
Abstract
The species-general life history (LH) principle posits that extrinsic morbidity-mortality risks accelerate organisms’ pace of life and promotes fast LH-related traits (e.g., earlier sexual maturation and reproduction). Humans, however, have experienced uniformly decelerated LH due to their evolved abilities and efforts in controlling environmental contingencies. The present study proposes that the reason for the existence of such human-specific deceleration of LH may be rooted in human parenting, hence aims to explore the roles of parenting within the relationships between environmental adversities and children’ LH calibration. The present study used data from the NIMH Data Archive (NDA) reported by 2014 children and their caregivers, and examined the effects of parental investment in calibrating children’s LH behavioral manifestations, as well as in moderating the impacts of environmental harshness and unpredictability on children’ development of LH. The findings showed that parental investment negatively predicts the development of fast LH behavioral profiles in children and moderates the impacts of environmental adversities directly imposed on children. We conclude that human parental investment serves as an important pivot in down-regulating environmental effects on children’s LH calibration, providing an alternative insight into the species-specific deceleration of LH.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Life history (LH) theory is an evolutionary biological framework that explains between- and within-species variation in the allocation of bioenergetic resources among components of fitness (e.g., growth, survival, reproduction; Del Giudice et al., 2015; Stearns, 1989, 1992). When allocating resources to fitness components, organisms are essentially subject to making tradeoffs, much owing to the biological reality that they have only a limited energy “budget” that cannot simultaneously maximize all the components (Del Giudice et al., 2015; Hill & Kaplan, 1999). One of the fundamental LH tradeoffs lies between parenting and mating effort, i.e., investing in existing offspring versus reproducing additional offspring. Harsh and unpredictable environmental conditions resulting in high and variable extrinsic morbidity-mortality rates (which are insensitive to individuals’ resource-allocation decisions) favor mating strategies that facilitate phenotypic diversity to brace environmental hazards (Ellis et al., 2009; Quinlan, 2007). Conversely, when environmental conditions are benign and stable, parenting strategies are selected for since they aid in increasing existing offspring’s survival and reproductive success. Parenting effort serves to channel the intergenerational calibration of LH. When the rates of extrinsic morbidity-mortality are low and stable, or are sensitive to parents’ resource-allocation decisions, increased parental investment should emerge to decelerate children’s LH (Ellis et al., 2009). Such intergenerational deceleration of LH can be construed as twofold. On the one hand, in addition to feeding and nurturing, parental care also serves as a “protecting belt” that reduces the impacts of environmental adversities directly imposed on children (Bowlby, 1982; Chisholm, 1993). On the other hand, increased parental investment makes room for children’s extended (brain and body) growth and fosters social learning, equipping children with higher competitiveness in terms of resource-capturing and reproduction (Chisholm, 1993; Geary, 2000; Geary & Flinn, 2001).
In contrast to our ancestors, modern humans have experienced slower paces of life owing to advances in medical care, sanitation, and food nutrition, representing evolved increased efforts in attenuating the rates of extrinsic morbidity-mortality (Hill, 1993; Lu et al., 2022b). Such evolved efforts may also manifest in human parenting. Human parenting varies greatly in form, encompassing almost exclusive maternal care, bi-parental cooperation, and alloparenting (i.e., care provided by relatives in addition to biological parents; Sear, 2016), as well as in degree, ranging from a more rejecting to a more responsive style (Rohner, 2004). The LH literature has established that parenting may serve a unique role in shaping children’s LH makeups. For instance, Draper and Harpending (1982) revealed that absence of father is associated with increased interest in competitive dominance in boys, as well as earlier sexual debut and less restrictive, discriminant sexuality in girls. Furthermore, this work was Belsky et al.'s (1991; for a review see also Belsky, 2019) psychosocial acceleration theory that identifies harsh, rejecting, neglectful, and inconsistent parenting as proxies of external adversity that accelerate children’s LH, which has been supported by later evidence (e.g., Belsky et al., 2007; Dunkel et al., 2015; Warren & Barnett, 2020). Putting the theorizing and observations together, the present research proposes that the calibration of children’s LH by parenting effort can be formulated in two pathways. First, harsh, rejecting, or neglectful parenting may shape children’s pace of LH in the same direction as early environmental harshness and unpredictability (i.e., accelerating children’s LH). Second, if parenting is responsive and/or supportive, it then may work in the opposite direction against early adversities (i.e., decelerating children’s pace of LH) and operate to alleviate direct environmental effects.
Environmental Contingencies and Evolution of LH Strategies
The principle of LH theory is that LH strategy is only considered as optimal if it involves a combination of resource-allocation decisions that maximizes individuals’ inclusive fitness under their living environmental conditions (Del Giudice et al., 2015; West & Gardner, 2013). To maximize fitness, resource allocation decisions must be coordinated in the same direction, rendering two types of LH strategies. One is to allocate resources to growth in the brain and physical body and development in terms of social and cognitive functioning, somatic maintenance such as metabolism, immune defense against pathogens, and cellular/DNA repairing, as well as parental investment in existing children’s growth and development. The other is to allocate resources to reproductive effort, such as engaging in early and frequent mating activities, delivering more children, yet investing less in existing children’s quality (Ellis et al., 2009; Geary, 2002). The two LH strategies have been understood as varying on a fast-slow continuum (Promislow & Harvey, 1990), with the former characterizing slow paces of life since it results in delayed reproduction, and the latter relatively fast paces of life since it promotes instant reproductive payoffs.
Organisms’ LH is largely contingent on environmental risks (e.g., predation, infectious pathogens, intraspecific violence) that impose threats to organisms’ survival and reproduction independent of individuals’ effort in resource allocation. The extent of environmental effects is indexed by the rate (environmental harshness) and the variance of the rate (environmental unpredictability) at which ecological conditions result in age-specific morbidity-mortality in a population (Chang & Lu, 2016; Ellis et al., 2009). In general, harsh and unpredictable environmental conditions would favor accelerated (fast) LH strategies such as early maturation and reproduction that spurs the rate of phenotypic iterations over that of extrinsic morbidity-mortality. In contrast, benign and stable environmental conditions foster population growth and intraspecific competition due to accordingly declining availability of resources (MacArthur & Wilson, 1967); in this case, decelerated (slow) LH strategies such as investing in (both organisms’ own and their offspring’s) growth and development would be favored as they enhance individuals’ reproductive competitiveness (Geary, 2000; Geary & Flinn, 2001).
