
Abstract
This work aimed to introduce a new technique for improving sustainable biodiesel from Microchloropsis salina by examining the impact of various nitrogen sources, salt stress, and carbon enrichment, individually or in combination, to determine the best treatment conditions for biomass and lipid productivity as well as biodiesel quality. M. salina effectively utilizes nitrate (NaNO3 and KNO3) and organic nitrogen (especially yeast extract and glycine) but not ammonium for growth. The highest biomass and lipid productivity were observed on yeast extract (0.11 g L−1 day−1 and 52.11 μgmL−1 day−1, respectively). Lipid production enhanced by 80% under 300 mM of NaCl, while growth increased only by 30%. Acetate could not be used to cultivate the alga mixotrophically; only glucose and glycerol were effective. The ideal organic carbon supply values for glucose and glycerol were 3 g L−1 and 2 g L−1, respectively. Nutrition by 2 g L−1 glycerol promoted biomass and lipid productivities by 62% and 95%, respectively, while biomass productivity was reduced by 16% under combined and lipid productivity increased by 80%. All treatments significantly improved the biomass and biodiesel quality-related fatty acid profile.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The need for fossil fuels, including oil, coal, and natural gases, has increased dramatically due to the rapid growth of the world population, economic growth, and rapid industrialization. Consequently, fossil fuels are running out more quickly. It is estimated that natural gas and oil reserves will be exhausted in between 40 and 64 years. By 2030, the global energy demand will have roughly a 60% increase [1]. Moreover, the risk of global warming due to anthropogenic CO2 emissions is related to coal, natural gas, and petrol combustion.
New renewable energy sources need to be developed to overcome all these problems. One of the most promising biofuels that has received much interest in recent years is biodiesel which considers a sustainable, eco-friendly, and CO2- emission-free alternative fuel. Microalgae’s quick growth, great photosynthetic efficiency, and lack of dependence on arable land make them a promising feedstock for manufacturing biodiesel. Also, microalgae can utilize CO2, nitrogen, and phosphate during their cultivation, leading to the greenhouse effect and water pollution mitigation. To produce 1 kg of biomass, approximately 1.8 kg of CO2 is required [2]. According to Liu et al. [3], up to 80% of the biomass that microalgae produce can be stored as lipids, and triacylglycerols (TAGs), which make up most of these lipids, can be used as a feedstock for the transesterification of lipid to create biodiesel. Since transportation expenses are generally low, microalgal oil is more affordable than biomass from trees and crops [4]. The fundamental properties of biodiesel made from microalgal oil are similar to those of conventional biodiesel, such as their high degree of saturation, cetane number, and viscosity [5].
There are other methods than large-scale algae production for increasing biodiesel generation. The metabolic process changes when the cultivation conditions are improved, which increases the generation of relative and mesh biodiesel. Numerous growth factors such as the availability of nutrients, light intensity, salinity, the presence of heavy metals, the pH scale, and the temperature significantly impact microalgal growth and metabolism, including lipid accumulation [6,7,8]. Under optimal growth conditions, microalgae produce fatty acids primarily for esterification into glycerol-based membrane lipids, while under unfavorable stressed conditions, many microalgae alter their fat biosynthesis pathways to synthesize and accumulate neutral lipids, primarily in the form of triacylglycerides, which make up about 5 to 20% of the cell dry weight (20–50% DW) [9, 10]. Organic carbon sources can serve as enhancer compounds and provide a mixotrophic condition preferable for algal growth and lipid production [11,12,13]. However, there is not much research addressing how combined stressors affect lipid productivity and fatty acid profile in biodiesel quality, whether in phototrophic or mixotrophic environments.
This study aimed to determine the ideal treatment scenario for M. salina in terms of biomass and lipid production as well as biodiesel quality by examining the effects of various nitrogen sources, salt stress, and carbon enrichment, both separately and in combination.
