
Abstract
High-risk human papillomaviruses are well-established drivers of several cancer types including cervical, head and neck, penile as well as anal cancers. While the E6 and E7 viral oncoproteins have proven to be critical for malignant transformation, evidence is also beginning to emerge suggesting that both host pathways and additional viral genes may also be pivotal for malignant transformation. Here, we focus on the role of host APOBEC genes, which have an important role in molecular editing including in the response to the viral DNA and their role in HPV-driven carcinogenesis. Further, we also discuss data developed suggesting the existence of HPV-derived miRNAs in HPV + tumors and their potential role in regulating the host transcriptome. Collectively, while recent advances in these two areas have added complexity to the working model of papillomavirus-induced oncogenesis, these discoveries have also shed a light onto new areas of research that will be required to fully understand the process.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The human papillomavirus (HPV) is an 8 kb double-stranded circular DNA virus that belongs to the large family of papillomaviruses (PVs). Currently, there are known more than 200 types of HPV but only a subgroup is predicted to cause cancer. These are classified by the International Agency for Research on Cancer (Wild et al. 2020) as high-risk types of HPV (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 63, 73, and 82) and probable cancer-related types (26, 53, and 66). The low-risk viruses can cause warts (1 and 2) or benign changes within genital organs (6, 11, 40, 42, 43, 44, 54, 61, 70, 81) with no or poorly proven relation to human pathology (Clifford et al. 2003; Muñoz et al. 2003; Burd 2003; Kiwerska et al. 2019). The high-risk viruses are well known for cervical cancer pathogenesis where almost 100% of the tumors are attributed to the oncogenic activity of the HPV 16 and 18. However, HPV-related tumor pathogenesis is observed also in other sites like the anus (90% of cases), tumors of the vagina or penis (50–75% of cases), and the head and neck region (about 10% of cases) (de Martel et al. 2020; Serrano et al. 2018; Wierzbicka et al. 2021). The NIH estimates that worldwide, high-risk HPV infections cause about 5% of all cancers emphasizing on one hand the importance of papillomaviruses in the pathogenesis of human tumors and on the other the relevance of HPV vaccination („https://www.cancer.gov/”, n.d.).
HPV 16-driven oncogenesis
Canonical oncogenic mechanism that involves the E5, E6, and E7 oncoproteins
The main mechanism of HPV 16 and 18-driven oncogenesis is already well known, and for its explanation, Harald zur Hausen was awarded the Nobel Prize in Medicine in 2008 (Dürst et al. 1983; Boshart et al. 1984). The main players in this process are the oncoproteins E6 and E7 encoded by the viral genome (Münger et al. 1989). The expression of the E6 and E7 proteins is initiated after infection from the episomal form albeit on low levels because of the inhibition by the viral E2 protein. The proteins become significantly higher expressed after integration of the virus into the host genome which requires the transition from the circular to linear form resulting in the disruption of the E2 gene (Janicek and Averette 2001). The E6 and E7 proteins interact with crucial proteins of the cellular machinery responsible for cell cycle regulation and tumor suppression (Z.-A.-S. Ghoreshi 2023). E6 binds the cellular TP53 protein of well-known suppressor function triggering its ubiquitination and subsequent degradation by the E6AP ligase activity (E6-associated protein ligase) (Scheffner et al. 1993). This leads in consequence to loss of the regulatory function of TP53 towards the cell cycle and reduced expression of proapoptotic proteins such as BAX which is physiologically mediated by TP53 (Martinez-Zapien et al. 2016; Fischer 2017; Ruttkay-Nedecky et al. 2013). On the other hand, the E7 protein interacts with the cellular RB1 protein leading to the release of the potent transcription factor E2F/DP that in the G1 phase is normally bound to RB1 and inactive. The viral protein mimics therefore the physiological conditions of the late G1 phase where RB1 is inactivated by phosphorylation what leads to E2F/DP release (Rubin 2013). The E2F/DP factor drives the transcription of several genes responsible for the G1 → S transition, accelerates the cell cycle, and leads to proliferation. Eventually, the activity of the oncoproteins triggers the process of oncogenesis (Fig. 1).

Oncogenic activity of the E6 and E7 proteins encoded by HPV 16 (based on Janicek and Averette (2001) and edited)
Several cancer-related pathways are involved in this process. PI3K/AKT/mTOR is a well-known oncogenic pathway induced by E6 and E7 in head and neck cancer (Molinolo et al. 2009). These viral oncoproteins are able to active Akt pathway via upstream regulators, such as RPTK (receptor protein tyrosine kinase) and PI3K (phosphoinositide 3-kinase) resulting in increased proliferation (Aguayo et al. 2023).
Furthermore, E6 and E7 oncoproteins activate the Wnt pathway causing the accumulation of β-catenin, which may increase transcription of cell proliferation genes. This effect may be associated with a decreased protein level of β-catenin degradation inducer—Siah-1 (Bello et al. 2015). Moreover, accumulation of nuclear β-catenin is found in human HPV-positive cancers (Shinohara et al. 2001).
Similarly, NOTCH is affected in HPV-infected cells. The influence of E6 oncoprotein on NOTCH pathway regulation occurs through disruption of the interaction of MAML (mastermind-like protein) and NICD proteins, a critical component of the Notch signaling pathway. This results in turn in the loss of NOTCH target gene expression, including HEY and HES, which function as potent transcriptional repressors (Das et al. 2021; Galloway and Laimins 2015).
