
Abstract
Stream restoration includes a number of different approaches intended to reduce sediment and nutrient export. Legacy sediment removal (LSR) and floodplain reconnection (FR) involve removing anthropogenically derived sediment accumulated in valley bottoms to reconnect incised streams to their floodplains. These projects also present an opportunity to create high-quality riparian and wetland plant communities and provide information about the early stages of wetland vegetation development and succession. We surveyed vegetation immediately after restoration at three sites and at three additional sites 1–3 years post-restoration to determine how LSR/FR affects riparian plant communities. Restoration increased the prevalence of hydrophytic herbaceous species at all sites, suggesting these projects successfully reconnected the stream to the floodplain. Pronounced decreases in woody basal area and stem density likely also influenced an increase in native and graminoid species after restoration. Only 16% of the indicator species identified for restored reaches were planted as part of the restoration, suggesting the local seed bank and other seed sources may be important for vegetation recovery and preservation of regional beta diversity. Although vegetation quality increased after restoration in reaches with initially low-quality herbaceous vegetation, vegetation quality did not improve or decreased after restoration in reaches with higher-quality vegetation before restoration. The practice of LSR/FR has the potential to improve the quality of some riparian vegetation communities, but the preservation of high-quality forested areas, even if they are atop legacy sediment terraces, should be considered, particularly if reductions in nutrient export do not offset losses in tree canopy.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Accumulation of anthropogenically derived floodplain sediment is observed globally and can occur as a result of intensive land clearing, agriculture, or mining activities (Flitcroft et al. 2022; James et al. 2020; Vauclin et al. 2020; Wohl et al. 2021). In the Mid-Atlantic United States, extensive forest clearing in the 1700s and early 1800s coincided with the construction of milldams and other impoundments. These impoundments often resulted in thick layers of anthropogenically derived legacy sediment being deposited atop pre-European floodplains and burying small anabranching stream systems (Langland et al. 2020; Merritts et al. 2011; Walter and Merritts 2008). Subsequent dam breaches caused streams to incise through these artificially aggraded floodplains, resulting in the entrenchment that is characteristic of many Mid-Atlantic streams (Walter and Merritts 2008). More recently, stream incision has been further exacerbated by increased impervious surface cover (ISC) in many watersheds (Walsh et al. 2005), which channels high volumes of surface runoff into streams, further increasing erosion and entrenchment (Hupp et al. 2013; McMillan et al. 2014; Walter and Merritts 2008).
Stream entrenchment is associated with changes to riparian plant community composition and the reduction of many critical ecosystem services performed by floodplains. Entrenchment creates elevated floodplains, resulting in lower water tables and decreased overbank flooding (Groffman et al. 2002) that disconnect the root systems of vegetation from the water table. In many areas, hydrophytic wetland and riparian plant species are replaced by upland species (Groffman et al. 2003; Voli et al. 2009). Lower groundwater tables are associated with reduced denitrification rates, decreasing the ability of these impaired systems to function as landscape-level nitrogen sinks (Gold et al. 2001; Groffman et al. 2002; Weitzman and Kaye 2017). In addition to reducing ecosystem services, legacy sediments are potentially a large source of suspended sediment (Hartranft et al. 2011; Hupp et al. 2013; Schenk and Hupp 2009), nitrogen (Forshay et al. 2022; Weitzman et al. 2014) and phosphorus that enter Maryland streams and, ultimately, the Chesapeake Bay (Inamdar et al. 2020; Walter et al. 2007).
In 2010, to address the negative impacts of excess sediment and nutrient inputs to the Chesapeake Bay, the US Environmental Protection Agency developed Total Maximum Daily Load (TMDL) regulations (Williams et al. 2017). Stream restoration has emerged as a popular Best Management Practice (BMP) to reduce nutrient and sediment loads, with the most common restoration techniques relying largely on stabilizing stream banks and deliberately designing channel forms to address erosion (Palmer et al. 2014b; Williams et al. 2017). However, recent reviews suggest these methods have not been as effective as anticipated (Meisenbach et al. 2012; Palmer et al. 2014a, 2014b). The combination of legacy sediment removal and floodplain reconnection (“LSR/FR,” also referred to herein as “restoration”) has emerged as a process-based restoration technique that alters floodplain geomorphology and is also designed to restore ecosystem structure and biological processes (Beechie et al. 2010; Palmer et al. 2014b). By removing as much of the anthropogenic legacy sediment as possible, the goal of this technique is to return the floodplain to its pre-European elevation, eliminate a major source of sediment input, and increase overbank flows and floodplain residence time (Hartranft et al. 2011; Langland et al. 2020). In 2020, the Chesapeake Bay Program Water Quality Goal Implementation Team approved legacy sediment removal as an accepted BMP for addressing Total Maximum Daily Loads in the Chesapeake Bay Watershed (Altland et al. 2020).
In the Chesapeake Bay region, research into the efficacy of stream restoration, including LSR/FR, has been focused largely on project goals pertaining to water quality, such as denitrification and sediment retention rates (Filoso and Palmer 2011; Forshay et al. 2022; Kaushal et al. 2008a; Langland et al. 2020; McMahon et al. 2021). However, these restoration projects also present the opportunity to reestablish hydrophytic species in floodplains and create diverse, high-quality riparian and wetland plant communities (Fraaije et al. 2019; Göthe et al. 2015). Conversely, the priority effects of revegetating many project sites with the same few species (Beauchamp et al. 2015; Kaase and Katz 2012; Lesage et al. 2018), coupled with the elimination of a site’s unique seed bank through sediment removal (DeBerry and Perry 2004) could cause regional homogenization of riparian habitats (Holl et al. 2022). Additionally, riparian ecosystems are disturbance-adapted systems that receive seeds from a wide area (Fryirs and Carthey 2022; Nilsson et al. 2010), making them particularly susceptible to colonization by invasive plant species (Catford and Jansson 2014; Cooper et al. 2017; Planty-Tabacchi et al. 1996; Richardson et al. 2007; Tabacchi et al. 2005; Vidra et al. 2006). This susceptibility to invasion may be exacerbated by the extreme disturbance involved with legacy sediment excavation (Larson et al. 2019; McLane et al. 2012; Wise 2017).
