
Abstract
Sweet sorghum bagasse (SSB) from food processing and agricultural industry has attracted the attention for uses in production of biofuel, enzymes and other products. The alteration in lignocellulolytic enzymes by use of supplements in fungal pretreatment of SSB to achieve higher lignin degradation, selectivity value and enzymatic hydrolysis to fermentable sugar was studied. Fungal strain Coriolus versicolor was selected for pretreatment due to high ligninolytic and low cellulolytic enzyme production resulting in high lignin degradation and selectivity value. SSB was pretreated with supplements of veratryl alcohol, syringic acid, catechol, gallic acid, vanillin, guaiacol, CuSO4 and MnSO4. The best results were obtained with CuSO4, gallic acid and syringic acid supplements. CuSO4 increased the activities of laccase (4.9-fold) and polyphenol oxidase (1.9-fold); gallic acid increased laccase (3.5-fold) and manganese peroxidase (2.5-fold); and syringic acid increased laccase (5.6-fold), lignin peroxidase (13-fold) and arylalcohol oxidase (2.8-fold) resulting in enhanced lignin degradations and selectivity values than the control. Reduced cellulolytic enzyme activities resulted in high cellulose recovery. Enzymatic hydrolysis of pretreated SSB yielded higher sugar due to degradation of lignin and reduced the crystallinity of cellulose. The study showed that supplements could be used to improve the pretreatment process. The results were confirmed by scanning electron microscopy, X-ray diffraction, Fourier transform infrared spectroscopy and thermogravimetric/differential thermogravimetric analysis of SSB.
Similar content being viewed by others

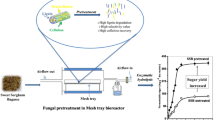

Avoid common mistakes on your manuscript.
Introduction
Increasing agro-industrial activity has led to the production of large quantities of lignocellulosic residues composed of cellulose, hemicellulose and lignin. Sweet sorghum bagasse (SSB) derived from food processing and agricultural industry has recently attracted much attention for its uses in the production of biofuel, enzymes and other useful products. The major problem associated with utilization of SSB is the rigid barrier of lignin that sandwiches the cellulose underneath its recalcitrant structure. There is need for an efficient pretreatment process of biomass to degrade lignin and expose the cellulose for hydrolysis by hydrolytic enzymes. The physical and chemical treatments of lignocellulosic biomass have potential problems, like high energy consumption and environmental pollution (Thakur et al. 2013; Zhang et al. 2007). The uses of biological pretreatments have advantages of being cheaper, safer, less energy consuming and environment-friendly.
Biological pretreatment using white rot fungi (WRF) has great potential for lignin degradation. WRF, due to their ability to produce ligninolytic enzymes laccase, lignin peroxidase (LiP), manganese peroxidase (MnP), polyphenol oxidase (PPO) and arylalcohol oxidase (AAO), can demineralize lignin into CO2 and H2O and decompose the network of lignin for obtaining cellulose (Meehnian et al. 2016). Non-selective lignin degradation, i.e., high cellulose consumption (Hatakka and Hammel 2010), long pretreatment time (Asgher et al. 2016) and cellulose crystallinity (Zhang et al. 2007) are the problems associated with fungal pretreatment of biomass. The high selectivity value (SV) of fungal strains, measured as a ratio of lignin degradation to cellulose consumption, was considered as an important parameter (Chang et al. 2012; Knezevic et al. 2013; Zhang et al. 2007). Pretreatment by Trametes versicolor (Chang et al. 2012), Pleurotus ostreatus (Thakur et al. 2013), Daedalea flavida (Meehnian et al. 2016) and Phanerochaete chrysosporium (Chang et al. 2012), reported to have high SV, varied with the ligninolytic and cellulolytic ability of the strain, type of substrate, fungal–substrate interaction (fungal growth) and culture conditions.
Ligninolytic and cellulolytic enzymes produced by fungi play a vital role in the fungal pretreatment of lignocellulosic biomass. Phenolic supplements (ferulic acid, xylidine, vanillic acid, cinnamic acid and guaiacol) (Liu et al. 2013) and CuSO4 (Meehnian et al. 2016) have been reported to enhance the laccase production, and metallic ion Mn2+ enhanced MnP activity (Song et al. 2013). Metallic salts CuSO4 and MnSO4 have been reported as inhibitors for cellulolytic enzymes (Geiger et al. 1998; Tejirian and Xu 2010). The effects of supplements have been studied in fermentations, mainly with reference to the production of certain ligninolytic enzymes. The effect of supplements on ligninolytic and cellulolytic enzyme activities during pretreatment of lignocellulosic biomass, lignin degradation and SV could be important, but has not been reported till date.
The objective of the present study was fungal pretreatment of SSB with metallic salts and phenolic supplements for a possible alteration in ligninolytic and cellulolytic enzyme profiles to achieve improvement in lignin degradation, SV and enzymatic hydrolysis to fermentable sugar. Fungal strain having high selectivity was screened initially out of different strains reported to be used in the pretreatment (Aguiar et al. 2014; Chang et al. 2012; Knezevic et al. 2013; Taniguchi et al. 2005; Zhang et al. 2007). The detailed studies on the alteration of the enzyme profiles have explained the effect on lignin degradation, SV and sugar yield after enzymatic hydrolysis.
Materials and methods
Materials
2,2′-Azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) was purchased from MP Biomedical (Mumbai, India). Cellulase (SaccariSEB EG) and β-glucosidase (SaccariSEB BG) were gifted from Advanced Enzyme Technologies Ltd. (Thane, India). p-Nitrophenyl-β-d-glucopyranoside, birchwood xylan, lignin sulfonic acid, catechol, vanillin and veratryl alcohol were purchased from Himedia Laboratory Pvt Ltd. (Mumbai, India). Azur-B and bovine serum albumin (BSA) were purchased from S.D. Fine Chem Ltd. (Mumbai, India). Cellulose, Gallic acid and syringic acid were purchased from Sigma Aldrich (Mumbai, India). Water was purified and deionized (DI) by the Milli-Q purification system (minimum resistivity 18 MΩ cm; Millipore, Billerica, MA, USA). Rest of the chemicals used were of analytical grade made by S.D. Fine Chem Ltd. (Mumbai, India).
