04 and 06, >80 %) and were abundant in the sites SOBR03 and 08, scattering into the natural, altered and impacted stretches (Fig. 3).
The shredders group only showed up at the station classified as natural (SOBR01). Gathering collectors were dominant only at the site SOBR05, and representative at the SOBR03 and 07 sites (>20 %), including the areas altered and impacted. At the altered stretches (SOBR07 and 08), the scrappers were representative with 34 and 28 % of the total biomass, respectively (Fig. 5b).
In relation to biomass in size classes within the FFG, significant differences were found concerning gathering collectors, filtering collectors and scrapers among the three categories according to the protocol (Kruskal–Wallis/Dunn test; p < 0.05). In the altered sites, the collectors and scrapers with sizes smaller than 10 mm were dominant, while in the impacted sites the collectors and scrapers were of sizes below 15 mm. Predator biomass was represented by larger organisms in all sites (>20 mm). Shredders found at the land parcel labeled as natural had sizes bigger than 10 mm(Figure 4).
In summary, the trophic organization was different along the stream and related to the environmental preservation degree. Shredders occurred only in the headwater, which was classified as a natural site (Phylloicus sp., Trichoptera). Predators were dominant in upstream sites of the urban area (>90 % of biomass). In the sites located near urban areas, an alternative predominance was observed between gathering collectors and predators, with the gathering collectors dominant in areas impacted by city sewage inflow (>90 %). There was a progressive participation of scrapers in altered stretches located downstream of the city.
Discussion
The trophic organization of the macrobenthic community was different along the stream and can be explained, in part, by their habitat requirements and also related to environmental preservation degree. The impact effect was evident from a comparison of upstream–downstream stretches and the ones near the urban region.
In protected areas of central Brazil, streams are characterized by low conductivity, pH and high values of dissolved oxygen, as seen in the headwater area of this study. But, as a peri-urban stream, its sub-basin is subjected to a continuous process of urbanization, and the anthropogenic pressure was evidenced by biological and environmental changes observed by this study, as discussed by Fonseca et al. (2014).
The rapid assessment protocol was considered an adequate tool to relate the aquatic community aspects with the degree of ecological conditions and the habitat diversity. This protocol permitted to establish the differences in trophic organization of the macrobenthic assemblages to be attributed to features of the stream within a human impact degree.
Shredders were registered in the headwater area, which is a typical taxon of running water (Wiggins 1996) and more abundant in leaf litter in riffle (Pes et al. 2005). In this study, the headwater area was considered a natural environment, with no direct and evident anthropic influence.
Streams impacted by anthropogenic sediments can result in the reduction of shredders’ density, as shown by Couceiro et al. (2011) in Amazonian streams of petroleum areas. This fact could have had a large effect on ecosystem function, since the reduction of the density of organisms in some feeding groups, such as shredders, can change the functioning of aquatic ecosystems, causing a trophic cascade effect (e.g., Chapin-III et al. 2000). Baer et al. (2001), for example, found that secondary production in streams in North Carolina (USA) was reduced by more than 70 % after the exclusion of litter, and, consequently a variety of changes occurred in the colonization dynamics of macroinvertebrates and in detritus processing.
The sites considered “altered” by the protocol had since only a moderate erosion of the margins (SOBR02), to evident anthropic alterations as observed in downstream sites of the urban region (SOBR07 and SOBR08), and, especially, related to organic pollution derived by sewage inflow. Despite of this, SOBR02 showed a high preservation level in response to availability of higher organic resource sources and bed stability of stream. Thus, these features permit to diversify habitats and substrate to be colonized by the aquatic macroinvertebrates.
Forest clearance, silting, cattle raising and pasture formation and especially also urban sewage inflow favored the homogenization of the bed of water course, reducing habitat and microhabitat diversity and thus affecting the biotic compartment of the stream (Dobson 1994; Aguiar et al. 2002; Boyero and Bosh 2004).
The areas with the lowest scores in the characterization protocol also obtained low richness of benthic organisms and with a dominance of tolerant species to urban impacts. This situation is relevant for SOBR05, which receives sewage inflow from Sobradinho City. The physical and chemical conditions were quite different among the three environmental protocol categories. This situation suggests that the human impacts had a local interference on the water quality along the Sobradinho stream sub-basin.