The disturbance and sediment removal associated with LSR/FR restoration projects provide an opportunity to examine the trajectory of the early stages of wetland succession (Mitsch and Gosselink 2015; Noon 1996; van der Valk 1981). While succession theory predicts that the early stages of vegetation recovery should be dominated by annual species, some studies of created wetlands have found a predominance of perennial species in early wetland succession (DeBerry and Perry 2004). Understanding how these sites develop after restoration and the potential role and time scale of seed dispersal into these sites could inform the active or passive revegetation strategies used in these projects.
In this project, we surveyed vegetation at six LSR/FR restoration sites near Baltimore, Maryland to characterize short-term changes to the vegetation community immediately following restoration (Fig. 1). A companion study investigated changes to nitrogen, phosphorus, and sediment exports from these sites (McMahon et al. 2021). If LSR/FR is a successful restoration approach, we anticipated that a shallower water table depth and an increase in the frequency of overbank flows would result in a shift from upland and facultative to more hydrophytic plant species (Booth and Loheide 2012; Fraaije et al. 2019; Hammersmark et al. 2010). If these projects result in biotic homogenization and invasion by exotic species (Kaase and Katz 2012; Lesage et al. 2018; Qian and Guo 2010), we expected to see decreased alpha and beta diversity and increased importance of exotic and invasive plant species in restored reaches when compared to unrestored reaches. We were also interested in comparing the importance of annual versus perennial species immediately and then 1–3 years after restoration at these sites.

First Mine Branch in September 2016 before restoration (top), in June 2017 as restoration was nearly finished (middle), and in May 2018 nearly one year after restoration (bottom). (Photos by V. Beauchamp and J. Moore)
Methods
Site Selection
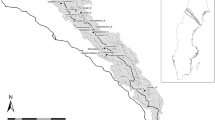
All six study sites were situated along third-order streams in the Piedmont region of Central Maryland (Fig. 2) and were selected to span a range of project length (63 – 1621 m) and watershed level ISC (0.5 – 48.6%). First Mine Branch, Plumtree Run, and Bear Cabin Branch were restored in 2017–2018, during the study period (Table 1). These three sites included a project reach sampled both pre-restoration and post-restoration and an additional unrestored “control” reach sampled concomitantly with the project reach, providing a Before-After-Control-Impact (BACI) design. Cabbage Run, Beetree Run, and North Stirrup Run were restored 1–3 years prior to this study, and each included an unrestored and a restored reach, providing a space-for-time substitution (SFT) design (Fig. 3). Reaches within a site were immediately adjacent, except for Beetree Run where the unrestored and restored reaches were separated by 4.8 km. Even at sites where unrestored reaches were adjacent to restored/project reaches, the reaches were often on different properties which may have had different land use histories. While the control reaches in the BACI sites and the unrestored reaches in the SFT sites are not exact matches to pre-restoration conditions of the restored reaches, together they represent the range of conditions we could have reasonably expected to be found before restoration.

Locations of study sites in Baltimore and Harford counties, Maryland

Project design schematic. Note: For figures in color, readers are referred to the online version of this paper
The control, pre-restoration and unrestored reaches either contained mature forest with floodplains dominated by Liriodendron tulipifera (tulip poplar), Salix nigra (black willow), Lindera benzoin (spicebush), Rosa multiflora (multiflora rose) and Rubus pensilvanicus (Pennsylvania blackberry) or were recently retired from agriculture with floodplains dominated by Microstegium vimineum (Japanese stiltgrass) and Phalaris arundinacea (reed canary grass). Streambanks before restoration ranged from 0.5 to 2.5 m high from toe slope to top of bank. All restoration projects were conducted by the same local consulting firm and involved extensive sediment excavation with a target bank height of 0.5 m, re-routing of the stream channel, and installation of toe wood and log vane structures. As a consequence of the legacy sediment excavation and removal, nearly all trees and understory vegetation were removed from within the limit of disturbance defined for each project (Fig. 1). Floodplains were actively revegetated with a mix of tree and shrub species planted on the floodplain as container-grown plants or installed along streambanks as live-stakes. At some sites, a mix of graminoid species was also seeded or planted along streambanks to enhance bank stabilization (Tables S1 and S2), Fig. 1b).
Field Sampling
Study reach length varied from 63 to 1621 m. To capture the inherent heterogeneity in each reach but still complete all seasonal sampling within 4–6 weeks, reaches were sampled with 3 to 18 transects spaced at 25 to 70 m intervals. Transects were oriented perpendicularly to the stream and spanned the floodplain to a maximum of 50 m on each bank. In some cases, the floodplain width sampled surpassed the limit of construction disturbance and extended into the adjacent undisturbed riparian area. Cover of all herbaceous species, regardless of height, and of woody individuals under 1 m tall was estimated to the nearest 10% in 1 m2 plots along each transect at 5 or 10 m intervals depending on transect length. Herbaceous layer vegetation was sampled in late spring and early fall to capture seasonal changes. Due to the timing of construction, fall sampling of the pre-restoration reaches at First Mine Branch and Plumtree Run was conducted in 2016. All other reaches were sampled in spring and fall 2017 and 2018. Species unfamiliar to the researchers were identified using the Flora of Virginia (Weakley et al. 2012).
Woody vegetation greater than 1 m tall was quantified, ideally, in 400 m2 plots (20 × 20 m) placed randomly along each transect, but plot dimensions and area were modified when needed due to topography. One plot was located on each bank except in cases where the stream abutted the upland slope on one side. All woody individuals greater than 1 m tall and greater than 0.5 cm stem diameter at base were identified and tallied by 0.5–2.0 cm or 2.1–5.0 cm diameter size classes. Diameter at breast height was measured on individuals larger than 5.0 cm diameter. Woody vegetation was sampled once on unrestored and restored reaches at SFT sites and before and after restoration on project reaches at BACI sites. Woody vegetation was also sampled once on control reaches at BACI sites to allow for future comparisons of community change, but those data are not reported for this study as we anticipated minimal change in woody vegetation at control reaches over one year. We did not distinguish between actively planted and passively recruited vegetation during sampling.
