
Abstract
Agricultural intensification has led to the decrease of the diversity of wild and domestic pollinators. For instance, honeybees declined by 59 % in 61 years in the USA. About 35 % of major crops in the world depend on pollination services, and 3–8 % of world crop production will disappear without pollinators. Indeed, pollination provides several ecosystem services such as enabling crop and honey productions, regulating weeds and other cultural services. Agricultural intensification has also decreased weed diversity by about 50 % in 70 years because massive herbicide sprays have reduced the competition between weeds and crops. Nevertheless, weeds are at the basis of agricultural foodwebs, providing food to many living organisms. In particular, weeds provide flowers for pollinating insects including honey and wild bees. Here, we review the decline of weeds and bees. We discuss the effect of bees and pollination on crop production. We describe the complex interactions between bee pollinators, e.g. honey and wild bees, and landscape habitats such as crop fields and semi-natural elements. For that, we focus on spatial and temporal effects on flower resources. We show that weed abundance can reduce crop yields, thus inducing conflict with farmers. But weed abundance enhances regulating services by ensuring the survival of honeybees in the absence of oil seed crops. Weed abundance also enhances pollination services and, in turn, honey yield for the benefit of beekeepers. Weed abundance has also improved the survival of wild flora and the socio-cultural value of landscapes, a major request from the public. From those findings, we present a conceptual framework allowing to define ecological engineering options based upon ecosystem services of weeds and pollinators.
Similar content being viewed by others


Contents
-
1.
Introduction
-
2.
The Effect of agricultural intensification on weeds and bees
-
2.1.
Agriculture and biodiversity
-
2.2.
Weeds and crops: ecological and agricultural debts of use of herbicides
-
2.3.
Collapses in wild and domestic bee populations in farmlands
-
2.1.
-
3.
Interacting networks: weeds, bees, pollination and crop yield
-
3.1.
Floral resources required for wild and honey bees: weeds are limiting
-
3.2.
Crop pollination by honeybees and its effect on yield
-
3.3.
Are crops pollinated by wild bees?
-
3.1.
-
4.
Mutualistic and antagonistic interactions between ecosystem services and stakeholders
-
4.1.
Interactions between wild and domestic pollinators in Weed-Crop-Pollinator webs
-
4.2.
Trade-offs between ecosystem services resulting from pollination
-
4.3.
Assessing competition for spatial use of landscape features
-
4.1.
-
5.
Management options for finding the best compromise: ecological engineering versus ecological intensification
-
5.1.
Agro-ecological infrastructures enhancing pollination services
-
5.2.
Critical knowledge gaps in managing trade-offs
-
5.3.
Future avenues for research: the way forward for managing landscapes to reduce conflict between stakeholders
-
5.1.
-
6.
References
-
7.
Figures
1 Introduction
The world population is still increasing, and consequently, the demand for food is growing. Over the last 40 years, the demand for food has been met by increased crop yields obtained through generalized use of external inputs such as fertilizers and pesticides and improved technology. However, agricultural intensification has had a direct effect on water, soils, landscapes and biodiversity. There is clear evidence that agricultural intensification has led to the loss of biodiversity (Geiger et al. 2010; Robinson and Sutherland 2002; Kleijn and Sutherland 2003; Tscharntke et al. 2005), a decline in landscape diversity with the disappearance of semi-natural elements of all types (Benton et al. 2003) and ultimately a reduction in ecosystem functioning (Cardinale et al. 2012). Loss of biodiversity does not simply alter ecosystems per se, it also results in the partial or complete loss of services provided to humans by these ecosystems (Costanza et al. 1997; Chapin et al. 2000). The demand for provisioning services over the past century has been met at the expense of other services, including, paradoxically, those indirectly profitable to crops, such as pest regulation and crop pollination, or those which affect the sustainability of food production such as prevention of soil erosion and preservation of genetic resources. In parallel, greater public awareness has raised public expectations, especially with regard to public goods (water, air) and cultural services such as the conservation of flagship species (Tilman et al. 2002). In the context of global change and relative uncertainty (depletion of non-renewable resources such as phosphorus, unstable agricultural prices and imprecision of climatic scenarios), the future of intensive farming systems may, therefore, appear challenging: new systems are required to take account of changing economic and environmental aims (reduction of pesticides, biodiversity conservation, health) and must be adapted to changes in land use and climate, as well as being acceptable to all stakeholders. The trade-off between food production and biodiversity is so critical that it no longer concerns farmers alone, especially because biodiversity is believed to support most ecosystem services (Gabriel et al. 2013; Phalan et al. 2011).
It is, therefore, essential to understand the consequences of biodiversity loss on the provision of ecosystem services (Ehrlich and Ehrlich 1981), which requires account to be taken of existing constraints or trade-offs between different ecosystem services (Foley et al. 2005; Bennett et al. 2009). Therefore, to go beyond the sole aim of producing food, the challenge is to find the best compromise between crop yields and societal benefits, either for the farmers themselves (e.g. health) or for those who live in the countryside (e.g. public goods). Promoting biodiversity may improve the provision of a range of services and compensate for the reduction in the use of pesticides and herbicides (e.g. the French Plan Ecophyto) through better biological control, a common basis for both agro-ecology and ecological intensification (Bommarco et al. 2013; Gaba et al. 2014). It is, therefore, important to identify the processes that explain why various species improve ecosystem functioning and services, in particular crop production, either directly or indirectly.
The relationship between weed flora, i.e. the agricultural wild plants, and crops is complex: crop production may conflict with weed flora abundance (Fried et al. 2008; Meiss et al. 2008), but weed flora also plays a functional role by producing seeds for granivores, maintaining flagship species and providing flowers for insects (Biesmeijer et al. 2006; Marshall et al. 2003), in particular pollinating insects such as bees. Bees provide the bulk of pollination services (Winfree et al. 2011), especially in farmland habitats (Klein et al. 2007; Ollerton et al. 2011). Bees rely on floral resources for their diet, either mass flowering crops (the availability of which is reduced in time to their period of flowering) or weeds (which provide less flowers than crops, but more constantly, spatially and temporally). In addition, the dependency of bees to crops versus weeds depends on their taxonomic group, i.e. honeybee, bumblebees or wild bees (Rollin et al. 2013). Weed abundance may thus have several consequences on bees and pollination, and ultimately in the delivery of ecosystem services. A study of annual cropping systems can, therefore, provide useful information on the interplay between services provided to various, possibly competing, stakeholders. First weed abundance may reduce yields and thus be in conflict with farmers. Weed abundance can, however, enhance regulating services by maintaining pollinators and pollination services. Indeed pollinators may improve crop yields, at least for some annual crops (Carvalheiro et al. 2011). Therefore, weed abundance may also benefit farmers. Second, by ensuring pollination, weed abundance may increase honey yields hence being a benefit for beekeepers. Finally, weed abundance may benefit for the general public by ensuring the persistence and survival of wild flora and improving the socio-cultural value of landscapes.
This paper focuses on the key service of pollination and considers (i) the interplay between the pollination of crops and wild plants, i.e. weeds; (ii) the competition between wild and honeybees and their associated pollination function; and (iii) the resulting conflicts or common interests between all those who benefit from pollination in farmlands, i.e. farmers, beekeepers and general public. We first review the literature on the decline of weeds and bees resulting from agricultural intensification (Sect. 2). We then scan the literature on the effects of bees, both wild and domestic, and pollination on crop production (Sect. 3). This in-depth literature review is then used as a basis for a new and original conceptual framework linking two interrelated ecological networks, “weeds and wild bees” and “crops and honey bees”. The many different aspects of pollination in farmland systems, which improves yields for farmers and beekeepers as well as affecting broader societal services, are used to demonstrate how these two networks interact with the network of farmers, general public and beekeepers (Sect. 4). Finally, we discuss ecological engineering options based on weeds and pollinator bees for sustainable management of biotic interactions to provide provisioning, regulation and cultural services within an agricultural landscape (Sect. 5).
2 The effect of agricultural intensification on weeds and bees
2.1 Agriculture and biodiversity
European agricultural landscapes have changed significantly over the past decades, under the influence of the Common Agricultural Policy (CAP), which set out to increase food production (Godfray et al. 2010; Pe'er et al. 2014). Agriculture intensification resulted in an increase in cultivated areas, a decrease in the semi-natural features in the landscape (forests, hedgerows, permanent grasslands) and a decline in land use heterogeneity (Benton et al. 2003; see Fig. 1 for an illustration). Crop yields have been improved by generalized use of fertilizers and pesticides (Tilman et al. 2002). Evidence has shown that this has resulted in a major loss of biodiversity in farmland landscapes, with a decline in all taxa (Donald et al. 2001), affecting ecosystem functioning (Tilman et al. 2012) and the provision of services (Cardinale et al. 2012). This decline has affected not only threatened species and species dependent on conservation measures (Donald et al. 2001; Bretagnolle et al. 2011) but also ordinary biodiversity (Green et al. 2005) and has more recently been shown to have affected functional biodiversity (Tscharntke et al. 2005; Biesmeijer et al. 2006). In particular, the pollination service provided by several insect families is currently threatened because these insects are disappearing from agricultural landscapes (Potts et al. 2010a, b). In parallel with the decline in the bee population, the economic activity that relies on honeybees, beekeeping, has also declined significantly in many parts of Western Europe (Ellis et al. 2010). The decline in weeds and the decline in bees (notably honeybees), which play a substantial role in the agricultural economy, as described below, may appear to be unrelated, but as will be seen in Sec. 3, are actually strongly interconnected.

Landscape simplification through loss of crop diversity and increase of field size over one human generation as illustrated by these four photographs taken between 1958 and 2010 of the Long Term Ecological Research “Zone Atelier Plaine & Val de Sèvre”. Photos have been georeferenced from IGN (Institut National de l'Information Géographique et Forestière) aerial photography by French National Center of Scientific Research (CNRS) in Chizé
2.2 Weeds and crops: ecological and agricultural debts of use of herbicides
Weed flora in arable fields is often considered to be a major constraint for crop production (Milberg and Hallgren 2004) because weeds use some of the resources that are essential for crop growth, e.g. water, nitrogen, light, and often cause high financial losses (Oerke 2006). Since the mid-19th century, chemical weed control, i.e. the application of herbicides, has reduced yield losses and controlled weeds. In parallel with the use of herbicides, improved seed cleaning techniques (Spahillari et al. 1999), the limited set of crop species sown by farmers (Knox et al. 2011) resulting in reduced diversity of crops, the loss of traditional crops such as flax (Mirek 1976) and the increasing application of mineral fertilizers (Robinson and Sutherland 2002), has caused a significant decline in the diversity of arable plants throughout Europe (Andreasen and Streibig 2011; Sutcliffe and Kay 2000; Baessler and Klotz 2006; Hyvonen 2007; Fried et al. 2009a, 2012; Storkey et al. 2010). The long-term survey between the 1950s/1960s and 2009, of 392 fields in 10 different study areas in central and northern Germany, showed a significant loss of diversity locally with a mean loss of 65 % (from 24 species to only 7) (Meyer et al. 2013). A significant loss of diversity has also been observed in the overall weed seed bank over recent decades in number of European countries (Robinson and Sutherland 2002; Roberts 1981; Roberts and Feast 1973; Chancellor 1986), though recent changes in agricultural management since the 1990s (e.g. organic farming and reduced pesticide input) may have helped to slow down the decline of the arable flora in terms of species number (Richner et al. 2015). The characteristic species or the threatened arable weeds are, however, still in decline (Richner et al. 2015).
