
Abstract
In this paper, the dynamical behaviors for a five-dimensional virus infection model with diffusion and two delays which describes the interactions of antibody, cytotoxic T-lymphocyte (CTL) immune responses and a general incidence function are investigated. The reproduction numbers for virus infection, antibody immune response, CTL immune response, CTL immune competition and antibody immune competition, respectively, are calculated. By using the Lyapunov functionals and linearization methods, the threshold conditions on the global stability of the equilibria for infection-free, immune-free, antibody response, CTL response and antibody and CTL responses, respectively, are established if the space is assumed as homogeneous. When the space is inhomogeneous, the effects of diffusion, intracellular delay and production delay are obtained by the numerical simulations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Mathematical models have been developed to explore mechanisms and dynamical behaviors in host virus infection process, and these provide insights into our understanding of HIV and other viruses; for example, HBV, HCV, influenza, SARS and Ebola are formulated and studied in many articles. Mathematical analysis for these models are necessary to obtain an integrated view for the virus dynamics in vivo. Nowak and Bangham (1996) pointed out that cytotoxic T-lymphocyte (CTL) immune responses play a critical part in antiviral defense by attacking virus-infected cells in most virus infections. They proposed the basic mathematical model describing immune responses against infected cells
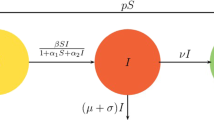
where the uninfected susceptible host cells u are produced at a rate \(\lambda \), die at rate d, and become infected at rate \(\beta \). Infected host cells, w, die at rate a and are killed by the CTL response at rate p. Free virus v are produced from infected cells at rate k and are removed at rate m. The variable z denotes the magnitude of the CTL response, which expands in response to viral antigen derived from infected cells at rate c, and decays in the absence of antigenic stimulation at rate b.
Usually the rate of infection in most virus infection models is assumed to be bilinear in the virus v and the uninfected cells u. However, the actual incidence rate is probably not linear over the entire range of v and u. Thus, it is reasonable to assume that the infection rate is given by the Beddington–DeAngelis functional response, \(\frac{\beta u(t)v(t)}{1+a_1u(t)+a_2v(t)}\), where \(a_1,a_2>0\) are constants. The functional response \(\frac{\beta u(t)v(t)}{1+a_1u(t)+a_2v(t)}\) was introduced by Beddington (1975) and DeAngelis et al. (1975). It is similar to the well-known Holling type II functional response but has an extra term \(a_2v\) in the denominator which models mutual interference between virus. When \(a_1>0; a_2=0\), the Beddington–DeAngelis functional response is simplified to Holling type II functional response (Li and Ma 2007). And when \(a_1=0\) and \(a_2>0\), it expresses a saturation response (Song and Neumann 2007). They obtained some criterion for the local asymptotic stability of the positive equilibrium of model (1) and gave the global stability of the positive equilibrium by constructing Lyapunov functions. Balasubramaniam et al. (2015) and Pawelek et al. (2012) performed detailed qualitative and bifurcation analysis such as the stability of equilibria and Hopf bifurcation.
Note that it is implicitly assumed that cells and viruses are well mixed, and the spatial mobility of cells and viruses has been ignored in model (1). Model (1) has been traditionally formulated in relation to the time evolution of uniform population distributions in a habitat and areas such governed by ordinary differential equations. However, as discussed by Wu (1996), in many biological systems, the species under consideration may disperse spatially as well as evolving in time. The mobility of susceptible cells, infected cells and immune cells is further neglected under normal conditions, but viruses move freely in body in McCluskey and Yang (2015), Gourley and So (2002), Xu and Ma (2009), Hattaf and Yousfi (2013, 2015), Wang et al. (2011, 2014) and Zhang and Xu (2014). They introduced the random mobility for viruses into model (1) and assume that the motion of virus follows the Fickian diffusion. Yang and Xu (2016) proposed the following virus infection model with spatial dependence
where \(u(x, t),\; w(x, t),\; v(x, t)\) and z(x, t) represent the densities of uninfected cells, infected cells, free virus and immune cells at location x and time t, respectively. The Laplacian operator and the diffusion coefficient are denoted by \(\triangle \) and D, respectively. It is demonstrated in model (2) that by constructing Lyapunov functionals and using LaSalle’s invariance principle, the global stability of the model is established. More recently, the global dynamics of diffusive virus dynamic models have been studied in McCluskey and Yang (2015), Gourley and So (2002), Xu and Ma (2009), Hattaf and Yousfi (2013, 2015), Wang et al. (2011, 2014) and Zhang and Xu (2014).
During viral infections, the immune system reacts against virus. The antibody and CTLs play the crucial roles in preventing and modulating infections. The antibody response is implemented by the functioning of immunocompetent B lymphocytes. The CTL response has the ability to suppress the virus replication in vivo. Hence, an effective vaccine to prevent virus infection needs both strong neutralizing antibody and CTL responses (Balasubramaniam et al. 2015; Wodarz 2003; Yan and Wang 2012; Wang et al. 2014). Therefore, some of the typical HIV infection models are described by delay differential equations, considering the dynamics of target cell, virus populations and immune response has been studied in recent years (Nelson and Perelson 2000; Yan and Wang 2012; Zhu and Zou 2009; Shu et al. 2013; Yuan and Zou 2013; Balasubramaniam et al. 2015; Wang et al. 2012, 2014; Pawelek et al. 2012; Huang et al. 2011; Ji 2015; Lu et al. 2015; Xiang et al. 2013). There are some models which include intracellular delay (Nelson and Perelson 2000; Yan and Wang 2012; Zhu and Zou 2009; Shu et al. 2013; Wang et al. 2012, 2014; Pawelek et al. 2012; Huang et al. 2011); some authors believe that time delays cannot be ignored in models for production viruses (Shu et al. 2013; Wang et al. 2014; Ji 2015; Xiang et al. 2013). Therefore, it is more realistic to investigate delayed virus infection models with antibody and CTL responses and nonlinear incidences. However, to our knowledge, there are few works on diffusive virus dynamics model with time delay and adaptive immune response.