Biomass
SSB was collected from the local agricultural farms in Jalandhar, Punjab, India. The collected SSB was dried at 40 °C for 72 h and ground to pass through a 5 mm sieve using a Wiley mill. The ground SSB (about 3% moisture) was stored in an airtight container at room temperature.
Fungal strain and maintenance
Coriolus versicolor (MTCC 138), Daedalea flavida (MTCC 145), Ganoderma lucidum (MTCC 1039), Phlebia radiata (MTCC 2791), Pleurotus eryngii (MTCC 1798), Pleurotus ostreatus (MTCC 1801), Stereum hirsutum (MTCC 1099) were procured from Microbial Type Culture Collection (MTCC), Chandigarh, India, and Pycnoporus cinnabarinus (NCIM 1181) from the National Collection of Industrial Microorganisms (NCIM) Pune, India. All strains were grown and maintained in 2% (w v−1) malt extract agar media in Petri plates at pH 4.5, 25 °C. All strains were sub-cultured periodically after every 15 days.
Lignocellulolytic abilities
Ligninolytic ability was detected using basal medium (BM) containing (g L−1) ammonium tartrate 5.0, yeast extract 0.1, potassium phosphate monobasic 1.0, calcium chloride 0.001, magnesium sulfate 0.5 and 1.6% (w v−1) agar (pH 4.5). Substrates, i.e., 0.1% (w v−1) ABTS, 0.1% (v v−1) guaiacol, 0.01% (w v−1) tannic acid, 0.25% (w v−1) lignin sulfonic acid and 0.01% (w v−1) azure B were used to determine the extent of ligninolytic ability (Pointing 1999). A single disc taken from actively growing 7 days old cultured fungal plate using sterile cork borer (10 mm diameter) was used as inoculum. Sterilized Petri plates containing BM and supplemented with substrates were inoculated with test strains and incubated (Innova 42R, Eppendorf Ltd., USA) at 27.5 °C for 5 days. The colorless agar medium turned green due to the oxidation of ABTS to ABTS-azine in the presence of laccase. The colorless medium containing guaiacol in the Petri plate turned brick red in the presence of laccase produced by the fungi. The appearance of a brown oxidation zone around the colonies in the Petri plates containing tannic acid signifies the production of polyphenol oxidase. In the lignin sulfonic acid plate, 1% (w v−1) aqueous solution of ferric chloride and 1% (w v−1) potassium ferricyanide were flooded, washed with DI water after 10 min and observed for clear zones of degraded lignin around colonies against the blue green color of undegraded lignin. In the Petri plate containing azure B, the clearance of blue color of agar showed the ability of lignin peroxidase enzyme production by the fungal strain. The diameters of these color zones around the colony were measured as the ligninolytic ability of the strains.
The cellulolytic ability of fungal strains was detected by the Congo red test (Teather and Wood 1982). Petri plates containing BM with carboxymethyl cellulose (1.1% (w v−1)) as substrate were sterilized. A single disc taken from actively growing 7 days old cultured fungal plate using a sterile cork borer (10 mm diameter) was inoculated and incubated at 27.5 °C for 5 days. The cultivated plates were flooded with aqueous Congo red solution (1 mg mL−1). After 15 min of exposure, the dye was drained and plates were washed three times with 1 M NaCl and observed for production of yellow zones around the colonies. For xylanolytic ability, xylan (4% (w v−1)) was supplemented with BM in Petri plates (Pointing 1999). Sterile plates were inoculated with test strains and incubated at 27.5 °C for 5 days. Plates were flooded with iodine stain (0.25% (w v−1) aqueous I 2 and 0.25% (w v−1) KI). After 5 min of exposure, the dye was drained and plates were washed with DI water and observed for the appearance of a yellow-opaque area against a blue/reddish purple color for undegraded xylan (signifying the release of xylanase). Measurement of the diameter of colored zones around the fungal disc was measured as cellulolytic and hemicellulolytic ability of strains.
Pretreatment of SSB in solid state fermentation (SSF)
Ten discs (using sterile cork borer of 10 mm diameter) of the actively growing 7 days old cultured plates were inoculated into a 500 mL capacity cotton-plugged Erlenmeyer flask containing 50 mL of 2% (w v−1) malt extract medium. After 7 days of incubation at 27.5 °C under static conditions, the culture was aseptically homogenized in a sterilized blender for 15 s × 3 cycles. Two mL (~2.4 × 106 spore mL−1) of blended culture was used as inoculum for 5 g of SSB.
Five grams of ground SSB (~3% moisture) was taken in 250 mL cotton-plugged Erlenmeyer flasks and 11.5 mL of distilled water was added to it (75% moisture). Moistened SSB in flasks were autoclaved at 121 °C for 15 min. The sterilized flasks were inoculated with 2 mL inoculum and incubated in a stationary condition at 27.5 °C for 30 days. Samples (entire content in flask) were taken for compositional analysis of treated biomass, lignocellulolytic enzyme activity assays and enzymatic hydrolysis of biomass.
Effect of supplements in SSF on lignin degradation
To study the effect of supplements on lignin degradation during pretreatment, various inorganic metallic salts and phenolic compounds such as CuSO4, MnSO4, veratryl alcohol, syringic acid, catechol, gallic acid, vanillin and guaiacol were incorporated in the fermentation medium. All the supplements were dissolved in DI water to make a 1 mM supplement solution. Five grams of SSB (~3% moisture) was taken in 250 mL cotton plugged Erlenmeyer flasks and mixed with 11.5 mL of supplement solution to attain a concentration of 2.2 µmol g−1 SSB in culture. The sterilized flasks were inoculated with 2 mL inoculum of the selected fungal strain. Supplements, significantly influencing the lignin degradation, were further studied at various concentrations ranging from 0.11 to 22.0 µmol g−1 SSB. SSB fermented/treated by fungal strain in the absence of the supplement was taken as control for the study. Fermentation and sampling of treated biomass was done as discussed above. All the experiments were done in triplicate.
Lignocellulolytic enzymes assays
50 mL of acetate buffer (pH 4.5, 50 mM) was added to each sample and shaken at 180 rpm at 27.5 °C for 1 h, and then filtered under vacuum to recover most of the water-soluble components and enzymes in the filtrate. The filtrate were stored at 4 °C and used for lignocellulolytic enzyme activity assays. The solid fractions from the pretreated SSB were dried in a hot air oven at 65 °C and used for composition analysis and enzymatic hydrolysis.