Although the linkage between environmental adversities and LH strategies has originally been proposed based on between-species observations (Stearns & Rodrigues, 2020), it should be applicable for explaining environment-induced LH variations at the within-species level (Del Giudice, 2020; Del Giudice & Belsky, 2011; Kuzawa & Bragg, 2012). Proxies of environmental risks widely adopted in human LH literature encompass socioeconomic status (SES), income-to-need ratio, parental absence and transition, family chaos, unsafe neighborhood, and parenting style (Brumbach et al., 2009; Chang et al., 2019b; Dunkel et al., 2015; Griskevicius et al., 2011; Griskevicius et al., 2011; Li et al., 2018; Lu et al., 2022b). Ample psychological and behavioral manifestations have been identified as important correlates of LH strategies, constituting species-specific LH profiles. For example, risk-taking, aggression, impulsivity, and associated deviant behaviors have been found associated with heightened early environmental adversities (Brumbach et al., 2009; Chang et al., 2019b; Kometani & Ohtsubo, 2022; Lu & Chang, 2019; Salas-Rodríguez et al., 2021). High extrinsic risk was found associated with poor socioemotional functioning (Li & Belsky, 2022; Li et al., 2018). Sociocultural factors, such as social attitudes toward and beliefs about gender, morality, and sexuality have also been linked to environmental adversities (Brumbach et al., 2009; Gladden et al., 2009; Zhu et al., 2018, 2022).
Parenting and Calibration of Offspring’s LH
Children first experience the external world through their interactions with caregivers (Bornstein, 2019; Warren & Barnett, 2020). The outcome of such child-caregiver interaction relies on the way in which parenting effort transmits specific information about the outside world to the child. It has been posited in the LH literature that cues about ecological conditions are conveyed to the child through the caregiver’s resource-allocation decisions and specific parenting behaviors (Chisholm, 1996; Del Giudice & Belsky, 2011; Simpson & Belsky, 2008). That said, in addition to the direct environmental contingencies on children’s LH development, parenting effort may enact as a key pivot that (1) undergoes and responds to environmental effects and transmits environmental cues to the child, (2) calibrates the child’s LH in parallel with environmental adversities, and (3) probably interacts with environmental contingencies on the child’s LH calibration.
As put earlier, harsh and variable environmental conditions shift individuals’ pace of life toward the fast end; that is, individuals are expected to mature early and reproduce as many and diverse offspring as they can, as such to offset the detrimental effects of extrinsic morbidity-mortality risks. This also implies that parental investment in each offspring would be relatively compromised, and such compromised parenting effort may exert a secondary effect on the children’s pace of life. As posited by Belsky et al. (1991), harsh, rejecting, inconsistent, and/or insensitive rearing during the first 5 to 7 years of life are associated with insecure attachment, a mistrustful internal work model, and opportunistic interpersonal orientations, and, subsequently, earlier sexual maturity and activity as well as short-term pair-bonding. Other literature revealed that benign and stable environments are in line with consistent and responsive parenting behaviors in parents and slow LH-related traits in their children; by contrast, adverse and stochastic environmental conditions are aligned with harsh, inconsistent, and/or unresponsive parenting and fast LH calibrations (e.g., Belsky et al., 2007, 2010; Brumbach et al., 2009; Chen & Chang, 2012; Del Giudice & Belsky, 2011; Dunkel et al., 2015; Ellis & Essex, 2007; Warren & Barnett, 2020).
Parenting effort may also be understood as a regulator of environmental contingencies on the child’s LH. First, parental care has evolved to operate as a “protecting belt” against juvenile-specific morbidity-mortality risks (Alonso-Alvarez & Velando, 2012; Bowlby, 1982), meaning that parenting effort may attenuate environmental effects on the child. Given that environmental adversities accelerate children’s pace of life, increased parental care and investment should decelerate children’s calibration of LH. Second, humans in modernized worlds are better able to manage extrinsic morbidity-mortality threats and are thus less subject to extrinsic population reductions, shifting human-specific LH to the slower end (Hill, 1993; Lu et al., 2021, 2022b). In this case, the main force of selection has shifted toward intraspecific competition (Alexander, 1990). Increased quality-focused parental investment would be favored in such competitive environments because (1) parents cannot afford allocating resources to a large number of children and (2) focused investments are more likely to pay off with increased competitiveness in each child (Sng & Ackerman, 2020; Sng et al., 2017). In other words, human-specific parental investment has the potential to independently decelerate the child’s pace of life. Building upon the literature and theorizing aforementioned, we propose that parenting investment may not only operate to directly shape children’s LH in parallel with environmental conditions, but also function to calibrate environmental contingencies on children’s LH development, resulting in decelerated LH-related manifestations. Such decelerating calibration can be achieved in two ways: (1) buffering the impacts of morbidity-mortality risks directly imposed on the child and (2) promoting slow LH-related traits that enhance the child’s competitiveness in the context of density-induced intraspecific competition.
The Present Study
The present research aimed to explore the role of parental investment in shaping children’s LH-related behavioral manifestations and moderating early environmental contingencies on children’s behavioral development. To examine our theorizing, we conducted a longitudinal study using data from the Adolescent Brain Cognitive Development study (ABCDSM release 4.0; https://abcdstudy.org) and the associated Social Development cohort study released by the NIMH Data Archive (NDA; https://nda.nih.gov). Specifically, we hypothesized that 1) while early environmental harshness and unpredictability would be positively associated with children’s fast LH profiles, parental investment would exhibit a negative relationship with children’s fast LH profiles, implying that parental investment may calibrate children’s LH in a different direction than environmental adversities. In addition, we hypothesized that parental investment would down-regulate the impacts of environmental harshness and unpredictability on children’s LH, demonstrating the decelerating effect of parenting on the accelerating calibration of environmental contingencies on fast LH profiles. To test this hypothesis, we generated interaction terms, environmental harshness × parental investment and environmental unpredictability × parental investment, at 1 standard deviation above and below the mean of parental investment. We expected stronger positive relationships between environmental harshness and unpredictability versus fast LH profiles when parental investment is low, and weaker relationships when parental investment is high.