Another interesting aspect of E6 oncogenicity is the ability to interact with cellular proteins containing PDZ domains, such as PSD-95/Dlg/ZO-1 and Scribble, via the PBM motif (PDZ domain binding motif), presents in the C-terminus of the protein. This interaction disrupts intercellular connections between epithelial cells leading to loss of cell polarity which is a known characteristic of neoplastic cells (Gardiol et al. 1999).
In addition to the oncogenic potential of the E6 and E7 proteins, viral integration into the host genome directly triggers genomic instability (Akagi et al. 2014). Moreover, the introduced structural abnormalities may result in the disruption of tumor suppressor genes in the host genome and possibly also upregulate the expression of oncogenes (Schmitz et al. 2012; Lu et al. 2014) and reviewed in Chen et al. (2017) thus altering the function of several tumor-related genes.
Interestingly, one of the factors associated with increased risk of HPV-related malignant transformation may derive from bacterial or viral infections which can trigger alterations in the E6 oncoprotein expression. In this regard, Szostek et al. showed that the presence of Ureaplasma urealyticum–stimulated E6 expression in SiHa cells derived from human cervical squamous cell carcinoma cell line (Szostek et al. 2014).
Although the most known HPV-associated oncoproteins are E6 and E7, the E5 oncoprotein also plays a role in carcinogenesis. Recent studies have confirmed the importance of E5 oncoprotein in the modulation of the immune system and cell transformation. E5 cooperates with E7 and E6 to fuel the development of malignant cells (de Freitas et al. 2017). E5 oncoprotein inhibits apoptosis by increasing the ubiquitination and proteasomal degradation of the pro-apoptotic protein BAX consequently promoting the accumulation of cells with mutations (Maufort et al. 2010). Moreover, as shown for HPV 18, E5 potentiates EGFR signaling leading to increased ERK/MAPK activity and via this pathway is implicated in cell cycle progression. An important function of E5 oncoprotein was also confirmed in the metastatic process and is mediated by the upregulation of the MET oncoprotein (Scott et al. 2018; Hemmat and Baghi 2018).
The non-canonical oncogenic mechanism related to the function of APOBEC enzymes
Recent reports suggest that the canonical mechanism of HPV-driven oncogenesis is not the only mechanism by which the virus contributes to tumor formation. The non-canonical mechanism of viral (including HPV) oncogenicity is related to the function of the APOBEC—apolipoprotein B mRNA editing enzymes. These enzymes belong to the class of APOBEC3s (A3A, A3B, A3C, A3DE, A3F, A3G, A3H) deaminases which become activated in the cell by an interferon-mediated signal cascade triggered by sensing foreign nucleic acids by the protein machinery of the cell (Stenglein et al. 2010). Several proteins like TLR9 and DAI, AIM2, and RNA polymerase III have been identified to sense single-stranded and double-stranded DNA, respectively (Uematsu and Akira 2007; Takaoka et al. 2007; Bürckstümmer et al. 2009; Chiu et al. 2009). Innate immune sensors play a crucial role in recognizing viral nucleic acids, including those from HPV viruses. They enable a rapid immune response to infections by secreting interferon (IFN), regulating IFN-stimulated genes, and activation of the inflammasome complex. For example, the nuclear DNA sensor IFI16 detects single and double-strand DNA particles upon which it moves to the cytoplasm and activates the TBK1-IRF3 and the pro-inflammatory NFκB signaling pathway. Interestingly, it contributes further to IFN expression through interaction with STING1, a major regulator of the innate immune response. It has been recently shown that STING1 can impact virtually all aspects of tumorigenesis (Z. Ghoreshi et al. 2022; Samson and Ablasser 2022).
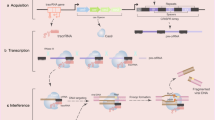
The overexpressed APOBEC enzymes mediate the deamination of cytidines to uridines in the foreign DNA. As uridines are atypical DNA nucleosides, they are recognized by the UNG2 glycosylase and turned to abasic lesions leading to downstream degradation of the foreign DNA by uracil excision mechanism (Stenglein et al. 2010) (Fig. 2). In fact, it has been demonstrated that this class of enzymes edits HPV DNA in infected cells (Z. Wang et al. 2014). Together, this formulates a protective mechanism that prevents DNA transmission between species.

Degradation of foreign genetic material by the APOBEC deaminases (based on Stenglein et al. (2010) and edited)
As the C → T transition is the predominant mutation in cancer genomes (Greenman et al. 2007), it allows to speculate that the virus-induced APOBEC enzymes might erroneously generate mutations in the genome of the host cell promoting malignant transformation (Suspène et al. 2011). In line with this presumption, the AID deaminase that is physiologically active during B-cell maturation in the germinal centers and mediates the process of somatic hypermutation (Maul and Gearhart 2010) was found involved in malignant transformation not only in B-cell lymphomas but also in nonlymphoid tumors (Okazaki et al. 2007). Moreover, the presence of typical mutations caused by deamination of cytosines introduced by APOBEC enzymes in a characteristic nucleotide context and thus having an APOBEC mutational signature has been demonstrated in genomes of multiple tumors (Alexandrov et al. 2013; Roberts et al. 2013). Interestingly, besides lymphoid neoplasms, APOBEC mutation signatures have been recognized in all HPV-related tumors including cervical cancers, esophageal cancers, or head and neck cancers suggesting an important role of this pathogenetics mechanism in these solid tumors. In fact, the frequent activating mutations E542K and E545K of the PIK3CA oncogene can be to a large extent attributed to the APOBEC-driven mutagenesis (Henderson et al. 2014) and reviewed in Chen et al. (2017). Faden et al. observed recently that APOBEC was the dominant mutational signature in somatic exomes in 79 HPV 16 + oropharyngeal squamous cell carcinoma cases analyzed. The authors assessed APOBEC mutational load in human and viral genomes concluding that there is a direct link in host and viral genome APOBEC-driven mutagenesis during infection. These provide an important link concerning APOBEC mutagenesis in host and virus and support a common mechanism driving APOBEC dysregulation (Faden et al. 2021).