Motivated by the works of Yang and Xu (2016), Yan and Wang (2012), Wang et al. (2014) and McCluskey and Yang (2015), we propose a delayed virus infection model with generalized incidence rate and spatial diffusion
for \(t>0,x\in \Omega \), where y(x, t) represents the densities of antibody cells at location x and time t, h represents the death rate of the antibody response, q is the antibody cells neutralize rate, g is the birth rate of the antibody response. And the other parameters are the same meaning as model (1).
In model (3), based on the epidemiological background, to incorporate the intracellular phase of the virus life cycle, we assume that virus production occurs after the virus entry by the intracellular delay \(\tau _1\). The recruitment of virus-producing cells at time t is given by the number of the uninfected cells that were newly infected at time \(t-\tau _1\) and are still alive at time t (Nelson and Perelson 2000; Yan and Wang 2012; Zhu and Zou 2009; Shu et al. 2013; Wang et al. 2012, 2014; Pawelek et al. 2012; Huang et al. 2011). The constant \(a_1\) is assumed to be the death rate for newly infected cells during time period \([t-\tau _1,t]\). \(\mathrm{e}^{-a_1\tau _1}\) denotes the surviving rate of infected cells during the delay period. Virus replication delay \(\tau _2\) represents the time necessary for the newly produced viruses to become mature and then infectious, that is, the maturation time of the newly produced viruses (Shu et al. 2013; Wang et al. 2014; Ji 2015; Xiang et al. 2013). The constant \(a_2\) is assumed to be the death rate for new virus during time period \([t-\tau _2,t]\). \(\mathrm{e}^{-a_2\tau _2}\) denotes the surviving rate of virus during the delay period.
We assume that the contacts between target cells, infected cells and viruses are given by an incidence function f(u, w, v), which is assumed to satisfy the following conditions:
\((A_1)\) Function \(f:\mathbb {R}_{+}^5\rightarrow \mathbb {R}_{+}\) is continuously differentiable; \(f(0,w,v)=0\) for all \(w\ge 0\) and \(v\ge 0;\) \(\frac{\partial f(u,w,v)}{\partial u}>0,\;\frac{\partial f(u,w,v)}{\partial w}\le 0\) and \(\frac{\partial f(u,w,v)}{\partial v}\le 0\) for all \(u\ge 0,\; w\ge 0\) and \(v\ge 0.\)
From assumption \((A_1)\), we easily obtain that there are no new infected cells (i.e., \(f(u,w,v)=0\)) without healthy cells (\(u= 0\)) or virus (\(v=0\)). If the total number of virus is constant, the more the amount of cell is, then the more the average number of cells which are infected by each virus in the unite time will be. If the total number of cells is constant, the more the amount of infected cells or virus is, then the less the average number of cells which are infected by each infected cell or virus in the unite time will be.
It is easy to check that class of functions f(u, w, v) satisfying \((A_1)\) include incidence functions such as \(f(u,w,v) =\frac{\beta uv}{1+bv}\) (Wang et al. 2013), \(f(u,w,v)=\frac{\beta uv}{1+au+bv}\) (Huang et al. 2011) and \(f(u,w,v) = \frac{\beta uv}{1+au+bv+cuv}\) (Zhou and Cui 2011), where constants \(\beta , a, b, c >0\).
We consider model (3) with initial conditions
and homogeneous Neumann boundary conditions
where \(\tau =\max \{\tau _1,\;\tau _2\},\) \(\Omega \) is a connected, bounded domain in \(\mathbb {R}^n\) with smooth boundary \(\partial \Omega .\) \(\frac{\partial }{\partial \vec {n}}\) denotes the outward normal derivative on \(\partial \Omega .\) \(\phi _i(x,\theta )(i=1,2,3,4,5)\) is H\(\ddot{o}\)lder continuous in \(\bar{\Omega }\times [-\tau ,0].\) The boundary conditions in (5) imply that the virus particles do not move across the boundary \(\partial \Omega \). \(\Delta \) is the Laplacian operator. D is the diffusion coefficient of the virus particles.
In this paper, our purpose is to investigate the dynamical properties of model (3), expressly the stability of equilibria. The reproduction numbers for viral infection, antibody immune response, CTL immune response, CTL immune competition and antibody immune competition, respectively, are calculated. By using Lyapunov functionals and LaSalle’s invariance principle, the threshold conditions for the global asymptotic stability of equilibria for infection-free \(E_0\), immune-free \(E_1\), antibody response \(E_2\), and infection only with CTL response \(E_3\) and infection with both antibody and CTL responses \(E_4\) are established, respectively. By using the linearization method, the instability of equilibria for \(E_0, E_1, E_2\) and \(E_3\), respectively, also is established.
The organization of this paper is as follows. In the next section, the basic properties of model (3) for the positivity and boundedness of solutions, the threshold values and the existence of equilibria are discussed. In Section 3, under the additional assumptions \((A_1)\)–\((A_2)\), the threshold conditions on the global stability and instability for \(E_0, E_1, E_2, E_3\) and \(E_4\) are stated and proved. In Sect. 4, the numerical simulations are given to further illustrate the dynamical behavior of the model. In the last section, we will give a conclusion.
2 Positivity, boundedness and equilibrium
In this section, we show the existence, positivity and boundedness of solutions of model (3)–(5) as they represent the densities of uninfected cells, infected cells, free virus, CTL immune cells and antibody cells. Further, we discuss the existence of equilibria of model (3).
Let \(C= C([-\tau ,0],X)\) be the Banach space of continuous functions from \([-\tau ,0]\) into X with the norm \(\parallel \phi \parallel \,=\,\max _{\theta \in [-\tau ,0]}\parallel \phi (\theta )\parallel _{X}\). In our case, X is the Banach space \(C(\overline{\Omega }, R^5)\) and C(E, F) denotes the space of continuous functions from the topological space E into the space F. For convenience, we identify an element \(\phi \in C\) as a function from \(\overline{\Omega }\times [-\tau ,0]\) into \(R^5\) defined by \(\phi (x,s)=\phi (s)(x)\).
For any continuous function \(\omega (\cdot ): [-\tau ,b)\rightarrow X\) for \(b>0\), we define \(\omega _t\in C\) by \(\omega _t(s) = \omega (t+s),\; s\in [-\tau ,0]\). It is not hard to see that \(t\rightarrow \omega _t\) is a continuous function from [0, b) to C.