Changes in the structure and composition of the aquatic macroinvertebrate community are due to changes in the trophic resource availability (Barbour et al. 1999). Along the Sobradinho stream basin, the human impacts prevailed due to urban and agriculture cover and these impacts can be related to changes in the biota. Changes in the macrobenthic assemblages along the stream can indicate the ability of groups and species to occupy different microhabitats and process organic material, as indicated by Buss et al. (2002) and Nedeau et al. (2003).
Gathering collectors were the FFG group with the greatest representation in terms of density in this study, whether natural, altered or impacted sites. Also, they were dominant in terms of biomass in the most impacted site (SOBR05). This group is considered rapid to be colonizers and also species tolerant to human impacts. Buss et al. (2002) also found gathering collectors in the streams located in the Atlantic forest (Brazil) independent of their water quality.
In areas submitted to urban activities, stream stretches were considered by protocol to be impacted. The type and degree of human impact can affect the environmental conditions and the biological diversity of the stream. For this, the use of community attributes, as FFG, should provide a better differentiation between degraded and reference stream reaches than taxonomic similarity, as postulated by Karr (1981, 1987) for benthic communities.
In polluted streams in southeastern Brazil, the prevalence of the dipterans chironomids, known as indicators of environmental stress, was reported (Marques et al. 1999). Chironomids are tolerant organisms and chiefly gathering collectors (Paul and Mayer 2001); their abundance may indicate degraded conditions (Gonzalez et al. 2003). These organisms are better adapted to extreme environmental conditions related with the contents of organic matter in the sediment, low dissolved oxygen concentration (Cranston 1995) and ammonium (Abílio et al. 2005). Our results confirm this tendency because Chironomid was dominant in stretch with high impact level (SOBR05), due to organic inflow from punctual pollution sources by the urban area, especially from a sewage treatment station.
The agriculture activities were predominant in stretches considered altered by the protocol. In these sites, there were alterations observed in the type of sediment, stream backwater areas, sediment deposition and riparian vegetation. These alterations were also observed in impacted stretches, with direct effect on the surrounding ecological conditions.
As view for FFG, the impact mainly derived to organic pollution favored the increase of the percentage for collector groups and, in contrast, diminished the contribution of predators and shredders (Kerans and Karr 1994). Specialized organisms such as shredders are more sensitive and can be found in greater densities in headwaters environments. Generalist organisms such as the collectors have wider range of feeding resource acceptability than the specialists; hence they are more tolerant to pollution since they are capable of modifying the availability of certain feeding sources (Cummins and Klug 1979). A similar response has been observed in other studies (Ortiz et al. 2005; Roy et al. 2003).
Based on the FFG community concept (Cummins et al. 2005; Merritt et al. 2008), this stream tended to be heterotrophic and had little association between shredders and riparian vegetation. Some habitats (e.g., leaves) can provide shelter (protection), favoring the occurrence of some macroinvertebrate groups, while other habitats (e.g., sand and silt) can impede the establishment of groups that need a firm substrate for fixation (Couceiro et al. 2010).
A lower density of predator in the sites subjected to impacts is expected. Nevertheless, the SOBR04 site presented an intermediate impact level, and the presence of the order Odonata as a dominant group of predator organisms may reflect this condition. Ferreira-Peruquetti and Fonseca-Gessner (2003) detected that the Odonata richness in areas altered by the riparian vegetation removal may be greatly augmented. However, important species loss for the system and the possibility of exotic species invasions into the altered system may also occur.
Despite the study of aquatic macroinvertebrate biomass, the differences between portions of the stream did not follow the same pattern observed for the density values. An example of the differences that can be obtained using biomass vs. number can be observed with chironomids larvae (Diptera: Chironomidade). Numerically, more than 50 % of macroinvertebrates in a sample are documented as chironomids, but their total biomass is abnormally inferior to 20 % (Arimoro et al. 2007).