It has been observed that in head and neck cancer patients, the APOBEC3B expression was significantly associated with treatment outcome (Janecka-Widła et al. 2021). In details, patients with high APOBEC3B expression detected in tumors had over two times higher risk of cancer progression than those with lower levels of this protein. In line with this observation, in the subgroup of HPV 16-positive HNSCC patients, those patients having tumors with low APOBEC3B expression survived significantly longer than those with high APOBEC3B expression (Janecka-Widła et al. 2021). Moreover, APOBEC3B overexpression differentiated HPV-negative low-grade oral epithelial dysplasia, which showed intermediate APOBEC3B expression, from oral cancers. The highest levels were found in grade II and III oral cancers suggesting that APOBEC3B may be used as a marker for advanced HPV-positive cancers (Argyris et al. 2021).
Thus, the erroneous functioning of the APOBEC enzymes induced by HPV infections may be an important mechanism of HPV oncogenicity, strictly related to the canonical mechanism described above. The continuous expression of oncoprotein E6 seems to be pivotal for APOBEC3B overexpression in HPV-associated cancers (Vieira et al. 2014). In line with this finding, Mori et al. demonstrated that E6-mediated degradation of the TP53 protein induces expression of the TEAD transcription factor, which is a positive regulator of APOBEC3B transcription (Mori et al. 2017). In addition, TEAD stabilizes the Yes-associated protein (YAP), an oncogenic component of the Hippo pathway (He et al. 2015).
The role of APOBEC enzymes in HPV-driven oncogenesis extends beyond the canonical mechanisms contributing to malignant transformation. Recent studies highlight the complexity and significance of APOBEC-related pathways in HPV-associated cancers; however, this non-canonical mechanism is likely secondary to the dominant mechanism based on HPV oncoproteins.
The putative oncogenic mechanism related to viral miRNAs
Virus-encoded miRNAs not only show auto-regulatory function but have also the potential to interfere with various physiological functions of the infected cell. Viral miRNAs are found in several viral genomes including herpesviruses (Grundhoff and Sullivan 2011; Dass et al. 2023), adenoviruses (Xu et al. 2007), polyomaviruses (Sullivan et al. 2005), and also plant viruses (Sharma and Singh 2016). Such viral miRNAs have been identified in EBV (Epstein-Barr virus) and HCMV (cytomegalovirus) (Pfeffer et al. 2004; Kuzembayeva et al. 2012). It has been shown that the encoded miRNAs have important functions in the process of infection and malignant transformation. For example, in EBV-infected B-cells viral miRNAs reduce immunogenicity by suppressing proinflammatory cytokines such as IL-12. This results in reduced recruitment of CD4( +) effector T cells and subsequent impaired elimination of the infected B-cells (Tagawa et al. 2016). The expression of EBV miRNAs is also brought together with worse survival of EBV-associated gastric cancer (Kang et al. 2017). Furthermore, EBV-encoded miRNAs contribute to Endemic Burkitt lymphoma clinical presentation, progression, and poor outcome (Oduor et al. 2017). Moreover, viral miRNAs were shown to contribute to cell cycle deregulation by posttranscriptional silencing of cell cycle-related genes like CCNE2 (cyclin E2), BRCC3, and EID1 targeted by miR-US25-1 encoded by the HCMV genome (Grey et al. 2010).
In light of the presented findings, it is intriguing whether HPV genomes harbor miRNAs of similar functionalities. A decade ago, two manuscripts were published wherein the authors bioinformatically as well as experimentally demonstrate that the HPV 16 genome encodes beside protein coding also miRNA genes that are expressed, albeit on low level, in human cervical lesions (Qian et al. 2013; Virtanen et al. 2016). The authors identified five miRNAs HPV 16-miR-H1, 2, 3, 5, and 6 dispersed in the viral genome and encoded on the plus or minus strand (Table 1, Fig. 3). These findings have been further validated in parallel studies of other groups (Thakur 2015) and by bioinformatic analysis of NGS data (Weng et al. 2018). Several functions promoting viral infection have been implicated for these miRNAs; however, experimentally validated data demonstrating the viral miRNA:host mRNA interaction is still lacking. Viral HPV 16-miR-H1-1 and miR-H2-1 were predicted to target several genes involved in immune system regulation and development. HPV 16-miR-H1-1 via putative targeting of BCL11A, CHD7, ITGAM, RAG1, and TCEA1 genes is able to inhibit T-cell activation leading to the inhibition of immune system development. Similarly, miR-H2-1 potentially inhibits genes involved in T-cell activation like PKNOX1, SP3, and XRCC4 as well as immune system development (JAK2, PKNOX1, SP3, XRCC4, FOXP1) (Qian et al. 2013). Also, typical cancer-related tumor-suppressive genes are predicted targets of H1 including CAV2, PTEN, and SEMA3F (Qian et al. 2013). The same holds true for the other delineated HPV miRNAs as reviewed by Gallo et al. (Gallo et al. 2020). Gutiérrez et al. identified approximately 900 target genes potentially regulated by HPV-encoded miRNAs. Within this cohort, the largest sub-group, that is 15% of the genes, was involved in cell proliferation followed by regulation of ribosomes (11%) and translation (11%) (Gutiérrez et al. 2018).