2 Materials and methods
2.1 Cultivation and growth condition
The strain Microchloropsis salina (formerly Nannochloropsis salina) was provided by the National Institute of Oceanography and Fisheries in Alexandria, Egypt. In 1-L Erlenmeyer flasks (flasks were stopped with cotton plugs), 800 mL of artificial seawater (ASW) fortified with F/2 nutrients had its NaNO3 concentration changed from 0.075 to 0.75 g L−1 [14]. The flasks were sterilized in an autoclave at 121 °C and 1.5 atom for 20 min. After cooling, the Erlenmeyer flasks were inoculated with a certain volume (10 mL) of microalgal strain pre-culture in 250-mL Erlenmeyer flasks. Aeration was given to the culture through silicon tubing, with one end inserted into the culture flask and the other into the bacterial filter connected to the aerator. Algal culture flasks were incubated under the continuous fluorescent light of 45 µmole m−2 s−1 and at a temperature of 25 ± 1 °C.
2.2 Optimization of media
2.2.1 Different nitrogen sources
In order to examine the effects of various nitrogen sources on the cell growth and lipid production of M. salina, potassium nitrate (KNO3, 0.892 g L−1), glycine (NH2CH2COOH, 0.662 g L−1), urea (CO (NH2)2, 0.265 g L−1), ammonium chloride (NH4Cl, 0.472 g L−1), and yeast extract (YE) (initial nitrogen concentration was the same, at 8.82 mM (123.5 mg N L−1) were used in place of sodium nitrate (NaNO3, 0.750 g L−1) in F/2 medium according to Kim et al.[15].
2.2.2 Salt stress
Four different sodium chloride (NaCl) concentrations (100, 200, 300, and 400 mM) were used to test the effect of salt stress.
2.2.3 Carbon enrichment
Different concentrations of three different carbon sources, glucose, glycerol, and sodium acetate (1, 2, and 3 g L−1), were used.
2.3 Measurement of microalgal growth and biomass
2.3.1 Optical density
Every 2 days, optical density at 680 nm was measured using a spectrophotometer to evaluate the growth of each algal culture. The resulting absorbance was used to plot the growth curve of respective algae cultures.
2.3.2 Dry weight measurement
A specific amount (30 mL) of the algal suspension was centrifuged at 4000 rpm for 15 min, followed by numerous thorough washes with distilled water. The precipitated biomass was dried overnight at 60 °C in an oven in pre-weighed Petri plates until constant weight after removing all salts from the medium. Data were given by g L−1 [16].
2.3.3 Estimation of biomass productivity
According to Eq. (1), which is discussed by Abomohra et al. (2013) [17], biomass productivity was calculated.
where CDWL (g L−1) is the cellular dry weight at the late exponential phase (tL) and CDWE (g L−1) is the cellular dry weight at the early exponential phase (tE).
2.4 Estimation of total lipids and productivity
This work adapted the Park procedure to use the SPV reaction to detect lipid content directly [18]. Initial preparation of the phospho-vanillin reagent involved constantly stirring until 0.06 g of vanillin was dissolved in 10 mL of deionized water, followed by the addition of 40 mL of concentrated phosphoric acid to the mixture. The biomass was extracted by centrifugation at 4000 RPM for 5 min using a known amount of biomass. The sample was treated with 2 mL of concentrated sulfuric acid (98%) and heated in a water bath at 100 °C for 10 min. The reaction mixture was then added to 5 mL of freshly made phospho-vanillin reagent once it had cooled, and it was all incubated for 15 min at 37 °C in the incubator. After that, a pink color emerged, and its absorbance was measured at 530 nm with a spectrophotometer to determine how much lipid was present in the sample. The standard curve was used to estimate the amount of lipids. The content of lipids was expressed as μg mL−1. According to Eq. (2), lipid productivity was calculated as discussed earlier by Abomohra et al. (2013) [17].
where LCL (μg mL−1) is the lipid content at the late exponential phase (tL) and LCE is the lipid content (μg mL−1) at the early exponential phase (tE).