The response of weeds to agricultural intensification has been associated with a decrease in the abundance of many species, even to the point of extinction, and the dominance of a small number of species (Meyer et al. 2013). There have also been weed community shifts with the selection of groups of species. The frequency of archaeophytes has generally declined (Preston et al. 2004), while the trend for the frequency of neophytes is less clear (some studies have reported an increase (Lososova and Simonova 2008) and others have shown a decrease (Meyer et al. 2013)). Species with a particular combination of traits have flourished as a result of the significant changes in crop rotation and increasing herbicide pressure (Storkey et al. 2010; Fried et al. 2012). Winter wheat has allowed the increase of small weeds, with fairly light seeds, that can germinate over a long-time frame during the growing period (Fried et al. 2012). There has also been an increase in the abundance of species with traits associated with the ability to escape herbicides either bylate germination (Fried et al. 2012) or by the development of resistance (e.g. Alopecurus myosuroides; Delye et al. 2007, 2010). The marked decline in arable weed diversity has also caused a decline in the functional biodiversity of agro-ecosystems. Most of the weed species that are known to be very important for farmland birds or insects (Marshall et al. 2003; Storkey 2006; Eraud et al. 2015), i.e. Chenopodium album, Fallopia convolvulus, Polygonum aviculare, Polygonum persicaria, Sinapis arvensis and Stellaria media, have decreased significantly over the last 30 years (Fried et al. 2009b).
Intensive weed management strategies are now questioned because of their harmful effects on the environment and biodiversity and also because of the rapid worldwide spread of herbicide resistance (Jasieniuk 1996; Neve and Vila-Aiub 2009; Manalil et al. 2011). Growers currently use a lower number of herbicides (Delye et al. 2013) increasing the selection pressure on weeds and hence increasing the number of resistant weeds which can survive the application of herbicides by a variety of mechanisms (Powles and Yu 2010; Beckie and Tardif 2012). Since non-chemical weeding can only be as effective as herbicides by combining several different methods (Bastiaans et al. 2008), this may lead to changes in the composition and structure of the arable plant community, probably increasing species abundance and diversity.
2.3 Collapses in wild and domestic bee populations in farmlands
There are several taxonomic groups of bee, including honeybees Apis mellifera, bumblebees (genus Bombus) and solitary bees (Michener 2007). In France for instance, there are about 35 species of bumblebee and approximately 950 additional species of wild, solitary bees (Kuhlmann et al. 2013; see Fig. 2 for some illustrations). All are currently on the decline in farmland landscapes (see Gonzalez-Varo et al. (2013) for a review): honeybees (VanEngelsdorp et al. 2008), bumblebees (Winfree et al. 2008; Cameron et al. 2011) and wild bees (Biesmeijer et al. 2006; Potts et al. 2010a, b). Honeybee populations are declining worldwide, in Europe (Potts et al. 2010b), Asia and Australia (Oldroyd and Nanork 2009) and North America (VanEngelsdorp et al. 2008; Ellis et al. 2010). In the latter, 59 % of honeybee hives disappeared in just 61 years. The decline in the wild bee populations over the past 120 years has been established using historic datasets (Burkle et al. 2013), and in Europe, 37–65 % of bee species are of conservation concern (Patiny et al. 2009). Many plant species, most of which are weeds, found in natural and semi-natural habitats are food resources for honeybees (Requier et al. 2015) as well as wild, solitary bees. The loss of natural habitats is, therefore, regarded as the primary cause of the decline of wild pollinators with both a decrease in nesting and foraging sites (Kremen et al. 2002; Winfree et al. 2009; Ricketts et al. 2008; Steffan-Dewenter et al. 2002; Hendrickx et al. 2007).

Various species of bees found in intensive farmland landscapes: a Bombus lapidarius; b Andraena spp. (male); c Eucera nigrescens and d honeybee Apis mellifera. All photos were made by Orianne Rollin except d (ACTA - “Le réseau des instituts des filières animales et végétales“)
The situation regarding honeybees, however, is more complex than for wild bees, for at least two reasons. First, the honeybee populations in farmland landscapes in Europe consist in domesticated, introduced honeybees for beekeeping, as well as feral or wild honeybees, i.e. the ancestral species or subspecies. Though it is often thought that the wild/feral populations are now virtually extinct in Western Europe (Jaffe et al. 2010) or have hybridized with introduced subspecies (in Germany for instance A. mellifera hybridized with A. carnica: Moritz 1991), the situation may be more complex and the feral population should probably best be seen as of unknown size, further interacting, through the large groups of males that gather as drones from many hives and feral colonies, with the domesticated bee population (Jaffe et al. 2010). Second, although honeybees and wild bees are both on the decline, honeybees are subject to a specific syndrome known as the colony collapse disorder (CCD). This was first described in North America in 2007 (Cox-Foster et al. 2007; VanEngelsdorp et al. 2009, 2008; Oldroyd 2007) but also reached Europe (possibly since 1998). CCD is characterized by an abnormal increase in mortality over the winter, commonly 25 % of hives but as high as 50 % of hives. Other associated symptoms (Aubert 2002; Saddier 2008; Winfree et al. 2009; Potts et al. 2010a; Neumann and Carreck 2010) result in both colony loss and loss of adult workers in spring and summer (VanEngelsdorp and Meixner 2010). Once considered as a syndrome explaining bee decline, especially winter loss, recent work eventually led to a new paradigm: bee decline results from a complex set of interacting drivers (Ratnieks and Carreck 2010; Di Pasquale et al. 2013; Alaux et al. 2010; VanEngelsdorp et al. 2009;Potts et al. 2010a), including honeybee husbandry practices (e.g. Varroa is an invasive species introduced from Asia through beekeeping practices: van Dooremalen et al. 2012). The drivers involved include pathogens (Whitehorn et al. 2013), environmental factors (e.g. flowers, pesticides), ecto-parasites (Meeus et al. 2011) and genetic factors (Cameron et al. 2011). Despite the acknowledged role of pathogens (especially Varroa: Ellis et al. 2010; Potts et al. 2010b) and honeybee management (Le Conte et al. 2010), environmental factors such as land use change (Steffan-Dewenter et al. 2002; Kremen et al. 2002), and pesticides, especially insecticides which have both lethal and sub-lethal effects (Henry et al. 2012; Desneux et al. 2007), and herbicides which further reduce floral resources (Gabriel and Tscharntke 2007) also play an important role.
3 Interacting networks: weeds, bees, pollination and crop yield
As we shall see now, both wild and honeybees rely to a considerable extent on flowers for survival and breeding (although some wild bees parasitize other bees: see Michener 2007). In intensive farmland landscapes, flowers are provided by mass-flowering crops, such as oilseed crops (rapeseed, sunflower) and to a lesser extent, legumes, by hedgerow plants, and by weed communities (in both crops and grasslands), especially those found in annual crops. This has been shown by a long-term study of the diet and foraging behaviour of honeybees and wild bees in intensive cereal landscapes (Rollin et al. 2013; Odoux et al. 2012; Requier et al. 2015).
3.1 Floral resources required for wild and honeybees: weeds are limiting
Bees depend exclusively on flowers for pollen and nectar supplies (although they also require other resources, such as water and nesting sites). Pollen is used for brood development (Brodschneider and Crailsheim 2010), since it contains proteins, fats, mineral salts, amino acids and vitamins (Campos et al. 2008; Manning 2001; Haydak 1970). Honeybees only store tiny quantities of pollen in the colony because it deteriorates rapidly (Pernal and Currie 2000). Nectar is used for the daily energy intake. It is the metabolic precursor for beeswax and is processed into honey, which is their food reserve for overwintering as bees do not forage in winter (at least not in temperate zones, Aronne et al. (2012)). Little information is available on the diet of honeybees in farmland landscapes, especially in intensive cereal systems (Requier et al. 2015). Honeybee colonies (in apiaries) comprise about 30,000 adults and a similar number of larvae and need a continuous influx of pollen since it is not stored, whereas the need for nectar is more seasonal (though a large quantity is still required). In cereal systems, mass-flowering crops, especially oilseed crops, provide the bulk of floral resources when they are in bloom. The main crops for honeybees in such systems include maize (for pollen only), sunflowers (both for pollen and nectar) and rapeseed (mainly for nectar) (Decourtye et al. 2011). Maize provides the highest amount of pollen (Charriere et al. 2011; Vaissiere and Vinson 1994; Odoux et al. 2004), while sunflowers have a lower quality pollen (Schmidt et al. 1995). However, mass-flowering crops provide a valuable source of food during a short period of time (Morandin and Winston 2006). As the blooming periods of oilseed crops (especially rapeseed and sunflower) are short and separated by a gap of about 2 months, honeybees have to rely on other resources, wild flowers, which are not usually as abundant and dense as the crops. Several studies have shown that honeybees then shift their attention to the weeds found in crops, as well as ligneous species, and may visit between one and two hundred species (around to one hundred species, e.g. Coffey and Breen 1997; Odoux et al. 2012; Pernal and Currie 2001). They also visit plant species from semi-natural habitats of forest fragments, although to a much lesser extent (Odoux et al. 2012; 2014), but they account for a minor part of pollen resources (Requier et al. 2015).
Although honeybees apparently select which flowers they visit (Aronne et al. 2012), they are typically considered as generalist foragers which use a wide variety of plants to satisfy their needs (Seeley and Visscher 1985). Their generalist behaviour seems to correspond to the various needs of the honeybee colony for amino acids, fats, vitamins and minerals, and since the quantities of these nutrients vary between different plants, honeybees forage on a wide variety of species in order to avoid a deficiency of any particular nutrient (Weiner et al. 2010). Aronne et al. (2012) further showed that honeybees usually select plants for their pollen content rather than for their nectar, which is not surprising given that pollen is not stored in hives. Indeed, many studies have shown that honeybee colonies perform better when pollen resources are varied (Alaux et al. 2010; Mayack and Naug 2009).
There is a strong correlation between plant diversity and wild bee diversity (e.g. Holzschuh et al. 2008; Potts et al. 2003; Biesmeijer et al. 2006; Hopwood 2008). Unlike honeybees, wild and solitary bees are not necessarily generalists, some species being specialist flower foragers (though they tend to pollinate a small number of families or genera rather a single species (Rollin et al. 2013)). Wild bees are, therefore, more selective in their choice of flower and usually forage on a limited diversity of plants, and some wild bees may pollinate only one plant species. Weeds are frequently visited by wild, solitary bees: cornflowers, for instance, attract tens of different species (Rollin 2013). Bumblebees occupy an intermediate position between honeybees and solitary wild bees (Rollin et al. 2013): they do not forage to any great extent in either semi-natural habitats or in oilseed crops: they are found with fairly uniform low abundance. Weeds are also a limiting factor for bumblebees: after the mass-flowering of rapeseed simplified landscapes contain significantly fewer flowers than complex landscapes. One study reported a sharp decline in bumblebees in simplified landscapes in late July with bumblebee abundance being positively correlated with the availability of herbaceous flowers (among other factors), suggesting that, in simplified landscapes, bumblebee abundance is limited by floral resources (Persson and Smith 2013).
Overall, therefore, at least in intensive cereal farming systems, (i) there is significant ecological segregation between wild and honeybees (Rollin et al. 2013; Carvalheiro et al. 2011), (ii) both wild and honeybees depend on weeds. Honeybees tend to be found more frequently and in greater abundance in mass-flowering oilseed crops whereas wild, solitary bees are more abundant in semi-natural features, grasslands and grassy strips. Bumblebees are found in both rapeseed and sunflower habitats but also in other habitats (oilseed rape and sunflower, Rollin et al. 2013). For both wild and honeybees, weeds are a limiting resource, although for different reasons: quantitatively for honeybees, especially between the mass-flowering periods of rapeseed and sunflower and qualitatively for the more selective wild bee foragers. The decline in wild bee diversity is, therefore, strongly correlated with the decline in weeds (and, more generally, wild flowers) (Biesmeijer et al. 2006; Carvell et al. 2006), though it is unclear which is the cause and which is the effect.