Method
Sample
The present research adopts tabulated assessment data from the ABCDSM study and the Social Development cohort study released by the NDA. The ethical review and approval of research protocol are mostly issued by the central Institutional Review Board (IRB) at the University of California, San Diego, with a few endorsed by local IRBs (Auchter et al., 2018). The ABCDSM dataset contains longitudinal data collected from 11,876 9- to 10-year-old youths and their caregivers tracked by 2 to 3 yearly follow-ups after the baseline collection. The Social Development cohort study involves data from 2300 youths and their parents collected at two annual waves. Data from the two studies were matched by the time of collection. Based on our variables of interest, participants with completely missing data were not proceeded to statistical analyses. The sample of the present study consists of 2014 caregivers (female = 88.6%; Mage = 40.85 years, SD = 7.48; Non-Hispanic White = 58.3%, Non-Hispanic Black = 26.2%, Hispanic = 6.6%, Asian = 2.8%, and Others = 6.0%) of 9- to 12-year-old youths (female = 48.5%; Mage = 10.44 years, SD = 0.65; Non-Hispanic White = 49.4%, Non-Hispanic Black = 26.4%, Hispanic = 9.2%, Asian = 1.6%, and Others = 13.5%). In caregivers, 83.2% are the biological mothers of their youth, and 11.3% are biological fathers; 60.8% of the caregivers are married, 18.4% are never married, 8.8% are divorced, 3.0% are separated, and 8.0% are living with a partner. In families, 10.5% made a household combined income less than $12,000; 7.8% fall between $12,000 and 24,999; 14.2% between $25,000 and 49,999; 24.2% between $50,000 and 99,999; and 43.3% greater than $100,000.
Measures
Unsafe Neighborhood
Unsafe neighborhood was measured by 3-item 5-point Likert questionnaire inquiring about participants’ subjective experience about their living neighborhood. Caregivers responded to “I feel safe walking in my neighborhood, day or night,” “Violence is not a problem in my neighborhood,” and “My neighborhood is safe from crime” with 1 = “Strongly Disagree” through 5 = “Strongly Agree.” The data used was collected in the year 1 of the present study. Items were reverse-coded and averaged to derive a total score for statistical analysis. The Cronbach’s α of this measure is .90.
Family Conflict
The Conflict subscale of the Family Environment Scale – Parent Form (FES; Moos & Moos, 1994) was used to measure caregivers’ self-reported family conflict. The conflict subscale contains a total of 9 items (e.g., “We fight a lot in our family,” “Family members often criticize each other,” “Family members sometimes hit each other”), of which four are reverse items (e.g., “Family members rarely become openly angry,” “Family members hardly lose their tempers”). Caregivers responded to the items with 1 = “Yes” or 0 = “No,” and the sum of their responses generates a total score, indexing the extent of the intensity of family conflict. The data was collected at the year 2 of the present study. Because the total score was derived by summing up the binary responses, the measure was not tested for the internal consistency reliability.
Unpredictable Life Events
Unpredictable life events was adapted from the Adverse Life Event Scale – Parent Form (Tiet et al., 1998), a 25-item questionnaire measuring youths’ experienced adverse events up to the reporting year that they had few or no control. Caregivers were asked to respond 1 = “Yes” or 0 = “No” to whether their child experienced, for example, “Someone in family died,” “Negative change in parent’s financial situation,” “Parents argued more than previously,” “Mother/father figure lost job,” “Parents separated in last 12 months,” “Got new stepbrother or stepsister,” which were followed by further inquiries about whether this experience was good or bad (1 = “Mostly good,” 2 = “Mostly bad”) to their child and how much this experience affected their child (0 = “Not at all,” 1 = “A little,” 2 = “Some,” 3 = “A lot”). The data used was collected at the year 2 of the present study. Only experienced events that the parent reported “Mostly bad” and that affected their child “A little”, “Some,” or “A lot” were summed up to generate a total score.
Victimization Experiences
Youths’ victimization experiences were measured using the Juvenile Victimization Questionnaire (JVQ; Hamby et al., 2004). The JVQ is a 31-item retrospective checklist inquiring youths’ direct and/or witnessed victimization experiences. Experiences of victimization were modulated across five domains: conventional crime, child maltreatment, peer and sibling victimization, sexual victimization, and witnessing and indirect victimization. Sample questions involve “At any time in your child’s life, did anyone hit or attack your child on purpose with an object or weapon?” “…, did anyone break or ruin any of your child’s things on purpose?” “…, did your child see anyone get attached or hit on purpose with a stick, rock, gun, knife, or other things that would hurt?” Caregivers responded to the questions on their child’s behalf with 1 = “Yes” and 0 = “No.” The data was collected in the year 3 of this study. Items scores were summed to derive a total score.
Parental Investment
Parental investment was measured using the Parental Involvement and Positive Parenting subscales of the Alabama Parenting Questionnaire – Child Form (APQ; Frick, 1991; Shelton et al., 1996). The two subscales were adopted in the present study because they were more suited for the evolutionary conceptualization of parental investment. The two selected subscales contain 25 question items, 19 for parental involvement (9 identical items for both parents) and 6 for positive parenting. Sample items of parental involvement involve “Your mom helps with some of your special activities (such as sports, boy/girl scouts, church youth groups),” “You play games or do other fun things with your mom,” “Your mom helps you with your homework;” Sample items of positive parenting includes “Your parents tell you that you are doing a good job,” “Your parents compliment you when you have done something well,” “Your parents tell you that they like it when you help out around the house.” Each item was rated on 5-point Likert scale (1 = “Never” to 5 = “Always”), and item ratings of each subscale were averaged to generate a final score of the dimension. A higher final score indicates a higher level of the dimension. The data used was collected in the year 3 of the present study. The internal consistency reliability estimates of the Parental Involvement and Positive Parenting subscales are 0.93 and 0.88, respectively.
Aggression
The Aggression subscale of the 6–18 version of the Child Behavior Checklist (CBCL/6-18; Achenbach & Rescorla, 2001) was used to register the extent of youths’ behavioral aggression. The CBCL is a component of the Achenbach System of Empirically Based Assessment (ASEBA). The CBCL Aggression subscale consists of twenty items measuring both direct (e.g., “Gets in many fights,” “Physically attacks people,” “Threatens other people”) and relational aggression (e.g., “Temper tantrums or hot temper,” “Screams a lot,” “Stubborn, sullen, or irritable”; Ligthart et al., 2005), of which each is rated on a 3-point scale (0 = “Absent” to 2 = “Occurs often”). The data used was collected in the year 3 of the present study. The internal consistency reliability estimate of the CBCL Aggression subscale is 0.90.
Delinquency
Youths’ delinquency was indexed based on caregivers’ report on a self-developed 48-item questionnaire by the ABCD Social Development cohort study that records youths’ prevalence of delinquent behaviors. This questionnaire encompasses five domains of delinquency—vandalism, theft, violence, hidden weapon carrying, and police contact. Sample questions involve “Has your child ever received an in-school detention?” “Has your child cheated on school tests or assignments?” “Has your child taken something from a store without paying for it?” “Has your child hit a teacher or another grown-up at school?” “Has your child been stopped and questioned by the police?” Responses to items were rated on a 3-point scale (0 = “Never,” 1 = “Once or twice,” 2 = “More often”). The data was collected in the year 3 of the present study. The reliability estimate of this measure is 0.76.
