
Abstract
Purpose
This study aims at designing a lactic starter for caper fermentation isolated from Tunisian fermented vegetables to improve the process and produce consistent and high-quality product.
Methods
In this study, the lactic starter was isolated by exploring the lactic acid bacteria (LAB) of Tunisian artisanal fermented vegetables. Identification was carried out by partial 16S rRNA gene sequencing. Screening was based on salt tolerance and antagonistic activities against Escherichia coli ATCC 10536 and Enterococcus faecalis ATCC 10541. Caper fermentation was optimized through a full factorial experimental design (23), by exploring three factors: starter inoculum size, NaCl concentration, and acetate content. Differences in pH values, Total aerobic mesophilic bacteria and LAB counts between the beginning and end of fermentation are selected as responses and corresponding regression coefficients were calculated.
Results
The lactic microbiota is mainly represented by Lactobacillus plantarum group. Based on salt tolerance and antimicrobial activity, the strain Lactobacillus plantarum F3 was selected as starter for caper fermentation. The effect of NaCl concentration, acetate content, and inoculum size on acidity, total aerobic mesophilic bacteria count, and LAB count after 1 week and 1 month of caper fermentation was studied. Depending on the fermentation time, either 1 week or 1 month, the initial conditions should comprise 0% acetate, 108 CFU/mL inoculum, and 5% NaCl for 1 week against 5% acetate, 107 CFU/mL inoculum, and 10% NaCl for 1 month lasting caper fermentation. A protocol for caper fermentation was set up ensuring hygienic quality and LAB viability.
Conclusion
Lb. plantarum F3 was selected as lactic starter for caper fermentation, and initial fermentation conditions were optimized through a full factorial design. This work has shown loss in LAB viability after 1 week of fermentation. Based on results obtained, an optimized fermentation protocol was set up. This protocol ensures LAB survival and high hygienic quality of the product.
Similar content being viewed by others


Introduction
In North African countries, traditional fermented fruits and vegetables are an important part of the diet. This importance is due to: (i) they provide variety in the flavors of existing staples, (ii) a cheap way of preservation, and (iii) improvement of the nutritional quality and digestibility of the raw food materials and made edible, some food that are barely edible (El Sheikha and Montet 2014). Fermentations have been the most important traditional techniques used to preserve many products available only at given seasons of the year such as olives, lemons, onions, green peppers, carrots, figs, grapes, prickly pears, and caper berries (Benkerroum 2013).
Caper berries are the fruits of Capparis species (mainly Capparis spinosa L.), largely cultivated in the Mediterranean basin for its flower buds (capers) and fruits (caper berries) that are consumed, after processing, for their aromatic, digestive, and medicinal properties (Pérez Pulido et al. 2005). In Tunisia, caper fruits are fermented in a traditional way, often in small enterprises or at home. Traditional fermented capers are highly appreciated for their unique organoleptic properties and are often included in culinary preparations. The picking of capers is manual and done between May and August (Douieb et al. 2010). The main procedures of caper fermentation in Tunisia are summarized in Fig. 1. These procedures going from immersion of the berries directly in brine (about 10% w/v NaCl) where they undergo complete or partial fermentation to the addition of vinegar. The spontaneous fermentation of caper depends mainly on the competitive activities of the indigenous microbiota (LAB and yeasts) together with a variety of contaminating microorganisms. However, spontaneous fermentation at the household level leads to variations in the sensory properties of the products and in poor hygienic quality (Benkerroum 2013). To improve fermentation and produce consistent and high-quality final products, a process control is necessary. The use of starter cultures would be an appropriate approach for the control and optimization of the fermentation process resulting in alleviation of the problems of variations in organoleptic and hygienic quality (Callewaert et al. 2000).

Different artisanal procedures used for caper fermentation in Tunisia
Therefore, it would be interesting to develop starter cultures having metabolic activities adapted to use the nutrients available in the food matrix and able to achieve a fast acidification. Furthermore, the antagonistic activity is a selective advantage of the starter against competitors in the fermentation, decreasing the risk of spoilage and contributing to product stability (Ruiz-Barba et al. 1994).
Thus, the aim of this study was to isolate and identify LAB microbiota present in artisanal Tunisian fermented vegetables, screening of a starter for caper fermentation and setting up a fermentation protocol that improved safety.
Materials and methods
Sample collection
Artisanal fermented mixed vegetables (23 samples) were collected from local markets in Tunisia and transported in isothermic bags to the laboratory. Composition of vegetable mixture and brine is indicated in Table 1. Caper berries (Capparis spinosa) were obtained from local market in the region of Tunis (Tunisia). The harvesting was made during the spring 2015.
