
Abstract
This study presents results from the first 5 years of an organic cropping trial in Ontario, Canada, where legume cover crops were the primary nitrogen source in a soybean-winter wheat/cover crop-corn rotation. Treatments included cover crop termination using moldboard plow (MP) or chisel plow (CP), a no-cover crop control under conventional production (CK-C), and four cover crops including summer-seeded crimson clover (CC, Trifolium incarnatum L.), summer-seeded hairy vetch (HV, Vicia villosa L. Roth), summer-seeded red clover (RCss, Trifolium pratense L.), and frost-seeded red clover (RCfs). Summer-seeding occurred after wheat harvest (July–August), and frost-seeding occurred in early spring (March–April). At cover crop termination, average aboveground cover crop biomass ranged from 5.9 to 8.1 Mg ha−1, while accumulated biomass nitrogen ranged from 155 to 193 kg ha−1. Corn grain yields were 11.6 Mg ha−1 for MP and 10.2 Mg ha−1 for CP tillage-termination method; and 13.3 Mg ha−1 for CK-C, 10.9 Mg ha−1 for RCfs, 10.6 Mg ha−1 for HV, 10.2 Mg ha−1 for CC, and 9.5 Mg ha−1 for RCss. Organic winter wheat yields were nitrogen-limited, averaging 27% lower than CK-C. Winter wheat yields were 10–15% lower in the RCfs than in other summer-seeded cover crop treatments. Soybean yields were largely unaffected by the treatments. It was concluded that summer-seeded legume cover crops are an effective primary nitrogen source for corn, but not as effective for the winter wheat phase of the soybean-winter wheat-corn rotation.
Similar content being viewed by others


Introduction
At present, organic agriculture production is experiencing sustained growth (e.g., USDA 2022; COTA 2021). It is believed that this is because of two main reasons: (i) growing consumer demand for food that is free of agri-chemical residues and produced using environmentally friendly practices (Stockdale and Watson 2009; Gomiero et al. 2011; Lynch 2017) and (ii) increasing realization by growers that organic production generates higher crop prices while reducing fertilizer and pesticide input costs (Archer et al. 2007; Smith et al. 2004; Moyer et al. 2017; Seidel et al. 2017). However, organic production can be challenging as yields are often reduced by sub-optimal levels of crop-available nitrogen (N) (Pang and Letey 2000; Spargo et al. 2011; Ponisio et al. 2015).
Organic farmers have several options for adding N to their soils, such as commercial organic N fertilizers, compost, manure, and leguminous cover crops which extract or “fix” N from the atmosphere (Canadian General Standards Board 2011; USDA 2018). Commercial organic N fertilizers are often expensive; however, while manure and compost are frequently limited in quantity and availability, and their use as a sole N source often results in overapplication of phosphorus (Sadeghpour et al. 2017). Although leguminous cover crops are well-established sources of N, their inclusion as a phase in a traditional cash crop rotation requires one or more years without a cash crop (Bruns 2012; McBride and Greene 2015). For example, adding a leguminous alfalfa cover crop in a corn-soybean-cereal rotation (e.g., corn-soybean-cereal/alfalfa-alfalfa) may supply sufficient N for the corn and cereal cash crops, but it results in no cash crop every 4 years during the alfalfa phase (Delate et al. 2013; McBride and Greene 2015; Hunt et al. 2019).
In southern Ontario, the most common legume cover crop is red clover (RC), which is typically “frost-sown” by broadcasting RC seed into the winter wheat phase of a corn-soybean-winter wheat rotation during early spring (OMAFRA 2018). This cover cropping practice relies on an adequate period of diurnal soil freeze–thaw conditions to both incorporate and germinate the RC seed, which is notoriously difficult because of weather-dependency and the requirement for precise timing (Blaser et al. 2006). Frost-seeding RC cover crops in southern Ontario is consequently very much a “hit-and-miss” practice, and it often does not achieve sufficient population density to provide adequate N to the following crops in the rotation (e.g., Hilker 2021).
In southern Ontario, drill-planting legume cover crops soon after winter wheat harvest could be a viable alternative to frost-seeding. Post-harvest drill-planting virtually ensures good germination of cover crop seed, and the ample crop heat units and precipitation accrued in southern Ontario between summer-seeding (mid-July to early August) and spring termination (May) usually leads to substantial accumulation of cover crop biomass and biomass N (Yang et al. 2019), effective weed suppression by the cover crop (Brust et al. 2014), and mobilization of phosphorus for the following crops (such as corn, Kamh et al. 2002). Although contribution of legume N to conventional corn has been studied in southern Ontario (Coombs et al. 2017; Yang et al. 2023), there is little information for this region on the use of summer-seeded leguminous cover crops as primary N source in the classic corn-soybean-winter wheat rotation.
Success in organic farming depends on how well crop rotation is designed and managed (Kalinova 2010). Yield loss during the usual 3-year transition from conventional to organic production can be substantial (Liebhardt et al. 1989; MacRae et al. 1990), and they linked this loss to be due primarily to reduced availability of N nutrient in the soil. Hence, corn and other high-value crops with large N demand are often not successful during the transition period (Liebhardt et al. 1989). The transition period for the field site in this study occurred during 2014–2017 and involved conversion of a conventional corn-soybean-winter wheat rotation to an organic rotation using leguminous cover crops as the N source (Yang et al. 2019). During the transition period, Yang et al. (2019) found that (i) average corn yields after hairy vetch and red clover cover crops were similar to the conventional control; (ii) 3-year average winter wheat yields were significantly greater for the conventional control (5.1 Mg ha–1) than for the cover crops (3.9 Mg ha–1) and organic control (3.7 Mg ha–1); and (iii) soybean yields were similar among both the cover crop treatments and the conventional control.