2.5 Extraction of total lipids
Following certain changes, the modified Folch technique [19] was used to extract the total lipid content. After centrifuging 50 mL of algal culture for 15 min at 4500 rpm, 40 mL of chloroform/methanol (2:1, v/v) solution was added to the dried algal cells. To separate the liquid phase, the mixture was filtered through filter paper (Whatman No. 1) after being shaken at 120 rpm for 48 h at room temperature. The liquid phase was transferred to a fresh flask, rinsed with 0.9% NaCl (w/v) solution, and the two phases were separated using a separatory funnel. In order to evaporate the solvent, the lower phase containing the lipids was taken in a pre-weighed glass vial and dried at 40 °C for 2 days. The difference between W2 (weight of glass vial containing lipids) and W1 (weight of glass vial) indicates the quantity of total lipid content.
2.6 Cultivation of Microchloropsis salina in combined conditions
The most effective growth and total lipid content–affecting elements from the earlier trials were chosen, merged, and approved as the ideal conditions for additional experimentation and analysis.
2.7 Fatty acid profiles determination
For the preparation of fatty acid methyl esters (FAMEs), 1.0 mL of n-hexane was added to 15 mg of oil, followed by 1.0 mL of sodium methoxide (0.4 mol), according to the modified method [20]. After being vortexed for 30 s, the mixtures were left to settle for 15 min. The upper phase that contained the FAMEs was recovered and analyzed using gas chromatography (Perkin Elmer model: flame ionization detector). A Perkin Elmer Auto System XL with a flame ionization detector (FID) was used for the GC analysis. The capillary column was a DB-Wax made of fused silica (60 m × 0.32 mm i.d.). The oven’s temperature was initially kept at 150 °C and was designed to rise by 3 °C/min to 220 °C. 1.1 ml/min of helium was used as the carrier gas. Temperatures for the injector and detector were 230 and 250 °C, respectively.
2.8 Evaluation of some biodiesel properties
The biodiesel properties were calculated using the FAME composition of the microalgae, including the Average Degree of Unsaturation (ADU percent), Iodine Value (IV, g I2.100 g−1 oil), Cetane Number (CN), Kinematic viscosity (ʋi, mm2 s−1), Cloud point (CP, °C), density (ρ), Higher Heating Value (HHV), Saponification Value (SV, mg KOH g−1), Long Chain Saturation Factor (LCSF, wt %), and Cold Filter Plugging Point (CFPP, °C) using the following Eqs. (3–12) discussed by [21,22,23,24,25].
where N is the number of carbon–carbon double bonds in fatty acid and Mf is the mass fraction of each fatty acid.
where Ni is the percentage of each FAME, and Mi is the molecular weight of each FAME.
where C16:0, C18:0, C20:0, C22:0, and C24:0 are the weight percentage of the corresponding fatty acids.
2.9 Statistical analysis
The mean and standard deviation (SD) of three replicates were used to express the results. Using the SPSS 23.0 program, the collected data were statistically evaluated by one-way analysis of variance (ANOVA), followed by Duncan’s multiple range testing for data with a significant difference, at p < 0.05.
3 Results and discussion
3.1 Growth and lipids production under different nitrogen sources of the microalga
It was found that M. salina efficiently utilizes organic nitrogen (particularly glycine and YE and, to a lesser degree, urea) and nitrate (NaNO3 and KNO3) for growth, but not ammonium. As shown in Fig. 1, the highest growth of M. salina was observed in Glycine (OD680 value was 2.40) and YE (OD680 value was 2.25) on the 12th day of cultivation followed by potassium nitrate (OD680 value was 1.95), sodium nitrate (OD680 value was 1.85), and urea (OD680 value was 1.39). Cell growth inhibition was observed in culture with ammonium chloride (OD680 value was 0.31).

Growth curves of M. salina grown on different sources of nitrogen sources determined by measuring optical density at 680 nm
The results in Table 1 reveal that YE resulted in the highest biomass and lipid productivity among all six nitrogen sources. This improvement might be caused by the fact that YE contains a variety of other substances in addition to nitrogen, such as amino acids, peptides, vitamins, and carbs. Because M. salina appears to use both nitrogen and organic molecules in YE through mixotrophic metabolism, cell growth and lipid production were enhanced. Glycine, which had the second highest cell concentration and lipids content, can provide the dissolved free amino acids in a culture medium that microalgae can easily utilize [26]. It was reported that Tetraselmis sp. had the maximum cell concentration and lipid productivity under YE, then comes glycine [15]. Although M. salina continued to grow effectively in the presence of urea supplementation, the final cell concentration and lipid output were somewhat lower than those seen when nitrate was present.