Isolation and identification of lactic acid bacteria from fermented vegetables
Brine samples were used to isolate LAB. Ten milliliters of brine from each fermented vegetable was homogenized in 90 mL of sterile saline solution; then, 10-fold serial dilutions were prepared. From each dilution, 0.1 mL was plated in triplicate on MRS agar (Scharlab, Barcelona) supplemented with 0.4 g/L sodium azide (Sigma, Madrid). Plates were incubated at 30 °C for 4 days in anaerobiosis (Anaerogen kit, Oxoid, Basingstoke, United Kingdom). Apparent colonies were randomly picked from MRS agar plates. Gram-positive, catalase-negative, non-motile rod, and cocci isolates were considered as LAB and used in further studies.
LAB isolated was identified by sequencing 16S rRNA genes. First, total DNA was extracted using the Qiagen DNeasy Tissue Kit (Qiagen, Courtaboeuf, France). The 16S rRNA genes was amplified using primer pairs FD1: (5′-AGAGTTTGATCCTGGCTCAG-3′), rd1: (5′-TAAGGAGGTGATCCAGCCGCC-3′) (Invitrogen, Cergy Pontoise, France) according to Weisburg et al. (1991). 16S rRNA gene sequence alignments were released using the multiple sequence alignment method and the identification of each strain was carried out by a BLAST search in GenBank (http://www.ncbi.nlm.nih.gov/genbank/). All isolates were stored as 20% glycerol stock cultures at − 80 °C.
Salt resistance test
LAB strains were tested for their ability to grow in presence of 10% (w/v) NaCl. LAB strains were grown on MRS agar supplemented by 10% NaCl and incubated 48 h at 37 °C. Strains growing after incubation were considered as salt resistant and retained for antagonistic activity test.
Antagonistic activity
Antagonistic activity of LAB strains against Escherichia coli ATCC 10536 and Enterococcus faecalis ATCC 10541 was evaluated by the agar well diffusion method of Todorov and Dicks (2005). The target bacteria were grown on Molten agar (7.5 g/L) and incubated at 37 °C for 24 h. Subsequently, LAB strains were inoculated on 10-mL MRS and incubated overnight at 37 °C. Molten agar containing each target bacteria was spread over the surface of agar plates containing TSG medium and left to solidify. After solidification, a hole was made in the center of the plate, and 80 μL of culture (16–18h) of the different strains was placed in the hole and allowed to diffuse at 4 °C for 2 h. After an overnight incubation at 37 °C, the plates were examined for halos around the hole.
Optimization of caper fermentation through a full factorial experimental design
The different fermentation batches were carried out in vats containing each 90 g of caper berries filled with 100 mL sterile water. The fermentation conditions were optimized through a full factorial experimental design (23) repeated twice, by exploring three factors which were chosen as independent quantitative variables, namely, starter inoculum size, NaCl concentration, and acetate content. The levels of the studied factors and the theoretical matrix, showing runs in standard order, are, respectively, illustrated in Table 2.
The starter used for caper fermentation was inoculated into MRS broth and grown at 37 °C for 24 h. Then, culture was harvested by centrifugation at 7500 rpm for 5 min, washed with sterile saline solution (0.9%), and the cell pellets were resuspended in sterile saline solution to produce suspensions of 109 CFU/mL to be inoculated as starter culture in caper fermentation according to the inoculum size mentioned in Table 2.
Fermentation occurred at ambient temperature for 1 month. Sampling from brine was performed from each vat under aseptic conditions at the beginning, after 1 week and 1 month of fermentation. A vat where only NaCl (10%) and acetic acid (5%) were used for the preparation of the brine (without starter inoculation) and incubated in the same conditions was used as control (run 17).
Differences in pH values (∆pH), total aerobic mesophilic bacteria count (∆TAMB), and LAB count (∆LAB) between the beginning and end of fermentation are selected as responses of the experimental design.
In order to evaluate the effect of the three operating factors on the three responses, the regression coefficients CpH for ∆pH, CTAMB for ∆TAMB count, and CLAB for ∆LAB count were calculated as follows:
Where
Aj: means either high (+) or low (−) level in experimental run i;
Xi: ∆TAMB count (CFU/mL)
Yi: ∆pH
Zi: ∆LAB count (CFU/mL)
Statistical analysis and mathematic models
The statistical analysis of the output variables obtained after running the experimental design and the calculation of the coefficients were done with NemrodW® software version 9901 and Minitab® software version 15.
To investigate the relationship between the three operating factors and the three responses, a first-order polynomial model studying the main effects of factors was adopted. The coded mathematical model for 23 factorial designs is expressed as follows:
Where
b0: squared effect term;
bi: regression coefficients corresponding to CTAMB, CpH, and CLAB described previously;
Y: the predicted response value.
Samples were analyzed in triplicate. Data were subjected to one-way ANOVA (SAS, 1985); pair-comparison of treatment means was achieved by Tukey’s procedure at P < 0.05, using the statistical software, Statistica for Windows (Statistica 6.0 per Windows 1998).