Theorem 2.1
For any given initial data \(\phi \in C\) satisfying the condition (4), there exists a unique solution of model (3)–(5) defined on \([0,+\infty )\) and this solution remains nonnegative and bounded for all \(t\ge 0\).
Proof
For any \(\phi = (\phi _1,\phi _2,\phi _3,\phi _4,\phi _5)^T\in C\) and \(x\in \overline{\Omega }\), we define \(F=(F_1,F_2,F_3,F_4, F_5):C\rightarrow X\) by
Then, model (3)–(5) can be rewritten as the following abstract functional differential equation:
where \(\omega = (u,w,v,z,y)^\mathrm{T} , \phi = (\phi _1,\phi _2,\phi _3,\phi _4,\phi _5)^\mathrm{T}\) and \(A\omega =(0,0,D\triangle v,0,0)^\mathrm{T}\). It is clear that F is locally Lipschitz in X. From Wu (1996), we deduce that model (6) admits a unique local solution on \([0, T_{\mathrm{max}})\), where \(T_{\mathrm{max}}\) is the maximal existence time for solution of model (6).
Therefore, we have \(u(x, t)\ge 0,\;w(x, t)\ge 0,\; v(x, t)\ge 0,\;z(x, t)\ge 0\) and \(y(x, t)\ge 0\) because 0 is a sub-solution of each equation of model (3).
Next, we prove the boundedness of solutions. Denote
So we have
where \(l_1= \min \{d,a,b\}\). Hence,
This implies that \(u,\;w\) and z are bounded for large t.
From the boundedness of w and (3)–(5), we deduce that v satisfies the following system
where \(\xi =\max (\frac{\lambda \mathrm{e}^{-a_1\tau _1}}{l_1},\max _{x\in \overline{\Omega }} \{\mathrm{e}^{-a_1\tau _1}\phi _1(x,\tau _1)+\phi _2(x,0)+\frac{p}{c}\phi _4(x,0)\}).\)
Let \(v_1(t)\) be a solution to the ordinary differential equation
Denote
So we can get
where \(l_2= \min \{m,h\}\). Hence,
Then \(v_1(t)\le \max (\frac{k\mathrm{e} ^{-a_2\tau _2}\xi }{l_2},\max _{x\in \overline{\Omega }} \{\phi _3(x,0)+\frac{q}{g}\phi _5(x,0)\}).\)
From the comparison principle Protter and Weinberger (1967), we get \(v(x, t)\le v_1(t)\). Hence,
From the above, we have proved that \(u(x,t),\;w(x, t),\;v(x, t),\;z(x, t)\) and y(x, t) are bounded on \(\overline{\Omega }\times [0, T_{\mathrm{max}})\). Therefore, it follows from the standard theory for semilinear parabolic systems (Henry 1993; Redlinger 1984) that \(T_{\mathrm{max}} = +\infty \). This completes the proof. \(\square \)
Now, we discuss the existence of equilibria of model (3). It is easy to know that any equilibrium \(E =(u, w, v, z,y)\) of model (3) satisfies
It is clear from (7) that model (3) always has a unique infection-free equilibrium \(E_0=(u_0,0,0,0,0)\) with \(u_0=\frac{\lambda }{d}.\)
The basic reproductive number of viral infection for model (3) is
If \(z=0\) and \(y=0,\) then we get the following equation
Since \(w\ge 0,\) we have \(u\le \frac{\lambda }{d}.\) Denote
We have
and
Because of \((A_1)\), we know that the function \(F_1(u)\) is strictly monotonically increasing with respect to u. When \(R_0 > 1,\) there exists a unique \(u_1\in (0,\frac{\lambda }{d})\) such that \(F_1(u_1)=0.\) Thus, we obtain a unique immune-free equilibrium \(E_1=(u_1,w_1,v_1,0,0)\) with \(u_1\in (0,\frac{\lambda }{d}), w_1=\frac{\lambda -\mathrm{d}u_1}{a \mathrm{e}^{a_1\tau _1}}\) and \(v_1=\frac{k(\lambda -\mathrm{d}u_1)}{am \mathrm{e}^{a_1\tau _1+a_2\tau _2}}.\)
If \(y\ne 0\) and \(z=0,\) we have \(v=\frac{h}{g}.\) From the first and second equations of (7), we have
Since \(y=\frac{kg(\lambda -\mathrm{d}u)-amh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}{aqh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}\ge 0,\) we get \(u\le \frac{\lambda }{d}-\frac{amh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}{kgd}.\) Denote
We have \(F_2(0)=-\frac{\lambda g}{h}<0\) and \(F_2'(u)=\frac{\partial f}{\partial v}-\frac{{ d}}{a\mathrm{e}^{a_1\tau _1}}\cdot \frac{\partial f}{\partial w}+\frac{{ d}g}{h}>0.\)
Now, we define the antibody immune reproductive number for model (3) given by
Note that when \(R_0>1\) model (3) has a unique immune-free equilibrium \(E_1=(u_1,w_1,v_1,\) 0, 0). This shows that virus infection is successful and the numbers of free viruses at equilibrium \(E_1\) is \(v_1\). Furthermore, we have that \(\frac{1}{h}\) is the average life span of antibody cells, g is birth rate of the antibody response. Hence, \(R_1\) denotes the average number of the antibody immune cells activated by virus when virus infection is successful and CTL responses have not been established.