Data Analysis—Plant Community Profiles
For each year of herbaceous layer sampling, the two seasonal data sets were combined using the maximum cover values for species recorded in both seasons. Plant communities in each reach were described by habit and duration, wetland indicator status, conservatism coefficient, and nativity. Habit, duration, and wetland indicator status were assigned according to the classifications in the USDA PLANTS database (USDA 2020). Plot level cover-weighted wetland indicator scores (WIS) were calculated by multiplying by the relative cover of each species in each plot by the numeric WIS for that species and summing all the scores in the plot (Wentworth et al. 1988). Invasive species were defined as those appearing in Swearingen and Fulton (2022), as these species have been identified as “highly invasive” by regional scientists and land managers. All other species identified as introduced in the USDA PLANTS database (USDA 2020) were classified as exotic. Coefficients of conservatism (CC) for all native species (Chamberlain and Ingram 2012) were taken from the Mid-Atlantic Piedmont database within The Universal Floristic Quality Assessment Calculator (Freyman et al. 2015). When CC values were unavailable for the Mid-Atlantic Piedmont region, the Mid-Atlantic Coastal Plain and Mid-Atlantic Ridge & Valley databases were used as supplements. Coefficients of conservatism range from 0 to 10 and represent an estimated sensitivity of a particular species to anthropogenic disturbance. All non-native species are assigned a score of 0 and CC scores increase as the range of a species' ecological tolerance decreases (Chamberlain and Ingram 2012). A Floristic Quality Index (FQI) is used to determine the ecological value of a site based on its species composition and was calculated for each plot by multiplying the average CC value of the plot by the square root of plot species richness (Chamberlain and Ingram 2012). Shannon diversity values were also calculated for each plot using the equation \(H=-\sum {p}_{i}*{\text{ln}}({p}_{i})\) where \({p}_{i}\) is the proportion of total plot cover composed of species i (Magurran 2004). We calculated importance values for the herbaceous species at each reach with the importance value function from the BiodiversityR package using only cover and frequency data. We also identified herbaceous species that, based on their relative abundance and occurrence, are significant indicators of unrestored or restored reaches. For this analysis we combined reaches across the BACI and SFT sites and used the multipatt function in the R package indicspecies.
To examine the effect of restoration on local herbaceous layer beta diversity, we calculated the compositional dissimilarity (q = 2, Morista-Horn) among all herbaceous plots within each reach using abundance data and the dissCqN function in the R package dissCqN. To examine the effect of restoration on regional beta diversity, the compositional similarity among reaches (q = 2) within each treatment group and year combination was calculated with the SimilarityMult function in the SpadeR package. These values were subtracted from 1 to convert them to a measure of dissimilarity. Standard errors were calculated from 200 bootstrap replications (Chao et al. 2008).
Biodiversity descriptors for woody vegetation were calculated using the same process as for the herbaceous vegetation, except for average richness and within and among reach beta diversity which could not be calculated as topography prevented us from making the plots a consistent size. Woody importance values were calculated with basal area and density expressed on a per m2 basis to account for the variation in plot size.
Data Analysis – Effect of Restoration
Linear mixed effect models were used to test for the effect of restoration on plot-level herbaceous richness (total, native, exotic, and invasive), the proportion of perennial species, WIS, FQI, and Shannon diversity. The function lme in the R package nlme was used to analyze responses for the BACI and SFT sites separately. Plots nested within transects and sites were entered as random effects, with treatment (control/unrestored or project/restored reach) and year of sample collection (year 1 or year 2) and their interaction as fixed effects. For BACI sites, a significant treatment*year interaction indicates that the project reach changed between the first and second year, independent of the control reach, presumably due to the restoration. For SFT sites, a significant treatment effect would show that unrestored and restored reaches differed significantly for the variable of interest over both years of sampling. Little year-to-year change was expected for woody vegetation on control reaches so the effects of restoration on plot-level woody variables was examined by comparing conditions between pre- and post-restoration reaches at BACI sites and restored and unrestored reaches at SFT sites. Linear models for woody data at BACI and SFT sites were run separately and consisted of the main effect of restoration status and the random effects of plots nested within transects and sites. The effect of restoration on woody richness could not be analyzed as neither woody plots nor reaches were a consistent size. Instead, we simply present the total number of woody species encountered at each reach.
We also used linear mixed-effect models to test for the effect of restoration on local herbaceous (within-reach) beta diversity. Because this value was calculated at the site level, the model contained only the random effect of site, while treatment, year, and their interaction were included as fixed effects. Responses for the BACI and SFT sites were analyzed separately. Significant differences in regional (among-reach) beta diversity among treatment and year combinations were determined using the function aovSufficient from the HH package which conducts ANOVA for groups using group means and within-group standard deviation.
The relationship between the plot-level change in each response variable (richness, diversity, FQI and WIS) and the average plot-level pre-restoration or unrestored value for each reach was examined with Pearson correlations, as was the relationship between the change in herbaceous and woody WIS and the change in woody basal area. We used Pearson correlations to determine if changes in any dependent variables were related to project length and Spearman correlations to determine if changes in any dependent variables were related to watershed-level ISC as ISC did not follow a normal distribution. All p-values were Bonferroni corrected for multiple comparisons.
Data Analysis—Community Composition
Non-metric multidimensional scaling (NMDS) ordination was used to visualize differences in plant community composition among reaches and years using species importance values calculated for both the herbaceous and woody layers. The herbaceous dataset consisted of the control and project reaches at the three BACI sites and the unrestored and restored reaches of the three SFT sites, each sampled in two consecutive years. The woody dataset consisted of the project reach at each BACI site sampled before and after restoration and the unrestored and restored reaches at the three SFT sites each sampled once. Each NMDS analysis was conducted using the metaMDS function in the R package vegan with a Bray distance and 100 random starts.