Physico-chemical analysis
The pH value of samples was determined at room temperature with an Inolab 740 pH-meter and combined glass electrode (Hanna Instruments, Germany) standardized with pH 4.01 and 7.01 standard buffer solutions (Scharlauchemie, Spain).
Microbial analysis
Serial dilutions of brine samples collected from each flask of fermented caper were used for enumeration of different microbial flora. LAB, TAMB, total coliforms, and yeast and molds were enumerated using the following media, respectively: MRS agar, plate count agar (PCA), Mc Conkey agar, and Sabouraud Choloramphenicol agar. All media are provided from Scharlab (Spain). For all the bacterial enumeration, 0.1/1 mL of suitable dilutions was plated on appropriate media in triplicate. Counts were obtained after 48 h (MRS, PCA, Sabouraud) and 24 h (Mc Conkey) of incubation at 30 °C. Results were calculated as the means of three determinations.
Results and discussion
LAB isolation, enumeration, and identification
As estimated by plating on MRS agar, cell numbers of presumptive LAB isolated from the different fermented vegetables ranged from 6.3 ± 0.02 Log CFU/mL to 8.7 ± 0.03 Log CFU/mL. These findings indicate that the microbiota can vary based on several factors including the origin of the vegetables, the variability of physico-chemical conditions, nutrient composition, and production methods. The cell densities in the fermented vegetables corresponded to those usually reported in the most common varieties (Buckenhüskes 1997).
Seventy-four isolates from fermented vegetables were collected. Out of these isolates, only 56 strains, including 43 rod-shaped bacteria (75.43%) and 13 cocci (24.56%), showing typical appearance of LAB on MRS were Gram-positive, catalase negative, and non-motile. Based on morphological aspect of the colonies, only 39 strains representing the lactic microflora were genotypically identified by partial sequence analysis of the 16S rRNA gene. Results are summarized in Table 1.
Lactic microbiota of fermented vegetables is mainly represented by Lactobacillus plantarum (69%) followed by Lactobocillus plantarum/pentosus group (8%). Four other species of LAB occurred in fermented vegetables with equal proportions (5%): Lactobacillus brevis, Leuconostoc mesenteroides, Pediococcus ethanolidurans, and Enterococcus durans. Lactobacillus alimentarius was also isolated at level of 3%.
The majority of identified species had been frequently found in vegetable matrixes. Lactobacillus plantarum represented the most commonly isolated species in fermented vegetable such as fermented olives, Narezushi, kimchi, and cauliflower (Mourad and Nour-Eddine 2006; Koyanagi et al. 2013; Park et al. 2012; Paramithiotis et al. 2010). The dominance of Lactobacillus plantarum in vegetable fermentations may be explained by its capacity to rapidly adapt to changing environmental conditions, including acid stress (Klaenhammer et al. 2005). In our study, other strains were also found: Lactobacillus alimentarius, Lactobacillus brevis, Leuconostoc mesenteroides, and Pediococcus ethanolidurans. Lactobacillus alimentarius and Lactobacillus brevis were rarely present in vegetable foods but they are found in meat product. Temmerman et al. (2004) had reported the presence of Lactobacillus alimentarius in fermented Sauerkraut and Lactobacillus brevis was also found in fermented fruits and vegetable of Asia (Swain et al. 2014). Leuconostoc spp., including Leuconostoc mesenteroides subsp. mesenteroides, mainly dominated the early spontaneous fermentation of carrots and cauliflower (Lyhs et al. 2004; Paramithiotis et al. 2010). In this research, Pediococcus ethanolidurans was represented by two isolates; this strain was occasionally isolated from fermented vegetable (Iuchi et al. 2012). The presence of Enterococcus durans in some samples of fermented vegetables is seen as poor hygienic quality of the product but it has been isolated from fresh carrot (Hu et al. 2008).
Screening of lactic starter
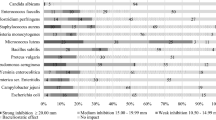
The capacity of a starter to complete with the autochthonous microbiota of the caper and to carry out the metabolic activities expected is conditioned by its growth and survival in the conditions prevailing in the brine, mainly the high salt concentration and low pH. Since caper fermentation occurred in brine with 10% of NaCl, LAB strains were tested for their resistance to this condition. Among the tested strains, only four, belonging to the Lb. plantarum group, tolerate 10% NaCl: Lb. plantarum F2, Lb. plantarum F3, Lb. plantarum F4, and Lb. plantarum G3. This result reflects the capacity of Lb. plantarum species to survive in brine. Salt tolerance for Lb. plantarum may be due to its ability to accumulate osmo- and cryoprotective solutes such as betaine and carnitine (Kets et al. 1994). However, this is not either a general characteristic for Lb. plantarum and variability in salt tolerance has been shown (Montaño et al. 1993).