The objectives of this study were to determine for the first 5 years (2018–2022) of the organic (i.e. post-transition) period: (i) cover crop performance in terms of biomass and biomass N production and (ii) corn, soybean, and winter wheat grain yields when nutrient N was supplied entirely by the biomass of spring and summer-seeded leguminous cover crops under two tillage treatments.
Materials and methods
Field site and experimental design
The field site was located at the Harrow Research and Development Centre, Harrow, Ontario (42.0°N, 82.9°W) on land that passed the 3-year (2014–2017) transition from conventional crop production to certified organic crop production (Yang et al. 2019). Mean annual air temperature and precipitation during the study years (2018–2022) were 10.9 °C and 801 mm, respectively (further details in Table S1). The soil type was Harrow sandy loam (Brunisolic Grey-Brown Podsol, Soil Classification Working Group 1998), with pH = 6.6 (distilled water), organic C = 13.0 g kg−1, sand = 623 g kg−1, silt = 287 g kg−1, and clay = 90 g kg−1 in the top 100 mm. The experimental area has been registered with Pro-Cert Organic Systems Ltd. and received Canada Organic Regime Certification since 2018.
The experimental design included three adjacent fields under a corn-soybean-winter wheat/cover crop rotation with each crop present each year. The treatments included two cover crop tillage-termination methods (spring moldboard plow, spring chisel plow) in factorial arrangement with five cover crop-seeding methods, i.e., summer-seeded crimson clover (CC, Trifolium incarnatum L., seeding rate = 25 kg ha−1), summer-seeded hairy vetch (HV, Vicia villosa L. Roth, seeding rate = 25 kg ha−1), summer-seeded red clover (RCss, Trifolium pratense L., seeding rate = 12 kg ha−1), frost-seeded red clover (RCfs, seeding rate = 12 kg ha−1), and a conventional no-cover crop control (CK-C). Summer-seeding occurred after wheat harvest (July–August), and frost-seeding occurred in early spring (March–April). The individual field plots were 9.2-m wide (to allow 12 corn rows) by 20-m long, and the treatments were arranged in a randomized complete block design with four replicates. Each of the three adjacent fields consequently had two tillage-termination methods × five cover crop-seeding methods × four replicates for a total of 40 individual experimental plots.
Agronomic operations
Spring tillage-termination of the cover crops (May–June) was conducted using moldboard plowing (all cover crop biomass incorporated into the soil) or chisel plowing (cover crop biomass partially incorporated and partially left on the soil surface). The corn phase of the rotation was established by offset disking and packing the plowed cover crop soil and then corn planting (May–June) using a John Deere 7000 planter. The soybean phase was established by spring moldboard plowing the post-harvest corn stubble, then offset disking and packing, and then soybean planting (May–June) using John Deere 7000 planter. The winter wheat phase was established by offset disking and packing the post-harvest soybean stubble, then wheat planting (October) using a Case International seed drill. Shortly after winter wheat harvest (July), the soil was moldboard or chisel plowed and packed, and legume cover crops were planted (July–August) using an International Grain drill.
Corn cultivar DL6207 was planted in 2018–2021, and DL5472 in 2022; soybean cultivar B138CO was planted in 2018–2020, B213CO in 2021, and AAC Wigle in 2022; Slyvite common #1 winter wheat was planted in 2018, 2019, and 2022, and Branson in 2020 and 2021. The cultivars varied among years because of weather-induced variation in acceptable planting conditions, and changes in cultivar availability. The cover crop cultivars were consistent among years and treatments and included untreated soft common crimson clover, common hairy vetch, and common red clover.
Conventional weed management (herbicide application) and fertilization (synthetic fertilizers) practices were used on CK-C, with 180 kg N ha−1 applied to corn, 110 kg N ha−1 applied to winter wheat, and phosphorus and potassium applied to both according to local soil test recommendations. Organic weed management (rotary hoe, cultivation) and organic fertilization (ground phosphate rock/Tek-Mac High Phos and sulfate of potash applied to according to OMAFRA 2018, recommendations) were applied to all cover crop treatments. No pest management operations occurred because there were no pest/disease issues during the study. Further detail on agronomic operations is given in Table S2.
Corn, soybean, and winter wheat grain yields were determined separately for each treatment (i.e., MP, CP, CK-C, CC, HV, RCss, RCfs). Corn grain yields were reported at 15.5 wt. % seed moisture content; soybean and winter wheat grain yields were reported at 14.0 wt. % seed moisture content.
Cover crop sample, soil sampling, and analyses
To determine cover crop biomass and biomass N accumulation, live aboveground biomass samples were collected from the cover crop treatments (20 cm × 75 cm area, n = 2 per cover crop plot) in the spring (May) before cover crop termination. To determine near-surface soil NH4+-N and NO3–-N concentrations, composited soil core samples (0–30-cm depth, n = 8) were collected from all corn treatments at planting (May–June), and from the CK-C treatment at 1 week and 4 weeks after side-dress application of 28% UAN.