If \(R_1>1,\) then \(v_1>\frac{h}{g},\) \(u_1<\frac{\lambda }{d}-\frac{amh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}{kdg}\) and
Thus, if \(R_1>1,\) there exists a unique infection equilibrium with only antibody response \(E_2=(u_2,w_2,v_2,0,y_2)\) with \(u_2\in (0,\frac{\lambda }{d}-\frac{amh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}{kgd}),\; w_2=\frac{\lambda -\mathrm{d}u_2}{a\mathrm{e}^{a_1\tau _1}},\;v_2=\frac{h}{g}\) and \(y_2=\frac{kg(\lambda -\mathrm{d}u_2)-amh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}{aqh\mathrm{e}^{a_1\tau _1+a_2\tau _2}}.\)
If \(y=0\) and \(z\ne 0\), we have \(w=\frac{b}{c}\) and \(v=\frac{kb\mathrm{e}^{-a_2\tau _2}}{cm}\). From the first equation of (7), we obtain
As \(z=\frac{c(\lambda -\mathrm{d}u)\mathrm{e}^{-a_1\tau _1}-ab}{pb}\ge 0\) then \(u\le \frac{\lambda }{d}-\frac{ab\mathrm{e}^{a_1\tau _1}}{cd}.\) Denote
We have \(F_3(0)=-\frac{\lambda cm}{kb\mathrm{e}^{-a_2\tau _2}}<0\) and \(F_3'(u)=\frac{\partial f}{\partial u}+\frac{cmd}{kb\mathrm{e}^{-a_2\tau _2}}>0.\) Denote
which \(R_2\) denotes the average number of the CTL immune cells activated by infected cells when virus infection is successful and antibody immune responses have not been established. Note that the number of infected cells at equilibrium \(E_1\) is \(w_1, \frac{1}{b}\) is the average life span of CTL cells and c is the rate at which the CTL responses are produced.
We see that \(R_2>1\) is equivalent to \(w_1>\frac{b}{c},\;u_1<\frac{\lambda }{d}-\frac{ab}{cd\mathrm{e}^{-a_1\tau _1}}\) and
Hence, \(R_2>1\), there exists a unique infection equilibrium with only CTL response \(E_3=(u_3,w_3,v_3,z_3,0)\) with \(u_3\in (0,\frac{\lambda }{d}-\frac{ab}{cd\mathrm{e}^{-a_1\tau _1}}),\;w_3=\frac{b}{c},\; v_3=\frac{kb\mathrm{e}^{-a_2\tau _2}}{cm}\) and \(z_3=\frac{c(\lambda -\mathrm{d}u)\mathrm{e}^{-a_1\tau _1}-ab}{pb}.\)
If \(z\ne 0\) and \(y\ne 0\), we have \(w=\frac{b}{c}\) and \(v=\frac{h}{g}\). From the first equation of (7), we have
According to \(z=\frac{(\lambda -\mathrm{d}u)\mathrm{e}^{-a_1\tau _1}-aw}{pw}\ge 0,\) we deduce that \(u\le \frac{\lambda }{d}-\frac{ab\mathrm{e}^{a_1\tau _1}}{cd}.\) Define
We have \(F_4(0)=-\frac{\lambda g}{h}<0\) and \(F_4'(u)=\frac{\partial f}{\partial u}+\frac{dg}{h}>0.\)
The CTL immune competitive reproductive number for model (3) is
In fact, when \(R_1>1\), model (3) has a unique infection equilibrium with only antibody response \(E_2=(u_2,w_2,v_2,0,y_2)\). This predicates that CTL immune responses have been established, and the number of infected cells at equilibrium \(E_2\) is \(w_2\). Hence, \(R_3\) denotes the average number of the CTL immune cells activated by infected cells under the condition that antibody immune responses have been established.
If \(R_3>1\), then \(w_2>\frac{b}{c},\; u_2<\frac{\lambda }{d}-\frac{ab\mathrm{e}^{a_1\tau _1}}{cd}\) and
Thus, there exists a unique \(u_4\in (0,\frac{\lambda }{d}-\frac{ab\mathrm{e}^{a_1\tau _1}}{cd})\) such that \(F_4(u_4)=0.\) From the third equation of (7), we obtain that \(y_4=\frac{m}{q}(R_4-1),\) where \(R_4\) is the antibody immune competitive reproductive number defined by
In fact, when \(R_2>1\), model (3) has a unique infection equilibrium with only CTL response \(E_3=(u_3,w_3,v_3,z_3,0)\). This predicates that antibody immune responses have been established, and the numbers of the viruses at equilibrium \(E_3\) is \(v_3\). Hence, \(R_4\) denotes the average number of the antibody immune cells activated by viruses under the condition that CTL immune responses have been established.
When \(R_3 > 1\) and \(R_4 > 1\), model (3) has a unique infection equilibrium with CTL and antibody response \(E_4=(u_4,w_4,v_4,z_4,y_4)\) with \(u_4\in (0,\frac{\lambda }{d}-\frac{ab\mathrm{e}^{a_1\tau _1}}{cd}),\) \(w_4=\frac{b}{c},\;v_4=\frac{h}{g}, z_4=\frac{(\lambda -\mathrm{d}u_4)\mathrm{e}^{-a_1\tau _1}-aw_4}{pw_4}\) and \(y_4=\frac{m}{q}(R_4-1).\)
3 Stability analysis
In this section, we discuss global stability of equilibria for infection-free, immune-free, antibody response, and infection only with CTL response and infection with both antibody and CTL responses, respectively.
We further introduce the following assumption
\((A_2)\) \((1-\frac{f(u,w,v)}{f(u,w_i,v_i)})(\frac{f(u,w_i,v_i)}{f(u,w,v)}-\frac{v}{v_i})\le 0\) for all \(u,\;w,\;v>0\), where \(w_i\) and \(v_i\) are the components of equilibrium \(E_i \;(i=1,2,3,4).\)
For convenience, for any solution (u(x, t), w(x, t), v(x, t), z(x, t), y(x, t)) of model (3) we let
3.1 Stability of equilibrium \(E_0\)
Theorem 3.1
-
(a)
If \(R_0 \le 1\), then the infection-free equilibrium \(E_0\) is globally asymptotically stable.
-
(b)
If \(R_0>1\), then the equilibrium \(E_0\) is unstable.