The retained strains were tested for their antagonistic activities against Escherichia coli ATCC10536 and Enterococcus faecalis ATCC1054. Indeed, antagonistic activity against E. coli is an interesting screening criterion since that presence of coliforms in brine has been reported (Pereira et al. 2008). Only the strain Lb. plantarum F3 showed antagonistic activity against E. coli with an inhibition zone diameter of 11 mm.
Several works studied the antimicrobial compounds produced by LAB such as organic acids, hydrogen peroxide, bacteriocins, fat, and amino acid metabolites (Choi et al. 2018). In our study and by using the well diffusion assay, organic acids may be key molecules for antagonistic activity of the studied strain against foodborne pathogenic bacteria.
Indeed, the growth-inhibiting activity of different LAB strains against pathogens was attributed to a pH reduction and/or to the production of organic acids, including lactic and acetic acid (Tejero-Sariñena et al. 2012). The organic acids secreted by LAB determine an environmental pH reduction that can be adverse for those microorganisms sharing the same niche (Tharmaraj and Shah 2009). In their undissociated form, organic acids can penetrate the cytoplasmic membrane of pathogenic microorganisms resulting in intracellular acidification and in the dissipation of the transmembrane proton motive force (Ricke 2003) or in the case of lactic acid, the antagonistic activity resulted from the permeabilization of the outer membrane of Gram-negative species, causing structural alterations in the phospholipid component (Alakomi et al. 2000).
Moreover, Arena et al. (2016) found a strong antimicrobial activity of Lb. plantarum strains against Listeria monocytogenes, Salmonella enteritidis, Escherichia coli O157:H7, and Staphylococcus aureus and suggested that the ability of Lb. plantarum strains to contrast pathogens growth in vitro was ascribed to a pH-lowering effect of supernatants and/or on the presence of organic acids. Pérez Pulido et al. (2007) reported that among all LAB isolated from fermented caper berries, only the strain Lb. plantarum Lb9 exhibited antagonistic activity against Listeria monocytogenes.
Based on the previous results, the strain Lb. plantarum F3, showing antagonistic activity against E. coli ATCC10536 and tolerance to high amount of NaCl, is more appropriate to be applied as starter for caper fermentation.
Optimization of the fermentation
Effect of fermentation conditions on the hygienic quality of fermented caper and viability of LAB
To insure safety of fermented caper and avoid spoilage during fermentation, three parameters were monitored: acidification, decrease in microbial load, and LAB count during fermentation. Thus, a full factorial experimental design was used. The variables investigated were salt concentration, starter inoculum size, and acetate content as indicated in Table 2. The effect of these variables on pH, TAMB count, and LAB count is represented in Fig. 2.

pH (a), total mesophilic aerobic bacteria count TMAB (b), and lactic acid bacteria count LAB (c) after 1 week (■) and 1 month (□) of caper fermentation; A, B, C, D, E, F, G, and H represent the means of two successive runs, T represent the control vat
After 1 week of fermentation, changes in pH occurred for all the runs (Fig. 2a). Values of pH decrease compared to initial values (P < 0.05). The highest decrease in pH was observed for the run H, and apparently, its conditions might be the most appropriate for rapid acidification (acetate, 107 CFU/mL inoculum, and 10% NaCl). Thus, high inoculum not necessarily leads to rapid decrease in pH; in contrary, acetate addition is needed.
Changes in pH after 1 month of fermentation are remarkable, with a decrease for the runs A, C, E, and G (P < 0.05) (Fig. 2a). The highest decrease was observed for the runs A and E, where the inoculum size is 107 CFU/mL. This fact confirms that, for prolonged fermentation, initial higher inoculum does not improve acidification as well as high salt concentration. The unexpected results were obtained for the runs B, D, F, and H where pH values increase. This fact can be explained by the reduction of lactic acid produced by the starter (Savard et al. 2002). For the control vat, pH remains steady after 1 month of fermentation.
TAMB count was determined after 1 week of fermentation and was significantly different (P < 0.05) (Fig. 2b). The lowest microbial load is obtained for the run B when the fermentation is conducted with addition of acetate, inoculated at 107 CFU/mL and in presence of 5% NaCl. This result confirms the fact observed for pH; high inoculum does not seem suitable to avoid microbial growth; in contrary, acetate addition is needed. Indeed, when lactate and acetate are present in a mixture in fermented vegetables, synergistic antimicrobial properties are often observed (Tsapatsaris and Kotzekidou 2004). For olive fermentation, Ruiz-Barba et al. (1994) suggested that for significant improvement of the fermentation process, the inoculum of starter should be ranged from 105 to 107 CFU/mL.