Lignin peroxidase (LiP) activity was determined by H2O2-dependent oxidation of azure B at 651 nm (ε 651 = 48 800 M−1 cm−1) (Archibald 1992). The reaction mixture of 2.6 mL contained 1 mL of 0.032 mM azure B, 500 µL of sodium tartrate (pH 3.0, 50 mM) and 500 µL enzyme extract. The reaction was initiated by adding 100 µL of 0.4 mM H2O2. The decrease in absorbance was monitored at 651 nm. One unit of LiP activity was defined as 1 µmol of the azur-B oxidized per minute. Laccase activity was determined by oxidation of ABTS to ABTS-azine at 420 nm (ε 420 = 36 000 M−1 cm−1) (Bourbonnais and Paice 1990). The 1 mL reaction mixture contained 200 µL of acetate buffer (pH 4.5, 0.1 mM), 600 µL of 1 mM ABTS and 200 µL crude enzyme. One unit of laccase activity was defined as 1 µmol of the ABTS oxidized to ABTS radicals per minute. Manganese peroxidase (MnP) activity was determined by phenol red oxidation (ε 610 = 22 000 M−1 cm−1) (Glenn and Gold 1985). Reaction mixtures of 5.5 mL contained 1.75 mL of sodium lactate (pH 4.5, 25 mM), 1.5 mL of sodium succinate buffer (pH 4.5, 20 mM), 500 µL 0.1 mM MnSO4, 750 µL of 1% (w v−1) bovine serum albumin, 750 µL of 0.1% (w v−1) phenol red and 250 µL enzyme extract. The reaction was started by the addition of 100 µL of 0.4 mM H2O2. After 1 min interval, 1 mL of reaction mixture was taken and the reaction was terminated by the immediate addition of 50 µL of 10% (w v−1) NaOH. The absorbance was read at 610 nm. One unit of MnP activity was defined as 1 µmol of the phenol red oxidized per minute. Polyphenol oxidase (PPO) was assayed by oxidation of catechol to quinone at 30 °C (Wong et al. 1971). The reaction mixture of 600 µL contained 100 µL crude enzyme with 500 µL 1.67 mM catechol in sodium phosphate buffer (pH 7.0, 50 mM) (ε 400 = 3450 M−1 cm−1). One unit of enzyme activity was defined as 1 µmol of the catechol transformed per minute. Aryl alcohol oxidase (AAO) activity was monitored using the oxidation of veratryl alcohol to veratrylaldehyde (Guillen et al. 1992). The assay mixture of 2.5 mL contained 1.8 mL of the crude enzyme, 0.1 mL of 50 mM veratryl alcohol and 0.6 mL of sodium tartrate buffer (pH 3.0, 0.5 M) (ε 310 = 9300 M−1 cm−1). One unit of enzyme activity was defined as 1 µmol of the veratryl alcohol oxidized to veratrylaldehyde per minute. All the enzymes were expressed as units per gram dry matter (U gdm−1).
Filter paper cellulase (FPase) activity was determined using Whatman No. 1 filter paper strip, 1.0 × 6.0 cm (equivalent to 50 mg) as substrate (Ghose 1987). 1.0 mL sodium citrate (pH 4.8, 50 mM) and 0.5 mL enzyme extract were added to the substrate and incubated at 50 °C for 30 min. The released sugar was measured at 540 nm using 3,5-dinitrosalicylic acid reagent and using glucose as standard. Carboxymethyl cellulase (CMCase) activity was estimated similar to FPase activity using 1% (w v−1) carboxymethyl cellulose in place of filter paper strips (Ghose 1987). One unit of activity was defined as the amount of enzyme required to produce 1 µmol reducing sugars per min. β-Glucosidase activity was assayed using p-nitrophenyl-β-d-glucopyranoside (p-NPG) as substrate (Wood and Bhat 1988). The reaction mixtures contained 1.8 mL of sodium acetate (pH 4.8, 0.1 M), 1 mL of 5 mM p-NPG in sodium acetate buffer and 0.2 mL of enzyme extract. The reaction mixtures were incubated at 50 °C for 30 min with gentle shaking. Hydrolysis was determined by monitoring the release of p-nitrophenyl at 405 nm with reference to a standard curve prepared using p-nitrophenol. One unit of activity was defined as the amount of enzyme required to produce 1 µmol p-nitrophenol per minute. Xylanase activity was assayed using 1% (w v−1) solution of birchwood xylan in 50 mM citrate buffer (pH 5.0) as the substrate and measuring at 520 nm using xylose as standard (Bailey et al. 1992). One unit of activity was defined as the amount of enzyme required to produce 1 µmol xylose per minute under standard condition of pH and temperature. All the enzymes were expressed as units per gram of dry matter (U gdm−1).
Composition analysis of SSB
Compositions of SSB were determined before and after pretreatment. The contents of lignin, cellulose and hemicelluloses of the SSB samples were determined by the protocol in the NREL report (NREL/TP-510-42618) (Sluiter et al. 2010) using two-stage acid hydrolysis. The first hydrolysis step used 72% (v v−1) sulfuric acid at 30 °C for 1 h. The samples were diluted immediately to 4% (v v−1) and autoclaved for 1 h. The autoclaved hydrolyzed solution was vacuum filtered through previously weighed filtering crucibles. The crucible and the acid-insoluble residue were dried at 105 °C until a constant weight was achieved. The aliquot sample collected from vacuum filtration was used to determine acid-soluble lignin, cellulose and hemicellulose contents. The acid-soluble lignin was measured by UV–Vis spectrophotometer (Kinetic, Biospectrometer, Eppendorf, Germany) using lignin sulfonic acid as standard. Cellulose and hemicellulose in samples were acid-hydrolyzed into monomeric sugars. Monomeric sugars were measured by HPLC (Agilent 1200 Series, MN, USA) equipped with Agilent Hi-Plex-H column and refractive index detector (RID) that were maintained at 80 and 55 °C, respectively. HPLC-grade water was used as the mobile phase, at a flow rate of 0.6 mL min−1. The content of cellulose in the sample was calculated from glucose and galactose obtained, while the content of hemicelluloses was calculated from the sum of xylose and arabinose obtained, and using an anhydro correction of 0.90 for C-6 sugars and 0.88 for C-5 sugars, respectively. The cellulose and hemicellulose contents were calculated using equations % Cellulose = (GR × 0.9)/w × 100 and % Hemicellulose = (XGA × 0.88)/w × 100, respectively, where GR is the glucose and galactose released (g); w the sample dry weight (g) and XGA the xylose and arabinose released (g). The percentage of lignin degradation, which is defined as the percentage of total lignin reduced after pretreatment, was calculated using equation % Lignin degradation = (1 − w(α + β)/(w o (α o + β o))) × 100, where w is the sample dry weight (g) of the pretreated SSB; w o is the initial dry weight (g) of the untreated SSB; α and β are the % of acid-soluble and acid-insoluble lignin in the pretreated SSB, respectively; α o and β o are the % of acid-soluble and acid-insoluble lignin in the untreated SSB, respectively. Dry mass loss was calculated as the percentage of loss of total solids following pretreatment after removing fungal biomass content.