At the downstream stretches near urban areas, predator biomass was more representative, followed by the scrapers. The contribution of this group was not noticeable when analyzed in terms of density of feeding functional groups. Decrease in the density of feeding groups may be caused by the presence of predators with sizes bigger than the primary consumers. In the case of chironomids (consumers), it may mean that, in spite of presenting higher biomass rotation and being constant in most habitats (Benke 2010), they are also more susceptible to predation (Wallace and Webster 1996), probably due to the smaller size (<10 mm in our study) and low mobility (Roque et al. 2004).
Other studies have been done in the Federal District to evaluate the fauna responses to organic residual discharge impact and urbanization (Martins-Silva and Barros 2001 and references). However, this work also aimed at studying the biomass density of aquatic macroinvertebrates at the feeding functional group level and its relationship with the characteristics of habitats. The study must be considered, above all, as an effort in attempting to understand the fauna responses to anthropogenic impacts.
Estimation of the diversity of habitats is an important tool in the aquatic environment quality approach due to the strong correlation between habitat availability and aquatic diversity (Galdean et al. 2000; Padovesi-Fonseca and Martins-Silva 2012). The use of the rapid assessment protocol along the stream was beneficial in this study. This analysis was also done for some lotic systems of the Federal District (Padovesi-Fonseca et al. 2010; Fonseca et al. 2014). Evaluation of a peri-urban stream, such as Sobradinho, was an important parameter for an environmental diagnosis of the studied region. By doing so, we were able to state that the Sobradinho stream is under different degrees of environmental impacts with lower preservation levels of its natural conditions, even in areas close to its headwater.
References
Abílio FJP, Fonseca-Gessner AF, Watanabe T, Lunginho LR (2005) Chironomidae and other aquatic insects of dams in semi-arid of Paraíba, Brazil. Entomol Vectores 12:255–264
Aguiar FC, Ferreira MT, Pinto P (2002) Relative influence of environmental variables on macroinvertebrate assemblages from in Iberian basin. J North Am Benthol Soc 21(10):43–53
APHA (2005) Standard methods for the examination of water and waste water, 21st edn. American Public Health Association, Washington
Arimoro FO, Ikomi RB, Iwegbue CMA (2007) Water quality changes in relation to dipteral community patterns and diversity measured at en inorganic effluent impacted stream in the Niger Delta, Nigeria. Ecol Indic 7:541–552
Baer SG, Siler ER, Eggert SL, Wallace B (2001) Colonization and production of macroinvertebrates on artificial substrata: upstream–downstream responses to a leaf litter exclusion manipulation. Freshwater Biol 46:347–365
Barbour MT, Gerritsen G, Snyder BD, Stribling JB (1999) Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish, 2nd edn. EPA 841-B-99-002. US Environmental Protection Agency; Office of Water, Washington
Benke AC (2010) Secondary production as part of bioenergetic theory—contributions from freshwater benthic science. River Res Appl 26:36–44
Boyero L, Bosch J (2004) The effect of riffle-scale environmental variability on macroinvertebrate assemblages in a tropical stream. Hydrobiologia 524:125–132
Buss DF, Baptista DF, Silveira MP, Nessimian JL, Dorvillé FLM (2002) Influence of water chemistry and environmental degradation on macroinvertebrate assemblages in a river basin in south-east Brazil. Hydrobiology 481:125–136
Chapin-III FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU, Lavorel S, Sala OE, Hobbie SE, Mack MC, Diaz S (2000) Consequences of changing biodiversity. Nature 405:234–242