Statistical Analysis
The data analysis involved the use of structural equation modeling with latent constructs rather than directly observed variables. Mplus 7.0 was employed for this analysis, utilizing full information maximum likelihood estimation to account for missing data. Model fit was assessed using recommended cut-off values, including the chi-square to degrees of freedom ratio (χ2/df < 10; Kline, 1998), Comparative Fit Index (CFI ≥ .90; Marsh et al., 1988), Tucker-Lewis Index (TLI ≥ .90; Marsh et al., 1988), Root Mean Squared Error of Approximation (RMSEA ≤ 0.08; Browne & Cudeck, 1993), Standardized Root Mean Square Residual (SRMR ≤ 0.08; Hu & Bentler, 1999), and minimum factor loading (loading > .30, Hair et al., 2006; loading > .32, Tabachnick & Fidell, 2013). An exploratory factor analysis was performed to identify and screen out indicator variables with a standardized loading less than .30 or loading more than one factor (Hair et al., 2006). The final theory-driven selection was comprised of two indicators of environmental harshness (e.g., unsafe neighborhood, victimization experiences), 2 indicators of environmental unpredictability (e.g., unpredictable life events, family conflict), 2 indicators of parental investment (e.g., parental involvement, positive parenting), and 2 loaded to fast LH behavioral profile (e.g., aggression, delinquency). The subsequent analysis included the interaction constructs representing the moderation effect of parental investment on the relationship between environmental harshness and unpredictability. The interaction constructs were computed using a log-likelihood ratio test, a default approach in Mplus (Muthén & Muthén, 2012). Instead of providing goodness of fit statistics, the log-likelihood ratio test generates a statistic called D, representing the difference in log-likelihood ratio value between the baseline model and the hypothesized interaction model.
Results
Table 1 presents the means, standard deviations, and zero-order correlations of the variables used in the current study. The intercorrelations were in line with our expectations that they were mostly statistically significant, supporting the LH hypotheses. In specific, indicators are positively correlated with one another within each latent construct. Between latent constructs, the proxies of environmental harshness (e.g., unsafe neighborhood, victimization experiences) and unpredictability (e.g., family conflict, unpredictable life events) are positively and significantly associated with that of fast LH profiles (e.g., aggression, delinquency). The indicators of parental investment (e.g., parental involvement, positive parenting) showed robust negative correlations with behavioral aggression and delinquency and partial negative correlations with environmental indicators. The intercorrelations between indicators laid the foundation for the following analysis of the structural relationships between environmental risks, parental investment, and the development of fast LH profiles.
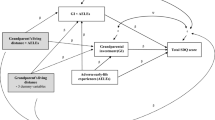
The initial model in Fig. 1, excluding the interaction terms, demonstrated acceptable fit statistics (χ2/df = 5.46, CFI = 0.98, TLI = 0.96, RMSEA = 0.047, SRMS = 0.023), meeting the recommended cut-off values for adequate model fit. As abovementioned, the present study employed a log-likelihood ratio test to determine the relative model fit between the baseline model and the hypothesized moderation model, referred to as D (Muthén & Muthén, 2012). In this study, the D value was 115.50, indicating a significant improvement in data fit by the hypothesized moderation model over the baseline model (p < .001).

The effects of environmental harshness, environmental unpredictability, parental investment, and their interactions on children’s fast LH behavioral profiles
In Fig. 1, all parameter estimates were statistically significant and aligned with the expected directions. Factor loadings, including the moderate ones involving environmental indicators, were adequate, meeting the minimum standards for measurement (Tabachnick & Fidell, 2013). The structural model’s parameter estimates were consistent with the hypotheses. Environmental harshness negatively influenced parental investment (β = −0.34, p < .001) and positively predicted fast LH behavioral profiles (β = 0.31, p < .001). Similarly, environmental unpredictability was negatively associated with parental investment (β = −0.47, p < .001) and positively associated with fast LH behavioral profiles (β = 0.22, p < .001). Parental investment demonstrated a negative relationship with fast LH profiles (β = −0.31, p < .001). The two interaction terms, representing parental investment in relation to environmental harshness (β = −0.24, p < .001) and unpredictability (β = − 0.59, p < .001), were statistically significant.
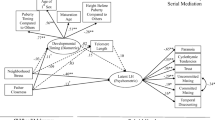
Figure 2 illustrates the simple slopes of environmental harshness and unpredictability on fast LH behavioral profiles at 1 standard deviation (SD) above (β = 0.25, p < .001; β = 0.30, p < .001) and 1 SD below the mean of parental investment (β = 0.39, p < .001; β = 0.44, p < .001). As predicted, parental investment attenuated the positive associations between environmental contingencies and fast LH strategies at the higher level of parental investment, while maintaining the associations at the lower level of parental investment.

Simple slopes of the regression of fast LH behavioral profiles on childhood environmental harshness (a) and childhood environmental unpredictability (b) at 1 SD above (dashed line) and 1 SD below (solid line) the mean of parental investment
Discussion
The present study examined the role of parental investment in formulating children’s LH-related behavioral profiles, as well as in moderating the relationships between environmental adversities (i.e., harshness and unpredictability) and children’s LH development. Consistent with previous literature (Brumbach et al., 2009; Chang et al., 2019a; Lu et al., 2022a), we found that higher levels of childhood environmental harshness and unpredictability predicted faster LH behavioral profiles. This finding is supported by the LH theoretical framework that environmental risks apply as a driving force that calibrates individuals’ LH strategies in response to their living ecological environments (Ellis, 2004; Ellis et al., 2009). Accelerated LH strategies would increase the likelihood of surviving to the age(s) of sexual maturity and first reproduction in harsh and variable environments. By contrast, investing in one’s own and the offspring’s growth and development, which enacts as to prime for future reproduction, would be more adaptive than investing for immediate reproductive returns in benign and stable environments (Chen & Kruger, 2017; Ellis et al., 2009). Although environmental harshness and unpredictability are conceptually distinct, they are highly correlated with and predicting LH-related traits in the same direction. This could be the reason why ample research employed either harshness or unpredictability as the main environmental construct, or treated indicators of both constructs jointly as that of a unified construct (environmental risks or adversities), as the hypothetical predictor of LH development. Previous research that conceptually distinguished the two constructs found that the impacts of harshness and unpredictability on LH calibration differed in terms of magnitude or statistical significance (e.g., Hartman et al., 2018; Li & Belsky, 2022; Szepsenwol et al., 2019). This is probably because this stream of research frequently adopted SES as the main indicator of harshness. The present research used different indicators of harshness (e.g., victimization experiences, unsafe neighborhood) and found that both harshness and unpredictability substantially affect children’s LH development, and such effects resemble in magnitude (β = 0.25, p < .001 and β = 0.28, p < .001).