Fungal growth estimation
Fungal growth in SSF was estimated in terms of N-acetyl glucosamine content (Aidoo et al. 1981). The glucosamine content of the fungal cell wall was used to monitor the fungal biomass. At the beginning, a glucosamine standard curve was obtained using N-acetyl glucosamine. Moreover, the glucosamine content of fungi (biomass vs. glucosamine) was determined through liquid culture cultivation. The fungal biomass was treated with 2 N HCl to convert fungal chitin into N-acetyl glucosamine. The glucosamine solution (1.0 mL) was mixed with 1 mL of acetylacetone reagent (1 mL of acetylacetone and 50 mL of 0.5 N sodium carbonate solution) and boiled vigorously for 20 min. After cooling to room temperature, 6 mL of ethanol was added followed by 1 mL of Ehrlich’s reagent (2.67 g of p-dimethylamino benzaldehyde in 100 mL of 1:1 mixture of ethanol and concentrated hydrochloric acid). The mixture was incubated at 65 °C for 10 min and was estimated colorimetrically at 530 nm against a reagent blank.
Characterization of SSB
Morphological, structural and chemical changes in SSB were characterized by scanning electron microscopy (SEM), X-ray diffraction (XRD), Fourier-transformed infrared spectroscopy (FTIR) and thermogravimetry/differential thermogravimetry (TGA/DTG). For SEM imaging, samples were dried in an oven at 45 °C for 48 h followed by gradual dehydration using acetone–water mixtures, with the acetone concentration increasing up to 100%. Samples were sputter-coated with Au–Pd and examined in SEM (EVO 40 EP-CARL ZEISS, Germany) at ×1500 and ×3000 magnification using an accelerating voltage of 20 kV. The sample was analyzed using FTIR (AGILENT CARY-630, USA) spectrometer in the attenuated total reflectance mode to compare the changes in the functional group of the SSB samples after lignin degradation in pretreatment. The samples were prepared by blending and grinding 1% (w w−1) SSB sample in KBr. Scans were taken for each sample from 4000 to 400 cm−1 at a resolution of 2 cm−1. To analyze the degradation of biomass, thermogravimetric (TG) analysis was conducted using TGA/DTG (SII EXSTAR 6000 [TG/DTA 6300], Japan) from 30 to 750 °C. Weight loss experiments were performed on the SSB samples weighing 5 mg. The selected heating rate was 10 °C min−1 in inert atmosphere (nitrogen flow of 20 mL min−1). To determine the crystallinity of the biomass, the XRD patterns of the SSB samples were collected by X-ray diffractometer (PANalytical [Xpert-Prompt, The Netherlands) using a copper X-ray source (k = 0.15–418 nm). The X-ray generator was operated at 45 kV and 40 mA. All samples were ground and sieved with a 60 mesh to make homogenous mixtures. Homogenous powders were dispersed in a thin film (~1 mm thickness) in a flat sample holder. The holder was filled with freeze-dried coffee powder and pressed into a compact material for the analysis. Scans were taken at 1 min from 5° to 50° 2θ. The percentage of crystallinity (%Cr) and crystallinity index (CI) were calculated by Segal et al. (1959) equations as %Cr = I crys/(I crys + I amor) × 100 and CI = (I crys − I amor)/I crys, where I crys is the overall intensity of the peak at 2θ of about 22° and I amor is the intensity at 2θ of about 18°.
Enzymatic hydrolysis of pretreated SSB
Enzymatic hydrolysis of cellulose, untreated and treated sample was carried out following the protocol in NREL/TP-510-42630 (Dowe and McMillan 2001). One gram of biomass (3% moisture) was transferred to a 250 mL Erlenmeyer flask and 100 mL of 50 mM citrate buffer (pH 4.8) was added. Cellulase (SaccariSEB EG, 10 filter paper unit dry g−1 solid) and β-glucosidase (SaccariSEB BG, 20 cellobiose unit dry g−1 solid) enzymes were added for the hydrolysis of the biomass. The flask was incubated in a shaking incubator at 165 rpm at 45 °C at for 108 h and analyzed for release of fermentable sugar. The sugar content was estimated using Agilent Hi-Plex-H column in HPLC (Agilent-1200 infinity series) as described earlier in “Composition analysis of SSB”.
Results and discussion
Screening of selective strains
The best lignin degrading strain was selected out of a total of eight strains reported for pretreatment in literature (Aguiar et al. 2014; Chang et al. 2012; Knezevic et al. 2013; Taniguchi et al. 2005; Zhang et al. 2007) based on the highest ligninolytic and low cellulolytic abilities (Online Resource 1) and maximum SV (lignin degradation/cellulose loss) during pretreatment of SSB in SSF (Online Resource 2).
Strains exhibited the different ligninolytic, cellulolytic and hemicellulolytic abilities, but Coriolus versicolor showed the highest ligninolytic ability and hemicellulolytic ability with low cellulolytic ability (Online Resource 1). Strains showed significant differences in enzyme activities, when grown on SSB in solid state fermentation (Online Resource 3). C. versicolor showed the highest activities of laccase, LiP, MnP and PPO. C. versicolor also produced AAO, but it was less than Ganoderma lucidum and Phlebia radiata. Daedalea flavida, Pleurotus ostreatus and Stereum hirsutum did not show any AAO activity. All the strains were unable to produce all ligninolytic enzymes, but C. versicolor, G. lucidium and P. radiata produced all ligninolytic enzymes (laccase, LiP, MnP, PPO and AAO). P. radiata showed high cellulolytic activity, i.e., CMCase, FPase and β-glucosidase, while C. versicolor showed the lowest CMCase activity and low FPase and β-glucosidase activity. Overall, C. versicolor produced the highest xylanase activity, i.e., capability to utilize hemicelluloses as carbon source, produced low levels of cellulolytic enzymes and high ligninolytic enzymes which significantly degraded lignin in SSB.