Cosser PR (1998) Macroinvertebrate community structure and chemistry of an organically polluted Creek in South-east Queensland. Aust J Mar Freshw Res 39:671–683
Couceiro SRM, Hamada N, Forsberg BR, Padovesi-Fonseca C (2010) Effects of anthropogenic silt on aquatic macroinvertebrates and abiotic variables in streams in the Brazilian Amazon. J Soils Sediments 10(1):89–103. doi:10.1007/s11368-009-0148-z
Couceiro SRM, Hamada N, Forsberg BR, Padovesi-Fonseca C (2011) Trophic structure of macroinvertebrates in Amazonian streams impacted by anthropogenic siltation. Austral Ecol 36:628–637
Cranston PS (1995) Medical significance. In: Armitage PD, Cranston PS, Pinder LCV (eds) The Chironomidae: biology and ecology of non-biting midges. Chapman & Hall, London
Cummins KW, Klug MJ (1979) Feeding ecology of stream invertebrates. Annu Rev Ecol Evol Syst 10:147–172
Cummins KW, Merritt RW, Andrade PCN (2005) The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Stud Neotrop Fauna Environ 40:69–89
da Silva FAM, Assad ED, Evangelista BA (2008) Caracterização climática do bioma Cerrado. In: Sano SM, Almeida SMP, Ribeiro JF (eds) Cerrado: Ecologia e Flora. Embrapa Informação Tecnológica, Brasília, pp 69–87 (in Portuguese)
Dobson M (1994) Microhabitat as a determinant of diversity: stream invertebrates colonizing leaf packs. Freshwater Biol 32:565–572
Dudgeon D (1992) Endangered ecosystems: a review of the conservation status of tropical Asian rivers. Hydrobiologia 248:167–191
Fenoglio S, Bandino G, Bona F (2002) Benthic macroinvertebrate communities as indicators of river environment quality: an experience in Nicaragua. Rev Biol Trop 50:125–1131
Ferreira-Peruquetti PS, Fonseca-Gessner AA (2003) Odonata (Insecta) in Cerrado and monoculture areas in the northeastern region of the São Paulo State, Brazil: relationship between land use and the fauna richness (in Portuguese). Rev Bras Zool 20:219–224
Fonseca BM, Mendonça-Galvão L, Padovesi-Fonseca C, Abreu LM, Fernandes ACM (2014) Nutrient baselines of Cerrado low-order streams: comparing natural and impacted sites in Central Brazil. Environ Monit Assess 186:19–36
Galdean N, Callisto M, Barbosa F (2000) Lotic ecosystems of Serra do Cipó, southeast Brazil: water quality and a tentative classification based on the benthic macroinvertebrate community. Aquat Ecosys Health Manage 3:545–552
Golterman HL, Clymo RS, Ohnstad MAM (1978) Methods for physical and chemical analysis of freshwaters. Blackwell Scientific Publications, Oxford, p 213 (v.I.B.P. Handbook. 8)
Gonzalez JM, Basaguren A, Pozo J (2003) Macroinvertebrate communities along a third-order Iberian stream. Ann Limnol-Int J Lim 39:287–296
Grubbs SA, Jacobsen RE, Cummins KW (1995) Colonization by Chironomidae (Insecta, Diptera) on three distinct leaf substrates in an Appalachian mountain stream. Ann Limnol-Int J Lim 31:105–118
Karr JR (1981) Assessment of biotic integrity using fish communities. Fisheries 6:21–27
Karr JR (1987) Biological monitoring and environmental assessment: a conceptual framework. Environ Manage 11:249–256
Kerans BL, Karr JR (1994) A benthic index of biotic integrity (B-IBI) for rivers of the Tennessee Valley. Ecol Appl 4:768–785
Marques GSM, Ferreira RL, Barbosa FAR (1999) The aquatic macroinvertebrate community and limnological features of Carioca and da Barra lakes, Parque Estadual do Rio Doce, MG. Rev Biol Trop 59:203–210
Marsalek J, Jiménez-Cisneros BE, Malmquist P-A, Karamouz M, Goldenfum J, Chocat B (2006) Urban water cycle processes and interactions. IHP-VI Technical Documents in Hydrology, vol 78. UNESCO, Paris
Martins-Silva MJ, Barros M (2001) Occurrence and distribution of fresh-water molluscs in the Riacho Fundo Creek Basin, Brasília, Brazil. Rev Biol Trop 49:791–796
Merritt RW, Cummins KW, Berg MB (2008) An introduction to the aquatic insects of North America, 4th edn. Kendall/Hunt Publishing Company, Dubuque