Our findings also suggest that increased parental investment is associated with decreased accelerated LH behavioral manifestations, which is in line with LH-based parenting literature. For instance, Belsky et al. (2007) exhibited that harsh maternal parenting is associated with faster pubertal development (e.g., earlier menarche in girls). Dunkel et al. (2015) showed that maternal, but not paternal, sensitivity and authoritativeness positively predicted a significant increase in children’s slow LH strategy. This cluster of evidence is consistent with psychosocial acceleration theory (Belsky, 2019; Belsky et al., 1991), which posits that children capture environmental cues through the parent-child relationship and hence particularly rely on parental investment to forecast environmental conditions. The theory speculates that harsh, neglectful, and inconsistent parenting, representing low levels of parental investment, provide cues to children about levels of adversity in the external world, to which children may respond with accelerated LH-related traits (e.g., early puberty, increased mating effort, decreased mental/physical health; Cabeza de Baca & Ellis, 2017). Relatively, responsive and supportive parenting, reflecting high levels of parental investment, convey cues about safe and reliable external conditions and prompt the development of decelerated LH-related traits. Therefore, the level of parental investment can be seen as (1) a proxy of environmental conditions that directly calibrates children’s pace of life and (2) a pivot between environmental adversities and children’s LH development that transmits environmental cues to children and shapes their LH development in addition to environmental adversities. Also noticeable is the different directions of the impacts of parental investment on children’s LH development. When parental investment is low, it may apply as a force aligned with environmental contingencies, representing as a proxy of environmental adversities. However, when parental investment remains at a high level, it may showcase as an influential force in contrast to environmental contingencies and offers an influence in an alternative direction on children’s LH.
Following this speculation is our evidence on the moderating role of parental investment between environmental contingencies and children’s LH development. The directions of the moderating effects of parental investment are consistent with our expectations. We found stronger relationships between environmental contingencies (i.e., harshness and unpredictability) versus fast LH behavioral profiles at the lower level of parental investment and weaker relationships at the higher level of parental investment. This finding might be contrary to the species-general LH principle that extrinsic risks give rise to the development of accelerated LH traits, but instead consistent with species-specific expectations. Alexander (1990) argued that humans have evolved to be “ecologically dominant” with increased abilities in managing environmental contingencies. This led to our consideration of the additional role of parenting between environmental contingencies and children’s LH. As addressed earlier, parental investment may operate to buffer against the direct effects of extrinsic risks on children (Alonso-Alvarez & Velando, 2012; Bowlby, 1982; Chisholm, 1993), as well as convey knowledge and skills to children, as such to equip children with enhanced competitiveness (Chisholm, 1993; Geary, 2000). Based on these arguments, we propose that increased parental investment should calibrate the child’s LH in a decelerating fashion, a contrary direction against the force of extrinsic risks. Most importantly, the two competing forces of calibration should form an interaction. Given that children are expected to respond to age-specific morbidity-mortality risks with the development of accelerated LH-related traits, increased parental investment arises as an opposite force that decelerates such accelerating development.
The present study is subject to several limitations. First, although the data set includes different informants in triangulating on more valid and reliable information, some of the variables, e.g., child delinquency, ended up being derived from indirect sources, i.e., reported by caregivers, potentially creating source bias. Second, the factor loadings within environmental latent constructs range from low to moderate levels (0.30 ~ 0..60), implying that the indicators might not have fully captured the characteristics of the constructs as they should have. This may impose another layer of limitation on the validity of the present study, and there is reason to believe that more consistent environmental indicators could have resulted in nuanced findings. Third, we did not differentiate the caregivers by gender or age due to our focus on understanding the roles of parental investment from a holistic perspective. However, gender differences in caregiving should call for additional consideration, especially from the perspective of sexual conflict. Females and males are fundamentally subject to differential costs and benefits of caregiving, depending on the adult sex ratio, operational sex ratio, and sex-specific mortality in the population (Klug et al., 2012; Kokko & Jennions, 2008). On the other hand, Haig (2010) speculated a substantial conflict between mothers’ fertility and offspring’s survival based on observations that paternally inherited genes favor increased demand for maternal care, whereas maternally inherited genes favor relatively less demand for maternal care and earlier sexual maturation. These imply that paternal and maternal cares may differ by sex-specific interest of fitness, and there is reason to believe that such a difference may drive children’s LH development into different directions. Despite these limitations, the present study marks one of the initial attempts to explore the roles of parental investment in calibrating children’s LH, as well as regulating environmental contingencies on children’s LH development, with the goal of broadening the current understanding of human-specific decelerated pace of life.
Data Availability
The ABCD data repository grows and changes over time. Data from the ABCD study that support the findings of the present research are available in https://doi.org/10.15154/1523041.
References
Achenbach, T. M., & Rescorla, L. A. (2001). Manual for the ASEBA school-age forms and profiles. University of Vermont, Research Center for Children, Youth, & Families.
Alexander, R. D. (1990). How did humans evolve? Reflections on the uniquely unique species (Special Publication No. 1). Museum of Zoology, 1–38.
Alonso-Alvarez, C., & Velando, A. (2012). Benefits and costs of parental care. In N. J. Royle, P. T. Smiseth, & M. Kölliker (Eds.), The evolution of parental care (pp. 40–61). Oxford University Press.