Fungal strains showed different SV, due to the varied lignin degradation, cellulose loss and hemicellulose loss depending on their ligninolytic and cellulolytic ability. C. versicolor showed maximum SV (Online Resource 2) and degradation of hemicellulose was high. The ability of the strain to degrade xylan could be an advantage for the dependency of the strains on hemicelluloses and to preserve cellulose during pretreatment. The degradation of the biomass with C. versicolor occurred in two phases. SV increased to 0.65 till 20 days in the first phase as lignin degraded selectively, and in the second phase, SV decreased to 0.59 till 30 days. This was due to the shielded structure of lignin which in the first phase acted as a barrier against the consumption of cellulose by the fungal strain. C. versicolor was selected for studies of the effect of supplements in the pretreatment of SSB.
Selection of supplements
In a preliminary study to observe the effect of the supplements on lignin degradation and SV, SSB was pretreated with 2.2 µmol g−1 SSB (initial) supplements of veratryl alcohol, syringic acid, catechol, gallic acid, vanillin, guaiacol, CuSO4 and MnSO4 (Table 1). CuSO4 resulted in maximum lignin degradation (24.8 ± 2.3% w w−1) followed by gallic acid (22.8 ± 1.0% w w−1), syringic acid (22.5 ± 1.0% w w−1), while it was lower for other supplements (18.5 to 22.0% w w−1). CuSO4 caused minimum consumption of cellulose (21.6 ± 1.01% w w−1) and the presence of syringic acid, catechol and gallic acid also resulted in low cellulose loss in biomass. CuSO4, gallic acid and syringic acid resulted in higher SV 1.15, 0.75 and 0.89, respectively, than the control (without supplements), and we further studied the effect on lignocellulolytic enzyme activities, lignin degradations and SV in pretreatment.
Effect of supplements
Fungal growth and degradation of biomass
The effect of variation in concentrations (0.11–22.0 µmol g−1 SSB) of CuSO4, gallic acid and syringic acid on biomass degradation and fungal growth during pretreatment of SSB was observed.
The composition of SSB was (w w−1): lignin 25.14%, cellulose 38.02% and hemicellulose 25.01%. CuSO4 (2.2 µmol g−1 SSB) resulted in maximum lignin degradation of 24.8 ± 2.3% w w−1 and cellulose loss was 21.6 ± 1.01% w w−1 with SV 1.15 (Fig. 1a). Gallic acid (4.4 µmol g−1 SSB) yielded 22.8 ± 2.1% w w−1 lignin degradation, 20.1 ± 1.8% w w−1 cellulose loss and SV 1.13 (Fig. 1b). Syringic acid (8.8 µmol g−1 SSB) resulted in high lignin degradation (28.7 ± 1.7% w w−1) with minimum cellulose consumption (16.6 ± 1.0% w w−1) and highest SV 1.77 (Fig. 1c). The higher concentration of supplements decreased the lignin degradation and SV, which might be due to the inhibition of fungal growth and ligninolytic enzyme activities.

Effect of a CuSO4, b gallic acid and c syringic acid at different concentrations on degradation (% w w−1) of SSB and selectivity value after 20 days of pretreatment with C. versicolor. Black filled bar, lignin; gray filled bar, cellulose; open bar, hemicelluloses; black filled square, selectivity value
SSB supplemented with syringic acid showed higher lignin degradation than CuSO4, gallic acid and control (Fig. 2a). Slow lignin degradation during the first 4–8 days of pretreatment was due to consumption of easily accessible small amounts of cellulose and hemicellulose for primary growth of fungi. Supplements resulted in low cellulose consumption in SSB (Fig. 2b), which might be due to reduced cellulolytic activities. The cellulose content in SSB pretreated with syringic acid was high, which could provide higher accessibility of carbohydrates for enzymatic saccharification and sugar yield (Moscon et al. 2014). In the case of CuSO4, SV was highest till 12 days and thereafter declined (Fig. 2c), likely due to consumption of exposed cellulose after lignin degradation at the latter phase. Similarly with gallic acid, SV reduced after 8 days of pretreatment.

Effect of supplements on degradation of a lignin, b cellulose, c variation of selectivity value, d degradation of hemicellulose in SSB, e variation of hemicelluloses loss to cellulose loss ratio, f growth of C. versicolor during pretreatment of SSB.  without supplement;
without supplement;  CuSO4 (2.2 µmol g−1 SSB);
CuSO4 (2.2 µmol g−1 SSB);  gallic acid (4.4 µmol g−1 SSB);
gallic acid (4.4 µmol g−1 SSB);  syringic acid (8.8 µmol g−1 SSB)
syringic acid (8.8 µmol g−1 SSB)
Syringic acid (8.8 µmol g−1 SSB) stimulated C. versicolor to feed on hemicelluloses than on cellulose, and the consumption of hemicellulose was maximum (28.2 ± 1.7% w w−1) (Fig. 2d). The ratio of hemicelluloses loss to loss of cellulose was higher in the case of syringic acid. The pattern of hemicellulose loss to cellulose loss (Fig. 2e) was similar to the pattern of SV. This was due to the effect of decreased consumption of cellulose with increase in the consumption of hemicellulose. The ratio of loss of hemicellulose to loss of cellulose (2.0 ± 0.09) was high in the presence of CuSO4 (2.2 µmol g−1 SSB) till 12 days and then decreased to 1.0 ± 0.04 (20th day), lower than at 8.8 µmol g−1 SSB syringic acid (1.69 ± 0.07). This was due to variation in lignocellulolytic enzyme profiles of fungal culture with time.