MiRNAs putatively encoded by the HPV 16 genome (based on Table 1)
Therefore, the cited literature data allow to carefully speculate that the putative HPV-encoded miRNAs, if real, might be involved in three crucial processes in the infected cell. That is (I) immune escape, (II) deregulation of cell cycle and attenuation of tumor suppressor genes leading to increased proliferation, and (III) takeover of the control of the cell’s transcriptional machinery. However, it needs to be stressed that the putative miRNA-driven oncogenic mechanism has a significantly inferior role compared to the function of the oncogenic virus-encoded proteins. Moreover, the functional validation of many of these findings is lacking; the proposed mechanism is speculative and not supported by direct target validation.
Controversies
Although several studies reported the identification of HPV-encoded miRNAs, mainly in human cervical lesions as reviewed before, other authors came to opposite conclusions. Wang et al. by cloning and sequencing of a small RNA library from the HPV 16 + CaSki cell line identified 174 host miRNAs but none of the reported HPV 16 miRNAs and therefore concluded that the HPV 16 virus in the analyzed cell line does not express viral miRNAs (X. Wang et al. 2008). Similarly, Cai et al. reported no viral miRNAs to be expressed from the HPV 31 genome, and Lui et al. reported the lack of HPV-encoded miRNAs in direct sequencing of HPV-positive cervical cancer cell lines (Lui et al. 2007; Cai et al. 2006).
Interestingly, Chirayil et al. developed an approach based on a library of overlapping genomic segments from an analyzed genome subcloned behind a RNA polymerase II promoter resulting in forced expression of the putative miRNAs (miDGE—miRNA discovery by forced genomic expression) (Chirayil et al. 2018). Thereafter, miRNA candidates were identified using small RNA sequencing and downstream assays. Using this approach, the authors screened 73 different PV genomes (most were human-PVs) and identified five viral miRNA candidates, 1 in HPV 17, 1 in HPV 37, 1 in HPV 41, and 2 in FcPV1. However, no miRNA candidates were identified in the genomes of high-risk HPV 16, 18, and others. The authors further screened transcriptomes of cervical carcinomas deposited in The Cancer Genome Atlas (TCGA) (Cancer Genome Atlas Research Network et al. 2017), but similarly did not observe expression of any miRNA from the high-risk HPV genomes (Chirayil et al. 2018). Another argument against the existence of HPV-encoded miRNAs is the fact that none of the predicted top putative mRNA targets suggested in the cited studies like BCL11A, ITGAM, RAG1, CAV2, PTEN, and SEMA3F was identified in the miRNA target search using miRDB performed by us (Table 1) thus showing large inconsistencies between studies.
Altogether, the reviewed findings raise several unanswered questions. What are the apparently HPV-derived sequences found by various groups? Why are the results of different researcher groups not reproduced? Might they derive from co-infection by other PVs or are they only inactive sequences found in the host genome that are remains from past infections? Assuming that these sequences do not encode functional miRNAs, why do they show such enrichment in potential targets encoding important tumor-related genes hardly possible by pure chance? Are these putative miRNAs expressed only at specific time points in the viral life cycle? Or why should high-risk HPVs, in contrast to several low-risk PVs and other viruses, lack miRNAs that facilitate the process of infection and immune evasion? What was the selective force during the evolutionary process? These and other questions need to be answered in further studies.
Conclusions
The mechanism of HPV-related carcinogenesis is much more complex than initially assumed. Since the discovery of the viral oncoproteins in the 80 s of the twentieth century, we witnessed the accumulation of experimental data which have significantly expanded the picture of how the virus infects the cell, takes over the control of its transcription machinery, and eventually leads to tumor formation. In this process, the function of the virus-encoded oncoproteins is supported by triggered genomic instability, accelerated mutagenesis via the APOBEC enzymes, and perhaps also by yet not fully understood viral miRNA sequences.