After 1 month of fermentation, despite that TAMB count is not significantly different (P > 0.05), there is an increase for the runs B, C, F, and H as well as the control vat T (Fig. 2b). This increase may be due to the absence of yeasts after 1 month of fermentation. In addition, the antimicrobial control caused by the starter, in the conditions of the runs B, C, F, and H, was effective during the first week than the autochthonous microflora increases. Similar fact was observed by Vega Leal-Sanchez et al. (2003) when applying Lb. plantarum LPCO10 for olive fermentation. TAMB count decreases only in the experiences D and G and remains stable for experiences A and E. The lowest microbial load after 1 month of fermentation was observed for the experience D with acetate addition, 5% NaCl, and inoculated with 108 CFU/mL.
Since the presence of lactic starter is desirable for caper fermentation to limit the negative effects of spoilage yeasts on the product quality and to insure fast acidification, the effects of different variables on LAB count are studied (Fig. 2c). Results showed that LAB are only present after 1 week and totally absent at the end of fermentation (30 days). A significant decrease was observed after 1 week for all the runs (1.5 to 3.0 Log CFU/mL), and the maximum decrease was retained for the run H (3.0 Log CFU/mL). Similar results were found by Vega Leal-Sanchez et al. (2003). Ruiz-Barba et al. (1994) found a decrease in the cell count of the starter, in olive fermentation, just after inoculation. Our results are similar to those reported by Francesca et al. (2016) during the fermentation of caper with Lb. pentosus OM13 for 45 days. LAB concentration was 7.56 Log CFU/mL after inoculation. From the 3rd day of fermentation, when LAB were found at 7.45 Log CFU/mL, their concentration decreased over time reaching the lowest value (3.06 Log CFU/mL) at 33 days. LAB were no more detected at the 45th day of fermentation.
LAB growth was totally inhibited after 1 month of fermentation; this inhibition could be a consequence of the initial conditions, which may be not favorable for starter growth. Indeed, phenolic compounds present at high concentrations in caper associated with NaCl may be responsible for the LAB decrease. Earlier studies have shown the effect of phenolic compounds on Lb. plantarum; Marsilio and Lanza (1998) described that Lb. plantarum growth was significantly reduced in the presence of p-coumaric acid (1 g/L), a phenolic compound detected in caper berries (Tlili et al. 2015), and the inhibitory activity increased in the presence of NaCl. Ruíz-Barba et al. (1990) studied the effect of phenolic compounds from olives on Lb. plantarum survival and indicated that the phenolic compounds present in the non-alkali treated green olive brines exhibit a bactericidal effect on Lb. plantarum. The bacteriostatic effect of these compounds was avoided by the use of phenolic inactivating agents. Durán et al. (1993) studied the survival of Lb. plantarum during the first days of olive fermentation. They reported that the inhibitory effect of diffused phenolic compounds on Lb. plantarum during these days was significant only when it was associated with NaCl in the brine (3%). The combined effect of 6% NaCl and phenolic compounds caused a marked decrease in survival in 7 days of olive fermentation. Salih et al. (2000) showed the inhibitory effect of hydroxycinnamic acid and p-coumaric acid (both present in caper) on Lb. plantarum growth, besides the inhibitory activity of p-coumaric acid increases in the presence on NaCl.
Total coliforms and yeasts present at the beginning of fermentation at similar level for all the runs, around 1.9 ± 0.09 Log CFU/mL and 1.3 ± 0.05 to 2.0 ± 0.11 Log CFU/mL respectively, are totally absent after 1 week and 1 month of fermentation. Indeed, in fermented vegetables, yeasts are regarded as food spoilage agent and appear after fermentation by autochthonous LAB. The repression of yeast growth in this study can be explained by the low initial load but also the fermentation conditions which associate lactic starter, acetate, and elevated concentrations of NaCl (Anderson et al. 1990; Savard et al. 2002). Massoud and Bassal (2007) found similar results after conservation of caper, coliforms, yeasts, and molds are absent in all the fermented samples. Özcan (2001) obtained the best results for fermented Capparis ovate with 5% brine in the presence of 1% acetic or citric acid.
Statistical analysis of responses and selection of the best initial brining conditions
The responses of the experimental design were analyzed using Nemrod software. Regression coefficients, after 7 days and 30 days of fermentation, are calculated and represented in Table 2. The mathematical models describing the variation Δ of pH, TAMB count, and LAB count are established according to the calculated regression coefficients C as follows:
7 days:
30 days:
After 1 week of fermentation, results indicated that effects of acetate addition, inoculum size, and NaCl concentration on pH decrease were not significant, which can suggest that variable levels need to be optimized. There is no correlation between decrease in pH and decrease in TAMB. The absence of acetate, the low inoculum size, and 5% of NaCl improved decrease of TAMB and the more influencing variable is the inoculum size. This fact can be explained by the antimicrobial activity of Lactobacillus plantarum F3 against autochthonous microflora of caper. High level of inoculum size exhibited a positive effect on LAB growth; meanwhile, the effect of acetate and NaCl concentration on LAB count is not significant.
After 1 month of fermentation, the effects of different factors on pH decrease were similar to those obtained after 1 week. High levels of acetate and NaCl enhance TAMB decrease. However, high level of inoculum size exhibits a negative effect on TAMB decrease.