On the other hand, it was reported that Isochrysis galbana be able to create the highest cell concentration in urea rather than nitrate or nitrite [27]. Cell growth was inhibited when ammonium chloride was supplemented, showing that the 8.82 mM concentration was toxic to cell growth. The excessive transfer of ammonium to the cells might interfere with the creation of ATP in the chloroplast, inhibiting photosynthesis, which is the cause of the inhibitory effect on cell growth [28]. Additionally, in some cases, excessive ammonium in the medium can cause a significant pH drop by releasing H+ ions, which prevents cell development and may even lead to cell lysis [29, 30].
3.2 Microalgal growth and lipids production under salt stress
As shown in Fig. 2, the highest growth of M. salina was observed at a concentration of 300 mM (OD 680 value was 2.41), followed by a concentration of 200 mM (OD 680 value was 2.19), control (OD 680 value was 1.85), the concentration of 100 mM (OD 680 value was 1.76), and concentration 400 Mm (OD 680 value was 1.66).

Growth curves of M. salina grown on different concentrations of sodium chloride determined by measuring optical density at 680 nm
Table 2 reveals that the best concentration of NaCl for growth and lipids accumulation was 300 Mm. These results are in agreement with that reported by Zulkifli et al. [31], on which 400 mM of NaCl showed the highest cell density of N. oculata as compared to other salinity concentration among other salinity concentrations.
These enhancements are due to some Nannochloropsis species being halophilic, and salinity strongly impacts their cellular physiology. Oxidative stress may be sodium chloride’s primary function in the algal medium, leading to the increment of the concentration of triacylglycerol in the cytoplasm [32, 33] and/or changes in lipid amount and composition [34]. The high lipid content at 300 mM NaCl may be due to adaptation under stress conditions, which help in the accumulation of lipid content in cells. In axenic conditions, salt stress was also used to maintain the culture leading to increased lipid production [35].
According to reports, M. salina’s biomass was maximum between salinities of 22 and 34%, whereas its highest lipid content was found at a salinity of 34% [36]. N. oculata can tolerate salinities between 10 and 35%, and the optimal salinity for their growth was 25% in nutrient-rich environments [37].
3.3 Microalgal growth and lipids production under carbon enrichment
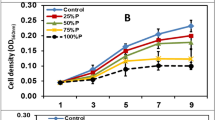
It was found that M. salina effectively took up and metabolized both glucose and glycerol but not acetate. The best concentration of glycerol was 2 g L−1 which showed the highest growth of M. salina (OD 680 value was 2.50) (Fig. 3a), while the best concentration of glucose was 3 g L−1 (OD 680 value was 2.27) (Fig. 3b). Cell growth inhibition was observed in culture using all three concentrations of acetate (Fig. 3c).

Growth curves of M. salina grown on different organic carbon sources determined by measuring optical density at 680 nm. a Different concentrations of glycerol. b Different concentrations of glucose. c Different concentrations of sodium acetate
Obtained results in Table 3 reveal that the final concentration of biomass and lipids accumulations of M. salina were higher than photoautotrophic control when grown mixotrophy on glycerol (2 g L−1) and glucose (3 g L−1). This was not the case when acetate was used. Although many microalgae are known to metabolize acetate [38, 39] easily, M. salina cannot. The results show that M. salina can utilize glycerol more effectively than glucose. Regarding lipid production, glycerol was a better organic carbon source than glucose at its best concentration of 3 g L−1 in the concentration range of (1:3) g L−1. It was found that the maximum biomass productivity in mixotrophic batch cultures using glucose or glycerol was identical at 170 mg L−1 day−1, being 30% higher than in control cultures. The optimal concentrations of the organic carbon sources for the growth of N. gaditana were 5 g L−1 for glucose and 1 g L−1 for glycerol. [40].