Proof
Consider conclusion (a). Define a Lyapunov functional \(L_1(t)=\int _{\Omega } (V_1(x,t)+V_2(x,t))\,\mathrm {d}x,\) where
and
By calculation, we have
Calculating the time derivative of \(L_1(t)\) along any positive solution of model (3) and noticing that \(u_0=\frac{\lambda }{d}\), we can obtain
Using the divergence theorem and the homogeneous Neumann boundary conditions, we get
Thus,
Obviously, if \(R_0 \le 1\), then \(\frac{\mathrm{d}L_1(t)}{\mathrm{d}t}\le 0\) for any (u, w, v, z, y). We have \(\frac{\mathrm{d}L_1(t)}{\mathrm{d}t}= 0\) if and only if \( u=u_0, v=0, z=0\) and \(y=0\). Let M be the largest invariant set of \(\{(x,y,v,z,w)\in R^5_+: \frac{\mathrm{d}L_1(t)}{\mathrm{d}t}=0\}\). From the third equation of model (3), we easily obtain \(M=\{E_0\}.\) It follows from LaSalle’s invariance principle Hale and Verduyn (1993) that the equilibrium \(E_0\) of model (3) is globally asymptotically stable when \(R_0\le 1\).
Next, we consider conclusion (b). To do so, we determine the characteristic equation about the equilibrium \(E_0\).
Let \(0 =\mu _1<\mu _2< \cdots< \mu _{n} < \cdots \) be the eigenvalues of the operator \(-\Delta \) on \(\Omega \) with the homogeneous Neumann boundary conditions, and \(E(\mu _{i})\) be the eigenfunction space corresponding to \(\mu _{i}\) in \(C^1(\Omega )\). Let \(\{\varphi _{ij} : j = 1, 2,\ldots , \text{ dim }E(\mu _{i})\}\) be an orthonormal basis of \(E(\mu _{i}), \mathbb {X} = [C^1(\Omega )]^5\), and \(\mathbb {X}_{ij} = \{c\varphi _{ij }: c \in \mathbb {R}^5\}\). Then
Let \(E^*(u^*,w^*,v^*,z^*,y^*)\) be an arbitrary equilibrium, and consider the following change
By substituting U(x, t), W(x, t), V(x, t), Z(x, t) and Y(x, t) into model (3) and linearizing, we obtain the following system
This system is equivalent to
where
We put \(\mathbb {L}\mathbb {Z} = \mathbb {D}\triangle \mathbb {Z}+\mathbb {A}\mathbb {Z}(x,t) +\mathbb {B}\mathbb {Z}(x,t-\tau _1)+\mathbb {C}\mathbb {Z}(x,t-\tau _2).\) For each \(i\ge 1, \mathbb {X}_{i}\) is invariant under the operator \(\mathbb {L}\), and s is an eigenvalue of \(\mathbb {L}\) if and only if it is a root of the characteristic equation det\((sI-\mathbb {A}-\mathbb {B}\mathrm{e}^{-a_1\tau _1}-\mathbb {C}\mathrm{e}^{-a_2\tau _2}+\mu _{i}\mathbb {D}) = 0\) for some \(i\ge 1\), in which case, there is an eigenvector in \(\mathbb {X}_{i}\).
From (13), by computing, we obtain the characteristic equation of the corresponding linearized system of model (3) at the equilibrium \(E_0\) as follows
where
Obviously, \(s_1=-d, s_2 =-b\) and \(s_3 =-h\) are the roots of this equation. It is easy to prove that Eq. (15) has a real positive root when \(R_0>1\).
When \(R_0>1\), we have \(f_1(0)=am(1-R_0)<0\), as \(\mu _1=0\) when \(i=1.\) Since \(\lim _{s\rightarrow +\infty }f_i(s)=+\infty \), there is a \(s^*>0\) such that \(f_i(s^*)=0\). Therefore, when \(R_0>1,\) the equilibrium \(E_0\) is unstable. This completes the proof. \(\square \)
Biologically, Theorem 3.1 shows that the viruses are cleared and the infection dies out.
3.2 Stability of equilibrium \(E_1\)
Theorem 3.2
Assume \((A_2)\) holds, if \(R_0>1\) (a) \(R_1\le 1\) and \(R_2\le 1,\) then the immune-free equilibrium \(E_1\) is globally asymptotically stable.
(b) If \(R_1>1\) or \(R_2>1\), then the equilibrium \(E_1\) is unstable.
Proof
Define firstly function \(H(\xi )=\xi -1-\ln \xi \). We have that \(H(\xi )\ge 0\) for all \(\xi >0\) and \(H(\xi )=0\) if and only if \(\xi =1\). Consider conclusion (a). Define a Lyapunov functional
where
and
It is obvious that \(L_2(t)>0\) for all \((u(t),w(t),v(t),z(t),y(t))>0\) and \((u(t),w(t),v(t),z(t),y(t))\ne (u_1,w_1,v_1,0,0)\).
Calculating the time derivative of \(V_1(x,t)\) and \(V_2(x,t)\) along any positive solution of model (3), we can obtain
and
By using
Since
we have
Obviously, we always have \(\frac{\mathrm{d}L_2(t)}{\mathrm{d}t}\le 0,\) and \(\frac{\mathrm{d}L_2(t)}{\mathrm{d}t}=0\) if and only if \(u(t)=u_1,w(t)=w_1,v(t)=v_1, z(t)=0\) and \(y(t)=0.\) From LaSalle’s invariance principle Hale and Verduyn (1993), we finally have that the immune-free equilibrium \(E_1\) of model (3) is globally asymptotically stable when \(R_0>1, R_1\le 1\) and \(R_2\le 1\).
Next, we consider conclusion (b). From (13), by computing, we obtain the characteristic equation of the corresponding linearized system of model (3) at the equilibrium \(E_1\) as follows
where
When \(R_1>1\), we have \(h-gv_1<0\). Hence, there is a positive root \(s_1=gv_1-h\). When \(R_2>1\), there is also a positive root \(s_2=cw_1-b\). Therefore, when \(R_1>1\) or \(R_2>1,\) the equilibrium \(E_1\) is unstable. This completes the proof. \(\square \)
Biologically, Theorem 3.2 implies that when \(R_0>1, R_1\le 1\) and \(R_2\le 1\) then the establishments of both CTLs and antibody immune responses are unsuccessful.
3.3 Stability of equilibrium \(E_2\)
Theorem 3.3
Assume \((A_2)\) holds, if \(R_0>1\) and \(R_1>1\) (a) If \(R_3\le 1,\) then the antibody response equilibrium \(E_2\) is globally asymptotically stable.
(b) If \(R_3>1\), then the equilibrium \(E_2\) is unstable.