Auchter, A. M., Hernandez Mejia, M., Heyser, C. J., Shilling, P. D., Jernigan, T. L., Brown, S. A., Tapert, S. F., & Dowling, G. J. (2018). A description of the ABCD organizational structure and communication framework. Developmental Cognitive Neuroscience, 32, 8–15. https://doi.org/10.1016/j.dcn.2018.04.003
Belsky, J. (2019). Early-life adversity accelerates child and adolescent development. Current Directions in Psychological Science, 28(3), Article 3. https://doi.org/10.1177/0963721419837670
Belsky, J., Steinberg, L., & Draper, P. (1991). Childhood experience, interpersonal development, and reproductive strategy: an evolutionary theory of socialization. Child Development, 62(4), 647–670. https://doi.org/10.1111/j.1467-8624.1991.tb01558.x
Belsky, J., Steinberg, L., Houts, R. M., Halpern-Felsher, B. L., & NICHD Early Child Care Research Network. (2010). The development of reproductive strategy in females: Early maternal harshness → earlier menarche → increased sexual risk taking. Developmental Psychology, 46(1), Article 1. https://doi.org/10.1037/a0015549
Belsky, J., Steinberg, L. D., Houts, R. M., Friedman, S. L., DeHart, G., Cauffman, E., Roisman, G. I., Halpern-Felsher, B. L., Susman, E., & The NICHD Early Child Care Research Network. (2007). Family rearing antecedents of pubertal timing. Child Development, 78(4), 1302–1321. https://doi.org/10.1111/j.1467-8624.2007.01067.x
Bornstein, M. H. (2019). Handbook of parenting (3rd ed., Vol. 1). Routledge.
Bowlby, J. (1982). Attachment and loss (2nd ed., Vol. 1). Basic Books.
Browne, M. W., & Cudeck, R. (1993). Alternative ways of assessing model fit. In K. A. Bollen & J. S. Long (Eds.), Testing structural equation models (pp. 136–162). Sage.
Brumbach, B. H., Figueredo, A. J., & Ellis, B. J. (2009). Effects of harsh and unpredictable environments in adolescence on development of life history strategies: a longitudinal test of an evolutionary model. Human Nature, 20(1), Article 1. https://doi.org/10.1007/s12110-009-9059-3
Cabeza de Baca, T., & Ellis, B. J. (2017). Early stress, parental motivation, and reproductive decision-making: applications of life history theory to parental behavior. Current Opinion in Psychology, 15, 1–6. https://doi.org/10.1016/j.copsyc.2017.02.005
Chang, L., & Lu, H. J. (2016). Environmental Risk. In V. Weekes-Shackelford, T. K. Shackelford, & V. A. Weekes-Shackelford (Eds.), Encyclopedia of Evolutionary Psychological Science (pp. 1–8). Springer International Publishing. https://doi.org/10.1007/978-3-319-16999-6_1918-1
Chang, L., Lu, H. J., Lansford, J. E., Bornstein, M. H., Steinberg, L., Chen, B. B., Skinner, A. T., Dodge, K. A., Deater-Deckard, K., Bacchini, D., Pastorelli, C., Alampay, L. P., Tapanya, S., Sorbring, E., Oburu, P., Al-Hassan, S. M., Di Giunta, L., Malone, P. S., Uribe Tirado, L. M., & Yotanyamaneewong, S. (2019a). External environment and internal state in relation to life-history behavioural profiles of adolescents in nine countries. Proceedings of the Royal Society B: Biological Sciences, 286(1917), Article 1917. https://doi.org/10.1098/rspb.2019.2097
Chang, L., Lu, H. J., Lansford, J. E., Skinner, A. T., Bornstein, M. H., Steinberg, L., Dodge, K. A., Chen, B. B., Tian, Q., Bacchini, D., Deater-Deckard, K., Pastorelli, C., Alampay, L. P., Sorbring, E., Al-Hassan, S. M., Oburu, P., Malone, P. S., Di Giunta, L., Tirado, L. M. U., & Tapanya, S. (2019b). Environmental harshness and unpredictability, life history, and social and academic behavior of adolescents in nine countries. Developmental Psychology, 55(4), Article 4. https://doi.org/10.1037/dev0000655
Chen, B. B., & Chang, L. (2012). Adaptive insecure attachment and resource control strategies during middle childhood. International Journal of Behavioral Development, 36(5), Article 5. https://doi.org/10.1177/0165025412445440
Chen, B. B., & Kruger, D. (2017). Future orientation as a mediator between perceived environmental cues in likelihood of future success and procrastination. Personality and Individual Differences, 108, 128–132. https://doi.org/10.1016/j.paid.2016.12.017
Chisholm, J. S. (1993). Death, hope, and sex: life-history theory and the development of reproductive strategies. Current Anthropology, 34(1), Article 1.
Chisholm, J. S. (1996). The evolutionary ecology of attachment organization. Human Nature, 7(1), Article 1. https://doi.org/10.1007/BF02733488
Del Giudice, M. (2020). Rethinking the fast-slow continuum of individual differences. Evolution and Human Behavior, 41(6), Article 6. https://doi.org/10.1016/j.evolhumbehav.2020.05.004
Del Giudice, M., & Belsky, J. (2011). The development of life history strategies: Toward a multi-stage theory. In D. M. Buss & P. H. Hawley (Eds.), The evolution of personality and individual differences (pp. 154–176). Oxford University Press.
Del Giudice, M., Gangestad, S. W., & Kaplan, S. (2015). Life history theory and evolutionary psychology. In D. M. Buss (Ed.), The Handbook of Evolutionary Psychology: Vol. Foundations (2nd ed., pp. 88–114). John Wiley & Sons, Inc.
Draper, P., & Harpending, H. (1982). Father absence and reproductive strategy: an evolutionary perspective. Journal of Anthropological Research, 38(3), 255–273. https://doi.org/10.1086/jar.38.3.3629848
Dunkel, C. S., Mathes, E. W., Kesselring, S. N., Decker, M. L., & Kelts, D. J. (2015). Parenting influence on the development of life history strategy. Evolution and Human Behavior, 36(5), Article 5. https://doi.org/10.1016/j.evolhumbehav.2015.02.006
Ellis, B. J. (2004). Timing of pubertal maturation in girls: an integrated life history approach. Psychological Bulletin, 130(6), Article 6. https://doi.org/10.1037/0033-2909.130.6.920
Ellis, B. J., & Essex, M. J. (2007). Family environments, adrenarche, and sexual maturation: a longitudinal test of a life history model. Child Development, 78(6), Article 6. https://doi.org/10.1111/j.1467-8624.2007.01092.x
Ellis, B. J., Figueredo, A. J., Brumbach, B. H., & Schlomer, G. L. (2009). Fundamental dimensions of environmental risk: the impact of harsh versus unpredictable environments on the evolution and development of life history strategies. Human Nature, 20(2), Article 2. https://doi.org/10.1007/s12110-009-9063-7
Frick, P. J. (1991). Alabama parenting questionnaire. In Unpublished rating scale.