To investigate the interaction and to visualize the effects of the studied factors on ∆pH and ∆TAMB, a graphical representation of contour plots was generated using Minitab (Figs. 3 and 4). The simultaneous effect of acetate and NaCl on ∆pH shows that acetate addition affects negatively decrease in pH regardless to NaCl concentration and initial inoculum size (Fig. 3a–c). After 1 month, the same interactions were observed; absence of acetate and low inoculum size seem to be more suitable for decrease of more than one pH unit (Fig. 3d). No observable correlation is shown between initial inoculum size and NaCl on ∆pH (Fig. 3e, f).

Contour plots of interaction effect between different variables in pH decrease after 7 days and 30 days of fermentation

Contour plots of interaction effect between different variables in TAMB decrease after 7 days and 30 days of fermentation
Decrease in TAMB is improved by 5% NaCl independently from acetate addition after 1 week of fermentation; however, 5% acetate and 10% NaCl seem to be more suitable for decrease in microbial load after 1 month of fermentation (Fig. 4a, b). Interaction between acetate content and inoculum size shows that presence of acetate and 108 CFU/mL inoculum size is needed to achieve a decrease of more than 2.5 108 CFU/mL of TAMB after 1 week of fermentation (Fig. 4c). To reach the same decrease after 1 month, high inoculum size is needed but in absence of acetate (Fig. 4d). The correlation between inoculum size and NaCl concentration on TAMB decrease shows that high inoculum size and 10% NaCl improve decrease in microbial load after 1 week of fermentation. However, this correlation is not observed after 1 month (Fig. 4e, f).
To conclude and based on the obtained results, no reproducibility of variable effects on studied responses was observed, and depending on fermentation time, two optimal initial conditions are possible: for 1 week lasting caper fermentation: 0% acetate, 108 CFU/mL inoculum, and 5% NaCl, and for 1 month lasting caper fermentation: 5% acetate, 107 CFU/mL inoculum, and 10% NaCl.
Based on above conclusions, a protocol for the manufacture of fermented caper was set up (Fig. 5). It included 30 days: 7 days of fermentation in brine containing 5% acetate and 10% NaCl followed by fermentation for 21 days after starter inoculation at 107 CFU/mL. This two-step fermentation protocol aims at favoring the decrease in pH and microbial load to avoid spoilage due to the synergic action of acetate and salt and maintaining LAB survival. The inoculation with starter is carried out after 7 days since that initial conditions are restrictive for LAB growth. Moreover, the late inoculation leads to a more efficient microbiological control of TAMB by Lb. plantarum F3, since that TAMB count increases after 1 week of fermentation. In spite of the decrease of LAB count, the process indicated in Fig. 5 allows the survival of lactic starter after 30 days of fermentation (LAB count: 3.2 102 CFU/mL). The total microbial load was very low; yeasts, molds, and coliforms were absent at the end of fermentation.

Optimized protocol for caper fermentation with lactic starter
Conclusions
Fermented caper is appreciated by consumers in North Africa and Mediterranean countries for its characteristic organoleptic properties. In this study, a lactic starter (Lb. plantarum F3) for caper fermentation was selected according to technological criteria and initial fermentation conditions were optimized through a full factorial design. This work has shown loss in LAB viability after 1 week of fermentation. Based on results obtained from the full factorial design, an optimized fermentation protocol was set up. This protocol ensures LAB survival after 1 month of fermentation and high hygienic quality of the product.