3.4 Cultivation of Microchloropsis salina under combined
The optimal level of each effect was used (optimized media) in accordance with the earlier findings, as M. salina was grown in F/2 medium with YE (123.5 mg N L−1), sodium chloride (300 mM), and glycerol (2 g L−1). The obtained results in Fig. 4 show that while the examined microalga’s total lipid content and lipid productivity significantly increased, their dry weight and biomass productivity decreased by 18 and 16%, respectively. M. salina’s lipid content and lipid productivity of M. salina increased by 72 and 80%, respectively.

Cellular dry weight (CDW) and lipids productivity (LP) of M. salina grown under control and combined conditions. a CDW. b LP. Error bar show the SD of three measurements
3.5 Gas chromatography (GC) analysis of fatty acids
The given data in Table 4 and Fig. 5 reveal that the fatty acid profiles differ significantly (quantity and quality level). It was found that SFAs in all conditions are higher than the control as SFAs of culture cultivated in optimized conditions are more than onefold than control. Also, SFAs dramatically increased when the culture was grown on NaCl at a concentration of 300 mM and glycerol at a concentration (2 g L−1) by 45% and 42%, respectively. SFAs moderately increased by 27% when the culture was grown on a nitrogen source (YE). Regardless of the culture’s condition, the main fatty acids were C16:0, C18:1, C18:2, and C18:3 (palmitic, oleic, linoleic, and linolenic acids).

Fatty acids percentage (%) of different treatments of M. salina (SFAs, saturated fatty acids; MUFAs, monounsaturated fatty acids; PUFAs, polyunsaturated fatty acids)
Oleic acid (C18:1) and palmitic acid (C16:0) were higher in the culture with YE, whereas the concentrations of linoleic acid (C18:2) and linolenic acid (C18:3) were lower than in the control culture. It was found that Tetraselmis sp. had linoleic acid (C18:2) and linolenic acid (C18:3) with higher concentrations than the culture with F/2 medium, and stearic acid (C18:0) content was significantly lower than those in the culture with YE [15].
Other fatty acids, such as lauric acid (C12:0), myristic acid (C14:0), pentadecanoic acid (C15:0), cis-10-heptadecanoic acid (C17:1), elaidic acid (C18:1), and linolelaidic acid (C18:2), are also found in addition to the major fatty acids under Nacl concentration 300 Mm. When Tetraselmis suecica cells are cultivated in 0.6 M NaCl, many additional fatty acids, including pentadecanoic acid (C15:0), cis-10-heptadecanoic acid (C17:1), elaidic acid (C18:1), and linolelaidic acid (C18:2), are discovered in addition to the two primary saturated fatty acids [41].
The fatty acid methyl ester profile (FAME) of M. salina grown mixotrophically using glycerol constitutes 86.5% of total fatty acids ranging between 16 and 18 carbon chain lengths which are the most abundant fatty acids needed for biodiesel application. FAME profile shows palmitic acid (C16:0) with a high percentage of 30.3%, followed by oleic acid (18:1) with 25.8%. These findings corroborated those of [42], who discovered that the FAME profile of E. gracilis under mixotrophic cultivation mode using glycerol contributes 93.46% of total fatty acids ranging between 16 and 18 carbon chain lengths and shows the presence of 24.17 and 25.8% of palmitic acid (C16:0) and oleic acid (18:1), respectively. Additionally, according to the C. vulgaris fatty acid profiles, the saturated fatty acid content increased from 16.91% in autotrophic conditions to 34.94% in the medium containing 5 g L−1 glycerol [43].
Under combined conditions, the content of SFAs greatly increased from 23.5 to 48.4%; and palmitic acid (C16:0) and stearic acid are the two main fatty acids (C18:0). It was reported that total fatty acids in N. oceanica increased under the combination of high salinity and high light [44]. In comparison to a single stress condition or a combination of more than three stress elements, lipid synthesis was significantly improved when two stress conditions were present. Although various research [45,46,47] have shown the positive effects of coupled stress factors on fat accumulation, the underlying mechanisms are hardly examined.