Fungus grew well with supplements in SSF (Fig. 2f) and prolific colonization was observed in 20 days. The fungal growth was slow during the first 4 days of pretreatment due to the lag phase. The fungal biomass were higher with phenolic supplements of gallic acid (413.8 ± 38.0 mg gdm−1) and syringic acid (442.2 ± 26.3 mg gdm−1) than in the control (384.5 ± 31.2 mg gdm−1) or with CuSO4 (328.3 ± 18.4 mg gdm−1). The presence of supplement reduced the exponential phase and stationary phase achieved earlier, after 16 days. The lower fungal biomass in case of CuSO4 than control might be due to oxidative stress.
Ligninolytic and cellulolytic enzyme production
Peak activities of ligninolytic enzymes laccase 11.9 ± 0.3, LiP 0.13 ± 0.01, MnP 8.1 ± 0.2, PPO 4.9 ± 0.7 and AAO 1.39 ± 0.04 U g−1 were detected in control (Fig. 3a) and activities increased with supplements (Fig. 3b–d). 2.2 µmol g−1 SSB CuSO4 increased the peak activities of laccase (4.9-fold) and PPO (1.9-fold), while LiP and AAO were little affected (Fig. 3b). This might be due to the induction effect of copper on laccase and related oxidative enzyme (PPO) at the transcription level (Kaur and Sudhakara 2011). Unexpectedly, significant increase in MnP activity was also observed. All ligninolytic enzymes showed peak activities on the 12th day except LiP on the 16th day (Fig. 3b). The stability of laccase activity was observed, likely due to inhibition of its degradation by extracellular proteolytic enzymes in the presence of Cu2+ (Kaur and Sudhakara 2011). Gallic acid (4.4 µmol g−1 SSB) resulted in high activities of laccase 42.1 ± 3.6, MnP 20.3 ± 1.1 and AAO 2.5 ± 0.1 U g−1 with optimum LiP. The positive effect of gallic acid on ligninolytic enzymes was due to its structural similarity with lignin components. Laccase, MnP and AAO peaked on the 16th, 12th and 20th day, respectively (Fig. 3c). Syringic acid (8.8 µmol g−1 SSB) significantly increased LiP 1.7 ± 0.08 U g−1 which was 13-fold higher than the activity in the control. This might be due to the threefold increase in AAO by the syringic acid-produced H2O2 in the culture, which significantly increased the LiP activity. Additionally, laccase activity 67.4 ± 7.7 U g−1 increased 5.6-fold and MnP activity 19.0 ± 1.1 U g−1 increased 2.5-fold with respect to the control. All ligninolytic enzymes showed their peak activities on the 16–20th day (Fig. 3d). The significant increase in enzyme activity might be due to the structural similarity of syringic acid with the syringyl unit of the lignin. The difference in syringic acid and syringyl unit of lignin is the functional group at the para site of the benzyl ring. The structural similarity could result in high inducing effect for release of ligninolytic enzyme (D’souza et al. 1999). The increase in ligninolytic enzyme activities supported the increased lignin degradation.

Lignocellulolytic enzyme activities during pretreatment of SSB by C. versicolor: a without supplement (control) and with supplements, b CuSO4 (2.2 µmol g−1 SSB), c gallic acid (4.4 µmol g−1 SSB), d syringic acid (8.8 µmol g−1 SSB). Ligninolytic enzymes (I):  laccase,
laccase,  MnP,
MnP,  PPO,
PPO,  LiP,
LiP,  AAO; cellulolytic and hemicellulolytic enzyme (II):
AAO; cellulolytic and hemicellulolytic enzyme (II):  β-glucosidase,
β-glucosidase,  CMCase,
CMCase,  FPase,
FPase,  xylanase activity
xylanase activity
The lignin in SSB is composed of phenylpropanoid p-hydroxyphenyl, guaiacyl and syringyl structural units to form polymer by ether and C–C linkages, predominantly, β-O-4′ aryl ether linkages, together with minor amounts of β-β′, β-5′, β-1′ and α,β-diaryl ether linkages (Sun et al. 2013). Ligninolytic enzymes can break linkages in the lignin structure by cleavage of Cα–Cβ bonds and Cα oxidation. All ligninolytic enzymes possess the ability to break these linkages, but LiP has the highest (E ′o ~ 1.2 V) and laccase has the lowest (E ′o = 0.5–0.8 V) redox potential. AAO (AAO produces H2O2; initiator for LiP) and LiP cleaved the Cα–Cβ bonds in inter-unit β-O-4′ and β–β′ linkages, initiating the depolymerization of the lignin structure. Partially degraded lignin and different types of aromatic monomers of lower molecular weights were further degraded by laccase, PPO and MnP (Bourbonnais and Paice 1990; Hatakka and Hammel 2010).
Supplements decreased the cellulolytic enzyme activities significantly (Fig. 3a–d). CMCase activity (3.0 ± 0.21 U g−1) reduced to half of control, while the FPase (10.0 ± 1.3 U g−1) and β-glucosidase activity (6.1 ± 0.8 U g−1) decreased with CuSO4. Activities of cellulolytic enzymes CMCase 2.2 ± 0.1, FPase 6.7 ± 0.9 and β-glucosidase 4.8 ± 0.2 U g−1 decreased to minimum with syringic acid. Reduced cellulolytic activities resulted in low consumption of cellulose and improved SV. The maximum xylanase activity 8.9 ± 0.5 U g−1 in the presence of syringic acid caused high consumption of hemicelluloses.
Characterization of SSB
SSB was brownish due to the presence of lignin and then changed to light yellow after pretreatment, indicating the exposed cellulose on the surface. The structure of a single strand of SSB (SEM images) was covered with small globules and the surface structure of lignocelluloses was intact (Fig. 4a). The surface of pretreated SSB became rugged due to removal of lignin and breaking of lignocellulose networks. A visible destruction in morphology and formation of holes and crevices on the biomass surfaces were observed in pretreated SSB (Fig. 4b). A maximum rugged surface and pores were visualized in SSB pretreated with syringic acid (8.8 µmol g−1 SSB). These intrinsic structural changes increase the biomass reactivity by enhancing cellulose accessibility toward enzymatic hydrolysis.