Plant-available N in the soil (i.e., mineral N as NH4+-N and NO3−-N) was determined via analysis of KCl extract using a Lachat autoanalyzer (Hach, Loveland, CO) and the protocol of Tel and Heseltine (1990). The KCL extract was obtained by weighing 20 g of field-moist soil into an Erlenmeyer flask, adding 100 mL 2 M KCl and shaking for 1 h, and then filtering the solution through Fisher Scientific Q5 filter paper. Total N concentrations in cover crop aboveground biomass and crop grain (after drying and grinding to 1 mm) was determined via dry combustion using a LECO-CN Analyzer (LECO, St. Joseph, MI). Corn chlorophyll status was measured at the R1 corn growth stage using a SPAD-502 leaf chlorophyll reader (Minolta Corporation, New Jersey, USA) and used as an estimate of corn N uptake. Further details on biomass and crop sampling are given in Table S2.
Statistical analyses
Analysis of variance was carried out using SAS Studio 3.81 (SAS Institute Inc, Cary, NC, USA). Cover crop biomass and biomass N were analyzed using a two-way completely randomized design with factorial arrangement (tillage-termination method × cover crop-seeding method)—both year and replicate were treated as random effects. Grain yields and seed N content for corn, soybean, and winter wheat were analyzed using a three-way completely randomized design (year × tillage-termination × cover crop-seeding) with replicate treated as a random effect. Significant differences among treatment means (P < 0.05) for biomass, biomass N, grain yield, and seed N content were determined using the least significant difference (LSD) test.
Results
Temperature and precipitation
Mean monthly air temperature at the field site ranged from 10.0–10.2 °C for 2018 and 2019, 10.9–11.1 °C for 2020 and 2021, and 12.5 °C for 2022 (Table S1). The first 2 years were similar to the 15-year average (10.3 °C); however, the last 3 years were appreciably warmer. Average air temperature was below freezing during January and February of each year, with 2018 and 2019 having the coldest January during the study period (− 4.2 °C and − 4.3 °C) and 2021 having the coldest February (− 5.0 °C). The 15-year average temperature for January and February were -3.0 °C and -3.4 °C, respectively (Table S1).
Annual field site precipitation varied among years (Table S1). The 2021 precipitation (972 mm) was similar to the 15-year average (950 mm), while the other 4 years were appreciably drier (676 to 854 mm). Growing season precipitation (June to September) followed roughly the same pattern as annual precipitation (Table S1); i.e., 2021 (447 mm) was somewhat a wetter than the 15-year average (362 mm), while the other years were appreciably drier (214 to 297 mm).
Cover crop biomass and biomass N
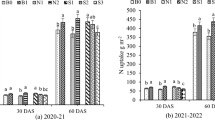
There was substantial variation in the amount of aboveground biomass and biomass N accumulated by the cover crops (Fig. 1). Over the 5-year study period, average annual aboveground biomass of the cover crops (i.e., biomass accumulated between summer planting and spring termination) followed the pattern CC (8.1 Mg ha−1) > RCfs (7.0 Mg ha−1) > RCss (6.7 Mg ha−1) > HV (5.9 Mg ha−1), while accumulated annual biomass N was RCfs ≈ CC ≈ HV (186–193 kg N ha−1) > RCss (155 kg N ha−1) (Table 1). These differences occurred because of annual weather variation (e.g., temperature, precipitation, degree of winter-kill), and because of different biomass characteristics among cover crop species, with varied N concentrations (23.4 g N kg−1 for CC and RCss, 27.8 g N kg−1 for RCfs, and 31.9 g N kg−1 for HV) and constant C concentrations (41.1–42.1 g C kg−1) (Table S3). Although the cover crop species are considered winter-hardy for southern Ontario, some degree of winter-kill always occurs, and this was particularly severe for CC in 2018, and for HV in both 2018 and 2019. Note also in Fig. 1 that accumulated aboveground biomass at cover crop termination was generally low in 2018, widely varying among cover crops in 2019, and relatively high in 2020–2022.

Legume aboveground biomass and aboveground biomass nitrogen before corn planting in spring 2018–2022. CC, summer-seeded crimson clover; HV, summer-seeded hairy vetch; RCss, summer-seeded red clover; RCfs, frost-seeded red clover
Soil inorganic N and corn N status
For 5-year average plant-available soil inorganic N, there were no tillage-termination effects or interactions between cover crop-seeding method and tillage-termination method; hence, only cover crop-seeding effects are reported (Fig. 2). Conventional control (CK-C) had significantly more mineral N (NO3− + NH4+) in the top 30 cm of soil than the cover crops at both 1 week after side-dress N application to corn (CK-C = 27.3 mg N kg−1 vs. cover crops = 13.1–16.3 mg N kg−1) and at 4 weeks after side-dress application (CK-C = 22.7 mg N kg−1 vs. cover crops = 8.9–10.5 mg N kg−1). This presumably reflects the fact that 180 kg of synthetic fertilizer N ha−1 was added to CK-C, but not to the cover crops. Interestingly, differences in SPAD corn plant N content at R1 growth stage (Fig. 3) were far less pronounced despite the above differences in soil inorganic N; i.e., MP (55.8a) > CP (54.1b), and CK-C (59.5a) > HV (54.7b) = RCfs (54.6b) > CC (53.4bc) > RCss (52.4c).