3.6 Biodiesel properties
Since saturated fatty acids positively impact transesterification and their biodiesel quality [7], the selected treatments showed high saturated and low unsaturated fatty acid content, which were recommended for good biodiesel quality. By comparing the obtained data with that of American standardization (ASTM D-6751) and European standardization (EN 14,214), cetane number (CN), iodine value (IV), density (ρ), and viscosity (V) which are considered limiting factors for the quality of biodiesel; they fall within the acceptable range (Table 5). The cetane number (CN) is a measure of how long it takes for fuel to ignite in diesel cycle engines. The ignition time decreases with increasing CN. The increase in CN is connected with a longer unbranched carbon chain in the FAME component parts [46].
On the other hand, a lower IV value is advantageous for the manufacture of Biodiesel [47], as this quality is influenced by the number and location of double bonds in the alkyl esters’ carbon chains. There is a greater chance of oxidation, deposit development, and lubricity degradation of the biodiesel with the IV. The density and viscosity of the biodiesel increase with the length of the carbon chain in the methyl esters, and these characteristics will drop as the number of double bonds rises [46]. SV value can reflect the highest fatty acid contents. Saturated fatty acids (SFAs) may also lessen the CFPP properties of biodiesel because they have substantially higher melting points than unsaturated fatty acids (USFAs) [25].
4 Conclusion
Organic nitrogen sources, salt stress, and mixotrophic nutrition by glycerol, either individually or in combination, have significantly improved the productivities of biomass and lipid in M. salina. The best treatment condition included YE as a nitrogen source, 300 mM sodium chloride, and glycerol (g L−1). Individually, YE enhanced lipid content by more than on fold compared to control with only 24% lipids. Also, 300 mM NaCl improved lipids productivity by 92%, and glycerol (g L−1) promoted lipid productivity by 83%. Under the combined condition, lipid productivity increased by 80%, while biomass productivity reduced by 16%.
In all treatments, the fatty acids profile revealed a notable increase in saturated fatty acids, a discernible drop in polyunsaturated fatty acids, and no notable changes in monounsaturated fatty acids. This study provides experimental results on increasing biomass and lipid productivities and improving the fatty acids profile of M. salina for viable biodiesel production. However, further research efforts should be encouraged to achieve the necessary developments to implement this approach on a large scale.
Data availability
All data and materials are supplied in the manuscript.
References
Liming H (2009) Financing rural renewable energy: a comparison between China and India. Renew Sustain Energy Rev 13(5):1096–1103
Rodolfi L, Chini Zittelli G, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol Bioeng 102:100–112
Liu J, Huang JC, Sun Z, Zhong YJ, Jiang Y, Chen F (2011) Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour Technol 102:106–110
Ahmad AL, Yasin NHM, Derek CJC, Lim JK (2011) Microalgae as a sustainable energy source for biodiesel production: a review. Renew Sustain Energy Rev 15:584–593
Knothe G (2009) Improving biodiesel fuel properties by modifying fatty ester composition. Energy Environ Sci 2:759–766
El-Sheekh MM, El-Mohsnawy E, Mabrouk M, Zoheir W (2020) Enhancement of biodiesel production from the green microalga Micractinium reisseri via optimization of cultivation regimes. Journal of Taibah University for Science 14(1):437–444
El-Sheekh M, Abomohra A, Hanelt D (2013) Optimization of biomass and fatty acid productivity of Scenedesmus obliquus as a promising microalga for biodiesel production. World J Microbiol Biotechnol 29(5):915–922
Beydeş H, Karapinar I (2018) Algal nutrient removal from wastewater in fed-batch operated photobioreactor. Int J Environ Res 12:303–311
Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert M, Darzins A (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54:621–639