References
Alakomi HL, Skyttä E, Saarela M, Mattila-Sandholm T, Latva-Kala K, Helander IM (2000) Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl Environ Microbiol 66:2001–2005. https://doi.org/10.1128/AEM.66.5.2001-2005.2000
Anderson RE, Erikson CJ, Salomonsson AC, Theander O (1990) Lactic acid fermentation of fresh and stored carrots: Chemical, microbial and sensory evaluation of products. LWT-Food Sci Technol 3:34–40
Arena MP, Silvain A, Normanno G, Grieco F, Drider D, Spano G, Fiocco D (2016) Use of Lactobacillus plantarum Strains as a bio-control strategy against food-borne pathogenic microorganisms. Front Microbiol 7:article 464. https://doi.org/10.3389/fmicb.2016.00464
Benkerroum N (2013) Traditional Fermented Foods of North African Countries: Technology and Food Safety Challenges With Regard to Microbiological Risks. Compr Rev Food Sci Food Saf 12:54–89. https://doi.org/10.1111/j.1541-4337.2012.00215.x
Buckenhüskes HJ (1997) Fermented vegetables. In: Doyle PD, Beuchat LR, Montville TJ (eds) Food Microbiology: Fundamentals and Frontiers. ASM Press, Washington DC, pp 595–609
Callewaert R, Hugas M, De Vuyst L (2000) Competitiveness and bacteriocin production of enterococci in the production of Spanish-style dry fermented sausages. Int J Food Microbiol 57:33–42. https://doi.org/10.1016/S0168-1605(00)00228-2
Choi AH, Kumar Patra J, Kim WJ, Kang SS (2018) Antagonistic activities and probiotic potential of Lactic Acid Bacteria derived from a plant-based fermented food. Front Microbiol 9:1963. https://doi.org/10.3389/fmicb.2018.01963
Douieb H, Benlemlih M, Errachidi F (2010) Improvement of the lactic acid fermentation of capers (Capparis spinosa L) through an experimental factorial design. Grasas Aceites 61:398–403. https://doi.org/10.3989/gya.010510
Durán MC, García P, Brenes M, Garrido A (1993) Lactobacillus plantarum survival during the first days of ripe olive brining. Syst Appl Microbiol 16:153–158. https://doi.org/10.1016/S0723-2020(11)80260-2
El Sheikha AF, Montet D (2014) African fermented foods: historical roots and real benefits. CRC Press, Taylor and Francis group, London, New York
Francesca N, Barbera M, Martorana A, Saiano F, Gaglio R, Aponte M, Moschetti GC, Settanni L (2016) Optimized method for the analysis of Phenolic compounds from caper (Capparis spinosa L.) berries and monitoring of their changes during fermentation. Food Chem 196:1172–1179. https://doi.org/10.1016/j.foodchem.2015.10.045
Hu CB, Zendo T, Nakayama J, Sonomoto K (2008) Description of durancin TW-49M, a novel enterocin B-homologous bacteriocin in carrot-isolated Enterococcus durans QU 49. JAM 105:681–690. https://doi.org/10.1111/j.1365-2672.2008.03798.x
Iuchi A, Haruguchi S, Mongkol Thanaruk W, Arima J, Nagase M, QuocKhanh H, Ichiyanagi T, Yamaguchi T, Shimomura N, Aimi T (2012) Characterization of Novel Amylase from Amylolytic Lactic Acid Bacteria Pediococcus ethanolidurans Isolated from Japanese Pickles (Nuka-zuke). Food Sci Technol Res 18:86–867. https://doi.org/10.3136/fstr.18.861
Kets EPW, Galinski EA, de Bont JAM (1994) Carnitine: a novel compatible solute in Lactobacillus plantarum. Arch Microbiol 62:243–248. https://doi.org/10.1007/BF00301845
Klaenhammer TR, Barrangou R, Buck BL, Azcarate-Peril MA, Altermann E (2005) Genomic features of lactic acid bacteria effecting bioprocessing and health. FEMS Microbiol Rev 29:393–409. https://doi.org/10.1016/j.fmrre.2005.04.007
Koyanagi T, Nakagawa A, Kiyohara M, Matsui H, Yamamoto K, Barla F, Take H, Katsuyama Y, Tsuji A, Shijimaya M al (2013) Pyrosequencing analysis of microbiota in Kaburazushi, a traditional medieval sushi in Japan. Biosci Biotechnol Biochem 77 : 2125-2130. https://doi.org/10.1271/bbb.130550
Lyhs U, Koort JK, Lundström HS, Björkroth KJ (2004) Leuconostoc gelidum and Leuconostoc gasicomitatum strains dominated the lactic acid bacterium population associated with strong slime formation in an acetic-acid herring preserve. Int J Food Microbiol 90:207–218. https://doi.org/10.1016/S0168-1605(03)00303-9
Marsilio V, Lanza B (1998) Characterization of an oleuropein degrading strain of Lactobacillus plantarum. Combined effects of compounds present in olive fermenting brines (phenols, glucose and NaCl) on bacterial activity. J Sci of Food and Agric 76:520-524
Massoud E, Bassal A (2007) Conservation du câpre. Ann Recherche Sci 7:111–118 hdl.handle.net/2042/41193
Montaño A, Bobillo M, Marshall VM (1993) Effect of sodium chloride on metabolism of two strains of Lactobacillus plantarum isolated from fermenting green olives. LAM 16:315–318. https://doi.org/10.1111/j.1472-765X.1993.tb00366.x
Mourad K, Nour-Eddine K (2006) Microbiological study of naturally fermented green olives: Isolation and identification of lactic acid bacteria and yeasts along with the effects of brine solutions obtained at the end of the olive fermentation. Grasas Aceites 57:292–300. https://doi.org/10.3989/gya.2006.v57.i3.51