Müller F, Hoffmann-Kroll R, Wiggering H (2000) Indicating ecosystem integrity—theoretical concepts and environmental requirements. Ecol Model 130:13–23
Nedeau EJ, Merritt RW, Kaufman MG (2003) The effect of an industrial effluent on an urban stream benthic community: water quality vs. habitat quality. Environ Pollut 123:1–13
Ortiz JD, Martí ME, Puig A (2005) Recovery of the macroinvertebrate community below a wastewater treatment plant input in a Mediterranean stream. Hydrobiologia 545:289–302
Padovesi-Fonseca C, Martins-Silva MJ (2012) Biologic integrity analysis as protection tool of Cerrado’s pristine areas. In: Bilibio C, Hensel O, Selbach JF (Orgs.) Sustainable water management in tropics and subtropics and case studies in Brazil, vol. 3. pp 685–721. UNIPAMPA/UNIKASSELL/PGCULT/UFMA, Jaguarão. http://books.google.com.br/books/about/Sustainable_water_management_in_the_trop.html
Padovesi-Fonseca C, Corrêa ACG, Leite GFM, Joveli JC, Costa LS, Pereira ST (2010) Diagnostic of ribeirão Mestre d’Armas sub-basin using two methods of rapid environmental assessment, Federal District, Central Brazil. Ambi-Agua 5:43–56 (in Portuguese)
Paul MJ, Mayer JL (2001) Stream in the urban landscape. Annu Rev Ecol Syst 32:333–365
Pennak RW (1989) Fresh-Water Invertebrates of the United States (Protozoa to Mollusca), 3ª edn. Wiley, New York, p 628
Pérez GR (1988) Guide to the study of aquatic macroinvertebrates of the Departamento de Antioquia. Colciencias, Universidade de Antioquia. Fondo Fen Colombia, Editorial Presencia Ltda. p 217
Pes AMO, Hamada N, Nessimian JL (2005) Identification keys of larvae for families and genera of Trichoptera (Insecta) in central Amazônia, Brazil. Rev Bras Entomol 49:181–204
Resh VH, Jackson JK (1993) Rapid assessment approaches in benthic macroinvertebrate biomonitoring studies. In: Resh VH, Rosenberg DM (eds) Freshwater biomonitoring and benthic macroinvertebrates. Chapman and Hall, New York
Roque FO, Trivinho-Strixino S, Jancso M, Fragoso EM (2004) Records of Chironomidae Larvae living on other aquatic animals in Brazil. Biota neotrop 4:1–9
Rosemberg DM, Resh VH (1993) Freshwater biomonitoring and benthic macroinvertebrates. Chapman and Hall, New York
Roy AH, Rosemond AD, Paul MJ, Leigh DS, Wallace JB (2003) Stream macroinvertebrate response to catchment urbanization (Georgia, USA). Freshw Biol 48:329–346
Vasconcelos MC, Melo AS (2008) An experimental test of the effects of inorganic sediment addition on benthic macroinvertebrates of a subtropical stream. Hydrobiologia 610:321–329
Wallace BJ, Webster JR (1996) The role of macroinvertebrates in stream ecosystem function. Annu Rev Entomol 41:115–139
Walsh CJ, Sharpe AK, Breen PF, Sonneman JA (2001) Effects of urbanization on streams of the Melbourne region, Victoria, Australia. I. Benthic Macroinvertebrate Communities. Freshw Biol 46:535–551
Wiggins GB (1996) Larvae of the North American Caddisfly Genera (Trichoptera), 2nd edn. University of Toronto Press, Toronto, p 401
Acknowledgments
This study is a result of the research program on aquatic biodiversity carried out by Nucleus of Limnological Studies (NEL). The authors are grateful to CAPES and CNPq for their scholarship to the first author and to the colleagues from the Laboratory of Limnology for help during field study. The present study was supported by Post-Graduate Program in Ecology/University of Brasilia, Brazil.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Moreyra, A.K., Padovesi-Fonseca, C. Environmental effects and urban impacts on aquatic macroinvertebrates in a stream of central Brazilian Cerrado. Sustain. Water Resour. Manag. 1, 125–136 (2015). https://doi.org/10.1007/s40899-015-0013-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40899-015-0013-8