Five-year average soil inorganic nitrogen in the Harrow sandy loam at the 1st (about 1 week after corn planting) and the 2nd (4 weeks after corn planting) sampling. CK-C, no-cover crop conventional control; CC, summer-seeded crimson clover; HV, summer-seeded hairy vetch; RCss, summer-seeded red clover; RCfs, frost-seeded red clover

Five-year average of chlorophyll meter readings for corn at the R1 growth stage with respect to cover crop species and termination-tillage method from 2018 to 2022. The red-dotted line indicates N deficiency critical threshold which was the 95% of the reference (CK-C) (Varvel et al. 1997)
Grain yields
Corn grain yields varied among years, tillage-termination method, and cover crop-seeding method (Table 2, Fig. 4). In 2018 and 2019, corn grain yields for both MP and CP were significantly lower in the cover crops than in CK-C. Moldboard plow tillage had 12.5% higher yield than CP. Corn grain yields averaged across tillage treatments over 5 years under the cover crop treatments had the pattern: CK-C (13.3a Mg ha−1) > RCfs (10.9b Mg ha−1) > HV (10.6bc Mg ha−1) > CC (10.2bc Mg ha−1) > the RCss (9.5c Mg ha−1). Relative to CK-C, cover crops yielded 18.2% less corn grain under RCfs, 23.5% less under CC, 20.0% less under HV, and 28.1% less under RCss.

Corn grain yields (15.5% moist) under different cover crop types and termination method in the first 5-year organic period in Harrow, Ontario (2018–2022). CK-C, no-cover crop conventional control; CC, summer-seeded crimson clover; HV, summer-seeded hairy vetch; RCss, summer-seeded red clover; RCfs, frost-seeded red clover
Aside from variation across years, there were no significant tillage-termination or cover crop-seeding effects on soybean yield (Table 2), and the 5-year average soybean yields were similar among cover crops (3.2–3.4 Mg ha−1) (Fig. 5). On the other hand, winter wheat grain yields exhibited both substantial annual variation and significant cover crop effects (Fig. 6), with CK-C (5.9a Mg ha−1) > CC (4.3b Mg ha−1) = RCss (4.2b Mg ha−1) = HV (4.1b Mg ha−1) > RCfs (3.7c Mg ha−1) (Table 2).

Soybean yields (14.0% moist) under different cover crop types in the first 5-year organic period in Harrow, Ontario (2018–2022). CK-C, no-cover crop conventional control; CC, summer-seeded crimson clover; HV, summer-seeded hairy vetch; RCss, summer-seeded red clover; RCfs, frost-seeded red clover

Winter wheat yields (14.0% moist) under different cover crop types in the first 5-year organic period in Harrow, Ontario (2018–2022). CK-C, no-cover crop conventional control; CC, summer-seeded crimson clover; HV, summer-seeded hairy vetch; RCss, summer-seeded red clover; RCfs, frost-seeded red clover
Grain N content
There were significant year-to-year variations in seed N concentration (P < 0.001) for all three crops, and cover crop treatments affected seed N concentration (P < 0.001) for corn and winter wheat, but not for soybean (Table 3, 4). The N concentration in corn seed was 9–12% lower in the cover crop treatments relative to CK-C, and N concentration in winter wheat seed was 11–15% lower in the cover crop treatments relative to CK-C (Table 4).
Discussion
Red clover, commonly used as an N source for corn, holds significant potential for reducing the need for fertilizer nitrogen in rotation systems in Ontario, Canada. Studies such as Vyn et al. (2000), Loucks et al. (2018), and Yang et al. (2019, 2023) have highlighted the benefits of red clover to corn. In Ontario, red clover is typically frost-seeded between late March and early April. However, limited data exists comparing the N contribution of frost-seeded red clover to summer-seeded red clover in subsequent crops.
Previous research suggests that frost-seeded red clover may fix more N (Mohler et al. 1994; Sainju et al. 2002) due to its longer growing season in spring and summer, enabling greater biomass production and N fixation. This study observed not only a higher biomass in frost-seeded red clover compared to summer-seeded red clover but also a higher N concentration (27.8 g N kg−1) in frost-seeded red clover compared to summer-seeded red clover (23.4 g N kg−1). Consequently, frost-seeded red clover contributed significantly more biomass N (193 kg N ha−1) than summer-seeded red clover (155 kg N ha−1), resulting in higher corn yields. It should be noted that the increased N contribution from frost-seeded red clover to corn came at the expense of winter wheat yields, which will be discussed later.
Crimson clover exhibited an increase in aboveground biomass N (186 kg N ha−1) compared to the transition period (119 kg N ha−1) (Yang et al. 2019). Hairy vetch showed relatively high average biomass N content (193 kg N ha−1) compared to red clover and crimson clover. However, it was lower than the average during the transition period (240 kg N ha−1, Yang et al. 2019) due to severe winter-kill of hairy vetch in 2018 and 2019. The N contents in the aboveground biomass of hairy vetch in this study were similar to the range of 175–288 kg N ha−1 found in spring-harvested hairy vetch biomass planted the previous October in North Carolina (Liebman et al. 2018). Conversely, in the same region as this study (~ 120 km away), relatively low hairy vetch biomass N levels (41–139 kg N ha−1) at freeze-up and minimal cover crop biomass regrowth in the following spring were observed (Van Eerd 2018).