Differences in community composition among reach types were examined in several ways. We compared species community composition among reach types using the adonis2 function in the vegan package in R, with the same data and groupings used for the NMDS and conducted post-hoc pairwise comparisons with the pairwise.adonis function. We looked at change in group importance by averaging species importance values across sites within each reach type (all unrestored reaches at BACI and SFT sites, restored reaches at BACI sites, restored reaches at SFT sites) and used Kruskal–Wallis tests and post-hoc Wilcoxon tests to compare median importance values by habit, duration, nativity, and wetland indicator status among reach types. P values from the Kruskal–Wallis and post-hoc Wilcoxon tests were adjusted for multiple comparisons. We also compared the composition of indicator species identified for unrestored and restored reaches. All statistical analyses were conducted in R version 4.2.3 (R Core Team 2023).
Results
The clearest consequence of stream restoration was an approximately 80% reduction in average woody basal area and a 75% reduction in woody stem density (Fig. 4a,b, Table S3), as the vast majority of the woody vegetation at these sites was removed during the sediment excavation process. This reduction in woody vegetation was apparent immediately following restoration (BACI sites) and persisted at sites that were 1 – 3 years post-restoration (SFT sites). This decrease in vegetation was accompanied by an increase in woody layer FQI values and a decrease in woody diversity immediately after restoration (BACI sites). This pattern persisted in sites with a longer time since restoration, but the difference in values between restored and unrestored reaches was no longer significant (Fig. 4c,d, Table S3). At the site level, overall woody richness decreased with restoration at BACI and SFT sites, primarily due to a decrease in invasive species richness (Table S3). There was no immediate effect of restoration on the woody vegetation layer wetland indicator score, but sites with a longer time since restoration had more hydrophytic vegetation (Fig. 4e, Table S3).

Response of woody layer (a) basal area (cm2m−2) (b) stem density (stems m−2) (c) plot-level floristic quality index, (d) Shannon diversity, and (e) wetland indicator score to stream restoration. Higher FQI and Shannon diversity values indicate higher-quality vegetation. Lower WIS values indicate a predominance of hydrophytic vegetation. Colors are as in Fig. 3. The grey line indicates that the Project: Pre-restoration reach in year one becomes the Project: Post-restoration reach in year 2. Means, standard errors, and results of statistical tests are presented in Table S1
The herbaceous vegetation community showed an immediate response to restoration in the form of a greater prevalence of hydrophytic vegetation (decrease in WIS), and this effect was still detectable several years post-restoration (Fig. 5a, Table S4). The change in herbaceous layer WIS score was not related to changes in woody layer basal area due to restoration (r = 0.49, p > 0.5). Regrowth of the herbaceous vegetation layer was also rapid, with no significant change in vegetation cover with restoration when compared to control reaches at BACI sites, and higher vegetation cover in restored reaches at SFT sites (Fig. 5b, Table S4). Although total cover was unchanged after restoration at BACI sites, native cover increased with restoration, invasive cover decreased, and exotic cover remained the same after restoration. The increase in herbaceous cover at the SFT sites was driven by a substantial increase in native cover while exotic cover was unchanged and invasive cover decreased (Fig. 5c-e, Table S4).

Initially (BACI sites) herbaceous species richness increased with restoration, with both native and exotic richness increasing, but invasive richness unchanged. In sites with a longer time since restoration (SFT sites), total richness was similar between restored and unrestored reaches, with restored reaches having higher native and exotic richness but lower invasive species richness (Fig. 6, Table S4). Changes in floristic quality and diversity followed similar patterns, with values for both increasing with restoration at the BACI sites but showing no significant effect of restoration at the SFT sites (Fig. 7a,b, Table S4). The proportion of perennial species in the flora remained high immediately after restoration (BACI reaches) but decreased with restoration at the SFT sites (Fig. 7c, Table S4). Beta diversity at both local (within site) and regional (among sites) scales did not change with restoration at BACI or SFT sites (Table S4).


Response of herbaceous layer (a) plot-level floristic quality index, (b) Shannon diversity, (c) proportion of perennial species, and (d) local beta diversity (within reach dissimilarity) to stream restoration. Colors are as in Fig. 3. Means, standard errors, and results of statistical tests are presented in Table S2
The average plot-level herbaceous layer FQI of unrestored reaches exhibited a significant negative correlation with the change in FQI after restoration; high-quality sites showed the largest decreases in quality with restoration (Fig. 8). A similar but non-significant trend was seen for FQI in the woody layer. There was no relationship between initial values and the change in those values with restoration for richness or Shannon diversity in either the herbaceous or woody layers. There was no relationship between project length or watershed ISC and the change in FQI, richness, or diversity in either the herbaceous or woody layer.

Relationship between herbaceous layer floristic quality index values in the unrestored/pre-restoration reach and change in these values with restoration. Higher FQI values indicate higher-quality vegetation. Values presented here are calculated at the plot level (1 m2) and are lower than those typically reported for entire habitats. The P value is calculated with a Bonferroni correction for multiple (4) comparisons
PerMANOVA of the effect of restoration on species composition showed significant differences in the herbaceous layer vegetation (df = 4, F = 2.57, p = 0.001), specifically between the unrestored and restored reaches in the SFT sites, but not between any of the reach types at the BACI sites. NMDS of the herbaceous layer vegetation resulted in a 2-dimensional solution with a final stress of 0.10 (Fig. 9a). A biplot with environmental variables shows control and unrestored reaches spread across the top and left side of the graph and suggests these sites are associated with higher richness, FQI, and basal area and less hydrophytic vegetation (higher WIS). These reaches are largely separated from the restored reaches, which are clustered at the bottom right. Along axis 1, the reaches stay grouped by site even after restoration, with North Stirrup Run, which had very little tree canopy and basal area, on the right side of the axis and Plumtree Run, which had the highest basal area and ISC on the left side of the axis. The woody layer NMDS had a 2-dimensional solution with a stress of 0.13 (Fig. 9b). There was no difference in woody layer species composition among any of the reach types. The NMDS of the woody layer shows no separation in woody community composition by treatment but does suggest that the woody composition for each site is similar before and after restoration as reaches within sites stay grouped along axis 1.