3.2 Crop pollination by honeybees and its effect on yield
There is strong evidence that insect-mediated pollination is declining worldwide (Gonzalez-Varo et al. 2013), owing to pressures related to global change. Although there has been no precise evaluation of the relative importance of the taxa that actually pollinate flowers, bees are the most important worldwide, partly because they are the most frequent visitors of flowers (Winfree et al. 2011; Neff and Simpson 1993). Pollination by insects is vital for both crops and wild plants (Ollerton et al. 2011): 84 % of European cultivated plants depend on insect pollination (Williams 1994) and 70 % of 57 crops grown worldwide (Klein et al. 2007). Wild and honeybees are the main pollinators of these crops (Klein et al. 2007; Garibaldi et al. 2013; Rader et al. 2012), and their pollination service to crops has been valued at 153 billion euros annually worldwide (Gallai et al. 2009), and around 22 billion euros annually for Europe (Gallai et al. 2009) and more than 18 billion dollars in the USA (Mader et al. 2011). A more recent study (Lautenbach et al. 2012) of 60 crops suggested 266 billion euros per year worldwide, and an estimated 3–8 % of world crop production could be lost in the absence of pollinators (Aizen et al. 2009).
Experimental work has been carried out to quantify pollination by bees of either crops or some wild flowers (see Fig. 3 for examples of phytometer experiments with crop plants and weeds). The dependence of crops on pollination, in particular by bees, varies considerably on the type of crop: wheat and maize, for instance, do not require any pollination, whereas many types of fruit tree rely on pollination (Klein et al. 2007). There is also considerable variation among annual crops, some depending on pollination, though the relative extent of wind versus insect pollination on yield remains to be determined (Hayter and Cresswell 2006). For instance, there is evidence that bees increase the yield from sunflowers (Carvalheiro et al. 2011). Rapeseed can be pollinated by wind, insects and autogamy (Delaplane and Mayer 2000; Garratt et al. 2014), and measurements of the pollination rate by bees and its effect on yield in cereal systems provided contradictory results (e.g. Garratt et al. (2014) for rapeseed), partly because the effect of bee pollination on yield is cultivar dependent (Steffan-Dewenter 2003; Morandin and Winston 2005). For rapeseed, Bommarco et al. (2012) found that there was an 18 % increase in seed weight when pollinators had access to rapeseed flowers (see also Morandin and Winston (2005)) but found no effect on the seed fructification rate. However, Stanley et al. (2013) found an increase of over 30 % in both seed production and seed weight. Other studies found no detectable effect, rapeseed being mass-pollinated by wind (review in Hayter and Cresswell (2006)). Furthermore, some rapeseed flower traits, such as pollen production, are more attractive to some pollinating insects than to others (Holzschuh et al. 2013; Stanley et al. 2013). However, although crop pollination by bees may significantly increase crop production, bee abundance might be too low to pollinate crops (Breeze et al. 2014).

Example of a phytometer experiment conducted on cornflower in winter barley in the Long Term Ecological Research “Zone Atelier Plaine & Val de Sèvre”. Note that only some flowers per cornflower individuals are bagged, in order to get control flowers
3.3 Are crops pollinated by wild bees?
Recent studies have shown that bees are affected both by the quantity of semi-natural features in the landscape and by the intensity of field management. Moreover, many studies, experimental or empirical, have shown that pollination of crops by bees depend on the landscape features, with greater pollination in landscapes with a higher density of semi-natural elements (Carvalheiro et al. 2010; Le Féon et al. 2013; Kennedy et al. 2013) although the effect also depends on the spatial scale (Steffan-Dewenter and Westphal 2008). Similarly, the pollination services provided to crops were shown to decrease with isolation from natural elements in the landscape, reducing both crop yields and their stability over time (Garibaldi et al. 2011; Chaplin-Kramer et al. 2013). In return, mass-flowering crops provide additional food resources and can increase the density of wild pollinators in adjacent habitats (Westphal et al. 2003).
While there is doubt about whether annual oilseed crops are pollinated by honeybees and, if so, to what extent, there is even greater discussion about the role of wild bees in crop pollination. Wild bees (including bumblebees) are certainly more efficient than honeybees in pollinating some perennial crops (e.g. raspberries and blueberries: Willmer et al. 1994; Javorek et al. 2002) and fruit (e.g. strawberries: Klatt et al. 2014), but little research has been carried out to determine their role in annual crop pollination. However, wild bees do forage on crops, and there is evidence that a diverse community of wild bees can increase crop production in some cases (e.g. Carvalheiro et al. 2011; Garibaldi et al. 2013). Diverse pollinator communities have been found to improve pollination services more than poorer communities (Klein et al. 2003; Hoehn et al. 2008; Klein et al. 2012). Indeed, wild bees and honeybees may have mutualistic effects on pollination: the behavioural interactions with wild bees may force honeybees to move on to another plant, which, in particular for crops which have male and female plants, may improve pollination and crop production (Greenleaf and Kremen 2006; Brittain et al. 2013). There does not, however, appear to be a general rule: in some cases, wild bees have been reported to pollinate crops without any effect on honeybees (Garibaldi et al. 2013), in other wild bees outcompete honeybees in crop pollination (Rader et al. 2012) and for some sunflower varieties, wild bees did not seem to complement or compensate for a lack of pollination by honeybees (Pisanty et al. 2014). It also remains to be established whether it is the bee diversity rather than their abundance or even presence (including honeybees) that drives crop pollination and yield.
4 Mutualistic and antagonistic interactions between ecosystem services and stakeholders
This in-depth literature review (Sects. 2 and 3) has shown that, in farmland landscapes, two a priori different networks involving bee pollinators and landscape habitats would seem to co-exist independently of each other: the ‘semi-natural elements - wild bees’ on the one hand and the ‘crops - honeybees’ on the other. This section describes the complex interactions between these networks, both in terms of dependency on limiting resources and by their pollination function, and suggests that the resulting delivery of services provided by bees goes far beyond crop pollination and honey production, with the various stakeholders competing for space within the landscape in conflicting or synergistic networks.
4.1 Interactions between wild and domestic pollinators in weed-crop-pollinator webs
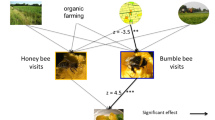
Honeybees mainly visit on oilseed crops, whereas most wild bees forage mainly on semi-natural features (Potts et al. 2003; Steffan-Dewenter and Tscharntke 2001; Rollin 2013). Given this marked segregation in habitat utilization between honeybees and wild bees in farmland landscapes, crops are pollinated mainly by honeybees and, to a lesser extent, bumblebees (see Sect. 3 above). However, this simplistic view is challenged by two important facts. First, by pollinating weeds, wild bees may interact indirectly with honeybees by providing floral ressources, since in the period between mass-flowering of oilseed crops in cereal systems, honeybees forage almost exclusively on weeds (Morandin and Winston 2006; Requier et al. 2015). Indeed, wild flowers strongly depend on pollinating insects (bees, butterflies and hoverflies) for reproduction: 78–94 % of flowering species rely on pollination (Ollerton et al. 2011; Winfree et al. 2011). Though honeybees actively collect pollen from wild flowers, the latter are pollinated by wild bees (Biesmeijer et al. 2006; Potts et al. 2010a), including rare weed species (Gibson et al. 2006), and so help to ensure the conservation of floral biodiversity in the landscape (Biesmeijer et al. 2006). These two networks are thus interconnected through the weed compartment, on which they both depend as a limiting resource, either in space (wild bees) or time (honey bees). Second, wild and domesticated bees also interact directly with each other in two different ways: competition by depletion (of resources) and competition by behavioural interference. Wild and honeybees may compete for floral resources, at least when honeybees forage on weeds between the mass-flowering periods of oilseed crops. Given the very large size of honeybee colonies, a spill-over effect has been suggested by Rollin (2013) and Blitzer et al. (2012), but this has not yet been quantified accurately. Honeybees and bumblebees may also interact through their foraging behaviour (Riedinger et al. 2014). Whether honeybees are antagonistic to other bee species was assessed by comparing the flower visitation rates of honeybees and wild bees (including bumblebees). Nielsen et al. (2012) found a positive correlation between honeybee and bumblebee visitation rates, whereas the relationship between honeybees and solitary bees varied from being positive, negative or insignificant, depending on the plant species. Interaction between honey and wild bees be even more complex: analysing the effects of mass-flowering rapeseed (a flower visited by short-tongued pollinators) on the relative abundance of long-tongued and short-tongued bumblebees, Diekötter et al. (2010) found that the density of long-tongued bumblebees visiting long-tubed flowers decreased as the amount of rapeseed increased, leading to a distortion in plant–pollinator interactions.
Honey and wild bees, therefore, interact in trophic networks in a rather complex manner and the type of trophic interaction (antagonistic or mutualistic) depends on the season (see Fig. 4). More importantly, these two interacting networks further affect crop pollination, depending on weed abundance. Carvalheiro et al. (2011) showed for instance that, when weeds are present in sufficient numbers or when weeds are growing sufficiently close to crops, the wild bee community is more abundant, pushing honeybees away to pollinate crop flowers which in turn increase crop production. Indeed, the presence of wild bees on flowers induces behavioural interference with honeybees, which, when disturbed, forage on other flowers and hence help to cross pollinate individual crop plants, increasing the success of pollination, as demonstrated in sunflower (Greenleaf and Kremen 2006). Similarly, in orchards with non-Apis bees, the foraging behaviour of honeybees changed and the visit from a single honeybee was shown to be more effective at pollination than in orchards without non-Apis bees (Brittain et al. 2013). Since increasing pollinator diversity may improve pollination services, through interactions between species that change the foraging behaviour, the increased diversity may enhance the functional quality of a dominant pollinator species and increase the pollination effectiveness of the individual species of pollinator. Consequently, as agricultural production relies to a large extent on pollination, increasing pollinator diversity is one way of sustainably improving pollinator-dependent crop yields.

Seasonal patterns of foraging for wild and honeybees. The grey rectangles indicate the mass-flowering blooming season for rapeseed and sunflower. The vertical lines delineate the temporal categories (called “month”) of seasonal patterns. Month periods are also indicated. Photos illustrate the habitat, i.e. rapeseed, winter wheat, sunflower and semi-natural habitats, in which wild and honeybees forage
4.2 Trade-offs between ecosystem services resulting from pollination
Pollination is involved in various agro-ecological networks, and therefore, there are various stakeholders with mutualistic or antagonistic interactions. First, crop pollination by honeybees increases the crop yield for some annual crops such as rapeseed and sunflower (see Sect. 3 above), providing provisioning services by increasing financial benefits for farmers. Honeybees also directly support beekeeping (Ellis et al. 2010). The survival of honeybees relies on weeds when floral resources are scarce between the mass-flowering periods of crops such as rapeseed and sunflower: poppies alone can account for up to 60 % of pollen resources for hives during late spring (Requier et al. 2015). These species of weed are usually found in winter cereals, and in smaller abundance in semi-natural habitats, and are often considered by farmers as pests that compete with the crops and can severely affect yields and revenue. Consequently, increasing pollination, i.e. by increasing weeds, to improve rapeseed and sunflower yields may significantly reduce winter cereal yields, i.e. due to the competition for resources between weeds and the crop, leading to conflicts between cereal farmers (within and between farms) and between farmers and beekeepers. Second, pollination provides regulating services as it governs the population of many species involved in functional biodiversity. Weeds ensure the maintenance of wild bees to whom they provide pollen and nectar, and in turn, pollination of weeds by wild bees ensures the reproduction, i.e. the persistence, of several weed species. Consequently, it impacts all species that depend on weeds, i.e. many birds and insects that control pest invertebrates (Marshall et al. 2003). Third, pollination is a cultural service since many species of bees are conservation-dependent and/or pollinate-threatened arable plant species (e.g. cornflowers) that have conservational and aesthetic value (Clergue et al. 2005). Indeed, the decline in pollinators has led to the loss of wild plants which depended on insect pollination (Biesmeijer et al. 2006; Carvell et al. 2006). Moreover, the presence of wild bees in a landscape depends on the proportion of semi-natural habitats which provide shelter and habitats for these insects but may also have aesthetic value themselves. For all these reasons, pollination and pollinators such as wild and honeybees, hoverflies and butterflies in farmland landscapes are involved in the provision of many different ecosystem services and may be sources of conflict between stakeholders.