El-Sheekh MM, Gheda S, El-Sayed AB, Abo Shady A, El-Sheikh M, Schagerl M (2019) Outdoor cultivation of the green microalga Chlorella vulgaris under culture stress conditions as a feedstock for biofuel. Environ Sci Pollut Res 26:18520–18532
Bharte S, Desai K (2019) The enhanced lipid productivity of Chlorella minutissima and Chlorella pyrenoidosa by carbon coupling nitrogen manipulation for biodiesel production. Environ Sci Pollut Res 26:3492–3500
Chandra R, Rohit MV, Swamy Y, Mohan SV (2014) Regulatory function of organic carbon supplementation on biodiesel production during growth and nutrient stress phases of mixotrophic microalgae cultivation. Bioresour Technol 165:279–287
Smith RT, Bangert K, Wilkinson SJ, Gilmour DJ (2015) Synergistic carbon metabolism in a fast-growing mixotrophic freshwater microalgal species Micractinium inermum. Biomass Bioenergy 82:73–86
Guillard RRL (1975) Culture of phytoplankton for feeding marine invertebrates. In: Smith WL, Chanley MH (eds) Culture of marine invertebrate animals. Plenum Press, New York, pp 26–60
Garam K, Ghulam M, Kisay L (2016) Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production Algae 2016, 31(3): 257–266
Ahmed A, Osman ME (1973) The influence of light on 14C02 -fixation by synchronous cultures of Chlorella pyrenoidosa. Egypt J Bot 16:319–327
Abomohra A, Wagner M, El-Sheekh M, Hanelt D (2013) Lipid and total fatty acid productivity in photoautotrophic fresh water microalgae: screening studies towards biodiesel production. J Appl Phycol 25:931–936. https://doi.org/10.1007/s10811-012-9917-y
Jaeyeon P, Hae J, Eun YY, Seung JM (2016) Easy and rapid quantification of lipid contents of marine dinoflagellates using the sulpho-phospho-vanillin method. Algae 31(4):391–340
Folch J, Lees M, Stanley G (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
Zahran HA, Tawfeuk HZ (2019) Physicochemical properties of new peanut (Arachishypogaea L.) varieties. OCL 26:19
Hoekman SK, Broch A, Robbins C, Ceniceros E, Natarajan M (2012) Review of biodiesel composition, properties, and specifications. Renew Sustain Energy Rev 16:143–169
Ma Y, Wang Z, Zhu M, Yu C, Cao Y, Zhang D, Zhou G (2013) Increased lipid productivity and TAG content in Nannochloropsis by heavy-ion irradiation mutagenesis. Bioresour Technol 136:360–367
Nascimento IA, Marques SSI, Cabanelas ITD, Pereira SA, Druzian JI, de Souza CO, Vich DV, de Carvalho GC, Nascimento MA (2013) Screening microalgae strains for biodiesel production: lipid productivity and estimation of fuel quality based on fatty acids profiles as selective criteria. Bioenergy Res 6:1–13
Song M, Pei H, Hu W, Ma G (2013) Evaluation of the potential of 10 microalgal strains for biodiesel production. Bioresour Technol 141:245–251
Francisco ÉC, Neves DB, Jacob-Lopes E, Franco TT (2010) Microalgae as feedstock for biodiesel production: carbon dioxide sequestration, lipid production and biofuel quality. J Chem Technol Biotechnol 85:395–403
Kumar V, Muthuraj M, Palabhanvi B, Ghoshal AK, Das D (2014) High cell density lipid rich cultivation of a novel microalgal isolate Chlorella sorokiniana FC6 IITG in a single-stage fed-batch mode under mixotrophic condition. Bioresour Technol 170:115–124
Fidalgo JP, Cid A, Torres E, Sukenik A, Herrero C (1998) Effects of nitrogen source and growth phase on proximate biochemical composition, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture 166:105–116
Ramanna L, Guldhe A, Rawat I, Bux F (2014) The optimization of biomass and lipid yields of Chlorella sorokiniana when using wastewater supplemented with different nitrogen sources. Bioresour Technol 168:127–135
Li T, Zheng Y, Yu L, Chen S (2013) High productivity cultivation of a heat-resistant microalga Chlorella sorokiniana for biofuel production. Bioresour Technol 131:60–67
Wu LF, Chen PC, Lee CM (2013) The effects of nitrogen sources and temperatures on cell growth and lipid accumulation of microalgae. Int Biodeterior Biodegrad 85:506–510