Non-metric multidimensional scaling of (a) herbaceous and (b) woody layer vegetation. Herbaceous layer vegetation was sampled for two years at control, unrestored, and restored reaches (paired symbols) and for one year on pre-restoration and post-restoration reaches (single symbols). Woody layer vegetation was sampled once for each reach
The median importance values (IV) of perennial herbaceous species, graminoids, native, and wetland species increased in BACI reaches compared to the unrestored reaches (Fig. 10, Table S1). This difference persisted in restored SFT reaches for graminoid IV. For all other comparisons, the median IV values seen in SFT restored reaches were similar to those in unrestored reaches and BACI restored reaches. There was no difference in median IV values by growth form, nativity, or wetland indicator score in the woody layer between unrestored and restored reaches (Table S2).

Median (± 1 SE) importance values (IV) for (a) herbaceous layer perennial species, (b) graminoid species, (c) native species, and (d) wetland species (OBL and FACW). Grey bars are median values across control-unrestored and project pre-restoration reaches at BACI sites and unrestored reaches at SFT sites. Red bars are median values in project post-restoration reaches at BACI sites and orange bars are median values in restored reaches at SFT sites
Significant herbaceous layer indicator species in unrestored reaches included the invasive Rosa multiflora, but also native species more indicative of wetlands or high to moderate-quality forest habitats (Table 2, Table S5). These included Symplocarpus foetidus (skunk cabbage), Circaea canadensis (enchanter's nightshade), Arisaema triphyllum (jack-in-the-pulpit), Parathelypteris noveboracensis (New York fern), and Polystichum acrostichoides (Christmas fern). For restored reaches, 79 herbaceous layer indicator species were identified, only 13 of which were intentionally planted during restoration (Table 2, Table S5). Of the 66 non-planted indicators, 56 are native species, 39 are wetland species (OBL or FACW) and 30 are graminoids. The only invasive indicator species of restored reaches was Arthraxon hispidus, which was found at all 6 restored reaches (project post-restoration and restored).
There were no significant woody layer indicator species of either unrestored or restored reaches; however, the five species with the largest decrease in indicator value with restoration were the invasive upland or facultative upland species Rubus phoenicolasius (Japanese wineberry), Lonicera maackii (bush honeysuckle), and Acer platanoides (Norway maple) and the native species Quercus alba (white oak) and Rubus occidentalis (black raspberry) (Table 3). The five woody layer species with the largest increase in indicator value with restoration were the upland species Cornus stolonifera and Sambucus canadensis, the facultative species Amelanchier canadensis, and the obligate wetland species Alnus serrulata, all of which are native, and the exotic facultative wetland species Salix purpurea. These five species were all planted at one to three of the six restored reaches and were not encountered at any of the unrestored reaches (Table 3).
Discussion
The disturbance due to extensive vegetation removal, earthmoving, and landscape sculpting involved in LSR/FR projects (Langland et al. 2020; Walter et al. 2013) occurs on a scale similar to that of many wetland creation projects implemented to compensate for U.S. wetland losses under Sect. 404 of the Clean Water Act (DeBerry and Perry 2012), and to other projects such as reprofiling of sandbars for flood control and navigation (Janssen et al. 2022), re-meandering of rivers in Northern Europe (Biggs et al. 1998; Lorenz et al. 2018; Pedersen et al. 2007a) and projects worldwide dealing with anthropogenic bottomland sedimentation (Allan James et al. 2020; Flitcroft et al. 2022; Vauclin et al. 2020; Wohl et al. 2021). While short-term vegetation responses to restoration may not be predictive of long-term outcomes or success at meeting design or permit criteria (Matthews et al. 2009; Pedersen et al. 2007a; Robertson et al. 2018; Zedler and Callaway 2002), this information can provide immediate feedback as to whether sites are meeting the primary goal of connecting the river to the floodplain (De Steven et al. 2015; Hartranft et al. 2011; Langland et al. 2020), are more susceptible to colonization by invasive species (Catford and Jansson 2014; Cooper et al. 2017; Planty-Tabacchi et al. 1996; Richardson et al. 2007; Tabacchi et al. 2005; Vidra et al. 2006), or are resulting in changes to site quality or to local and regional biodiversity (Beauchamp et al. 2015; Holl et al. 2022; Matthews and Spyreas 2010; Matthews et al. 2009). Characterizing the vegetation that colonizes these sites immediately and then several years after restoration provides insight into the early stages of wetland vegetation development and succession (DeBerry and Perry 2004; Mitsch and Gosselink 2015; Noon 1996; Sarneel et al. 2019; van der Valk 1981).
In the projects studied here, restoration by LSR/FR resulted in rapid changes in the composition of both the herbaceous and woody layers of restored reaches. Although we did not monitor groundwater depth or the frequency of overbank flooding, the immediate and sustained increase in the prevalence of hydrophytic herbaceous species suggests that the surveyed LSR/FR projects successfully increased vegetation access to groundwater and connected the stream to the floodplain (De Steven et al. 2015; Fraaije et al. 2019; Hammersmark et al. 2010; Lorenz et al. 2018; McMillan et al. 2014). Changes to the composition of the herbaceous vegetation community are also likely influenced by the substantial decrease in woody basal area and species richness, an expected consequence of the excavation accompanying these projects (Violin et al. 2011; Wood et al. 2022). At these sites, the combination of a higher water table, increased overbank flooding, and decreased shading provides an environment that favors hydrophytic, native, and graminoid species immediately after restoration (McKown et al. 2021). The rapid herbaceous layer vegetation response of these sites was expected as riparian vegetation often responds quickly to restoration involving extensive floodplain remodeling that increases floodplain connection (Fraaije et al. 2019; Kail et al. 2015; Lorenz et al. 2018; Pedersen et al. 2007b; Sarneel et al. 2019). The slower response of the woody layer to the altered hydrology at these sites is likely due to the time required for planted and recruited wetland trees to grow into the woody layer (> 1m height) and the gradual die-off of remaining upland trees and shrubs in what is now the active floodplain (DeBerry and Perry 2012; Robertson et al. 2018).