Proof
Consider conclusion (a). Define a Lyapunov functional \(L_3(t)\) as follows
where
and
It is obvious that \(L_3(t)>0\) for all \((u(t),w(t),v(t),z(t),y(t))>0\) and \((u(t),w(t),v(t), z(t),y(t))\ne (u_2,w_2,v_2,0,y_2)\), where \(u_2,w_2,v_2\) and \(y_2\) satisfy the following equations
Calculating the time derivative of \(L_3(t)\) along any positive solution of model (3), we can obtain
Obviously, we always have \(\frac{\mathrm{d}L_3(t)}{\mathrm{d}t}\le 0,\) and \(\frac{\mathrm{d}L_3(t)}{\mathrm{d}t}=0\) if and only if \(u(t)=u_2,w(t)=w_2,v(t)=v_2, z(t)=0\) and \(y(t)=y_2.\) From LaSalle’s invariance principle (Hale and Verduyn 1993), we finally have that the equilibrium \(E_2\) of model (3) is globally asymptotically stable when \(R_0>1, R_1\le 1\) and \(R_2\le 1\).
Next, we consider conclusion (b). From (13), by computing, we obtain the characteristic equation of the corresponding linearized system of model (3) at the equilibrium \(E_2\) as follows
where
where
When \(R_3>1\), we have \(s=cw_2-b>0.\) Therefore, when \(R_3>1\) equilibrium \(E_2\) is unstable. This completes the proof. \(\square \)
Biologically, Theorem 3.3 implies that when \(R_0>1, R_1>1\) and \(R_3\le 1\), the antibody response can be established, but the infected cells are too weak so that it cannot stimulate CTL immune response.
3.4 Stability of equilibrium \(E_3\)
Theorem 3.4
Assume \((A_2)\) holds, if \(R_0>1\) and \(R_2>1\) (a) If \(R_4\le 1\), then the infection equilibrium \(E_3\) with only CTL response is globally asymptotically stable.
(b) If \(R_4>1\), then the equilibrium \(E_3\) is unstable.
Proof
Consider conclusion (a). Define a Lyapunov functional \(L_4(t)\) as follows
where
and
We easily prove that \(L_4(t)>0\) for all \((u(t),w(t),v(t),z(t),y(t))>0\) and \((u(t),w(t), v(t),z(t),y(t))\ne (u_3,w_3,v_3,z_3,0)\).
By using
Calculating the time derivative of \(L_4(t)\) along any positive solution of model (3), we can obtain
Obviously, we always have \(\frac{\mathrm{d}L_4(t)}{\mathrm{d}t}\le 0,\) and \(\frac{\mathrm{d}L_4(t)}{\mathrm{d}t}=0\) if and only if \(u(t)=u_3,w(t)=w_3,v(t)=v_3, z(t)=z_3\) and \(y(t)=0.\) From LaSalle’s invariance principle (Hale and Verduyn 1993), we finally have that the equilibrium \(E_3\) of model (3) is globally asymptotically stable when \(R_0>1, R_2>1\) and \(R_4\le 1\).
Next, we consider conclusion (b). From (13), by computing, we obtain the characteristic equation of the linearization system of model (3) at the equilibrium \(E_3\) as follows
where
where
When \(R_4>1\), we have there is a positive root \(s_1=gv_3-h\). Therefore, when \(R_4>1\) equilibrium \(E_3\) is unstable for any \(\tau _1\ge 0\) and \(\tau _2\ge 0\). This completes the proof. \(\square \)
Biologically, Theorem 3.4 implies that, when \(R_0>1, R_2>1\) and \(R_4\le 1\), the CTL immune response can be determined, but the viral loads are so small that it cannot activate the antibody responses.
3.5 Stability of equilibrium \(E_4\)
Theorem 3.5
Assume \((A_2)\) holds, if \(R_0>1, R_1>1, R_3>1\) and \(R_4>1\), then the infection equilibrium with CTL and antibody responses \(E_4\) is globally asymptotically stable.
Proof
Define a Lyapunov functional \(L_5(t)\) as follows
where
and
It is obvious that \(L_5(t)>0\) for all \((u(t),w(t),v(t),z(t),y(t))>0\) and \((u(t),w(t),v(t), z(t),y(t))\ne (u_4,w_4,v_4,z_4,y_4)\).
Calculating the time derivative of \( L_5(t) \) along any positive solution of model (3), we can obtain
Obviously, we always have \(\frac{\mathrm{d}L_5(t)}{\mathrm{d}t}\le 0,\) and \(\frac{\mathrm{d}L_5(t)}{\mathrm{d}t}=0\) if and only if \(u=u_4,\;w=w_4,\;v=v_4.\) From the LaSalle’s invariance principle Hale and Verduyn (1993), we finally have that the equilibrium \(E_4\) of model (3) is globally asymptotically stable when \(R_0>1, R_1>1, R_3>1\) and \(R_4>1\). This completes the proof. \(\square \)
Biologically, Theorem 3.5 implies that, if CTL immune response has not any delay, then the susceptible cells, infected cells, free virus, CTL immune response and antibody immune response can coexist in vivo.
4 Numerical simulations
In this section, we perform some numerical simulations to illustrate the results obtained in Sect. 3. We consider model (3) under the homogeneous Neumann boundary conditions
and initial conditions
In model (3), we choose a nonlinear incidence \(f(u,w,v)=\frac{\beta u}{1+m_1u+n_1v+m_1n_1uv}\). Furthermore, \(\beta , g, h, \tau _1, \tau _2, c\) and b are chosen as free parameters and all remaining parameters are fixed as in Table 1.
In Figs. 1, 2, 3, 4 and 5a–e are denoted time series figures of u(x, t), w(x, t), v(x, t), z(x, t) and y(x, t).