The conventional CK-C treatment exhibited higher soil mineral N levels at the first and second sampling times compared to the other treatments. This difference can be attributed to the one-time annual addition of N fertilizer in the CK-C treatment and the slower decomposition of legume biomass in the cover crop treatments. In Ontario, it is recommended to have pre-sidedress soil nitrate N (NO3−-N) levels of 18 mg N kg−1 or higher in the top 30 cm of soil for optimal corn yield without need for additional fertilizer N (OMAFRA 2018). In this study, the CK-C treatment demonstrated sufficient N in the soil approximately 1 week and even 4 weeks after corn N side-dressing, assuming continuous conversion of urea- and ammonium-N to nitrate. However, the cover crop treatments showed lower soil mineral N levels at both sampling dates, especially during the second sampling. These differences were clearly reflected in the chlorophyll levels of corn leaves, which were measured around 9 weeks after corn planting. Corn leaf chlorophyll levels in well-fertilized controls (e.g., 180 kg N ha−1 in the CK-C) were used as the reference N level. Nitrogen deficiency in the cover crop treatments, particularly in CC and RC, was identified when their chlorophyll levels fell below 95% of the reference level (Schepers 1994; Varvel et al. 1997). The variation in chlorophyll levels also corresponded to differences in corn yields. Although the average results over the 5-year period showed generally low soil mineral nitrogen, low chlorophyll readings, and low corn grain yields for all cover crop treatments, the chlorophyll readings exhibited significant year-to-year variations. For example, the cover crop treatments displayed higher chlorophyll levels (~ 60.0) relative to the CK-C treatment (58.5) under the MP in 2022 (Table S4). It is worth noting that the decomposition of legume biomass releases approximately equal amounts of inorganic and organic N (Yang et al. 2020), and the crop can utilize both forms of nitrogen (Miller et al. 2007; Zhang et al. 2017).
The yield of organic corn grain can vary significantly due to various factors, including soil quality, weather conditions, and crop management practices. In a study by Delbridge (2014), organic corn fields yielded an average of 75% compared to conventional corn fields on the same ground. In the current study, the average organic corn yield ranged from 72 to 82% of the conventional corn yield. However, when using MP termination, our organic corn yield was higher, reaching approximately 84 to 87% of the CK-C yield. On the other hand, under CP termination, the organic corn yield was lower, especially for CC and RC, at around 66 to 69% of the CK-C yield. Overall, the organic corn yields observed in our study (10.4 Mg ha−1) were greater than the average organic corn yield (8.4 Mg ha−1) reported by the United States Department of Agriculture (USDA National Agricultural Statistics Service 2020). A survey conducted among 526 organic corn farmers in Ohio, Indiana, Michigan, and Pennsylvania by Kumarappan et al. (2019) revealed an average organic grain corn yield of 8.4 Mg ha−1 in 2017, with a standard deviation of 2.7 Mg ha−1. In comparison, our average organic corn yield was higher, surpassing the yields of neighboring states above-mentioned. Additionally, the organic corn yields (8.6–11.6 Mg ha−1) observed in our study were higher than the organic corn grain yields (8.1–10.2 Mg ha−1) reported in a 3-year average study conducted by Champagne et al. (2021) following the transition to certified organic land using hairy vetch + triticale or red clover + timothy at Pennsylvania State. While there is limited data available on legume-fed organic corn in Canada, a study by Lavergne et al. (2021) demonstrated that legume cover crops (specifically pure pea as the primary nitrogen source) increased organic corn grain yield (5.6 Mg ha−1) compared to a no-cover crop control (4.0 Mg ha−1) in Québec, Canada.
In post-transition organic systems, the organic soybean yields are generally comparable to those achieved under conventional management (Kuegger 2003). However, a study by Porter (2003) in Minnesota reported that soybean yields in organic systems were 16–19% lower compared to conventional yields within a 2- to 4-year rotation. In a separate study conducted in New York by Cox et al. (2019), organic soybean yields were similar to conventional yields in 2015 and 2016 but approximately 11% lower in 2017 and 2018. In our study, organic soybean yields varied significantly over the years, ranging from 2.5 Mg ha−1 in 2020 to 3.9 Mg ha−1 in 2021. However, these yields were similar to conventional yields across these years. The average soybean yield during the organic period (2018–2022) was 3.3 Mg ha−1, which was lower than the yield during the transition period (4.1 Mg ha−1, 2014–2017) in the same trial (Yang et al. 2019). Nevertheless, it exceeded the 10-year average yield of soybeans (2.5 Mg ha−1) in a green manure–based organic production experiment (corn-soybean-oat/alfalfa-alfalfa) conducted in Minnesota by Mahoney et al. (2004). Furthermore, the soybean yield in our current study was higher than the soybean yield (2.7 Mg ha−1) reported during the 4-year transition period of a corn-soybean-winter spelt/red clover rotation in central New York (Caldwell et al. 2014).
The winter wheat yields not only varied over the years but also showed differences among the treatments, with lower yields observed for organic practices compared to conventional methods. In contrast to the consistent decrease in winter wheat yields for all treatments during the transition period as noted by Yang et al. (2019), the organic period saw a stabilization and even an increase in winter wheat yield. The average organic winter wheat yield of 4.1 Mg ha−1 in our study is slightly higher than the average of 3.9 Mg ha−1 during the transition period reported by Yang et al. (2019) and significantly greater than the 5-year average winter wheat yield of 2.9 Mg ha−1 (2015–2019) in the USA (Langemeier and O’Donnell 2020). Our findings align with Gopinath et al. (2008), who reported initial yield losses in organic winter wheat during the conversion period. However, the decline followed by an increase in organic winter wheat yields observed in our study differs from the results presented by Eltun et al. (2002), who suggested that yield losses may persist for longer durations.
Previous studies have shown that frost-seeding red clover into an established winter wheat stand can fix significant amounts of N without negatively affecting winter wheat yield (Hartl 1989; Hesterman et al. 1992; Vyn et al. 2000; Schipanski and Drinkwater 2010). However, our study found that frost-seeding red clover led to a 10–15% reduction in wheat yield compared to no frost-seeding. We believe this yield reduction is due to competition between red clover and wheat for soil moisture and nutrients, especially in our organic fields where N availability is limited. Queen et al. (2009) suggested that this effect is a result of light, moisture, and nutrient competition from underseeded red clover in winter wheat.