A common consequence of disturbance is a higher abundance of disturbance-adapted exotic and invasive plant species (MacDougall and Turkington 2005; Wise 2017), with riparian systems particularly susceptible to colonization by exotic species (Catford and Jansson 2014; Cooper et al. 2017; Planty-Tabacchi et al. 1996; Richardson et al. 2007; Tabacchi et al. 2005; Vidra et al. 2006). Many of the urban riparian forests in the Mid-Atlantic are highly invaded (Johnson et al. 2020) and degraded riparian areas are more likely to contain higher amounts of exotic propagules in the seed bank (Cockel and Gurnell 2011; O'Donnell et al. 2016). Proximity to urban areas and increased levels of available nitrate, a common condition of the sites chosen for restoration by LSR/FR (McMahon et al. 2021), can also lead to increased colonization of invasive species in restored sites. The extensive disturbance involved in LSR/FR projects has the potential to increase the susceptibility of these sites to colonization by exotic and invasive plant species (Hunter and DeBerry 2023), which could decrease the quality of these sites and lead to a loss of diversity, quality, and local or regional biotic homogenization (Holl et al. 2022; Matthews et al. 2009; Robertson et al. 2018). Data from the six sites we surveyed only partially supports this hypothesis. In the herbaceous layer, exotic species richness increased immediately with restoration and stayed elevated for 1–3 years after project completion, but invasive species richness was initially unaffected and later decreased with restoration. Herbaceous layer invasive cover decreased immediately with restoration and remained low 1–3 years after restoration. In the woody layer, exotic richness was low in all reaches and invasive richness decreased with restoration. The lack of expected invasive species colonization is likely due to the composition of the species pool in the immediate area. Across all reaches surveyed, none of the invasive species encountered were classified as obligate or facultative wetland species (Tables S1, 2), decreasing the chance they would succeed once wetland hydrology was restored (Hutchinson et al. 2020; Reinartz and Warne 1993). In addition to a lack of significant increase in invasive species, we also found that restoration was not associated with a decrease in woody or herbaceous FQI, or a decrease in the importance of native herbaceous species, herbaceous layer richness, or Shannon diversity; indicating that, on average, plant community quality and diversity do not significantly decline, at least in the first few years after project completion (Göthe et al. 2015; Kuglerová et al. 2016).
We also found little early evidence of biotic homogenization with restoration. The priority effects from revegetating restoration sites with a limited suite of species (Beauchamp et al. 2015; Kaase and Katz 2012; Lesage et al. 2018), combined with the elimination of the seed bank through sediment removal (Beas et al. 2013; DeBerry and Perry 2012) could cause areas within restored reaches to become more similar. At a larger scale, the combination of these processes could result in restored reaches across the landscape converging on a similar suite of species (Holl et al. 2022; Robertson et al. 2018). Recognizing that our results only cover a short period post-restoration, beta diversity may increase over time as generalist species with wide ecological niches decline and specialists grow to occupy narrow portions of environmental gradients (Matthews and Spyreas 2010) or, alternatively could decrease over time if non-native species or competitive native species increase in dominance (Aronson and Galatowitsch 2008; Robertson et al. 2018).
Early shifts in herbaceous layer species composition can also add to the understanding of the early stages of wetland ecosystem development (Mitsch et al. 2012). In LSR/FR projects much of the existing seed bank is presumably removed with the excavation of legacy sediment deposits, making conditions at these sites similar to the primary succession expected in wetland creation projects (DeBerry and Perry 2004). However, evidence from our study and others shows that vegetation colonization occurs rapidly (Kail et al. 2015) with seed potentially originating from several nearby sources (De Steven et al. 2015; Pedersen et al. 2007b) including legacy sediments left on the site (Merritts et al. 2011; Wegmann et al. 2012), seed inputs from local riparian or forested habitat outside the limit of construction disturbance (Battaglia et al. 2008; Ruzicka et al. 2010), or dispersal via hydrochory from upstream riparian and wetland habitat (Fryirs and Carthey 2022; Johansson et al. 1996; Nilsson et al. 2010). Instead of being dominated by quick-dispersing annual plant species as would be predicted and sometimes realized in the early stages of succession (Janssen et al. 2022; Noon 1996; van der Valk 1981), the proportion of annual species in the flora of restored reaches was similar to or lower than that seen in unrestored reaches. Our results are similar to those of other studies in created or restored wetlands (DeBerry and Perry 2004; Reinartz and Warne 1993) and colonization of in-stream gravel bars (Dostálek et al. 2022) that found early herbaceous colonizers are often perennial species. In these heavily disturbed settings, revegetation of newly created substrate may be a random process, highlighting the importance of seed dispersal into these sites from a variety of pathways (DeBerry and Perry 2012).
Contributions from existing seedbanks and nearby seed sources may also be responsible for the increase in graminoid importance and the preservation of regional beta diversity among restored sites (Leck 1989). Significant indicators of restored reaches included many native species that were not intentionally planted (Table 2), suggesting that each site has a unique seed bank or regional species pool that can increase regeneration potential and decrease regional homogenization (Goodson et al. 2001; Gurnell et al. 2006; O'Donnell et al. 2016; Vosse et al. 2008). In the NMDS of the herbaceous layer (Fig. 9a), axis 1 shows a consistent separation of reaches by site, both before and after restoration, clearly indicating that restoration alters species composition but not to the point that sites lose their unique herbaceous-layer identity after restoration.