Özcan M (2001) Pickling caper flower buds. J Food Qual 24:261–269. https://doi.org/10.1111/j.1745-4557.2001.tb00607.x
Paramithiotis S, Hondrodimou OL, Drosinos EH (2010) Development of the microbial community during spontaneous cauliflower fermentation. Food Res Int 43:1098–1103. https://doi.org/10.1016/j.foodres.2010.01.023
Park EJ, Chun J, Cha CJ, Park WS, Jeon CO, Bae JW (2012) Bacterial community analysis during fermentation of ten representative kinds of kimchi with bar coded pyrosequencing. Food Microbiol 30:197–204. https://doi.org/10.1016/j.fm.2011.10.011
Pereira AP, Pereira JA, Bento A, Estevinho ML (2008) Microbiological characterization of table olives commercialized in Portugal in respect to safety aspects. Food Chem Toxicol 46:2895–2902. https://doi.org/10.1016/j.fct.2008.05.033
Pérez Pulido R, Benomar N, Abriouel H, López RL, Cañamero M, Gálvez A (2005) Microbiological study of lactic acid fermentation of caper berries by molecular and culture-dependent methods. Appl Environ Microbiol 71:7872–7879. https://doi.org/10.1128/AEM.71.12.7872-7879.2005
Pérez Pulido R, Benomar N, Abriouel H, Lucas Lopez R, Martínez Canamero M, Guyot JP et al (2007) Characterization of lactobacilli isolated from caper berry fermentations. JAM 102:583–590. https://doi.org/10.1111/j.1365-2672.2006.03067.x
Ricke SC (2003) Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult Sci 82:632–639
Ruíz-Barba JL, Rios-Sánchez RM, Fedriani-Iriso C, Olias JM, Rios JL, Jiménez-Díaz R (1990) Bactericidal effect of phenolic compounds from green olives on Lactobacillus plantarum. Syst Appl Microbiol 13:199–205. https://doi.org/10.1016/S0723-2020(11)80170-0
Ruiz-Barba JL, Cathcart DP, Warner PJ, Jimenez-Díaz R (1994) Use of Lactobacillus plantarum LPC010, a bacteriocin producer, as a starter culture in Spanish-style green olive fermentations. Appl Environ Microbiol 60:2059–2064 PMC201601
Salih AG, Le Quéré JM, Drilleau JF (2000) Action des acides hydroxycinnamiques libres et esterifiés sur la croissance des bactéries lactiques. Sci Aliment 20:537–560. https://doi.org/10.3166/sda.20.537-560
Savard T, Beaulieu C, Gardner NJ, Champagne CP (2002) Characterization of spoilage yeasts isolated from fermented vegetables and inhibition by lactic, acetic and propionic acids. Food Microbiol 19:363–373. https://doi.org/10.1006/fmic.2002.0483
Swain MR, Anandharaj M, Ray R, Rani R (2014) Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol Res Int 1:1–19. https://doi.org/10.1155/2014/250424
Tejero-Sariñena S, Barlow J, Costabile A, Gibson GR, Rowland I (2012) In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: evidence for the effects of organic acids. Anaerobe 18:530–538. https://doi.org/10.1016/j.anaerobe.2012.08.004
Temmerman R, Huys G, Swings J (2004) Identification of lactic acid bacteria: culture-dependent and culture independent methods. Trends Food Sci Technol 15:348–350. https://doi.org/10.1016/j.tifs.2003.12.007
Tharmaraj N, Shah NP (2009) Antimicrobial effects of probiotics against selected pathogenic and spoilage bacteria in cheese-based dips. Int Food Res J 16:261–276
Tlili N, Mejri H, Feriani A, Saadaoui E, Khaldi A, Nasri N (2015) Phenolic profile and antioxidant activity of Capparis spinosa seedsharvested from different wild habitats. Ind Crop 76:930–953. https://doi.org/10.1016/j.indcrop.2015.07.040
Todorov SD, Dicks LM (2005) Optimization of bacteriocin ST311LD production by Enterococcus faecium ST 311LD, isolated from spoiled black olives. J Microbiol 43:370–374
Tsapatsaris S, Kotzekidou P (2004) Application of central composite design and response surface methodology to the fermentation of olive juice by Lactobacillus plantarum and Debaryomyces hansenii. Int J Food Microbiol 95:157–168. https://doi.org/10.1016/j.ijfoodmicro.2004.02.011
Vega Leal-Sanchez MV, Ruiz-Barba JL, Sanchez AH, Rejano L, Jimenez-Diaz R, Garrido A (2003) Fermentation profile and optimization of green olive fermentation using Lactobacillus plantarum LPCO10 as a starter culture. Food Microbiol 20:421–430. https://doi.org/10.1016/S0740-0020(02)00147-8
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991) 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173:697–703
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Research involving human participants and/or animals
N/A.
Informed consent
N/A.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Manel Ziadi and Taroub Bouzaiene contribute equally to this work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ziadi, M., Bouzaiene, T., Lakhal, S. et al. Screening of lactic starter from Tunisian fermented vegetables and application for the improvement of caper (Capparis spinosa) fermentation through an experimental factorial design. Ann Microbiol 69, 1373–1385 (2019). https://doi.org/10.1007/s13213-019-01519-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13213-019-01519-x