Organic farming is expected to promote soil health and enhance nutrient production through practices such as crop rotation, cover cropping, and the use of compost/manure and other organic fertilizers. However, our results showed lower N contents in organic corn and winter wheat compared to conventional grains, whereas the N content was similar in both organic and conventional soybeans. A long-term agricultural research (LTAR) experiment demonstrated similar N contents for soybeans and corn between organic and conventional production systems (Delate et al. 2013). In Europe, organic production of winter wheat has generally been associated with low grain protein contents (Gooding et al. 1993; Lueck et al. 2006), with N supply being identified as a contributing factor influencing grain protein content (Lueck et al. 2006). However, it is important to note that apart from farming practices, the N content of grains can be influenced by other factors, including soil and climate. Hence, additional research is necessary to gain a comprehensive understanding of the variations in nitrogen content between organic and conventional grains and to determine if any of these variances might have implications for other aspects of grain quality, such as protein or amino acid composition.
Conclusions
Summer-seeded legume cover crops, including hairy vetch, red clover, and crimson clover, following winter wheat harvest in southern Ontario, can serve as a reliable nitrogen source for organic soybean-winter wheat-corn production. In the first 5 years of organic farming after the transition period, these legume cover crops accumulated substantial aboveground biomass nitrogen before corn planting. On average, summer-seeding hairy vetch accumulated 193 kg N ha−1, crimson clover accumulated 186 kg N ha−1, red clover accumulated 155 kg N ha−1, and frost-seeded red clover accumulated 193 kg N ha−1. These cover crops provided satisfactory N levels for subsequent corn production. During the first 5 years of organic farming, corn yields under moldboard plow termination of legumes ranged from 11.2 to 11.5 Mg ha−1 for hairy vetch, crimson clover, and frost-seeded red clover, which was approximately 13–16% lower than conventional corn yields. In comparison, corn yields under chisel plow termination ranged from 9.2 to 10.0 Mg ha−1 for hairy vetch, crimson, and frost-seeded red clover, representing a decrease of about 23–31% compared to conventional corn yields. Among the cover crop treatments, summer-seeded red clover resulted in the lowest corn yields. Soybean yields varied over the years during the first 5 years of organic production, but they were similar in the cover crop treatments compared to conventionally managed plots. Winter wheat yields were significantly higher for conventionally managed plots (5.9 Mg ha−1) compared to summer-seeded legumes (4.2 Mg ha−1), with the lowest yields observed in frost-seeded red clover (3.7 Mg ha−1) likely due to competition between the early seeded red clover and winter wheat for moisture and nitrogen. The results suggest that moldboard plow termination of cover crops can suppress weeds and maximize biomass N mineralization. Additionally, the study indicates that frost-seeded red clover had a greater impact on corn yields compared to summer-seeded RC, although this benefit came at the expense of winter wheat yields. Overall, this research highlights the effectiveness of summer-seeding legumes after winter wheat harvest as a primary nitrogen source in a soybean-winter wheat-corn rotation in southern Ontario, Canada.
Data availability
Please contact the correspondence author.
References
Archer DW, Jaradat AA, Johnson JM-F, Weyers SL, Gesch RW, Forcella F, Kludze HK (2007) Crop productivity and economics during the transition to alternative cropping systems. Agron J 99:1538–1547
Blaser BC, Gibson LR, Singer JW, Jannink JL (2006) Optimizing seeding rates for winter cereal grains and frost-seeded red clover intercrops. Agron J 98:1041–1049
Bruns HA (2012) Concepts in crop rotations. In: Aflakpui G (ed) Agricultural science. InTech. Available from: https://www.intechopen.com/books/agricultural-science/conceptsin-crop-rotation
Brust J, Claupein W, Gerhards R (2014) Growth and weed suppression ability of common and new cover crops in Germany. Crop Protec 63:1–8
Caldwell B, Mohler CL, Ketterings QM, DiTommaso A (2014) Yields and profitability during and after transition in organic grain cropping systems. Agron J 106:871–880
Canada Organic Trade Association (COTA) (2021) Quick facts about organic in Canada. https://canada-organic.myshopify.com/products/2021-organic-quick-facts
Canadian General Standards Board (2011) Organic production systems general principles and management standards. Government of Canada, Ottawa, ON. https://www.tpsgc-pwgsc.gc.ca/ongc-cgsb/programme-program/normes-standards/internet/bio-org/documents/032-0310-2008-eng.pdf (Accessed on April 17, 2018)
Champagne RJ, Wallace JM, Curran WS, Barbercheck ME (2021) Rotational no-till and tillage-based organic corn produce management tradeoffs in the Northeast. Agron J 113:5348–5361. https://doi.org/10.1002/agj2.20823
Coombs C, Lauzon JD, Deen B, Van Eerd LL (2017) Legume cover crop management on nitrogen dynamics and yield in grain corn systems. Field Crops Res 201:75–85