References
Aguayo F, Perez-Dominguez F, Osorio JC, Oliva C, Calaf GM (2023) PI3K/AKT/mTOR signaling pathway in HPV-driven head and neck carcinogenesis: therapeutic implications. Biology 12(5):672. https://doi.org/10.3390/biology12050672
Akagi K, Li J, Broutian TR, Padilla-Nash H, Xiao W, Jiang B, Rocco JW et al (2014) Genome-wide analysis of HPV integration in human cancers reveals recurrent, focal genomic instability. Genome Res 24(2):185–99. https://doi.org/10.1101/gr.164806.113
Alexandrov LB, Nik-Zainal S, Wedge DC, Aparicio SAJR, Behjati S, Biankin AV, Bignell GR et al (2013) Signatures of mutational processes in human cancer. Nature 500(7463):415–21. https://doi.org/10.1038/nature12477
Argyris PP, Wilkinson PE, Jarvis MC, Magliocca KR, Patel MR, Vogel RI, Gopalakrishnan R, Koutlas IG, Harris RS (2021) Endogenous APOBEC3B overexpression characterizes HPV-positive and HPV-negative oral epithelial dysplasias and head and neck cancers. Mod Pathol Off J U S Can Acad Pathol Inc 34(2):280–90. https://doi.org/10.1038/s41379-020-0617-x
Bello JOM, Nieva LO, Paredes AC, Gonzalez AMF, Zavaleta LR, Lizano M (2015) Regulation of the Wnt/β-catenin signaling pathway by human papillomavirus E6 and E7 oncoproteins. Viruses 7(8):4734–55. https://doi.org/10.3390/v7082842
Boshart M, Gissmann L, Ikenberg H, Kleinheinz A, Scheurlen W, zur Hausen H (1984) A new type of papillomavirus DNA, its presence in genital cancer biopsies and in cell lines derived from cervical cancer. EMBO J 3(5):1151–57. https://doi.org/10.1002/j.1460-2075.1984.tb01944.x
Bürckstümmer T, Baumann C, Blüml S, Dixit E, Dürnberger G, Jahn H, Planyavsky M et al (2009) An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nat Immunol 10(3):266–72. https://doi.org/10.1038/ni.1702
Burd EM (2003) Human papillomavirus and cervical cancer. Clin Microbiol Rev 16(1):1–17. https://doi.org/10.1128/CMR.16.1.1-17.2003
Cai X, Li G, Laimins LA, Cullen BR (2006) Human papillomavirus genotype 31 does not express detectable microRNA levels during latent or productive virus replication. J Virol 80(21):10890–93. https://doi.org/10.1128/JVI.01175-06
Cancer Genome Atlas Research Network, Albert Einstein College of Medicine, Analytical Biological Services, Barretos Cancer Hospital, Baylor College of Medicine, Beckman Research Institute of City of Hope, Buck Institute for Research on Aging et al (2017) Integrated genomic and molecular characterization of cervical cancer. Nature 543(7645):378–84. https://doi.org/10.1038/nature21386
Chen L, Qiu X, Zhang N, Wang Y, Wang M, Li D, Wang L, Du Y (2017) APOBEC-mediated genomic alterations link immunity and viral infection during human papillomavirus-driven cervical carcinogenesis. Biosci Trends 11(4):383–88. https://doi.org/10.5582/bst.2017.01103
Chirayil R, Kincaid RP, Dahlke C, Kuny CV, Dälken N, Spohn M, Lawson B, Grundhoff A, Sullivan CS (2018) Identification of virus-encoded microRNAs in divergent papillomaviruses. PLoS Pathog 14(7):e1007156. https://doi.org/10.1371/journal.ppat.1007156
Chiu Y-H, Macmillan JB, Chen ZJ (2009) RNA polymerase III detects cytosolic DNA and induces type I interferons through the RIG-I pathway. Cell 138(3):576–91. https://doi.org/10.1016/j.cell.2009.06.015
Clifford GM, Smith JS, Plummer M, Muñoz N, Franceschi S (2003) Human papillomavirus types in invasive cervical cancer worldwide: a meta-analysis. Br J Cancer 88(1):63–73. https://doi.org/10.1038/sj.bjc.6600688
Das T, Zhong R, Spiotto MT (2021) Notch signaling and human papillomavirus-associated oral tumorigenesis. Adv Exp Med Biol 1287:105–22. https://doi.org/10.1007/978-3-030-55031-8_8
Dass D, Dhotre K, Chakraborty M, Nath A, Banerjee A, Bagchi P, Mukherjee A (2023) miRNAs in herpesvirus infection: powerful regulators in small packages. Viruses 15(2):429. https://doi.org/10.3390/v15020429
de Freitas AC, de Oliveira THA, Barros MR, Venuti A (2017) hrHPV E5 oncoprotein: immune evasion and related immunotherapies. J Exp Clin Cancer Res: CR 36(1):71. https://doi.org/10.1186/s13046-017-0541-1
de Martel C, Georges D, Bray F, Ferlay J, Clifford GM (2020) Global burden of cancer attributable to infections in 2018: a worldwide incidence analysis. Lancet Glob Health 8(2):e180-90. https://doi.org/10.1016/S2214-109X(19)30488-7
Dürst M, Gissmann L, Ikenberg H, zur Hausen H (1983) A papillomavirus DNA from a cervical carcinoma and its prevalence in cancer biopsy samples from different geographic regions. Proc Natl Acad Sci USA 80(12):3812–15. https://doi.org/10.1073/pnas.80.12.3812