While the results of the restoration projects we surveyed were largely positive, with rapid regrowth of herbaceous layer vegetation, an increase in hydrophytic plant species, and no decrease in herbaceous-layer richness, diversity, or average native community quality, there also are several potential drawbacks with respect to the vegetation community. While tree removal is an inescapable result of this process, loss of leaf litter from trees and from Symplocarpus foetidus, a quintessential wetland species that, at least initially, is nearly eradicated by sediment excavation, may deprive floodplains of critical carbon sources that are essential for nitrogen amelioration via denitrification (Henriksen and Kirkhusmo 2000; McCarty et al. 2006; McMillan et al. 2014; Rusanen et al. 2004). In a companion project, McMahon et al. (2021) found that restoration had no immediate effect on the flux of total dissolved nitrogen at any of these six sites, likely due to a limited supply of dissolved organic carbon and the legacy effects of nitrogen in groundwater.
We also found that low-quality sites showed the most improvement with restoration, as measured by the herbaceous layer community FQI, but high-quality sites improved the least or even decreased in quality after restoration (Fig. 8). We saw no relationships between project length or watershed ISC and site recovery in terms of herbaceous or woody layer FQI, nativity score, richness, and diversity. For these variables, selecting larger projects or areas with more or less ISC did not affect the outcome of restoration in terms of creating quality riparian and wetland habitat during the 1–3 years post-restoration time frame we investigated.
Over the short span of site recovery monitored for this study we found that although some vegetation responses like the increase in hydrophytic herbaceous layer vegetation occurred quickly at BACI sites and were sustained at the SFT sites, other changes were either temporary or limited in spatial scope. Of the vegetation responses we measured, we found that increases in total herbaceous layer richness, FQI, and diversity; woody layer FQI; and median importance values of perennial, native, and wetland species were seen at BACI sites and not at SFT sites. This suggests that either the response of these variables is not universal across sites or that these changes were reversed as vegetation continued to develop at these sites.
There were several differences between the unrestored reaches at BACI and SFT sites, including a different species composition (Fig. 9) and higher invasive herbaceous cover and more hydrophytic vegetation (lower WIS) at the SFT sites (Fig. 5), that could have affected the response of these sites to restoration. Additionally, we do not know the initial conditions of the restored sites at the SFT reaches, which is one of the drawbacks of a space-for-time design and may affect our interpretation of restoration effects at these sites. It is also possible that the difference in response of these sites is due to the difference in time since restoration. A study of 29 mitigation wetlands in Illinois also found that peaks in some measures of vegetation quality occurred early after restoration (Matthews et al. 2009). If these qualities are desired aspects of restored vegetation communities, active management or other interventions may be needed to sustain these qualities after restoration.
While we did not see a significant increase in invasive species with restoration, we did find that Arthraxon hispidus, the only invasive species identified as an indicator species of restored reaches, increased in importance in BACI and SFT restored reaches. This species is considered invasive in the Mid-Atlantic region and may require management if it continues to increase in cover and frequency as these sites age (Hunter and DeBerry 2023; Robertson et al. 2018; Swearingen and Fulton 2022), but may also decrease as canopy cover increases (White et al. 2020). Another potentially concerning species is the non-native Salix purpurea which was planted at three of the six restored reaches. While this non-native species is not considered invasive, priority effects from planting are expected to dominate restored woody communities (Beauchamp et al. 2015; Kaase and Katz 2012). If propagule dispersal of woody species is inhibited at these sites, as it can be at wetland creation and restoration projects (Battaglia et al. 2008), negative effects of introducing this species to these sites may manifest in the future (Cremer 2003).
As state and local governments in the Chesapeake Bay watershed increasingly turn to stream restoration to meet regulatory stormwater management and TMDL goals (Inamdar et al. 2023; Williams et al. 2017), local groups have started to object to the loss of trees and forest habitat that accompanies these projects (Condon 2023; Olivo 2020; Wheeler 2020). Data to settle the debate surrounding these tradeoffs in water quality improvement versus forest canopy preservation is sparse (Bernhardt et al. 2005; Kaushal et al. 2023; Lammers and Bledsoe 2017), and a better understanding of the ecological costs and benefits of LSR/FR and other disturbance-intensive restoration practices such as natural channel design and regenerative stormwater conveyance is urgently needed. These stream restoration methods can be effective in reducing surface water nitrogen, phosphorus, and sediment concentrations at some sites, but variability in initial site conditions, hydrology, and landscape setting of these projects can affect their ability to act as sinks for nutrients and sediments (Kaushal et al. 2023, 2008b; Lammers and Bledsoe 2017; Mayer et al. 2022; Newcomer Johnson et al. 2016; Williams et al. 2017). In some areas, legacy sediment terraces are habitat for native forest understory plant species. Our study showed that while these upland or ecotonal communities are being replaced with what is, at least initially, good quality riparian habitat, not all sites improve with restoration. Preservation of high-quality forest areas, even if they are atop legacy sediment terraces, should be considered, particularly if losses in tree canopy are not being offset by gains in nutrient cycling.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Allan James L, Beach TP, Richter DD (2020) Floodplain and terrace legacy sediment as a widespread record of anthropogenic geomorphic change. Ann Am Assoc Geogr 111:742–755. https://doi.org/10.1080/24694452.2020.1835460