Cox W, Cherney J, Sorrells M (2019) agronomic comparisons of organic and conventional soybean with recommended and high inputs during the first 4 years of organic management. Agron 9:602. https://doi.org/10.3390/agronomy9100602
Delate K, Cambardella C, Chase C, Johanns A, Turnbull R (2013) The long-term agroecological research (LTAR) experiment supports organic yields, soil quality, and economic performance in Iowa. Crop Manag 12:1–13
Delbridge TA (2014) Profitability and adoption of organic agriculture: essays on the decision to transition. Retrieved from The University of Minnesota Digital Conservancy. https://hdl.handle.net/11299/165304
Eltun R, Korsæt A, Nordheim O (2002) A comparison of environmental, soil fertility, yield, and economical effects in six cropping systems based on an 8-year experiment in Norway. Agric Ecosyst Environ 90:155–168
Gomiero T, Pimental D, Paoletti MG (2011) Environmental impact of different agricultural management practices: conventional vs. organic agriculture. Crit Rev Plant Sci 30:95–124
Gooding MJ, Davies WP, Thompson AJ, Smith SP (1993) The challenge of achieving breadmaking quality in organic and low input wheat in the UK-A review. Asp Appl Biol 36:189–198
Gopinath KA, Mina BL, Kundu S, Gupta HS (2008) Influence of organic amendments on growth, yield and quality of wheat and soil properties during transition to organic production. Nutr Cycling Agroecosyst 82:51–60
Hartl W (1989) Influence of undersown clovers on weeds and on the yield of winter wheat in organic farming. Agric Ecosyst Environ 27:389–396
Hesterman OB, Griffin TS, Williams PT, Harris GH, Christenson DR (1992) Forage legume-small grain intercrops: nitrogen production and response of subsequent corn. J Prod Agric 5:340–348. https://doi.org/10.2134/jpa1992.0340
Hilker B (2021) Examinations of red clover (Trifolium pratense L.) as a cover crop. M.S. degree dissertation. Department of Plant Agriculture, University of Guelph. https://hdl.handle.net/10214/24100
Hunt ND, Hill JD, Liebman M (2019) Cropping system diversity effects on nutrient discharge, soil erosion, and agronomic performance. Environ Sci Technol 53:1344–1352
Kalinova J (2010) Chapter 14: Allelopathy and organic farming. In: Lichtfouse E (ed) Sociology, organic farming, climate change and soil science. Sustainable agriculture reviews (vol 3, pp 379–419). https://doi.org/10.1007/978-90-481-3333-8_14
Kamh M, Abdou M, Chude V, Wiesler F, Horst WJ (2002) Mobilization of phosphorus contributes to positive rotational effects of leguminous cover crops on maize grown on soils from northern Nigeria. J Plant Nutri Soil Sci 165:566–572
Kuegger G (2003) Organic soybean production. National Center for Appropriate Technology. https://coolbean.info/pdf/soybean_research/library/forage_and_food_production/Organic_Soybean_Production.pdf
Kumarappan S, Jackson-Smith D, Brock C (2019) A profile of organic corn production in the Midwest and Northeast. PART II: Economics of Organic Corn Production. The Ohio State University. https://go.osu.edu/orgcorn_report2
Langemeier M and O’Donnell M (2020) Conventional Vs. organic grains: 5-year comparison of return. https://www.agfax.com/2020/09/08/conventional-vs-organic-grains-5-year-comparison-of-returns/
Lavergne S, Vanasse A, Thivierge M-N, Halde C (2021) Nitrogen content of pea-based cover crop mixtures and subsequent organic corn yield. Agron J 113:3532–3547
Liebhardt WC, Andrews RW, Culik MN, Harwood RR, Janke RR, Radke JK, Rieger-Schwartz SL (1989) Crop production during conversion from conventional to low-input methods. Agron J 81:150–159
Liebman AM, Grossman J, Brown M, Wells MS, Reberg-Horton SS, Shi W (2018) Legume cover crops and tillage impact nitrogen dynamics in organic corn production. Agron J 110:1046–1057
Loucks CES, Deen W, Gaudin ACM, Earl HJ, Bowley RS, Martin RC (2018) Genotypic differences in red clover (Trifolium pratense L.) response under severe water deficit. Plant Soil 425:401–414
Lueck L, Schmidt CS, Cooper JM, Hall CM, Shotton PN, Leifert C (2006) Effect of organic, low-input and conventional production systems on yield and quality of winter wheat. Asp Appl Biol 80:135–140
Lynch DH (2017) Nutrient cycling and soil health in organic cropping systems - importance of management strategies and soil resilience. Sustain Agric Res 4:80–88
MacRae RD, Hill SB, Mehuys GR, Henning J (1990) Farm-scale agronomic and economic conversion from conventional to sustainable agriculture. Adv in Agron 43:155–198
Mahoney PR, Olson KD, Porter PM, Huggins DR, Perillo CA, Crooksto RK (2004) Profitability of organic cropping systems in southwestern Minnesota. Renew Agric Food Syst 19:35–46
McBride WD, Greene C (2015) Foreman L and Ali M (2015) The profit potential of certified organic field crop production, ERR-188. U.S. Department of Agriculture, Economic Research Service
Miller AE, Bowman WD, Suding KN (2007) Plant uptake of inorganic and organic nitrogen: neighbor identity matters. Ecology 88:1832–1840
Mohler CL, Johnson NM, Ostlie MH (1994) Nitrogen fixation by red clover frost-seeded into corn. Agron J 86:948–953
Moyer J, Nichols K, Bhosekar V (2017) A fifteen year review summarizing effects of. conventional and organic farming systems on. soils, nutrition, environment, economics and yields (1981–1995). Ansian J Sci Tech 8:4628–4634
Ontario Ministry of Agriculture Food and Rural Affairs (OMAFRA) (2018) Agronomy guide for field crops. In: Brown C (ed) Publication 811, Queen’s Printer for Ontario, Toronto, Canada