Faden DL, Kuhs KAL, Lin M, Langenbucher A, Pinheiro M, Yeager M, Cullen M et al (2021) APOBEC mutagenesis is concordant between tumor and viral genomes in HPV-positive head and neck squamous cell carcinoma. Viruses 13(8):1666. https://doi.org/10.3390/v13081666
Fischer M (2017) Census and evaluation of P53 target genes. Oncogene 36(28):3943–3956. https://doi.org/10.1038/onc.2016.502
Gallo A, Miceli V, Bulati M, Iannolo G, Contino F, Conaldi PG (2020) Viral miRNAs as active players and participants in tumorigenesis. Cancers 12(2):358. https://doi.org/10.3390/cancers12020358
Galloway DA, Laimins LA (2015) Human papillomaviruses: shared and distinct pathways for pathogenesis. Curr Opin Virol 14(październik):87–92. https://doi.org/10.1016/j.coviro.2015.09.001
Gardiol D, Kühne C, Glaunsinger B, Lee SS, Javier R, Banks L (1999) Oncogenic human papillomavirus E6 proteins target the discs large tumour suppressor for proteasome-mediated degradation. Oncogene 18(40):5487–96. https://doi.org/10.1038/sj.onc.1202920
Ghoreshi Z-A-S (2023) The role of DNA viruses in human cancer. Cancer Inform 22:11769351231154186. https://doi.org/10.1177/11769351231154186
Ghoreshi Z-A, Nakhaee M, Samie M, Zak MS, Arefini N (2022) Innate immune sensors for detecting nucleic acids during infection. J Lab Med. https://doi.org/10.1515/labmed-2021-0173
Greenman C, Stephens P, Smith R, Dalgliesh GL, Hunter C, Bignell G, Davies H et al (2007) Patterns of somatic mutation in human cancer genomes. Nature 446(7132):153–58. https://doi.org/10.1038/nature05610
Grey F, Tirabassi R, Meyers H, Wu G, McWeeney S, Hook L, Nelson JA (2010) A viral microRNA down-regulates multiple cell cycle genes through mRNA 5’UTRs. PLoS Pathog 6(6):e1000967. https://doi.org/10.1371/journal.ppat.1000967
Grundhoff A, Sullivan CS (2011) Virus-encoded microRNAs. Virology 411(2):325–43. https://doi.org/10.1016/j.virol.2011.01.002
Gutiérrez DA, Varela-Ramírez A, Rodríguez-Esquivel M, Mendoza-Rodríguez MG, Ayala-Sumuano JT, Pineda D, Garrido-Guerrero E et al (2018) Predicting human miRNA-like sequences within human papillomavirus genomes. Arch Med Res 49(5):323–34. https://doi.org/10.1016/j.arcmed.2018.10.008
He C, Mao D, Hua G, Lv X, Chen X, Angeletti PC, Dong J et al (2015) The Hippo/YAP pathway interacts with EGFR signaling and HPV oncoproteins to regulate cervical cancer progression. EMBO Mol Med 7(11):1426–49. https://doi.org/10.15252/emmm.201404976
Hemmat N, Baghi HB (2018) Human papillomavirus E5 protein, the undercover culprit of tumorigenesis. Infect Agents Cancer 13:31. https://doi.org/10.1186/s13027-018-0208-3
Henderson S, Chakravarthy A, Su X, Boshoff C, Fenton TR (2014) APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep 7(6):1833–41. https://doi.org/10.1016/j.celrep.2014.05.012
https://www.cancer.gov/ (n.d.)
Janecka-Widła A, Majchrzyk K, Mucha-Małecka A, Biesaga B (2021) EGFR/PI3K/Akt/mTOR pathway in head and neck squamous cell carcinoma patients with different HPV status. Pol J Pathol: Off J Pol Soc Pathol 72(4):296–314. https://doi.org/10.5114/pjp.2021.113073
Janicek MF, Averette HE (2001) Cervical cancer: prevention, diagnosis, and therapeutics. CA: Cancer J Clin 51(2):92–114. https://doi.org/10.3322/canjclin.51.2.92. quiz 115–18
Kang BW, Choi Y, Kwon OK, Lee SS, Chung HY, Yu W, Bae HI et al (2017) High level of viral microRNA-BART20–5p expression is associated with worse survival of patients with epstein-barr virus-associated gastric cancer. Oncotarget 8(9):14988–94. https://doi.org/10.18632/oncotarget.14744
Kiwerska K, Jozefiak A, Markowska J, Kedzia W, Jackowska J, Wierzbicka M (2019) Oral-genital human papillomavirus infection in Polish couples: frequent detection of HPV 42. BMC Infect Dis 19(1):122. https://doi.org/10.1186/s12879-018-3645-0
Kuzembayeva M, Chiu Y-F, Sugden B (2012) Comparing proteomics and RISC immunoprecipitations to identify targets of Epstein-Barr viral miRNAs. PloS One 7(10):e47409. https://doi.org/10.1371/journal.pone.0047409
Li S-C, Shiau C-K, Lin W-C (2008) Vir-Mir Db: prediction of viral microRNA candidate hairpins. Nucleic Acids Res 36(Database issue):D184-189. https://doi.org/10.1093/nar/gkm610
Lu X, Lin Q, Lin M, Duan P, Ye L, Chen J, Chen X, Zhang L, Xue X (2014) Multiple-integrations of HPV16 genome and altered transcription of viral oncogenes and cellular genes are associated with the development of cervical cancer. PloS One 9(7):e97588. https://doi.org/10.1371/journal.pone.0097588
Lui W-O, Pourmand N, Patterson BK, Fire A (2007) Patterns of known and novel small RNAs in human cervical cancer. Cancer Res 67(13):6031–43. https://doi.org/10.1158/0008-5472.CAN-06-0561