Thus, besides being an important ecosystem service, pollination in a farmland landscape is essential to a wide range of stakeholders such as beekeepers, farmers and the general public (Fig. 5). Beekeepers depend upon pollination services and benefit from the presence of habitats with flowers, such as rapeseed and sunflower crops, as well as winter cereals and semi-natural habitats that shelter weeds. However, beekeepers are not directly involved in the management of agricultural landscapes: it is rather cereal farmers who define land use and have to manage the trade-offs. The provisioning services provided by winter cereals could be increased by growing more of this crop with an intensive weed control to reduce crop yield losses, hence reducing weed availability and replacing semi-natural habitats by crop land. Increasing winter cereal yields may, however, in turn reduce rapeseed and sunflower yields, by reducing pollination due to the decline of bees between mass-flowering periods. The public on the other hand depends directly upon pollination for provisioning (food), regulation and cultural services. Their income may come from specific ecosystem services, such as nature tourism, which depend on the proportion of semi-natural habitats and weeds in the landscape (Wratten et al. 2012). More weeds and more bees may lead to higher profits for beekeepers or social value for the public, whereas fewer weeds may be better for cereal farmers. Management strategies for multiple ecosystem services may have opposite effects (reducing one service while enhancing another), leading to trade-offs, where one service is increased at the expense of another, and competition for use of landscape features.

A summarised view of the interacting pollinator webs and antagonistic stakeholder network. Antagonistic and synergic relationships are indicated by red and green arrows, respectively. The relationship between honey and wild bees which remains to be established is indicated by the orange arrow. This figure shows that even if each stakeholder interest seems to be independent from the other, they are indeed related through the plant-pollinator trophic network
4.3 Assessing competition for spatial use of landscape features
Land use changes can alter the spatial structure of wild plant populations, which may in turn affect the attractiveness of flower aggregations to different groups of pollinators at different spatial scales (Nielsen et al. 2012). Bees may be affected both locally by farm management and by the surrounding landscape. Modelling the relative effects of landscape composition (nesting and floral resources), landscape configuration (shape, connectivity) and farm management (e.g. organic farming) on wild bee abundance and richness for 39 cropsystems, Kennedy et al. (2013) found that bee abundance and richness were higher in diversified, organically managed fields and in landscapes with more high-quality habitats, i.e. suitable habitats for nesting and nearby floral resource. Pollinator persistence depends, therefore, on maintaining high-quality habitats around farms and on local management practices that may offset the impact of intensive monoculture (Deguines et al. 2014). There is a correlation between landscape complexity (including semi-natural features), floral diversity and availability and bee diversity (Duelli and Obrist 2003; Le Féon et al. 2010). Furthermore, crop pollination relies on honeybee abundance which, to some extent, relies on weed abundance and diversity and, to a lesser extent, on wild bee diversity, which in turn relies on semi-natural features within the landscape (Steffan-Dewenter and Westphal 2008). However, the spatial scale at which these features affect pollinator abundance and pollination function has not been fully established (Benjamin et al. 2014).
The importance of the spatial and temporal scales used for the analysis and valuation of ecosystem services has been widely recognized in both economics and ecology (Thies et al. 2005; Kennedy et al. 2013). The spatial scale is particularly critical, since most land use management is determined at farm or even field scale, and very rarely at landscape scale (although it may be coordinated between adjacent farms), which means that the management may not be optimum. Furthermore, many different services are provided by many different organisms, even for pollination, and so, many different optimum spatial scales should be expected. These have been shown to vary with body size in wild bees (Benjamin et al. 2014). The process of pollination takes place across the landscape scale. Honeybees can travel long distances in search of desirable floral rewards and tend to forage within 2 km of their hives if there are attractive floral resources in the vicinity (Osborne et al. 2001), although foraging distances of up to 6 km have been suggested (Beekman and Ratnieks 2000). Consequently, the main factor affecting honeybee pollination is the temporal dynamics of flowering which must ensure the presence of floral resources, i.e. weeds during the pollination period. Therefore, between crop mass-flowering, honeybees are affected by the proportion of semi-natural habitats in the landscape rather than by their configuration (i.e. distribution within the landscape). Flower strips, adjacent to pollination-dependent crops, have proved to be effective in increasing yields (e.g. Blaauw and Isaacs 2014), but these reduce the area available for crops. The foraging distance of wild pollinators, however, is usually limited to a few dozen of metres (except for bumble bees and solitary carpenter bees, e.g. Rao and Strange 2012). Both the presence and the distribution of semi-natural habitats affect the diversity of wild pollinators and are likely to have an indirect effect on honeybees by increasing weed diversity and abundance. However, since wild and honeybees forage on similar resources between the mass-flowering periods of rapeseed and sunflower, pollinators may be expected to compete for resources. This raises the question of the extent to which favouring honeybees for crop or honey production might be detrimental to wild bees. This is an important issue since reducing wild bee communities may reduce the abundance of weeds that are not pollinated by honeybees and hence reduce ultimately the abundance of honeybees. Understanding the dynamics of this complex network and how the spatio-temporal composition of the landscape affects relationships within this network remains a challenge for agro-ecosystem management.
5 Management options for finding the best compromise: ecological engineering versus ecological intensification
Section 4 proposed a conceptual framework for enabling agriculture to benefit from weed functionalities to increase crop yield through pollination (see also Carvalheiro et al. 2011) as well as providing regulation and cultural services (Wratten et al. 2012). Increasing weed abundance may benefit beekeepers and the general public but may have an adverse effect for cereal farmers. There is also a trade-off between semi-natural features and cultivated areas. Making these trade-offs explicit should be a core aim of ecosystem assessments. However, because weeds can be both beneficial and harmful and the ecological, agronomic, socio-economic processes involved are extremely complex, diverse and interact at different spatial scales, designing agro-ecological cropping systems that ensure and maximise these services is far from straightforward. Moreover, because of the apparently conflicting aims of the stakeholders involved, only management at landscape scale can effectively resolve trade-offs between ecosystem services in the long term. The benefits to the various stakeholders must be considered at landscape or regional scale to assess the outcomes in terms of total food production and economic or societal benefits. This section analyses ecological engineering options for agro-ecosystem management, from local field scale to landscape scale, and describes the remaining challenges for ecosystem service research and management.
Many management strategies and policy initiatives have been set up in recent years, especially within intensively farmed landscapes, to halt the decline in bee abundance by increasing the availability of pollen and nectar resources (review in Decourtye et al. 2011; Holzschuh et al. 2010). These include reducing the use of pesticides, changing cropping systems, introducing Agri-Environment Schemes (AES) or flower-rich strips and managing or improving semi-natural features at landscape scale. Most of these measures, however, may have a significant effect on crop production, either directly by reducing yields or indirectly by reducing the area used for the annual crops that provide the highest income (Ghazoul 2007). We give a brief review of agricultural practices and agro-ecological infrastructures that are favourable for bees (including landscape management by stakeholders other than farmers) and discuss the trade-offs between these measures and crop production.
5.1 Agro-ecological infrastructures enhancing pollination services
Since intensive agriculture (and land use changes) has led to a worldwide decline in bees, less intensively managed agriculture should increase bee populations (see Winfree et al. 2011 for a meta-analysis). Pesticides (including insecticides, fungicides and herbicides) have been shown to have an adverse effect on both wild and honeybees (Desneux et al. 2007), and so, reducing the application of pesticides would help to conserve the species richness as well as abundance of bees. At field level, cultivating melliferous crops will increase the carrying capacity of the landscape (Decourtye et al. 2011; Rollin et al. 2013). There is also some evidence that reducing field size may improve pollination, at least by wild bees (Isaacs and Kirk 2010) which have short foraging distances. The most significant changes in agricultural practices are to be found in organic farming: waggle dance studies showed that honeybees had a significant preference for organically managed and AES land (Couvillon et al. 2014). Organic farming was also found to be beneficial for cavity-nesting bees, wasps and their parasitoids (Holzschuh et al. 2010). Agroforestry was shown (at least in tropical systems) to sustain larger wild bee communities than primary forest or agricultural land (Hoehn et al. 2010), although there is no documented evidence for temperate cereal agroforestry systems.
Alternatively, the conservation of semi-natural habitats and the use of flower strips (Whittingham et al. 2007) have been shown to restore wild pollinator populations by increasing resources, an indirect but increasingly clear indication that the availability of floral resources may act as a limiting factor for bee populations. Recent studies have shown that AES enhances the abundance and species richness of bees (Kleijn and Sutherland 2003; Kleijn et al. 2004). All types of semi-natural or natural features within the landscape are critical for the survival of bees and the pollination service. There are also several examples of actions taken by private or public bodies that have increased pollination services through better management and restoration of semi-natural features. For instance, a study of the effects of garden habitats on wild bees by Samnegard et al. (2011) found that a native plant (Campanula persicifolia) was pollinated to a greater extent when semi-natural features were present or close-by. When semi-natural habitats are too small and/or dispersed in intensively farmed agricultural landscapes, they cannot support viable populations of butterflies and bumblebees, which rely on dispersal from larger patches of semi-natural grassland (Ockinger and Smith 2007). Wild plants in field margins and hedgerows are important sources of alternative forage for pollinating insects, even during mass-flowering, and conservation of field margins and hedgerows, which provide alternative foraging habitats for pollinators, appears to be essential for the provision of pollination services to both crops and wild plants (Garibaldi et al. 2011; Stanley and Stout 2014).
Overall, therefore, such measures could increase pollinator populations, and ultimately pollination, by enhancing floral resources. Such strategies also provide secondary benefits for farmers and the surrounding landscape (Wratten et al. 2012), such as pest regulation, soil protection, improved water quality and more attractive landscapes.
5.2 Critical knowledge gaps in managing trade-offs
The management options described above, however, incur costs, either for farmers (directly or indirectly) or for other stakeholders. Restoring plant diversity in farmland landscapes is advocated in order to increase pollinators, but farmers rarely adopt such practices and instead kill weeds to the point of extinction (Ghazoul 2007). As yet, no study has accurately quantified the two aspects of the trade-offs between ecosystem services and stakeholders (as described in Sect. 4.2) and the only studies so far published explored only parts of the compromise (Steffan-Dewenter and Westphal 2008). In order to be fully efficient and applicable, these alternative management strategies should, in addition to maintaining yields or at least farmers’ incomes, (i) enable the maintenance of weeds at landscape scale while controlling competition on crop yield, (ii) provide semi-natural features in the landscape to allow the reproduction of wild bees, (iii) limit potential competition between honey and wild bees and (iv) limit potential competition between stakeholders for spatial use of landscape features. One way to meet these challenges is to set up spatial configurations that would allow for trade-offs between crops (rapeseed, winter cereal, sunflower) and semi-natural features along a land sparing/land sharing continuum (Green et al. 2005; O'Farrell and Anderson 2010; Tscharntke et al. 2012). Assessing the value of these strategies requires explicit spatial modelling such as dynamic Bayesian networks and Markov random fields to represent stochastic spatial interactions between any structure in the spatial interaction network (regular or not) (Tixier et al. 2013). Such a model coupled with decision-making models (e.g. Factored Markov Decision Processes, Tixier et al. 2013) could provide an effective means for analyzing the effect of proposed management decisions on ecosystem services resulting from different decisions related to stakeholders’ aims. This requires a better understanding of the ecological processes and a precise knowledge of the relationships between biodiversity, ecosystem services and crop production. Stakeholders’ aims also need to be precisely defined in terms of criteria (acceptable thresholds). However, the information required is not available, and the links between biodiversity and ecosystem services remain to be determined.