Zulkifli AF, Ramli A, Lim JW, Lam MK (2018) Effect of NaNO3 and NaCl concentration on Nannochloropsisoculata cell biomass and FAME composition for biodiesel production. IOP Conf Series: J Phys: Conf Series 1123:012071. https://doi.org/10.1088/1742-6596/1123/1/012071
Takagi M (2006) Effect of salt concentration on intracellular accumulation of lipids and triacylglycerides in marine microalgae Dunaliella cells. J Biosci Bioeng. 223–226
Kan G, Shi C, Wang X (2012) Acclimatory responses to high-salt stress in Chlamydomonas (Chlorophyta. Chlorophyceae from Antarctica Acta Oceanol Sin 31:116–124
López-Pérez L, Martínez-Ballesta M, Maurel C (2009) Changes in plasma membrane lipids, aquaporins and proton pump of broccoli roots, as an adaptation mechanism to salinity. Phytochem 70:492–500
Bartley ML, Boeing WJ, Corcoran AA, Holguin FO, Schaub T (2013) Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenerg 54:83–88
Gu H, Nagle N, Pienkos PT, Posewitz MC (2015) Nitrogen recycling from fuel-extracted algal biomass: residuals as the sole nitrogen source for culturing Scenedesmus acutus. Bioresour Technol 184:153–160
Chapman SP, Paget CM, Johnson GN, Schwartz JM (2015) Flux balance analysis reveals acetate metabolism modulates cyclic electron flow and alternative glycolytic pathways in Chlamydomonas reinhardtii. Front Plant Sci 6:474
Smith RT, Bangert K, Wilkinson SJ, Gilmour DJ (2015) Synergistic carbon metabolism in a fast growing mixotrophic freshwater microalgal species Micractinium inermum. Biomass Bioenergy 82:73–86
Menegol T, Romero-Villegas GI, López-Rodríguez M, Navarro-López E, López-RosalesL YC, Cerón-García C, Molina-Grima E (2019) Mixotrophic production of polyunsaturated fatty acids and carotenoids by the microalga Nannochloropsis gaditana. J Appl Phycol 31:2823–2832
Adel WA, Hussein ET (2017) Effect of salinity and pH on fatty acid profile of the green algae Tetraselmissuecica. J Pet Environ Biotechnol 8:3. https://doi.org/10.4172/2157-7463.1000333
Anwesha K, Shrasti V, Monika PR (2017) Glycerol on lipid enhancement and FAME characterization in algae for raw material of biodiesel. International Journal of Renewable Energy Research 7(4):2017
Hee-Jeong C, Sung-Whan Y (2015) Influence of crude glycerol on the biomass and lipid content of microalgae. Biotechnol Biotechnol Equip 29(3):506–513. https://doi.org/10.1080/13102818.2015.1013988
Li ZR, Wakao S, Fischer BB (2009) Niyogi KK (2009) Sensing and responding to excess light. Annu Rev Plant Biol 60:239–260
Yeesang C, Cheirsilp B (2011) Effect of nitrogen, salt, and iron content in the growth medium and light intensity on lipid production by microalgae isolated from freshwater sources in Thailand. Bioresour Technol 102:3034–3040
Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006) Optimization of Nannochloropsis oculata growth using the response surface method. J Chem Technol Biotechnol 81:1049–1056
Knothe G (2005) Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process Technol 86:1059–1070
Yodsuwan N, Sawayama S, Sirisansaneeyakul S (2017) Effect of nitrogen concentration on growth, lipid production and fatty acid profiles of the marine diatom Phaeodactylum tricornutum. Agric Nat Resour 51:190–197
Acknowledgements
The authors would like to thank the Academy of Scientific Research & Technology (ASRT) for the fellowship awarded to Heba Mansour. The authors greatly appreciate the efforts of Dr. Mostafa Elshobary, Botany Department, Faculty of Science, Tanta University, Tanta, Egypt, for his valuable advice.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
Mostafa El-Sheekh: supervised the work, writing, and editing of the final manuscript, Heba Mansour: carried out the experimental work and prepared the draft of the manuscript, Mohamed Bedaiwy: supervised the work, Rania Elshenoudy: helped in the experimental and draft.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
El-Sheekh, M.M., Mansour, H.M., Bedaiwy, M.Y. et al. Influence of nutrient supplementation and stress conditions on the biomass and lipid production of Microchloropsis salina for biodiesel production. Biomass Conv. Bioref. (2022). https://doi.org/10.1007/s13399-022-03434-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13399-022-03434-9