Martinez-Zapien D, Ruiz FX, Poirson J, Mitschler A, Ramirez J, Forster A, Cousido-Siah A et al (2016) Structure of the E6/E6AP/P53 complex required for HPV-mediated degradation of P53. Nature 529(7587):541–45. https://doi.org/10.1038/nature16481
Maufort JP, Shai A, Pitot HC, Lambert PF (2010) A role for HPV16 E5 in cervical carcinogenesis. Cancer Res 70(7):2924–31. https://doi.org/10.1158/0008-5472.CAN-09-3436
Maul RW, Gearhart PJ (2010) AID and somatic hypermutation. Adv Immunol 105:159–91. https://doi.org/10.1016/S0065-2776(10)05006-6
Molinolo AA, Amornphimoltham P, Squarize CH, Castilho RM, Patel V, Gutkind JS (2009) Dysregulated molecular networks in head and neck carcinogenesis. Oral Oncol 45(4–5):324–34. https://doi.org/10.1016/j.oraloncology.2008.07.011
Mori S, Takeuchi T, Ishii Y, Yugawa T, Kiyono T, Nishina H, Kukimoto I (2017) Human papillomavirus 16 E6 upregulates APOBEC3B via the TEAD transcription factor. J Virol 91(6):e02413-16. https://doi.org/10.1128/JVI.02413-16
Münger K, Phelps WC, Bubb V, Howley PM, Schlegel R (1989) The E6 and E7 genes of the human papillomavirus type 16 together are necessary and sufficient for transformation of primary human keratinocytes. J Virol 63(10):4417–21. https://doi.org/10.1128/JVI.63.10.4417-4421.1989
Muñoz N, Bosch FX, de Sanjosé S, Herrero R, Castellsagué X, Shah KV, Snijders PJF, Meijer CJLM, International Agency for Research on Cancer Multicenter Cervical Cancer Study Group (2003) Epidemiologic classification of human papillomavirus types associated with cervical cancer. N Engl J Med 348(6):518–27. https://doi.org/10.1056/NEJMoa021641
Oduor CI, Movassagh M, Kaymaz Y, Chelimo K, Otieno J, Ong’echa JM, Moormann AM, Bailey JA (2017) Human and Epstein-Barr virus miRNA profiling as predictive biomarkers for endemic Burkitt lymphoma. Front Microbiol 8:501. https://doi.org/10.3389/fmicb.2017.00501
Okazaki I-m, Kotani A, Honjo T (2007) Role of AID in tumorigenesis. Adv Immunol 94:245–73. https://doi.org/10.1016/S0065-2776(06)94008-5
Pfeffer S, Zavolan M, Grässer FA, Chien M, Russo JJ, Ju J, John B et al (2004) Identification of virus-encoded microRNAs. Science (New York, N.Y.) 304(5671):734–36. https://doi.org/10.1126/science.1096781
Qian K, Pietilä T, Rönty M, Michon F, Frilander MJ, Ritari J (2013) Identification and validation of human papillomavirus encoded microRNAs. PLoS One 8(7):e70202
Qureshi A, Thakur N, Monga I, Thakur A, Kumar M (2014) VIRmiRNA: a comprehensive resource for experimentally validated viral miRNAs and their targets. Database: J Biol Databases Curation 2014:bau103. https://doi.org/10.1093/database/bau103
Roberts SA, Lawrence MS, Klimczak LJ, Grimm SA, Fargo D, Stojanov P, Kiezun A et al (2013) An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat Genet 45(9):970–76. https://doi.org/10.1038/ng.2702
Rubin SM (2013) Deciphering the retinoblastoma protein phosphorylation code. Trends Biochem Sci 38(1):12–19. https://doi.org/10.1016/j.tibs.2012.10.007
Ruttkay-Nedecky B, Jimenez Jimenez AM, Nejdl L, Chudobova D, Gumulec J, Masarik M, Adam V, Kizek R (2013) Relevance of infection with human papillomavirus: the role of the P53 tumor suppressor protein and E6/E7 zinc finger proteins (Review). Int J Oncol 43(6):1754–62. https://doi.org/10.3892/ijo.2013.2105
Samson N, Ablasser A (2022) The cGAS–STING pathway and cancer. Nat Cancer 3:1452–1463. https://doi.org/10.1038/s43018-022-00468-w
Scheffner M, Huibregtse JM, Vierstra RD, Howley PM (1993) The HPV-16 E6 and E6-AP complex functions as a ubiquitin-protein ligase in the ubiquitination of P53. Cell 75(3):495–505. https://doi.org/10.1016/0092-8674(93)90384-3
Schmitz M, Driesch C, Beer-Grondke K, Jansen L, Runnebaum IB, Dürst M (2012) Loss of gene function as a consequence of human papillomavirus DNA integration. Int J Cancer 131(5):E593-602. https://doi.org/10.1002/ijc.27433
Scott ML, Coleman DT, Kelly KC, Carroll JL, Woodby B, Songock WK, Cardelli JA, Bodily JM (2018) Human papillomavirus type 16 E5-mediated upregulation of met in human keratinocytes. Virology 519(czerwiec):1–11. https://doi.org/10.1016/j.virol.2018.03.021
Serrano B, Brotons M, Bosch FX, Bruni L (2018) Epidemiology and burden of HPV-related disease. Best Pract Res Clin Obstet Gynaecol 47(luty):14–26. https://doi.org/10.1016/j.bpobgyn.2017.08.006
Sharma N, Singh SK (2016) Implications of non-coding RNAs in viral infections. Rev Med Virol 26(5):356–68. https://doi.org/10.1002/rmv.1893