Characterization of untreated and pretreated SSBs. a SEM (untreated), b SEM (pretreated with supplement syringic acid), c FTIR, d XRD, e TGA.  Avicel;
Avicel;  untreated SSB; pretreated
untreated SSB; pretreated  without supplement; with
without supplement; with  CuSO4 (2.2 µmol g−1 SSB),
CuSO4 (2.2 µmol g−1 SSB),  gallic acid (4.4 µmol g−1 SSB),
gallic acid (4.4 µmol g−1 SSB),  syringic acid (8.8 µmol g−1 SSB)
syringic acid (8.8 µmol g−1 SSB)
FTIR spectroscopy due to functional groups showed broad peaks at 3418 cm−1 (hydrogen bonded O–H stretching), 2930 cm−1 (C–H stretching), 1638 cm−1 (C=C stretching of aromatic ring), 1435 cm−1 (C–H stretching) and 1130 cm−1 (C–O stretching) (Zeng et al. 2011) for lignin (Online Resource 4) and untreated SSB (Fig. 4c). Pretreatment of SSB resulted in decrease in the respective peaks and intensity at 1638 cm−1 (C=C stretching for aromatic ring mainly originating from lignin) (Fig. 4c). The largest decrease in the intensities of lignin was observed in SSB pretreated with syringic acid (8.8 µmol g−1 SSB).
X-ray diffraction analysis displayed change in percentage crystallinity (%Cr) and the crystallinity index (CI) of pretreated SSB, taking pure micro-crystalline cellulose (Avicel) as reference (Fig. 4d; Online Resource 5). The %Cr and CI of Avicel was 88.1% and 0.86. The %Cr and CI of untreated SSB, pretreated SSB without supplement, SSB pretreated with CuSO4 (2.2 µmol g−1 SSB), gallic acid (4.4 µmol g−1 SSB) and syringic acid (8.8 µmol g−1 SSB) were 75.2, 73.9, 73.1, 72, 66.4% and 0.72, 0.65, 0.63, 0.60 and 0.50, respectively. The decrease in %Cr and CI indicated that the pretreatment was effective in lowering the crystallinity of SSB due to degradation of lignin (Yoshida et al. 2008).
The thermal degradation characteristics using TGA/DTG curves showed that degradation of SSB occurred in three steps (Fig. 4e; Online Resource 6): (i) dehydration and volatilization at <180 °C, (ii) degradation between 190 and 500 °C corresponding to depolymerization of hemicelluloses and cellulose components and (iii) charcoal formation at >500 °C by slow decomposition of the solid residues of lignin. DTG curves shifted toward the lower temperature range, percentage mass loss was higher and the intensities of DTG peaks increased after pretreatment due to lower thermal stability and reduction in cellulose crystallinity.
Enzymatic hydrolysis
Pretreatment with supplements affected the enzymatic hydrolysis of SSB (Fig. 5). The glucose yield on enzymatic hydrolysis of pure cellulose, untreated SSB, pretreated SSB without supplement and SSB pretreated with CuSO4 (2.2 µmol g−1 SSB), gallic acid (4.4 µmol g−1 SSB) and syringic acid (8.8 µmol g−1 SSB) were 764.0 ± 37, 162.7 ± 6.6, 189.3 ± 9.2, 214.3 ± 8.8, 256.4 ± 10.5 and 312.3 ± 12.8 mg g−1, respectively. Pretreatment with supplements increased the lignin degradation and porosity of SSB that enabled the penetration of hydrolytic enzymes and increased the glucose yield. The sugar yield from SSB pretreated with syringic acid supplement was ~1.91 and 1.65 times higher than untreated SSB and control (pretreatment without supplement).

Effect of pretreatment on sugar yield after enzymatic hydrolysis of SSB.  untreated;
untreated;  control; pretreated with supplements
control; pretreated with supplements  syringic acid (8.8 µmol g−1 SSB),
syringic acid (8.8 µmol g−1 SSB),  CuSO4 (2.2 µmol g−1 SSB),
CuSO4 (2.2 µmol g−1 SSB),  gallic acid (4.4 µmol g−1 SSB);
gallic acid (4.4 µmol g−1 SSB);  cellulose
cellulose
Conclusion
Coriolus versicolor gave high degradation of lignin and SV in the pretreatment of SSB. CuSO4, gallic acid and syringic acid supplements increased ligninolytic and decreased cellulolytic enzyme activities resulting in improvement of lignin degradation and SV. CuSO4 increased the activities of laccase (4.9-fold) and PPO (1.9-fold); gallic acid increased laccase (3.5-fold) and MnP (2.5-fold); and syringic acid increased laccase (5.6-fold), LiP (13-fold) and AAO (2.8-fold) than control. Syringic acid resulted in maximum lignin degradation with highest SV and enzymatic hydrolysis SSB after pretreatment yielded higher (~1.9 times) sugar. Supplements could be used to improve the fungal pretreatment process.