Geary, D. C. (2000). Evolution and proximate expression of human paternal investment. Psychological Bulletin, 126(1), 55–77. https://doi.org/10.1037/0033-2909.126.1.55
Geary, D. C. (2002). Sexual selection and human life history. In Advances in Child Development and Behavior (Vol. 30, pp. 41–101). Elsevier. https://doi.org/10.1016/S0065-2407(02)80039-8
Geary, D. C., & Flinn, M. V. (2001). Evolution of human parental behavior and the human family. Parenting, 1(1–2), 5–61. https://doi.org/10.1080/15295192.2001.9681209
Gladden, P. R., Figueredo, A. J., & Jacobs, W. J. (2009). Life history strategy, psychopathic attitudes, personality, and general intelligence. Personality and Individual Differences, 46(3), Article 3. https://doi.org/10.1016/j.paid.2008.10.010
Griskevicius, V., Delton, A. W., Robertson, T. E., & Tybur, J. M. (2011). Environmental contingency in life history strategies: the influence of mortality and socioeconomic status on reproductive timing. Journal of Personality and Social Psychology, 100(2), Article 2. https://doi.org/10.1037/a0021082
Griskevicius, V., Tybur, J. M., Delton, A. W., & Robertson, T. E. (2011). The influence of mortality and socioeconomic status on risk and delayed rewards: a life history theory approach. Journal of Personality and Social Psychology, 100(6), Article 6. https://doi.org/10.1037/a0022403
Haig, D. (2010). Transfers and transitions: parent–offspring conflict, genomic imprinting, and the evolution of human life history. Proceedings of the National Academy of Sciences, 107(suppl_1), 1731–1735. https://doi.org/10.1073/pnas.0904111106
Hair, J. F., Black, W. C., Babin, B. J., Anderson, R. E., & Tatham, R. L. (2006). Multivariate data analysis. Pearson.
Hamby, S. L., Finkelhor, D., Ormrod, R., & Turner, H. (2004). The juvenile victimization questionnaire: Administration and scoring manual. Crimes Against Children Research Center. Retrieved August 17, 2023, from https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=a9f6b044502318b0631014af3b3c9aed2aadb10a
Hartman, S., Sung, S., Simpson, J. A., Schlomer, G. L., & Belsky, J. (2018). Decomposing environmental unpredictability in forecasting adolescent and young adult development: a two-sample study. Development and Psychopathology, 30(4), Article 4. https://doi.org/10.1017/S0954579417001729>
Hill, K. (1993). Life history theory and evolutionary anthropology. Evolutionary Anthropology: Issues, News, and Reviews, 2(3), 78–88. https://doi.org/10.1002/evan.1360020303
Hill, K., & Kaplan, H. (1999). Life history traits in humans: theory and empirical studies. Annual Review of Anthropology, 28(1), Article 1. https://doi.org/10.1146/annurev.anthro.28.1.397
Hu, L., & Bentler, P. M. (1999). Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Structural Equation Modeling: A Multidisciplinary Journal, 6(1), 1–55. https://doi.org/10.1080/10705519909540118
Kline, R. B. (1998). Principles and practice of structural equation modeling. Guilford Press.
Klug, H., Alonzo, S. H., & Bonsall, M. B. (2012). Theoretical foundations of parental care. In N. J. Royle, P. T. Simiseth, & M. Kölliker (Eds.), The evolution of parental care (pp. 21–39). Oxford University Press.
Kokko, H., & Jennions, M. D. (2008). Parental investment, sexual selection and sex ratios. Journal of Evolutionary Biology, 21(4), 919–948. https://doi.org/10.1111/j.1420-9101.2008.01540.x
Kometani, A., & Ohtsubo, Y. (2022). Can impulsivity evolve in response to childhood environmental harshness? Evolutionary Human Sciences, 4, e21. https://doi.org/10.1017/ehs.2022.22
Kuzawa, C. W., & Bragg, J. M. (2012). Plasticity in human life history strategy: implications for contemporary human variation and the evolution of genus Homo. Current Anthropology, 53(S6), Article S6. https://doi.org/10.1086/667410
Li, Z., & Belsky, J. (2022). Indirect effects, via parental factors, of income harshness and unpredictability on kindergarteners’ socioemotional functioning. Development and Psychopathology, 34(2), Article 2. https://doi.org/10.1017/S095457942100136X
Li, Z., Liu, S., Hartman, S., & Belsky, J. (2018). Interactive effects of early-life income harshness and unpredictability on children’s socioemotional and academic functioning in kindergarten and adolescence. Developmental Psychology, 54, 2101–2112. https://doi.org/10.1037/dev0000601
Ligthart, L., Bartels, M., Hoekstra, R. A., Hudziak, J. J., & Boomsma, D. I. (2005). Genetic contributions to subtypes of aggression. Twin Research and Human Genetics, 8(5), 483–491. https://doi.org/10.1375/twin.8.5.483
Lu, H. J., & Chang, L. (2019). Aggression and risk‐taking as adaptive implementations of fast life history strategy. Developmental Science, e12827. https://doi.org/10.1111/desc.12827
Lu, H. J., Liu, Y. Y., & Chang, L. (2022a). Child attachment in adjusting the species-general contingency between environmental adversities and fast life history strategies. Development and Psychopathology, 34(2), Article 2. https://doi.org/10.1017/S0954579421001413
Lu, H. J., Liu, Y. Y., O, J., Guo, S., Zhu, N., Chen, B. B., Lansford, J. E., & Chang, L. (2021). Disease history and life history predict behavioral control of the COVID-19 pandemic. Evolutionary Psychology, 19(1), Article 1. https://doi.org/10.1177/14747049211000714
Lu, H. J., Wang, X. R., Liu, Y. Y., & Chang, L. (2022b). Disease prevalence and fatality, life history strategies, and behavioral control of the COVID pandemic. Evolutionary Psychological Science, 8(1), Article 1. https://doi.org/10.1007/s40806-021-00306-9
MacArthur, R. H., & Wilson, E. O. (1967). The theory of island biogeography: Monographs in population biology. Princeton University Press. Retrieved August 17, 2023, from https://www.jstor.org/stable/j.ctt19cc1t2
Marsh, H. W., Balla, J. R., & McDonald, R. P. (1988). Goodness-of-fit indexes in confirmatory factor analysis: the effect of sample size. Psychological Bulletin, 103(3), Article 3. https://doi.org/10.1037/0033-2909.103.3.391
Moos, R. H., & Moos, B. S. (1994). Family environment scale manual: development, applications, and research (3rd ed.). Consulting Psychologists Press.