Pang XP, Letey J (2000) Organic farming: challenge of timing nitrogen availability to crop nitrogen requirements. Soil Sci Soc Am J 64:247–253
Ponisio LC, M’gonigle LK, Mace KC, Palomino J, De Valpine P, Kremen C (2015) Diversification practices reduce organicto conventional yield gap. Proc Royal Soc B 282:20141396. https://doi.org/10.1098/rspb.2014.1396
Porter PM (2003) Organic and other management strategies with two- and four-year crop rotations in Minnesota. Agron J 95:233–244
Queen A, Earl H, Deen W (2009) Light and moisture competition effects on biomass of red clover underseeded to winter wheat. Agron J 101:1511–1521
Sadeghpour A, Ketterings QM, Godwin GS, Czymmek KJ, Vermeylen F (2017) Agro-environmental consequences of shifting from nitrogen- to phosphorus-based manure management of corn. Soil Sci Soc Am J 81:1127–1138
Sainju UM, Singh BP, Whitehead WF, Wang S (2002) Winter cover crop residue and tillage effects on corn nitrogen uptake and soil nitrogen availability. Agron J 9:501–509
Schepers JS (1994) New diagnostic tools for tissue testing. Commun Soil Sci Plant Anal 25:817–826
Schipanski ME, Drinkwater LE (2010) Nitrogen fixation of red clover interseeded with winter cereals across a management-induced fertility gradient. Nutr Cycl Agroecos 90:105–119
Seidel R, Moyer J, Nichols K, Bhosekar V (2017) Studies on long-term performance of organic and conventional cropping systems in Pennsylvania. Org Agric 7:53–61
Smith EG, Clapperton MJ, Blackshaw R (2004) Profitability and risk of organic production systems in the northern Great Plains. Renew Agric Food Syst 19:152–158
Soil Classification Working Group (1998) The Canadian system of soil classification. Publ. 1646. Agriculture and Agri-Food Canada, Ottawa, ON
Spargo JT, Cavigelli MA, Mirsky SB, Maul JE, Meisinger JJ (2011) Mineralizable soil nitrogen and labile soil organic matter in diverse long-term cropping systems. Nutr Cycl Agroecosyst 90:253–266
Stockdale EA, Watson CA (2009) Biological indicators of soil quality in organic farming systems. Renew Agri Food Syst 24:308–318
Tel DA, Heseltine C (1990) The analyses of KCL soil extracts for nitrate, nitrite and ammonium using a TRAACS 800 analyzer. Commun Soil Sci Plant Analy 21:1681–1688
United States Department of Agriculture (USDA) (2018) National organic program. https://www.ams.usda.gov/about-ams/programs-offices/national-organic-program
USDA National Agricultural Statistics Service (2020) Organic Production. Retrieved from https://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Organic_Production/index.php
USDA (2022) Certified Organic Survey - 2021 Summary. National Agricultural Statistics Service. Release December 15, 2022. https://downloads.usda.library.cornell.edu/usda-esmis/files/zg64tk92g/2z10z137s/bn99bh97r/cenorg22.pdf
Van Eerd LL (2018) Nitrogen dynamics and yields of fresh bean and sweet corn with different cover crops and planting dates. Nutr Cycl Agroecosyst 111:33–46
Varvel GE, Schepers JS, Francis DD (1997) Ability for in-season correction of nitrogen deficiency in corn using chlorophyll meters. Soil Sci Soc Am J 61:1233–1239
Vyn TJ, Faber JG, Janovicek KJ, Beauchamp EG (2000) Cover crop effects on nitrogen availability to corn following wheat. Agron J 92:915–924
Yang XM, Drury CF, Reynolds WD, Reeb MD (2019) legume cover crops provide nitrogen to corn during a three-year transition to organic cropping. Agron J 111:3253–3264
Yang XM, Drury CF, Reynolds WD, Phillips LA (2020) Nitrogen release from shoots and roots of crimson clover, hairy vetch, and red clover. Can J Soil Sci 100:179–188
Yang XM, Reynolds WD, Drury CF, Reeb MD (2023) Impacts of summer-seeded legume cover crops and termination-tillage method on nitrogen availability to corn in rotation. Front Soil Sci 2:1082940. https://doi.org/10.3389/fsoil.2022.1082940
Zhang L, Yang XM, Gao D, Wang L, Li J, Wei Z, Shi YL (2017) Effects of poly-γ-glutamic acid (γ-PGA) on plant growth and its distribution in a controlled plant-soil system. Sci Rep 7:6090. https://doi.org/10.1038/s41598-017-06248-2
Acknowledgements
This study was conducted as part of the Organic Science Cluster 3, an Industry-led Research and Development Program supported by Agriculture and Agri-Food Canada (AAFC). We would like to express our gratitude to AAFC and the Grain Farmers of Ontario (c2017ag24) for their financial support. We extend our sincere appreciation to the Harrow Research and Development Centre Farm Crew, Dr. T. Oloya, and the summer and co-op students for their valuable technical assistance throughout the study.
Funding
Open Access funding provided by Agriculture & Agri-Food Canada. This work was supported by the Agriculture and Agri-Food Organic Science Cluster-3 program under Grant [ASC-13] and Grain Farmers of Ontario under Grant [c2017ag24].
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by M-AR and XY. The first draft of the manuscript was written by XY, and all authors commented on posteriorly versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
The authors have no ethical conflict and waivers to declare.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yang, X., Drury, C.F., Dan Reynolds, W. et al. Legume cover crop as a primary nitrogen source in an organic crop rotation in Ontario, Canada: impacts on corn, soybean and winter wheat yields. Org. Agr. 14, 19–31 (2024). https://doi.org/10.1007/s13165-023-00452-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13165-023-00452-3