5 Discussion
In this paper, we have discussed a delayed virus infection model (3) with diffusion, adaptive immune responses and general incidence rate. During viral infection, CTL immune responses which attack infected cells, and antibody responses which attack viruses. Hence, we assume that the production of CTL immune response depends on the infected cells and CTL immune responses. We see that similar assumption also is given in Nowak and Bangham (1996), Yan and Wang (2012), Zhu and Zou (2009), Shu et al. (2013), Wang et al. (2013, 2014, 2012) and Balasubramaniam et al. (2015). Similarly, the production of antibody response depends on the virus and antibody (Yan and Wang 2012; Wang et al. 2013; Balasubramaniam et al. 2015; Wang et al. 2014). Assumptions \((A_1)\) and \((A_2)\) for nonlinear function f(u, w, v)v are introduced and a combination of the basic reproduction number for viral infection \(R_0\), for CTL response \(R_1\), for antibody immune response \(R_2\), for CTL immune competition \(R_3\) and for humoral immune competition \(R_4\) defined by (8)–(12), respectively, also are defined. Under \((A_1)\) and \((A_2)\), the global stability and instability of the equilibria of model (3) by utilizing the method of constructing suitable Lyapunov functionals which are motivated by recent works of Pawelek et al. (2012), Zhu and Zou (2009), Shu et al. (2013), Yuan and Zou (2013) and Huang et al. (2011) are completely determined by the basic reproduction numbers \(R_0, R_1, R_2, R_3\) and \(R_4\).

Taking \(\beta =0.01,\; c=0.1,\; b=0.15,\; g=1.5,\; h=0.1,\;\tau _1=10,\;\tau _2=5\), we have \(R_0=0.2087<1\), the infection-free equilibrium \(E_0(1000,0,0,0,0)\) is asymptotically stable

Taking \(\beta =0.15,\; c=0.01,\; b=0.2,\; g=0.5,\; h=1.5,\;\tau _1=3,\;\tau _2=15\), we have \(R_0=3.0373>1,\;R_1= 0.7098<1\) and \(R_2=0.9277<1\), the immune-free equilibrium \(E_1(44.0253,18.5544,2.1293,0,0)\) is asymptotically stable
By the analysis, we have shown that when \(R_0\le 1\), the infection-free equilibrium \(E_0\) is globally asymptotically stable, which means that the viruses are cleared and the infection dies out. When \(R_0>1, R_1\le 1\) and \(R_2\le 1\) the immune-free equilibrium \(E_1\) is globally asymptotically stable, which means that immune response would not be activated and viral infection becomes vanished. When \(R_0>1, R_1>1\) and \(R_3\le 1\), the infection equilibrium with only antibody cells response \(E_2\) is globally asymptotically stable. As respect to the analysis of infection equilibrium \(E_3\) with only CTL response, when \(R_0>1,\) \(R_2>1\) and \(R_4\le 1, E_3\) is globally asymptotically stable, which means that the antibody response would not be activated and viral infection becomes vanished. About the stability of infection equilibrium \(E_4\) with both CTL and antibody response we have obtained that when \(R_3>1\) and \(R_4>1, E_4\) is globally asymptotically stable. We see that \((A_1)\) is basic for model (3). Particularly, when \(f(u,w,v)=\frac{\beta u}{1+m_1u+n_1v+m_1n_1uv}\) then \((A_1)\) naturally hold. But \((A_2)\) is a mathematical assumption. It is only used in the proofs of theorems on the global stability of equilibria \(E_1, E_2, E_3\) and \(E_4\) to obtain \(\frac{dL_n(t)}{dt}\) for the Lyapunov function \(L_n\) (see the proofs of Theorems 3.2–3.5). Furthermore, the numerical simulations given in Sect. 4 show the stability. Moreover, the effect of diffusion is considered as an important factor, which will be closer to reality. Compared to the case without diffusion, the approach is to construct Lyapunov functionals for partial differential equations (PDEs) or delayed partial differential equations (DPDEs) using Lyapunov functionals for ordinary differential equations (ODEs) or delayed differential equations (DDEs). Research on diffusion will be more complicated. Moreover, all the five state variables are influenced by multi-time delays and diffusion can better impact the virus infection problems. Therefore, research in this paper can be seen as an improvement and a supplementary of model (2), and it might be helpful to understand the virus infection model. Finally, under homogeneous Neumann boundary conditions, our results imply that diffusion, the intracellular delay and virus replication delay have no effect on the global behaviors of such virus dynamics model.

Taking \(\beta =0.25,\; c=0.01,\; b=0.18,\; g=1.5,\; h=1,\;\tau _1=10,\;\tau _2=5\), we have \(R_0=5.2164>1,\;R_1= 3.3682>1\) and \(R_3=0.8899<1\), the infection equilibrium only with CTL immune response \(E_2(114.8758,16.0179,0.6667,0,6.1420)\) is asymptotically stable

Taking \(\beta =0.35,\; c=0.1,\; b=0.15,\; g=1.5,\; h=1,\;\tau _1=10,\;\tau _2=5\), we have \(R_0=7.3030>1,\;R_2= 11.8885>1\) and \(R_4=0.2854<1\), the infection equilibrium only with antibody response \(E_3(455.1241,1.5000,0.1902,2.7868,0)\) is asymptotically stable

Taking \(\beta =0.45,\; c=0.15,\; b=0.15,\; g=0.1,\; h=0.01,\;\tau _1=2,\;\tau _2=5\), we have \(R_0=10.1716>1,\;R_3= 7.5777>1\) and \(R_4=1.2683>1\), the infection equilibrium with both antibody and CTL immune responses \(E_4(613.4595,1.0000,0.1000,3.2889,0.8049)\) is asymptotically stable
Observing all obtained results in this paper, we can directly put forward the following open question which need to be further studied in the future.
In this paper, we only discuss a five-dimensional diffusive virus infection model with intracellular delay, virus replication delay and general incidence rate. Based on different practical backgrounds, the immune response delay and mitotic proliferation terms for both uninfected and infected target cells are considered in modeling the viral infection of disease. Therefore, whether the results obtained in this paper also can be extended to five-dimensional diffusive virus infection model with mitosis transmission and immune delay. In other words, with immune delay as a bifurcation parameter, whether we also can obtain that the global asymptotic stability of equilibria for infection-free, immune-free, antibody response, infection with CTL response and infection with both antibody and CTL response, respectively, will also be a very estimable and significative subject.