Muthén, L. K., & Muthén, B. O. (2012). Mplus: Statistical analysis with latent variables user’s guide (7th ed.). Authors. Retrieved September 23, 2023, from http://www.statmodel.com/download/LV%20Interaction.pdf
Promislow, D. E. L., & Harvey, P. H. (1990). Living fast and dying young: a comparative analysis of life-history variation among mammals. Journal of Zoology, 220(3), Article 3. https://doi.org/10.1111/j.1469-7998.1990.tb04316.x
Quinlan, R. J. (2007). Human parental effort and environmental risk. Proceedings of the Royal Society B: Biological Sciences, 274(1606), Article 1606. https://doi.org/10.1098/rspb.2006.3690
Rohner, R. P. (2004). The parental “Acceptance-Rejection syndrome”: universal correlates of perceived rejection. American Psychologist, 59(8), Article 8. https://doi.org/10.1037/0003-066X.59.8.830
Salas-Rodríguez, J., Gómez-Jacinto, L., & Hombrados-Mendieta, M. I. (2021). Life history theory: evolutionary mechanisms and gender role on risk-taking behaviors in young adults. Personality and Individual Differences, 175, 110752. https://doi.org/10.1016/j.paid.2021.110752
Sear, R. (2016). Beyond the nuclear family: an evolutionary perspective on parenting. Current Opinion in Psychology, 7, 98–103. https://doi.org/10.1016/j.copsyc.2015.08.013
Shelton, K. K., Frick, P. J., & Wootton, J. (1996). Assessment of parenting practices in families of elementary school-age children. Journal of Clinical Child Psychology, 25(3), 317–329. https://doi.org/10.1207/s15374424jccp2503_8
Simpson, J. A., & Belsky, J. (2008). Attachment theory within a modern evolutionary framework. In Handbook of attachment: theory, research, and clinical applications (2nd ed., pp. 131–157). The Guilford Press.
Sng, O., & Ackerman, J. M. (2020). Too many people, women, men? The psychological effects of population density and sex ratio. Current Opinion in Psychology, 32, 38–42. https://doi.org/10.1016/j.copsyc.2019.06.015
Sng, O., Neuberg, S. L., Varnum, M. E. W., & Kenrick, D. T. (2017). The crowded life is a slow life: population density and life history strategy. Journal of Personality and Social Psychology, 112(5), 736–754. https://doi.org/10.1037/pspi0000086
Stearns, S. C. (1989). Trade-offs in life-history evolution. Functional Ecology, 3(3), Article 3. https://doi.org/10.2307/2389364
Stearns, S. C. (1992). The evolution of life histories. Oxford University Press.
Stearns, S. C., & Rodrigues, A. M. M. (2020). On the use of “life history theory” in evolutionary psychology. Evolution and Human Behavior, 41(6), Article 6. https://doi.org/10.1016/j.evolhumbehav.2020.02.001
Szepsenwol, O., Zamir, O., & Simpson, J. A. (2019). The effect of early-life harshness and unpredictability on intimate partner violence in adulthood: a life history perspective. Journal of Social and Personal Relationships, 36(5), Article 5. https://doi.org/10.1177/0265407518806680
Tabachnick, B. G., & Fidell, L. S. (2013). Using multivariate statistics (6th ed.). Pearson.
Tiet, Q. Q., Bird, H. R., Davies, M., Hoven, C., Cohen, P., Jensen, P. S., & Goodman, S. (1998). Adverse life events and resilience. Journal of the American Academy of Child & Adolescent Psychiatry, 37(11), 1191–1200. https://doi.org/10.1097/00004583-199811000-00020
Warren, S. M., & Barnett, M. A. (2020). Effortful control development in the face of harshness and unpredictability. Human Nature, 31(1), Article 1. https://doi.org/10.1007/s12110-019-09360-6
West, S. A., & Gardner, A. (2013). Adaptation and inclusive fitness. Current Biology, 23(13), Article 13. https://doi.org/10.1016/j.cub.2013.05.031
Zhu, N., Chen, B. B., Lu, H. J., & Chang, L. (2022). Life-history calibration of social hierarchies: childhood adversity predicts leadership preference through relational social investment. Personality and Individual Differences, 188, 111482. https://doi.org/10.1016/j.paid.2021.111482
Zhu, N., Hawk, S. T., & Chang, L. (2018). Living slow and being moral: life history predicts the dual process of other-centered reasoning and judgments. Human Nature, 29(2), Article 2. https://doi.org/10.1007/s12110-018-9313-7
Acknowledgements
Data used in the preparation of this article were obtained from the Adolescent Brain Cognitive Development (ABCDSM) Study (https://abcdstudy.org), held in the NIMH Data Archive (NDA). This is a multisite, longitudinal study designed to recruit more than 10,000 children age 9–10 and follow them over 10 years into early adulthood. The ABCD Study® is supported by the National Institutes of Health and additional federal partners under award numbers U01DA041048, U01DA050989, U01DA051016, U01DA041022, U01DA051018, U01DA051037, U01DA050987, U01DA041174, U01DA041106, U01DA041117, U01DA041028, U01DA041134, U01DA050988, U01DA051039, U01DA041156, U01DA041025, U01DA041120, U01DA051038, U01DA041148, U01DA041093, U01DA041089, U24DA041123, U24DA041147. A full list of supporters is available at https://abcdstudy.org/federal-partners.html. A listing of participating sites and a complete listing of the study investigators can be found at https://abcdstudy.org/consortium_members/. ABCD consortium investigators designed and implemented the study and/or provided data but did not necessarily participate in the analysis or writing of this report. This manuscript reflects the views of the authors and may not reflect the opinions or views of the NIH or ABCD consortium investigators.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data analysis was performed by HJL. The first draft of the manuscript was written by ATY, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics Approval
The ABCD research sites are subject to the ethical review and approval by the central Institutional Review Board (IRB) at the University of California, San Diego, and IRBs at local institutions.
Consent to Participate
Informed consent to participate was not necessary because all data used for the present study came from the National Institute of Mental Health Data Archive.
Consent to Publish
Informed consent to publish was not necessary because all data used for the present study came from the National Institute of Mental Health Data Archive.
Competing Interests
The authors declare no competing interests.
Disclaimer
This manuscript reflects the views of the authors and may not reflect the opinions or views of the NIH or ABCD consortium investigators.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yang, A.T., Lu, H.J. & Chang, L. Environmental Harshness and Unpredictability, Parenting, and Offspring Life History. Evolutionary Psychological Science 9, 451–462 (2023). https://doi.org/10.1007/s40806-023-00375-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40806-023-00375-y