The framework proposed above was based on the goods and services that ecosystems can provide to stakeholders and the role that biodiversity, i.e. bees and weeds, may play in producing them. However, as described in Sect. 2, intensive agriculture has resulted in the significant decline in the diversity of bees and weeds. It is not easy to reverse this trend. For instance, fertilizers decrease diversity and it will take a long time for plant diversity to recover from sustained high rates of N enrichment (hysteresis) and it may not recover simply by reducing N input (irreversibility) (Isbell et al. 2013). A weed regulation method is required to ensure that weeds are sufficiently abundant for pollinators but not too abundant to reduce crop yield. The literature provides increasing evidence that post-dispersal seed predation may substantially contribute to reducing the weed seed bank (Westerman et al. 2008; White et al. 2007), hence providing biological control of weeds. However, although both vertebrates (birds and small mammals) and invertebrates have been reported as weed seed predators (O'Rourke 2006; Evans et al. 2011), few studies have assessed their relative capacity to regulate weed populations significantly (Whelan et al. 2008) and this is still a matter for discussion. There are significant gaps in our understanding of the processes that govern seed removal by weed seed predators and the consequences on weed assembly which limit the development of efficient weed-control strategies in agro-ecosystems.
5.3 Future avenues for research: the way forward for managing landscapes to reduce conflict between stakeholders
This chapter has discussed a conceptual framework, based on a literature review, showing the potentially key, counterintuitive, role of weeds in increasing crop production through the regulation of both honey and wild bees. Although this framework is appealing, it also raises many important questions which still need to be answered. Overall, there is still a lack of basic information to guide sound management decisions at landscape scale. There is some evidence that the diversity of wild bees and abundance of honeybees are positively correlated with the abundance and diversity of weeds at field and landscape scales (e.g. Winfree et al. 2011) but there are still many unanswered questions. Although crop production appears to be related to wild bee diversity and honeybee abundance, there is still no quantitative estimate of the relationship between weed diversity in arable fields, the diversity of wild pollinators (in semi-natural habitats adjacent to fields), crop production and the visitation rates and pollen consumption by honeybees. Many basic questions remain unanswered: Which species of bees pollinate crops? What fraction of crops are pollinated based on experimental evidence? How does pollination affect crop production (number of seeds, seed size, fat content)? Can weed biodiversity be maintained in agro-ecosystem without population outbreaks? How can ecological and stakeholder conflicts be resolved at landscape scale?
Although there is a general call for ecological intensification in farmland landscapes (e.g. Bommarco et al. 2013; Dore et al. 2011), such major knowledge gaps should preclude any recommendations or statements on management at this stage and a more prudent strategy relying on ecological engineering should be adopted. Experiments conducted at landscape scale, in parallel with modelling exercises (see, e.g. Devaux et al. 2008) are perhaps the best course of action in the short term. Experimental tests may involve monitoring well-designed, broad, large-scale herbicide reduction plans (on the lines ofthe French Ecophyto Plan), at least at farm scale, in order to test basic predictions such as increased pollination services, improved biological control and better conservation of threatened species. Additionally, careful quantitative measurement of crop yields, and farmers’revenues from pollination-dependant crops and a broader economic assessment of ecosystem services at landscape level will provide invaluable information for setting up measures to adapt management of farmland landscapes for the benefit of all stakeholders.
References
Aizen MA, Garibaldi LA, Cunningham SA et al (2009) How much does agriculture depend on pollinators? Lessons from long-termtrends in crop production. Ann Bot 103:1579–1588
Alaux C, Ducloz F, Crauser D, Le Conte Y (2010) Diet effects on honeybee immunocompetence. Biol Lett 6(4):562–565. doi:10.1098/rsbl.2009.0986
Andreasen C, Streibig JC (2011) Evaluation of changes in weed flora in arable fields of Nordic countries - based on Danish long-term surveys. Weed Res 51:214–226
Aronne G, Giovanetti M, Guarracino MR, de Micco V (2012) Foraging rules of flower selection applied by colonies of Apis mellifera: ranking and associations of floral sources. Funct Ecol 26(5):1186–1196. doi:10.1111/j.1365-2435.2012.02017.x
Aubert M (2002) Analyse des phénomènes d' affaiblissement des colonies d'abeilles. AFSSA
Baessler C, Klotz S (2006) Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric Ecosyst Environ 115(1–4):43–50
Bastiaans L, Paolini R, Baumann DT (2008) Focus on ecological weed management: what is hindering adoption? Weed Res 48(6):481–491
Beckie H, Tardif FJ (2012) Herbicide cross resistance in weeds. Crop Prot 35:15–28
Beekman M, Ratnieks FLW (2000) Long-range foraging by the honey-bee, Apis mellifera L. Funct Ecol 14(4):490–496. doi:10.1046/j.1365-2435.2000.00443.x
Benjamin FE, Reilly JR, Winfree R (2014) Pollinator body size mediates the scale at which land use drives crop pollination services. J Appl Ecol 51(2):440–449. doi:10.1111/1365-2664.12198
Bennett EM, Peterson GD, Gordon LJ (2009) Understanding relationships among multiple ecosystem services. Ecol Lett 12:1394–1404. doi:10.1111/j.1461-0248.2009.01387.x
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? TREE 18(4):182–188. doi:10.1016/S0169-5347(03)00011-9
Biesmeijer JC, Roberts SPM, Reemer M, Ohlemuller R, Edwards M, Peeters T, Schaffers AP, Potts SG, Kleukers R, Thomas CD, Settele J, Kunin WE (2006) Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313(5785):351–354. doi:10.1126/science.1127863
Blaauw BR, Isaacs R (2014) Flower plantings increase wild bee abundance and the pollination services provided to a pollination-dependent crop. J Appl Ecol 51(4):890–898
Blitzer EJ, Dormann CF, Holzschuh A, Klein AM, Rand TA, Tscharntke T (2012) Spillover of functionally important organisms between managed and natural habitats. Agric Ecosyst Environ 146(1):34–43. doi:10.1016/j.agee.2011.09.005
Bommarco R, Marini L, Vaissiere BE (2012) Insect pollination enhances seed yield, quality, and market value in oilseed rape. Oecologia 169(4):1025–1032. doi:10.1007/s00442-012-2271-6
Bommarco R, Kleijn D, Potts SG (2013) Ecological intensification: harnessing ecosystem services for food security. TREE 28(4):230–238. doi:10.1016/j.tree.2012.10.012
Breeze TD, Vaissière BE, Bommarco R, Petanidou T, Seraphides N, Kozak L, Scheper J, Biesmeijer JC, Kleijn D, Gyldenkærne S, Moretti M, Holzschuh A, Steffan-Dewenter A, Stout JC, Pärtel M, Zobel M, Potts SG (2014) Agricultural Policies Exacerbate Honeybee Pollination Service Supply-Demand Mismatches Across Europe. PLoS ONE 9(1):e82996. doi:10.1371/journal.pone.0082996
Bretagnolle V, Villers A, Denonfoux L, Cornulier T, Inchausti P, Badenhausser I (2011) Rapid recovery of a depleted population of Little Bustards Tetrax tetrax following provision of alfalfa through an agri-environment scheme. Ibis 153(1):4–13. doi:10.1111/j.1474-919X.2010.01092.x
Brittain C, Williams N, Kremen C, Klein AM (2013) Synergistic effects of non-Apis bees and honey bees for pollination services. Proc R Soc B-Biol Sci 280 (1754). doi:10.1098/rspb.2012.2767
Brodschneider R, Crailsheim K (2010) Nutrition and health in honey bees. Apidologie 41(3):278–294. doi:10.1051/apido/2010012
Burkle LA, Marlin JC, Knight TM (2013) Plant-Pollinator Interactions over 120 Years: Loss of Species, Co-Occurrence, and Function. Science 339(6127):1611–1615. doi:10.1126/science.1232728
Cameron SA, Lozier JD, Strange JP, Koch JB, Cordes N, Solter LF, Griswold TL (2011) Patterns of widespread decline in North American bumble bees. PNAS 108(2):662–667. doi:10.1073/pnas.1014743108
Campos MGR, Bogdanov S, Bicudo de Almedia-Muradian L, Szczesna T, Mancebo Y, Frigerio C, Ferreira F (2008) Pollen composition and standardisation of analytical methods. J Apicul Res Bee World 47:154–161
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486(7401):59–67. doi:10.1038/nature11148
Carvalheiro LG, Seymour CL, Veldtman R, Nicolson SW (2010) Pollination services decline with distance from natural habitat even in biodiversity-rich areas. J Appl Ecol 47(4):810–820. doi:10.1111/j.1365-2664.2010.01829.x
Carvalheiro LG, Veldtman R, Schenkute AG, Tesfay GB, Pirk CWW, Donaldson JC, Nicolson SW (2011) Natural and within-farmland biodiversity enhances crop productivity. Ecol Lett 14(3):251–259. doi:10.1111/j.1461-0248.2010.01579.x
Carvell C, Roy DB, Smart SM, Pywell RF, Preston CD, Goulson D (2006) Declines in forage availability for bumblebees at a national scale. Biol Conserv 132(4):481–489. doi:10.1016/j.biocon.2006.05.008
Chancellor RJ (1986) Decline of arable weed seeds during 20 years in soil under grasse and the periodicity of seedling emergence after cultivation. J Appl Ecol 23(2):631–637. doi:10.2307/2404041
Chapin FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU, Lavorel S, Sala OS, Hobbie SE, Mack M, Díaz S (2000) Consequences of changing biodiversity. Nature 405:234–242. doi:10.1038/35012241
Chaplin-Kramer R, de Valpine P, Mills NJ, Kremen C (2013) Detecting pest control services across spatial and temporal scales. Agric Ecosyst Environ 181:206–212. doi:10.1016/j.agee.2013.10.007
Charriere JD, Imdorf A, Koenig C, Gallmann S, Kuhn R (2010) Do sunflowers influence the development of honey bee, Apis mellifera, colonies in areas with diversified crop farming? J Apicult Res 49(3):227–235. doi:10.3896/ibra.1.49.3.01
Clergue B, Amiaud B, Pervanchon F, Lasserre-Joulin F, Plantureux S (2005) Biodiversity: function and assessment in agricultural areas. A rev Agron Sustain Dev 25(1):1–15. doi:10.1051/agro:2004049
Coffey MF, Breen J (1997) Seasonal variation in pollen and nectar sources of honey bees in Ireland. J Apic Res 36:63--76
Costanza R, d’Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, Oneill RV, Paruelo J, Raskin RG, Sutton P, vanden Belt M (1997) The value of the world's ecosystem services and natural capital. Nature 387(6630):253–260. doi:10.1038/387253a0