References
Aguiar A, Gavioli D, Ferraz A (2014) Metabolite secretion, Fe3+-reducing activity and wood degradation by the white-rot fungus Trametes versicolor ATCC 20869. Fungal Biol 118:935–942
Aidoo KE, Hendry R, Wood BJB (1981) Estimation of fungal growth in a solid-state fermentation system. Eur J Appl Microbiol Biotechnol 12:6–9
Archibald FS (1992) A new assay for lignin-type peroxidases employing the dye azure B. Appl Environ Microbiol 58:3110–3116
Asgher M, Wahab A, Bilal M, Iqbal HMN (2016) Lignocellulose degradation and production of lignin modifying enzymes by Schizophyllum commune IBL-06 in solid-state fermentation. Biocatal Agric Biotechnol 6:195–201
Bailey MJ, Biely P, Poutanen K (1992) Interlaboratory testing of methods for assay of xylanase activity. J Biotechnol 23:257–270
Bourbonnais R, Paice MG (1990) Oxidation of non-phenolic substrates: an expanded role for laccase in lignin biodegradation. FEBS Lett 267:99–102
Chang AJ, Fan J, Wen X (2012) Screening of fungi capable of highly selective degradation of lignin in rice straw. Int Biodeterior Biodegrad 72:26–30
D’souza TM, Merritt CS, Reddy CA (1999) Lignin-modifying enzymes of the white rot basidiomycete Ganoderma lucidum. Appl Environ Microbiol 65:5307–5313
Dowe N, McMillan J (2001) SSF experimental protocols: lignocellulosic biomass hydrolysis and fermentation: laboratory analytical procedure (LAP). National Renewable Energy Laboratory, Golden
Geiger G, Brandl H, Furrer G, Schulin R (1998) The effect of copper on the activity of cellulase and β-glucosidase in the presence of montmorillonite or Al-montmorillonite. Soil Biol Biochem 30:1537–1544. doi:10.1007/978-1-4615-4683-2_31
Ghose T (1987) Measurement of cellulase activities. Pure Appl Chem 59:257–268
Glenn JK, Gold MH (1985) Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch Biochem Biophys 242:329–341
Guillen F, Martinez AT, Martinez MJ (1992) Substrate specificity and properties of the aryl-alcohol oxidase from the ligninolytic fungus Pleurotus eryngii. Eur J Biochem 209:603–611
Hatakka A, Hammel K (2010) Fungal biodegradation of lignocelluloses. In: Hofrichter M (ed) The mycota: industrial applications, vol 10. Springer, Berlin, pp 319–340. doi:10.1007/978-3-642-11458-8_15
Kaur H, Sudhakara KM (2011) Effect of carbon, nitrogen sources and inducers on ligninolytic enzyme production by Morchella crassipes. World J Microbiol Biotechnol 27:687–691. doi:10.1007/s11274-010-0507-3
Knezevic A, Milovanovic I, Stajic M, Vukojevic J (2013) Potential of Trametes species to degrade lignin. Int Biodeterior Biodegrad 85:52–56. doi:10.1016/j.ibiod.2013.06.017
Liu Y, Sun J, Luo Z, Rao S, Su Y, Yang Y (2013) Effect of supplements Mn2+, Cu2+, and aromatic compounds and Penicillium decumbens on lignocellulosic enzyme activity and productivity of Catathelasma ventricosum. J Microbiol Biotechnol 23:565–571. doi:10.4014/jmb.1211.11007
Meehnian H, Jana AK, Jana MM (2016) Effect of particle size, moisture content, and supplements on selective pretreatment of cotton stalks by Daedalea flavida and enzymatic saccharification. 3 Biotech 6:235
Moscon JM et al (2014) Comparison of conventional and alternative technologies for the enzymatic hydrolysis of rice hulls to obtain fermentable sugars. Biocatal Agric Biotechnol 3:149–154
Pointing SB (1999) Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Divers 2:17–33
Segal L, Creely J, Martin A, Conrad C (1959) An empirical method for estimating the degree of crystallinity of native cellulose using the X-ray diffractometer. Text Res J 29:786–794
Sluiter A, Hames B, Ruiz R, Scarlata C, Sluiter J, Templeton D, Crocker D (2010) Determination of structural carbohydrates and lignin in biomass: laboratory analytical procedure (LAP). National Renewable Energy Laboratory, National Renewable Energy Laboratory, Golden
Song L, Ma F, Zeng Y, Zhang X, Yu H (2013) The promoting effects of manganese on biological pretreatment with Irpex lacteus and enzymatic hydrolysis of corn stover. Bioresour Technol 135:89–92. doi:10.1016/j.biortech.2012.09.004
Sun SL, Wen JL, Ma MG, Li MF, Sun RC (2013) Revealing the structural inhomogeneity of lignins from sweet sorghum stem by successive alkali extractions. J Agric Food Chem 61:4226–4235. doi:10.1021/jf400824p
Taniguchi M, Suzuki H, Watanabe D, Sakai K, Hoshino K, Tanaka T (2005) Evaluation of pretreatment with Pleurotus ostreatus for enzymatic hydrolysis of rice straw. J Biosci Bioeng 100:637–643. doi:10.1263/jbb.100.637
Teather RM, Wood PJ (1982) Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol 43:777–780
Tejirian A, Xu F (2010) Inhibition of cellulase-catalyzed lignocellulosic hydrolysis by iron and oxidative metal ions and complexes. Appl Environ Microbiol 76:7673–7682. doi:10.1128/aem.01376-10
Thakur S, Shrivastava B, Ingale S, Kuhad RC, Gupte A (2013) Degradation and selective ligninolysis of wheat straw and banana stem for an efficient bioethanol production using fungal and chemical pretreatment. 3 Biotech 3:365–372
Wong TC, Luh BS, Whitaker JR (1971) Isolation and characterization of polyphenol oxidase isozymes of clingstone peach. Plant Physiol 48:19–23
Wood TM, Bhat KM (1988) Methods for measuring cellulase activities. Methods Enzymol 160:87–112
Yoshida M et al (2008) Effects of cellulose crystallinity, hemicellulose, and lignin on the enzymatic hydrolysis of Miscanthus sinensis to monosaccharides. Biosci Biotechnol Biochem 72:805–810. doi:10.1271/bbb.70689
Zeng J, Singh D, Chen S (2011) Biological pretreatment of wheat straw by Phanerochaete chrysosporium supplemented with inorganic salts. Bioresour Technol 102:3206–3214. doi:10.1016/j.biortech.2010.11.008
Zhang X, Xu C, Wang H (2007) Pretreatment of bamboo residues with Coriolus versicolor for enzymatic hydrolysis. J Biosci Bioeng 104:149–151. doi:10.1263/jbb.104.149
Acknowledgements
Mrs. Vartika Mishra gratefully acknowledges the Ministry of Human Resource Development (MHRD), Government of India, for providing the fellowship during the study. All authors are highly thankful to the National Institute of Technology (NIT), Jalandhar, for providing grants and administrative support for the study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
Ligninolytic and cellulolytic ability of white rot fungi measured by diameter of characteristic zones (PDF 249 kb)
Online Resource 2
Degradation of lignin (LIG), cellulose (CEL), hemicellulose (HEM) and variation of selectivity value (SV) during pretreatment of SSB (PDF 156 kb)
Online Resource 3
Peak ligninolytic, cellulolytic and hemicellulolytic enzyme activities during pretreatment by fungal strains (PDF 291 kb)
Online Resource 4
FTIR spectra of lignin (PDF 128 kb)
Online Resource 5
Percentage crystallinity (%Cr) and crystallinity index (C.I.) of Avicel, untreated and pretreated SSB (PDF 354 kb)
Online Resource 6
Thermal degradation of untreated and pretreated SSB (PDF 281 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mishra, V., Jana, A.K., Jana, M.M. et al. Fungal pretreatment of sweet sorghum bagasse with supplements: improvement in lignin degradation, selectivity and enzymatic saccharification. 3 Biotech 7, 110 (2017). https://doi.org/10.1007/s13205-017-0719-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-017-0719-4