Shinohara A, Yokoyama Y, Wan X, Takahashi Y, Mori Y, Takami T, Shimokawa K, Tamaya T (2001) Cytoplasmic/nuclear expression without mutation of Exon 3 of the beta-catenin gene is frequent in the development of the neoplasm of the uterine cervix. Gynecol Oncol 82(3):450–55. https://doi.org/10.1006/gyno.2001.6298
Stenglein MD, Burns MB, Li M, Lengyel J, Harris RS (2010) APOBEC3 proteins mediate the clearance of foreign DNA from human cells. Nat Struct Mol Biol 17(2):222–29. https://doi.org/10.1038/nsmb.1744
Sullivan CS, Grundhoff AT, Tevethia S, Pipas JM, Ganem D (2005) SV40-encoded microRNAs regulate viral gene expression and reduce susceptibility to cytotoxic T cells. Nature 435(7042):682–86. https://doi.org/10.1038/nature03576
Suspène R, Aynaud M-M, Guétard D, Henry M, Eckhoff G, Marchio A, Pineau P, Dejean A, Vartanian J-P, Wain-Hobson S (2011) Somatic hypermutation of human mitochondrial and nuclear DNA by APOBEC3 cytidine deaminases, a pathway for DNA catabolism. Proc Natl Acad Sci USA 108(12):4858–63. https://doi.org/10.1073/pnas.1009687108
Szostek S, Zawilińska B, Biernat-Sudolska M, Kopeć J, Kłeszcz E, Koprynia M, Rojek-Zakrzewska D, Kosz-Vnenchak M (2014) Differences in the expression of human papillomavirus type 16 (HPV-16) E6 oncogene mRNA in SiHa cell line inoculated with CMV, HSV or ureaplasmas. Folia Biol 62(1):73–78. https://doi.org/10.3409/fb62_1.73
Tagawa T, Albanese M, Bouvet M, Moosmann A, Mautner J, Heissmeyer V, Zielinski C et al (2016) Epstein-Barr viral miRNAs inhibit antiviral CD4+ T cell responses targeting IL-12 and peptide processing. J Exp Med 213(10):2065–80. https://doi.org/10.1084/jem.20160248
Takaoka A, Wang Z, Choi MK, Yanai H, Negishi H, Ban T, Lu Y et al (2007) DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 448(7152):501–5. https://doi.org/10.1038/nature06013
Thakur S (2015) Gynecologic and obstetric pathology. Mod Pathol 28(2):271–318. https://doi.org/10.1038/modpathol.2015.17
Uematsu S, Akira S (2007) Toll-like receptors and type I interferons. J Biol Chem 282(21):15319–23. https://doi.org/10.1074/jbc.R700009200
Vieira VC, Leonard B, White EA, Starrett GJ, Temiz NA, Lorenz LD, Lee D et al (2014) Human papillomavirus E6 triggers upregulation of the antiviral and cancer genomic DNA deaminase APOBEC3B. mBio 5(6):e02234-14. https://doi.org/10.1128/mBio.02234-14
Virtanen E, Pietilä T, Nieminen P, Qian K, Auvinen E (2016) Low expression levels of putative HPV encoded microRNAs in cervical samples. SpringerPlus 5(1):1856. https://doi.org/10.1186/s40064-016-3524-3
Wang X, Tang S, Le S-Y, Lu R, Rader JS, Meyers C, Zheng Z-M (2008) Aberrant expression of oncogenic and tumor-suppressive microRNAs in cervical cancer is required for cancer cell growth. PloS One 3(7):e2557. https://doi.org/10.1371/journal.pone.0002557
Wang Z, Wakae K, Kitamura K, Aoyama S, Liu G, Koura M, Monjurul AM, Kukimoto I, Muramatsu M (2014) APOBEC3 deaminases induce hypermutation in human papillomavirus 16 DNA upon beta interferon stimulation. J Virol 88(2):1308–17. https://doi.org/10.1128/JVI.03091-13
Weng S-L, Huang K-Y, Weng JT-Y, Hung F-Y, Chang T-H, Lee T-Y (2018) Genome-wide discovery of viral microRNAs based on phylogenetic analysis and structural evolution of various human papillomavirus subtypes. Brief Bioinform 19(6):1102–14. https://doi.org/10.1093/bib/bbx046
Wierzbicka M, Klussmann JP, San Giorgi MR, Wuerdemann N, Dikkers FG (2021) Oral and laryngeal HPV infection: incidence, prevalence and risk factors, with special regard to concurrent infection in head, neck and genitals. Vaccine 39(17):2344–50. https://doi.org/10.1016/j.vaccine.2021.03.047
Wild CP, Weiderpass E, Stewart BW (2020) World cancer report: cancer research for cancer prevention. International Agency for Research on Cancer: Lyon, France,. 2020
Xu N, Segerman B, Zhou X, Akusjärvi G (2007) Adenovirus virus-associated RNAII-derived small RNAs are efficiently incorporated into the Rna-induced silencing complex and associate with polyribosomes. J Virol 81(19):10540–49. https://doi.org/10.1128/JVI.00885-07
Funding
The study was supported by the National Science Centre, Poland (grant number 2016/23/B/NZ2/03041 to M.G and grant number 2016/21/N/NZ5/01422 to J.J); J.C.B was partially supported by the American Cancer Society.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by: Ewa Ziętkiewicz
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Janiszewska, J., Kostrzewska-Poczekaj, M., Wierzbicka, M. et al. HPV-driven oncogenesis—much more than the E6 and E7 oncoproteins. J Appl Genetics (2024). https://doi.org/10.1007/s13353-024-00883-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13353-024-00883-y