Couvillon MJ, Schurch R, Ratnieks FLW (2014) Dancing Bees Communicate a Foraging Preference for Rural Lands in High-Level Agri-Environment Schemes. Curr Biol 24(11):1212–1215. doi:10.1016/j.cub.2014.03.072
Cox-Foster DL, Conlan S, Holmes EC, Palacios G, Evans JD, Moran NA, Quan PL, Briese T, Hornig M, Geiser DM, Martinson V, vanEngelsdorp D, Kalkstein AL, Drysdale A, Hui J, Zhai JH, Cui LW, Hutchison SK, Simons JF, Egholm M, Pettis JS, Lipkin WI (2007) A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318(5848):283–287. doi:10.1126/science.1146498
Decourtye A, Mader E, Desneux N (2011) Landscape enhancement of floral resources for honey bees in agro-ecosystems. Apidologie 41(3):264–277. doi:10.1051/apido/2010024
Deguines N, Jono C, Baude M, Henry M, Julliard R, Fontaine C (2014) Large- scale trade- off between agricultural intensification and crop pollination services. Front Ecol Environ 12(4):212–217. doi:10.1890/130054
Delaplane KS, Mayer NF (2000) Crop Pollination by Bees. CABI Publishing, Wallingford
Delye C, Menchari Y, Guillemin JP, Matejicek A, Michel S, Camilleri C, Chauvel B (2007) Status of black grass (Alopecurus myosuroides) resistance to acetyl-coenzyme A carboxylase inhibitors in France. Weed Res 47(2):95–105
Delye C, Michel S, Berard A, Chauvel B, Brunel D, Guillemin JP, Dessaint F, Le Corre V (2010) Geographical variation in resistance to acetyl-coenzyme A carboxylase-inhibiting herbicides across the range of the arable weed Alopecurus myosuroides (black-grass). New Phytol 186(4):1005–1017. doi:10.1111/j.1469-8137.2010.03233.x
Delye C, Jasieniuk M, Le Corre V (2013) Deciphering the evolution of herbicide resistance in weeds. Trends Genet 29(11):649–658. doi:10.1016/j.tig.2013:06.001
Desneux N, Decourtye A, Delpuech JM (2007) The sublethal effects of pesticides on beneficial arthropods. In: Annual Review of Entomology. Ann Rev Entomol 52:81–106. doi:10.1146/annurev.ento.52.110405.091440
Devaux C, Klein EK, Lavigne C, Sausse C, Messéan A (2008) Environmental and landscape effects on cross-pollination rates observed at long distance among French oilseed rape (Brassica napus) commercial fields. J Appl Ecol 45:804–813. doi:10.1111/j.1365-2664.2007.01400.x
Di Pasquale G, Salignon M, Le Conte Y, Belzunces LP, Decourtye A, Kretzschmar A, Suchail S, Brunet J-L, Alaux C (2013) Influence of pollen nutrition on honey bee health: do pollen quality and diversity matter? PLoS One 8(8):e72016. doi:10.1371/journal.pone.0072016
Diekötter T, Kadoya T, Peter F, Wolters V, Jauker F (2010) Oilseed rape crops distort plant-pollinator interactions. J Appl Ecol 47(1):209–214. doi:10.1111/j.1365-2664.2009.01759.x
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe's farmland bird populations. Proc R Soc B-Biol Sci 268(1462):25–29. doi:10.1098/rspb.2000.1325
Dore T, Makowski D, Malezieux E, Munier-Jolain N, Tchamitchian M, Tittonell P (2011) Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. Eur J Agronom 34(4):197–210. doi:10.1016/j.eja.2011.02.006
Duelli P, Obrist MK (2003) Biodiversity indicators: the choice of values and measures. Agric Ecosyst Environ 98(1–3):87–98. doi:10.1016/s0167-8809(03)00072-0
Ehrlich PR, Ehrlich AH (1981) Extinction: The Causes and Consequences of the Disappearance of Species. Random House, New York
Ellis EJD, Evans JD, Pettis J (2010) Colony losses, managed colony population decline, and Colony Collapse Disorder in the United States. J Apic Res 49:134–136
Eraud C, Cadet E, Powolny T, Gaba S, Bretagnolle F, Bretagnolle V (2015) Weed seeds, not grain, contribute to the diet of wintering skylarksin arable farmlands of Western France. Eur J Wildl Res 61(1):151–161. doi:10.1007/s10344-014-0888-y
Evans DM, Pocock MJO, Brooks J, Memmott J (2011) Seeds in farmland food-webs: Resource importance, distribution and the impacts of farm management. Biol Conserv 144:2941–2950. doi:10.1016/j.biocon.2011.08.013
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski JH, Holloway T, Howard EA, Kucharik CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309:570–574. doi:10.1126/science.1111772
Fried G, Norton LR, Reboud X (2008) Environmental and management factors determining weed species composition and diversity in France. Agric Ecosyst Environ 128(1–2):68–76. doi:10.1016/j.agee.2008.05.003
Fried G, Chauvel B, Reboud X (2009a) A functional analysis of large-scale temporal shifts from 1970 to 2000 in weed assemblages of sunflower crops in France. J Veget Sci 20(1):49–58. doi:10.1111/j.1654-1103.2009.05284.x
Fried G, Petit S, Dessaint F, Reboud X (2009b) Arable weed decline in Northern France: Crop edges as refugia for weed conservation? Biol Conserv 142(1):238–243. doi:10.1016/j.biocon.2008.09.029
Fried G, Kazakou E, Gaba S (2012) Trajectories of weed communities explained by traits associated with species' response to management practices. Agric Ecosyst Environ 158:147–155. doi:10.1016/j.agee.2012.06.005
Gaba S, Bretagnolle F, Rigaud T, Philippot L (2014) Managing biotic interactions for ecological intensification of agroecosystem. Front Ecol Evol 2. doi:10.3389/fevo.2014.00029
Gabriel D, Tscharntke T (2007) Insect pollinated plants benefit from organic farming. Agric Ecosyst Environ 118(1–4):43–48. doi:10.1016/j.agee.2006.04.005
Gabriel D, Sait SM, Kunin WE, Benton TG (2013) Food production vs. biodiversity: comparing organic and conventional agriculture. J Appl Ecol 50(2):355–364. doi:10.1111/1365-2664.12035
Gallai N, Salles JM, Settele J, Vaissiere BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol Econom 68(3):810–821. doi:10.1016/j.ecolecon.2008.06.014
Garibaldi LA, Aizen MA, Klein AM, Cunningham SA, Harder LD (2011) Global growth and stability of agricultural yield decrease with pollinator dependence. PNAS 108:5909–5914. doi:10.1073/pnas.1012431108
Garibaldi LA, Steffan-Dewenter I, Winfree R, Aizen MA, Bommarco R, Cunningham SA, Kremen C, Carvalheiro LG, Harder LD, Afik O, Bartomeus I, Benjamin F, Boreux V, Cariveau D, Chacoff NP, Dudenhoffer JH, Freitas BM, Ghazoul J, Greenleaf S, Hipolito J, Holzschuh A, Howlett B, Isaacs R, Javorek SK, Kennedy CM, Krewenka KM, Krishnan S, Mandelik Y, Mayfield MM, Motzke I, Munyuli T, Nault BA, Otieno M, Petersen J, Pisanty G, Potts SG, Rader R, Ricketts TH, Rundlof M, Seymour CL, Schuepp C, Szentgyorgyi H, Taki H, Tscharntke T, Vergara CH, Viana BF, Wanger TC, Westphal C, Williams N, Klein AM (2013) Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 339(6127):1608–1611. doi:10.1126/science.1230200
Garratt MPD, Breeze TD, Jenner N, Polce C, Biesmeijer JC, Potts SG (2014) Avoiding a bad apple: Insect pollination enhances fruit quality and economic value. Agric Ecosyst Environ 184:34–40. doi:10.1016/j.agee.2013.10.032
Geiger F, Bengtsson J, Berendse F, Weisser WW, Emmerson M, Morales MB, Ceryngier P, Liira J, Tscharntke T, Winqvist C, Eggers S, Bommarco R, Pärt T, Bretagnolle V, Plantegenest M, Clement LW, Dennis C, Palmer C, Oñate JJ, Guerrero I, Hawro V, Aavik T, Thies C, Flohre A, Hänke S, Fischer C, Goedhart PW, Inchausti P (2010) Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl Ecol 11(2):97–105. doi:10.1016/j.baae.2009.12.001
Ghazoul J (2007) Challenges to the uptake of the ecosystem service rationale for conservation. Conserv Biol 21(6):1651–1652. doi:10.1111/j.1523-1739.2007.00758.x
Gibson RH, Nelson IL, Hopkins GW, Hamlett BJ, Memmott J (2006) Pollinator webs, plant communities and the conservation of rare plants: arable weeds as a case study. J Appl Ecol 43(2):246–257. doi:10.1111/j.1365-2664.2006.01130.x
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Pretty J, Robinson S, Thomas SM, Toulmin C (2010) Food Security: The Challenge of Feeding 9 Billion People. Science 327(5967):812–818. doi:10.1126/science.1185383
Gonzalez-Varo JP, Biesmeijer JC, Bommarco R, Potts SG, Schweiger O, Smith HG, Steffan-Dewenter I, Szentgyorgyi H, Woyciechowski M, Vila M (2013) Combined effects of global change pressures on animal-mediated pollination. TREE 28(9):524–530. doi:10.1016/j.tree.2013.05.008
Green RE, Cornell SJ, Scharlemann JPW, Balmford A (2005) Farming and the fate of wild nature. Science 307(5709):550–555. doi:10.1126/science.1106049
Greenleaf SS, Kremen C (2006) Wild bees enhance honey bees' pollination of hybrid sunflower. PNAS 103(37):13890–13895. doi:10.1073/pnas.0600929103
Haydak MH (1970) Honey Bee Nutrition. Ann Rev Entomol 15:143–156. doi:10.1146/annurev.en.15.010170.001043
Hayter KE, Cresswell JE (2006) The influence of pollinator abundance on the dynamics and efficiency of pollination in agricultural Brassica napus: implications for landscape-scale gene dispersal. J Appl Ecol 43(6):1196–1202. doi:10.1111/j.1365-2664.2006.01219.x
Hendrickx F, Maelfait JP, Van Wingerden W, Schweiger O, Speelmans M, Aviron S, Augenstein I, Billeter R, Bailey D, Bukacek R, Burel F, Diekotter T, Dirksen J, Herzog F, Liira J, Roubalova M, Vandomme V, Bugter R (2007) How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. J Appl Ecol 44(2):340–351. doi:10.1111/j.1365-2664.2006.01270.x
Henry M, Beguin M, Requier F, Rollin O, Odoux JF, Aupinel P, Aptel J, Tchamitchian S, Decourtye A (2012) A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 336(6079):348–350. doi:10.1126/science.1215039
Hoehn P, Tscharntke T, Tylianakis JM, Steffan-Dewenter I (2008) Functional group diversity of bee pollinators increases crop yield. Proc R Soc B-Biol Sci 275(1648):2283–2291. doi:10.1098/rspb.2008.0405
Hoehn P, Steffan-Dewenter I, Tscharntke T (2010) Relative contribution of agroforestry, rainforest and openland to local and regional bee diversity. Biodivers Conserv 19(8):2189–2200. doi:10.1007/s10531-010-9831-z
Holzschuh A, Steffan-Dewenter I, Tscharntke T (2008) Agricultural landscapes with organic crops support higher pollinator diversity. Oikos 117(3):354–361. doi:10.1111/j.2007.0030-1299.16303.x
Holzschuh A, Steffan-Dewenter I, Tscharntke T (2010) How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J Anim Ecol 79(2):491–500. doi:10.1111/j.1365-2656.2009.01642.x
Holzschuh A, Dudenhoffer JH, Tscharntke T (2013) Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biol Conserv 153:101–107. doi:10.1016/j.biocon.2012.04.032
Hopwood JL (2008) The contribution of roadside grassland restorations to native bee conservation. Biol Conserv 141(10):2632–2640. doi:10.1016/j.biocon.2008.07.026
Hyvonen T (2007) Can conversion to organic farming restore the species composition of arable weed communities? Biol Conserv 137(3):382–390. doi:10.1016/j.biocon.2007.02.021