References
Balasubramaniam P, Tamilalagan P, Prakash M (2015) Bifurcation analysis of HIV infection model with antibody and cytotoxic T-lymphocyte immune responses and Beddington–DeAngelis functional response. Math Methods Appl Sci 38:1330–1341
Beddington JR (1975) Mutual interference between parasites or predators and its effect on searching efficiency. J Anim Ecol 44:331–340
Culshaw RV, Ruan S, Spiteri RJ (2004) Optimal HIV treatment by maximising immune response. J Math Biol 48:545–562
DeAngelis DL, Goldstein RA, ÓNeill RV (1975) A model for trophic interaction. Ecology 56:881–892
Gourley SA, So JWH (2002) Dynamics of a food-limited population model incorporating nonlocal delays on a finite domain. J Math Biol 44:49–78
Hale JK, Verduyn SML (1993) Introduction to functional differential equations. Springer, New York
Hattaf K, Yousfi N (2013) Global stability for reaction–diffusion equations in biology. Comput Math Appl 66:1488–1497
Hattaf K, Yousfi N (2015) A generalized HBV model with diffusion and two delays. Comput Math Appl 69:31–40
Henry D (1993) Geometric theory of semilinear parabolic equations. In: Lecture notes in mathematics. Springer, Berlin
Huang G, Ma W, Takeuchi Y (2011) Global analysis for delay virus dynamics model with Beddington–DeAngelis function response. Appl Math Lett 24:1199–1203
Ji Y (2015) Global stability of a multiple delayed viral infection model with general incidence rate and an application to HIV infection. Math Biosci Eng 12:525–536
Li D, Ma W (2007) Asymptotic properties of an HIV-1 infection model with time delay. J Math Anal Appl 335:683–691
Lu X, Hui L, Liu S, Li J (2015) A mathematical model of HIV-1 infection with two time delays. Math Biosci Eng 12:431–449
McCluskey CC, Yang Y (2015) Global stability of a diffusive virus dynamics model with general incidence function and time delay. Nonlinear Anal RWA 25:64–78
Nelson P, Perelson JM (2000) A model of HIV-1 pathogenesis that includes an intracelluar delay. Math Biosci 163:201–215
Nowak MA, Bangham CRM (1996) Population dynamics of immune response to persistent viruses. Science 272:74–79
Pawelek KA, Liu S, Pahlevani F, Rong L (2012) A model of HIV-1 infection with two time delays: mathematical analysis and comparison with patient data. Math Biosci 235:98–109
Perelson AS, Kirschner DE, Boer RD (1993) Dynamics of HIV infection of CD4\(^+\) T cells. Math Biosci 114:81–125
Protter MH, Weinberger HF (1967) Maximum principles in differential equations. Prentice Hall, Englewood Cliffs
Redlinger R (1984) Existence theorems for semilinear parabolic systems with functionals. Nonlinear Anal TMA 8:667–682
Shu H, Wang L, Watmoughs J (2013) Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL immune responses. SIAM J Appl Math 73:1280–1302
Song X, Neumann A (2007) Global stability and periodic solution of the viral dynamics. J Math Anal Appl 329:281–297
Wang S, Feng X, He Y (2011) Global asymptotical properties for a diffused HBV infection model with CTL immune response and nonlinear incidence. Acta Math Sci 31:1959–1967
Wang X, Elaiw A, Song X (2012) Global properties of a delayed HIV infection model with CTL immune response. Appl Math Comput 218:9405–9414
Wang Y, Zhou Y, Brauer F, Heffernan JM (2013) Viral dynamics model with CTL immune response incorporating antiretroviral therapy. J Math Biol 67:901–934
Wang F, Huang Y, Zou X (2014) Global dynamics of a PDE in-host viral model. Appl Anal 93:2312–2329
Wang J, Pang J, Kuniya T, Enatsu Y (2014) Global threshold dynamics in a five-dimensional virus model with cell-mediated, humoral immune responses and distributed delays. Appl Math Comput 241:298–316
Wodarz D (2003) Hepatitis C virus dynamics and pathology: the role of CTL and antibody responses. J Gen Virol 84:1743–1750
Wu J (1996) Theory and applications of partial functional differential equations. Springer, NewYork
Xiang H, Feng L, Huo H (2013) Stability of the virus dynamics model with Beddington–DeAngelis functional response and delays. Appl Math Model 37:5414–5423
Xu R, Ma Z (2009) An HBV model with diffusion and time delay. J Theor Biol 257:499–509
Yan Y, Wang W (2012) Global stability of a five-dimensional model with immune responses and delay. Discrete Contin Dyn Syst B 17:401–416
Yang Y, Xu Y (2016) Global stability of a diffusive and delayed virus dynamics model with Beddington–DeAngelis incidence function and CTL immune response. Comput Math Appl 71:922–930
Yuan Z, Zou X (2013) Global threshold dynamics in an HIV virus model with nonlinear infection rate and distributed invasion and production delays. Math Biosci Eng 10:483–498
Zhang Y, Xu Z (2014) Dynamics of a diffusive HBV model with delayed Beddington–DeAngelis response. Nonlinear Anal RWA 15:118–139
Zhou X, Cui J (2011) Global stability of the viral dynamics with Crowley–Martin functional response. Bull Korean Math Soc 48:555–574
Zhu H, Zou X (2009) Dynamics of a HIV-1 infection model with cell-mediated immune response and intracellular delay. Discrete Contin Dyn Syst Ser B 12:511–524
Acknowledgements
This work was supported by the Natural Science Foundation of Shanxi University of Finance and Economics (Starting Fund for the Shanxi University of Finance and Economics doctoral graduates research, Grant No. Z18116), the National Natural Science Foundation of China (Grant nos. 11771373 and 11661076), the Natural Science Foundation of Xinjiang (Grant no. 2016D03022) and the Doctorial Subjects Foundation of the Ministry of Education of China (Grant no. 2013651110001).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Geraldo Diniz.
Rights and permissions
About this article

Cite this article
Miao, H., Teng, Z., Abdurahman, X. et al. Global stability of a diffusive and delayed virus infection model with general incidence function and adaptive immune response. Comp. Appl. Math. 37, 3780–3805 (2018). https://doi.org/10.1007/s40314-017-0543-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40314-017-0543-9
Keywords
- Virus infection model
- Delay
- Adaptive immune response
- Diffusion
- General incidence function
- Global stability