Isaacs R, Kirk AK (2010) Pollination services provided to small and large highbush blueberry fields by wild and managed bees. J Appl Ecol 47:841–849. doi:10.1111/j.1365-2664.2010.01823.x
Isbell F, Reich PB, Tilman D, Hobbie SE, Polasky S, Binder S (2013) Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. PNAS 110(29):11911–11916. doi:10.1073/pnas.1310880110
Jaffe R, Dietemann V, Allsopp MH, Costa C, Crewe RM, Dall'olio R, de la Rua P, El-Niweiri MAA, Fries I, Kezic N, Meusel MS, Paxton RJ, Shaibi T, Stolle E, Moritz RFA (2010) Estimating the Density of Honeybee Colonies across Their Natural Range to Fill the Gap in Pollinator Decline Censuses. Conserv Biol 24(2):583–593. doi:10.1111/j.1523-1739.2009.01331.x
Jasieniuk M (1996) The evolution and genetics of herbicide resistance in weeds. Weed Sci 44:176–193
Javorek SK, Mackenzie KE, Vander Kloet SP (2002) Comparative pollination effectiveness among bees (Hymenoptera : Apoidea) on lowbush blueberry (Ericaceae : Vaccinium angustifolium). Ann Entomol Soc Am 95(3):345–351. doi:10.1603/0013-8746(2002)095[0345:cpeabh]2.0.co;2
Kennedy CM, Lonsdorf E, Neel MC, Williams NM, Ricketts TH, Winfree R, Bommarco R, Brittain C, Burley AL, Cariveau D, Carvalheiro LG, Chacoff NP, Cunningham SA, Danforth BN, Dudenhoffer JH, Elle E, Gaines HR, Garibaldi LA, Gratton C, Holzschuh A, Isaacs R, Javorek SK, Jha S, Klein AM, Krewenka K, Mandelik Y, Mayfield MM, Morandin L, Neame LA, Otieno M, Park M, Potts SG, Rundlof M, Saez A, Steffan-Dewenter I, Taki H, Viana BF, Westphal C, Wilson JK, Greenleaf SS, Kremen C (2013) A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol Lett 16(5):584–599. doi:10.1111/ele.12082
Klatt BK, Holzschuh A, Westphal C, Clough Y, Smit I, Pawelzik E, Tscharntke T (2014) Bee pollination improves crop quality, shelf life and commercial value. Proc R Soc B-Biol Sci 281 (1775). doi:10.1098/rspb.2013.2440
Kleijn D, Sutherland WJ (2003) How effective are European agri-environment schemes on conserving and promoting biodiversity? J Appl Ecol 40:947–970
Kleijn D, Berendse F, Smit R, Gilisen N, Smit J, Brak B, Groeneveld R (2004) Ecological Effectiveness of Agri-Environment Schemes in Different Agricultural Landscapes in The Netherlands. Conservation Biology, 18:775–786. doi:10.1111/j.1523-1739.2004.00550.x
Klein AM, Steffan-Dewenter I, Tscharntke T (2003) Fruit set of highland coffee increases with the diversity of pollinating bees. Proc R Soc B-Biol Sci 270(1518):955–961. doi:10.1098/rspb.2002.2306
Klein AM, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, Tscharntke T (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc B-Biol Sci 274(1608):303–313. doi:10.1098/rspb.2006.3721
Klein AM, Brittain C, Hendrix SD, Thorp R, Williams N, Kremen C (2012) Wild pollination services to California almond rely on semi-natural habitat. J Appl Ecol 49(3):723–732. doi:10.1111/j.1365-2664.2012.02144.x
Knox OGG, Leake AR, Walker RL, Edwards AC, Watson CA (2011) Revisiting the multiple benefits of historical crop rotations within contemporary UK agricultural systems. J Sustain Agri 35:163–179
Kremen C, Williams NM, Thorp RW (2002) Crop pollination from native bees at risk from agricultural intensification. PNAS 99(26):16812–16816. doi:10.1073/pnas.262413599
Kuhlmann M, et al (2013) Checklist of the Western Palaearctic Bees (Hymenoptera: Apoidea: Anthophila). http://westpalbees.myspecies.info
Lautenbach S, Seppelt R, Liebscher J, Dormann CF (2012) Spatial and Temporal Trends of Global Pollination Benefit. PLoS ONE 7 (4). doi:10.1371/journal.pone.0035954
Le Conte Y, Ellis M, Ritter W (2010) Varroa mites and honey bee health: can Varroa explain part of the colony losses? Apidologie 41:353–363
Le Féon V, Schermann-Legionnet A, Delettre Y, Aviron S, Billeter R, Bugter R, Hendrickx F, Burel F (2010) Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric Ecosyst Environ 137(1–2):143–150. doi:10.1016/j.agee.2010.01.015
Le Féon V, Burel F, Chifflet R, Henry M, Ricroch A, Vaissière BE, Baudry J (2013) Solitary bee abundance and species richness in dynamic agricultural landscapes. Agric Ecosyst Environ 166:94–101
Lososova Z, Simonova D (2008) Changes during the 20th century in species composition of synanthropic vegetation in Moravia (Czech Republic). Preslia 80:291–305
Mader E, Shepherd M, Vaughan M, Hoffman Black S, LeBuhn G (2011) Attracting native pollinators: protecting North America’s bees and butterflies: the Xerces Society guide. Storey Pub, North Adams
Manalil S, Busi R, Renton M, Powles SB (2011) Rapid Evolution of Herbicide Resistance by Low Herbicide Dosages. Weed Sci 59:210–217
Manning R (2001) Fatty acids in pollen: a review of their importance for honey bees. Bee World 82(2):60–75
Marshall EJP, Brown VK, Boatman ND, Lutman PJW, Squire GR, Ward LK (2003) The role of weeds in supporting biological diversity within crop fields. Weed Res 43:77–89
Mayack C, Naug D (2009) Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J Invert Pathol 100(3):185–188. doi:10.1016/j.jip.2008.12.001
Meeus I, Brown MJF, De Graaf DC, Smagghe G (2011) Effects of Invasive Parasites on Bumble Bee Declines. Conserv Biol 25(4):662–671. doi:10.1111/j.1523-1739.2011.01707.x
Meiss H, Munier-Jolain N, Henriot F, Caneill J (2008) Effects of biomass, age and functional traits on regrowth of arable weeds after cutting. J Plant Diseas Prot 21:493–499
Meyer S, Wesche K, Krause B, Leuschner C (2013) Dramatic losses of spcialist arable plants in cenral Germany since the 1950/60s - a cross-regional analysis. Diver Distri 19:1175–1187. doi:10.1111/ddi.12102
Michener CD (2007) The Bees of the World, vol 2nd Revised edition. Hopkins University Press
Milberg P, Hallgren E (2004) Yield loss due to weeds in cereals and its large-scale variability in Sweden. Field Crop Res 86(2–3):199–209. doi:10.1016/j.fcr.2003.08.006
Mirek Z (1976) The extinction of flax weed Camelina alysum (Mill.) Thell. in Poland. Phytocoenosis 5:227–236
Morandin LA, Winston ML (2005) Wild bee abundance and seed production in conventional, organic, and genetically modified canola. Ecol Appl 15(3):871–881. doi:10.1890/03-5271
Morandin LA, Winston ML (2006) Pollinators provide economic incentive to preserve natural land in agroecosystems. Agric Ecosyst Environ 116(3–4):289–292. doi:10.1016/j.agee.2006.02.012
Moritz RFA (1991) The limitations of biometric control on pure race breeding in Apis mellifera. J Apic Res 30(2):54–59
Neff J, Simpson B (1993) Bees, pollination systems and plant diversity. In: LaSalle J, Gauld I (eds) Hymenoptera and Biodiversity. CAB International, Wallingford, pp 143–167
Neumann P, Carreck NL (2010) Honey bee colony losses. J Apicul Res 49(1):1–6. doi:10.3896/ibra.1.49.1.01
Neve P, Vila-Aiub M (2009) Evolutionary-thinking in agricultural weed management. New Phytol 184:783–793
Nielsen A, Dauber J, Kunin WE, Lamborn E, Jauker B, Moora M, Potts SG, Reitan T, Roberts S, Sober V, Settele J, Steffan-Dewenter I, Stout JC, Tscheulin T, Vaitis M, Vivarelli D, Biesmeijer JC, Petanidou T (2012) Pollinator community responses to the spatial population structure of wild plants: A pan-European approach. Bas Appl Ecol 13(6):489–499. doi:10.1016/j.baae.2012.08.008
Ockinger E, Smith HG (2007) Semi-natural grasslands as population sources for pollinating insects in agricultural landscapes. J Appl Ecol 44(1):50–59. doi:10.1111/j.1365-2664.2006.01250.x
Odoux JF, Lamy H, Aupinel P (2004) L'abeille récolte-t-elle du pollen de maïs et de tournesol? LSA 201(5–6):187–193
Odoux JF, Feuillet D, Aupinel P, Loublier Y, Tasei JN, Mateescu C (2012) Territorial biodiversity and consequences on physico-chemical characteristics of pollen collected by honey bee colonies. Apidologie 43(5):561–575. doi:10.1007/s13592-012-0125-1
Odoux JF, Aupinel P, Gateff S, Requier F, Henry M, Bretagnolle V (2014) ECOBEE: a tool for long-term bee colony monitoring at landscape scale in West European intensive agrosystems. J Apicult Res 53(1):57–66. doi:10.3896/IBRA.1.53.1.05
Oerke E-C (2006) Crop losses to pests. J Agric Sci 144(1):31–43. doi:10.1017/S0021859605005708
O'Farrell PJ, Anderson PML (2010) Sustainable multifunctional landscapes: a review to implementation. Curr Opin Environ Sustain 2(1–2):59–65. doi:10.1016/j.cosust.2010.02.005
Oldroyd BP (2007) What's killing American honey Bees? Plos Biol 5(6):1195–1199. doi:10.1371/journal.pbio.0050168
Oldroyd BP, Nanork P (2009) Conservation of Asian honey bees. Apidologie 40(3):296–312. doi:10.1051/apido/2009021
Ollerton J, Winfree R, Tarrant S (2011) How many flowering plants are pollinated by animals? Oikos 120(3):321–326. doi:10.1111/j.1600-0706.2010.18644.x
O'Rourke E (2006) Biodiversity and land use change on the Causse Mejan, France. Biodivers Conserv 15(8):2611–2626. doi:10.1007/s10531-005-5402-0
Osborne JL, Williams IH, Marshall AH, Michaelson-Yeates TPT (2001) Pollination and gene flow in white clover, growing in a patchy habitat. In: Benedek P, Richards KW (eds) Proceedings of the Eight International Pollination Symposium Pollination: Integrator of Crops and Native Plant Systems. Acta Horticulturae. pp 35-40
Patiny S, Rasmont P, Michez D (2009) A survey and review of the status of wild bees in the West-Palaearctic region. Apidol 40(3):313–331. doi:10.1051/apido/2009028
Pe'er G, Dicks LV, Visconti P, Arlettaz R, Baldi A, Benton TG, Collins S, Dieterich M, Gregory RD, Hartig F, Henle K, Hobson PR, Kleijn D, Neumann RK, Robijns T, Schmidt J, Shwartz A, Sutherland WJ, Turbe A, Wulf F, Scott AV (2014) AGRICULTURE POLICY EU agricultural reform fails on biodiversity. Science 344(6188):1090–1092. doi:10.1126/science.1253425
Pernal SF, Currie RW (2000) Pollen quality of fresh and 1-year-old single pollen diets for worker honey bees (Apis mellifera L.). Apidol 31(3):387–409
Pernal SF, Currie RW (2001) The influence of pollen quality on foraging behavior in honeybees (Apis mellifera L.). Behav Ecol Soc 51:53–68
Persson AS, Smith HG (2013) Seasonal persistence of bumblebee populations is affected by landscape context. Agric Ecosyst Environ 165:201–209. doi:10.1016/j.agee.2012.12.008
Phalan B, Onial M, Balmford A, Green RE (2011) Reconciling Food Production and Biodiversity Conservation: Land Sharing and Land Sparing Compared. Science 333(6047):1289–1291. doi:10.1126/science.1208742
Pisanty G, Klein AM, Mandelik Y (2014) Do wild bees complement honeybee pollination of confection sunflowers in Israel? Apidol 45(2):235–247. doi:10.1007/s13592-013-0242-5
Potts SG, Vulliamy B, Dafni A, Ne'eman G, Willmer P (2003) Linking bees and flowers: how do floral communities structure pollinator communities? Ecology 84(10):2628–2642. doi:10.1890/02-0136
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010a) Global pollinator declines: trends, impacts and drivers. TREE 25(6):345–353. doi:10.1016/j.tree.2010.01.007
Potts SG, Roberts SPM, Dean R, Marris G, Brown MA, Jones R, Neumann P, Settele J (2010b) Declines of managed honey bees and beekeepers in Europe. J Apicult Res 49(1):15–22. doi:10.3896/ibra.1.49.1.02
Powles SB, Yu Q (2010) Evolution in action: plants resistant to herbicides. Annu Rev Plant Physiol Plant Mol Biol 61:317–347
Preston CD, Pearman DA, Hall AR (2004) Archaeophytes in Britain. Bot J Linnean Soc 145:257–294
Rader R, Howlett BG, Cunningham SA, Westcott DA, Edwards W (2012) Spatial and temporal variation in pollinator effectiveness: do unmanaged insects provide consistent pollination services to mass flowering crops? J Appl Ecol 49(1):126–134. doi:10.1111/j.1365-2664.2011.02066.x
Rao S, Strange JP (2012) Bumble Bee (Hymenoptera: Apidae) Foraging Distance and Colony Density Associated With a Late-Season Mass Flowering Crop. Environ Entomol 41(4):905–915
Ratnieks FLW, Carreck NL (2010) Clarity on Honey Bee Collapse? Science 327(5962):152–153. doi:10.1126/science.1185563
Requier F, Odoux JF, Tamic T, Moreau N, Henry M, Decourtye A, Bretagnolle V (2015) Honey-bee diet in intensive farmland habitats reveals an unexpected flower richness and a critical role of weeds. Ecol Appl. doi:10.1890/14-1011.1
Richner N, Holderegger R, Linder HP, Walter T (2015) Reviewing change in the arable flora of Europe: a meta-analysis. Weed Res 55:1–13. doi:10.1111/wre.12123
Ricketts TH, Regetz J, Steffan-Dewenter I, Cunningham SA, Kremen C, Bogdanski A, Gemmill-Herren B, Greenleaf SS, Klein AM, Mayfield MM, Morandin LA, Ochieng A, Potts SG, Viana BF (2008) Landscape effects on crop pollination services: are there general patterns? Ecol Lett 11(10):1121. doi:10.1111/j.1461-0248.2008.01241.x
Riedinger V, Renner M, Rundlof M, Steffan-Dewenter I, Holzschuh A (2014) Early mass-flowering crops mitigate pollinator dilution in late-flowering crops. Landsc Ecol 29(3):425–435. doi:10.1007/s10980-013-9973-y
Roberts HA (1981) Seed banks in soils. Adv Appl Biol 6:1–55
Roberts HA, Feast PM (1973) Changes in the numbers of viable weed seeds in soil under different regimes. Weed Res 13:298–303
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39:157–176
Rollin O (2013) Etude Multi-Echelle Du Patron De Diversite Des Abeilles Et Utilisation Des Ressources Fleuries
Rollin O, Bretagnolle V, Decourtye A, Aptel J, Michel N, Vaissière BE, Henry M (2013) Differences of floral resource use between honey bees and wild bees in an intensive farming system. Agric Ecosyst Environ 179:78–86. doi:10.1016/j.agee.2013.07.007
Saddier M (2008) Pour une filière apicole durable-L'abeille et les pollinisateurs sauvages. Député de Haute-Savoie
Samnegard U, Persson AS, Smith HG (2011) Gardens benefit bees and enhance pollination in intensively managed farmland. Biol Conserv 144(11):2602–2606. doi:10.1016/j.biocon.2011.07.008
Schmidt LS, Schmidt JO, Rao H, Wang WY, Xu LG (1995) Feeding preference and survival young worker honey bees fed rape, sesame, sunflower pollen. J Econom Entomol 88(6):1591–1595
Seeley TD, Visscher PK (1985) Survival of honeybees in cold climates - The critical timing of colony growth and reproduction. Ecol Entomol 10(1):81–88. doi:10.1111/j.1365-2311.1985.tb00537.x
Spahillari M, Hammer K, Gladis T, Diederichsen A (1999) Weeds as part of agriculture. Outlook Agric 28:227–232
Stanley DA, Stout JC (2014) Pollinator sharing between mass-flowering oilseed rape and co-flowering wild plants: implications for wild plant pollination. Plant Ecol 2014(215):315–325
Stanley DA, Knight ME, Stout JC (2013) Ecological Variation in Response to Mass-Flowering Oilseed Rape and Surrounding Landscape Composition by Members of a Cryptic Bumblebee Complex. PLoS ONE 8 (6). doi:10.1371/journal.pone.0065516
Steffan-Dewenter I (2003) Importance of habitat area and landscape context for species richness of bees and wasps in fragmented orchard meadows. Conserv Biol 17(4):1036–1044. doi:10.1046/j.1523-1739.2003.01575.x
Steffan-Dewenter I, Tscharntke T (2001) Succession of bee communities on fallows. Ecography 24(1):83–93. doi:10.1034/j.1600-0587.2001.240110.x
Steffan-Dewenter I, Westphal C (2008) The interplay of pollinator diversity, pollination services and landscape change. J Appl Ecol 45(3):737–741. doi:10.1111/j.1365-2664.2008.01483.x
Steffan-Dewenter I, Munzenberg U, Burger C, Thies C, Tscharntke T (2002) Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83(5):1421–1432. doi:10.1890/0012-9658(2002)083[1421:SDEOLC]2.0.CO;2
Storkey J (2006) A functional group approach to the management of UK arable weeds to support biological diversity. Weed Res 46:513–522. doi:10.1111/j.1365-3180.2006.00528.x
Storkey J, Moss SR, Cussans JW (2010) Using Assembly Theory to Explain Changes in a Weed Flora in Response to Agricultural Intensification. Weed Sci 58(1):39–46. doi:10.1614/WS-09-096.1
Sutcliffe OL, Kay QON (2000) Changes in the arable flora of central southern England since the 1960s. Biol Conserv 93:1–8. doi:10.1016/S0006-3207(99)00119-6
Thies C, Roschewitz I, Tscharntke T (2005) The landscape context of cereal aphid-parasitoid interactions. Proc R Soc B-Biol Sci 272(1559):203–210. doi:10.1098/rspb.2004.2902
Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S (2002) Agricultural sustainability and intensive production practices. Nature 418(6898):671–677. doi:10.1038/nature01014
Tilman D, Reich PB, Isbell F (2012) Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. PNAS 109(26):10394–10397. doi:10.1073/pnas.1208240109
Tixier P, Peyrard N, Aubertot J-N, Gaba S, Radoszycki J, Caron-Lormier G, Vinatier F, Mollot G, Sabbadin R (2013) Modelling Interaction Networks for Enhanced Ecosystem Services in Agroecosystems. Adv Ecol Res 49:437–480. doi:10.1016/B978-0-12-420002-9.00007-X
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity - ecosystem service management. Ecol Lett 8:857–874. doi:10.1111/j.1461-0248.2005.00782.x
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A (2012) Global food security, biodiversity conservation and the future of agricultural intensification. Biol Conserv 151(1):53–59. doi:10.1016/j.biocon.2012.01.068
Vaissiere BE, Vinson SB (1994) Pollen morphology and its effect on pollen collection by Honey-bees, Apis mellifera (Hymenoptera, Apidae), with special reference to upland cotton, Gossypium-Hirsutum L. (Malvaceae). Grana 33(3):128–138
van Dooremalen C, Gerritsen L, Cornelissen B, van der Steen JJM, van Langevelde F et al (2012) Winter Survival of Individual Honey Bees and Honey Bee Colonies Depends on Level of Varroa destructor Infestation. PLoS ONE 7(4):e36285. doi:10.1371/journal.pone.0036285
VanEngelsdorp D, Meixner MD (2010) A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J Invertebr Pathol 103(Suppl 1):S80–S95. doi:10.1016/j.jip.2009.06.011
VanEngelsdorp D, Hayes J, Underwood RM, Pettis J (2008) A Survey of Honey Bee Colony Losses in the US, Fall 2007 to Spring 2008. Plos One 3 (12). doi:10.1371/journal.pone.0004071
VanEngelsdorp D, Evans JD, Saegerman C, Mullin C, Haubruge E, Nguyen BK, Frazier M, Frazier J, Cox-Foster D, Chen YP, Underwood R, Tarpy DR, Pettis JS (2009) Colony Collapse Disorder: A Descriptive Study. PLoS One 4(8):17. doi:10.1371/journal.pone.0006481
Weiner J, Andersen SB, Wille WKM, Griepentrog HW, Olsen JM (2010) Evolutionary Agroecology: the potential for cooperative, high density, weed-suppressing cereals. Evol Applic 3(5–6):473–479. doi:10.1111/j.1752-4571.2010.00144.x
Westerman PR, Borza JK, Andjelkovic J, Liebman M, Danielson B (2008) Density-dependent predation of weed seeds in maize fields. J Appl Ecol 45:1612–1620. doi:10.1111/j.1365-2664.2008.01481.x
Westphal C, Steffan-Dewenter I, Tscharntke T (2003) Mass flowering crops enhance pollinator densities at a landscape scale. Ecol Lett 6(11):961–965. doi:10.1046/j.1461-0248.2003.00523.x
Whelan CJ, Wenny DG, Marquis RJ (2008) Ecosystem services provided by birds. In: Ostfeld RS, Schlesinger WH (eds) Year in Ecology and Conservation Biology. Ann NY Academ Sci. 1134: 25-60. doi:10.1196/anndis.1439.003
White SS, Renner KA, Menalled FD, Landis DA (2007) Feeding preferences of weed seed predators and effect on weed emergence. Weed Sci 55:606–612
Whitehorn PR, Tinsley MC, Brown MJF, Goulson D (2013) Investigating the impact of deploying commercial Bombus terrestris for crop pollination on pathogen dynamics in wild bumble bees. J Apic Res 52(3):149–157. doi:10.3896/ibra.1.52.3.06
Whittingham MJ, Krebs JR, Swetnam RD, Vickery JA, Wilson JD, Freckleton RP (2007) Should conservation strategies consider spatial generality? Farmland birds show regional not national patterns of habitat association. Ecol Lett 10:25–35. doi:10.1111/j.1461-0248.2006.00992.x
Williams IH (1994) Bees for pollination - conclusions and recommendations of the EC workshop on bees for pollination held in Brussels, 2-3 March 1992. Bee World 75(1):46–48
Willmer PG, Bataw AAM, Hughes JP (1994) The superiority of bumblebee to honeybees as pollinators - Insect vists to Raspberry flowers. Ecol Entomol 19(3):271–284. doi:10.1111/j.1365-2311.1994.tb00419.x
Winfree R, Williams NM, Gaines H, Ascher JS, Kremen C (2008) Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J Appl Ecol 45(3):793–802. doi:10.1111/j.1365-2664.2007.01418.x
Winfree R, Aguilar R, Vazquez DP, LeBuhn G, Aizen MA (2009) A meta-analysis of bees' responses to anthropogenic disturbance. Ecol 90(8):2068–2076. doi:10.1890/08-1245.1
Winfree R, Gross BJ, Kremen C (2011) Valuing pollination services to agriculture. Ecol Econ 71:80–88. doi:10.1016/j.ecolecon.2011.08.001
Wratten SD, Gillespie M, Decourtye A, Mader E, Desneux N (2012) Pollinator habitat enhancement: Benefits to other ecosystem services. Agric Ecosyst Environ 159:112–122. doi:10.1016/j.agee.2012.06.020
Acknowledgments
The authors wish to acknowledge in particular two PhD students, Fabrice Requier and Orianne Rollin, for the many fruitful discussions they had with them on bee ecology. Many thanks also to Jean-François Odoux and Axel Decourtye for sharing their knowledge on honeybees, and to Mickael Henry for collaboration on ECOBEE. The English language of this manuscript has been corrected by Tony Tebby. This analysis is part of the ECOBEE platform, and we thank the program DEPHY-abeilles and the ANR AgrobioSE for partial funding “ANR-13-AGRO-0001”.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Bretagnolle, V., Gaba, S. Weeds for bees? A review. Agron. Sustain. Dev. 35, 891–909 (2015). https://doi.org/10.1007/s13593-015-0302-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13593-015-0302-5