Altland D, Becraft C, Berg J, Brown T, Burch J, Clearwater D, Coleman J, Crawford S, Doll B, Geratz J, Hanson J, Hartranft J, Hottenstein J, Kaushal S, Lowe S, Mayer P, Noe G, Oberholzer W, Parola A, Scott D, Stack B, Sweeney J, White J (2020) Consensus Recommendations to Improve Protocols 2 and 3 for Defining Stream Restoration Pollutant Removal Credits. Chesapeake Stormwater Network
Aronson MFJ, Galatowitsch S (2008) Long-term vegetation development of restored prairie pothole wetlands. Wetlands 28:883–895. https://doi.org/10.1672/08-142.1
Battaglia LL, Pritchett DW, Minchin PR (2008) Evaluating Dispersal Limitation in Passive Bottomland Forest Restoration. Restor Ecol 16:417–424. https://doi.org/10.1111/j.1526-100X.2007.00319.x
Beas BJ, Smith LM, Hickman KR, LaGrange TG, Stutheit R (2013) Seed bank responses to wetland restoration: Do restored wetlands resemble reference conditions following sediment removal? Aquat Bot 108:7–15. https://doi.org/10.1016/j.aquabot.2013.02.002
Beauchamp VB, Swan CM, Szlavecz K, Hu J (2015) Riparian community structure and soil properties of restored urban streams. Ecohydrology 8:880–895. https://doi.org/10.1002/eco.1644
Beechie TJ, Sear DA, Olden JD, Pess GR, Buffington JM, Moir H, Roni P, Pollock MM (2010) Process-based Principles for Restoring River Ecosystems. Bioscience 60:209–222. https://doi.org/10.1525/bio.2010.60.3.7
Bernhardt ES, Palmer MA, Allan JD, Alexander G, Barnas K, Brooks S, Carr J, Clayton S, Dahm C, Follstad-Shah J, Galat D, Gloss S, Goodwin P, Hart D, Hassett B, Jenkinson R, Katz S, Kondolf GM, Lake PS, Lave R, Meyer JL, O’Donnell TK, Pagano L, Powell B, Sudduth E (2005) Ecology. Synthesizing U.S. river restoration efforts. Science 308:636–637. https://doi.org/10.1126/science.1109769
Biggs J, Corfield A, Grøn P, Hansen HO, Walker D, Whitfield M, Williams P (1998) Restoration of the rivers Brede, Cole and Skerne: a joint Danish and British EU-LIFE demonstration project, V—short-term impacts on the conservation value of aquatic macroinvertebrate and macrophyte assemblages. Aquat Conserv: Mar Freshwat Ecosyst 8:241–255. https://doi.org/10.1002/(SICI)1099-0755(199801/02)8:1%3c241::AID-AQC269%3e3.0.CO;2-9
Booth EG, Loheide SP (2012) Hydroecological model predictions indicate wetter and more diverse soil water regimes and vegetation types following floodplain restoration. Journal of Geophysical Research: Biogeosciences 117:n/a-n/a. https://doi.org/10.1029/2011jg001831
Catford JA, Jansson R (2014) Drowned, buried and carried away: effects of plant traits on the distribution of native and alien species in riparian ecosystems. New Phytol 204:19–36. https://doi.org/10.1111/nph.12951
Chamberlain SJ, Ingram HM (2012) Developing coefficients of conservatism to advance floristic quality assessment in the Mid-Atlantic region. J Torrey Bot Soc 139(416–427):412. https://doi.org/10.3159/TORREY-D-12-00007.1
Chao A, Jost L, Chiang SC, Jiang YH, Chazdon RL (2008) A two-stage probabilistic approach to multiple-community similarity indices. Biometrics 64:1178–1186. https://doi.org/10.1111/j.1541-0420.2008.01010.x
Cockel CP, Gurnell AM (2011) An investigation of the composition of the urban riparian soil propagule bank along the River Brent, Greater London, UK, in comparison with previous propagule bank studies in rural areas. Urban Ecosyst 15:367–387. https://doi.org/10.1007/s11252-011-0203-6
Condon C (2023) Environmental groups concerned by upcoming construction along Herring Run in Northeast Baltimore. The Baltimore Sun, October 13, 2023. Baltimore, MD
Cooper DJ, Kaczynski KM, Sueltenfuss J, Gaucherand S, Hazen C (2017) Mountain wetland restoration: The role of hydrologic regime and plant introductions after 15 years in the Colorado Rocky Mountains, U.S.A. Ecol Eng 101:46–59. https://doi.org/10.1016/j.ecoleng.2017.01.017
Cremer KW (2003) Introduced willows can become invasive pests in Australia. Biodiversity 4:17–24. https://doi.org/10.1080/14888386.2003.9712705
De Steven D, Faulkner SP, Keeland BD, Baldwin MJ, McCoy JW, Hughes SC (2015) Understory vegetation as an indicator for floodplain forest restoration in the Mississippi River Alluvial Valley, U.S.A. Restor Ecol 23:402–412. https://doi.org/10.1111/rec.12210
DeBerry DA, Perry JE (2004) Primary Succession in a Created Freshwater Wetland. Castanea 69:185–193. https://doi.org/10.2179/0008-7475(2004)069%3c0185:Psiacf%3e2.0.Co;2
DeBerry DA, Perry JE (2012) Vegetation dynamics across a chronosequence of created wetland sites in Virginia, USA. Wetlands Ecol Manage 20:521–537. https://doi.org/10.1007/s11273-012-9273-3
Dostálek J, Frantík T, Pavlů L (2022) Passive restoration of vegetation on gravel/sand bars in the city: a case study in Prague, Czech Republic. Urban Ecosyst 25:1265–1277. https://doi.org/10.1007/s11252-022-01225-8
Filoso S, Palmer MA (2011) Assessing stream restoration effectiveness at reducing nitrogen export to downstream waters. Ecol Appl 21:1989–2006. https://doi.org/10.1890/10-0854.1
Flitcroft RL, Brignon WR, Staab B, Bellmore JR, Burnett J, Burns P, Cluer B, Giannico G, Helstab JM, Jennings J, Mayes C, Mazzacano C, Mork L, Meyer K, Munyon J, Penaluna BE, Powers P, Scott DN, Wondzell SM (2022) Rehabilitating Valley Floors to a Stage 0 Condition: A Synthesis of Opening Outcomes. Frontiers in Environmental Science 10. https://doi.org/10.3389/fenvs.2022.892268
Forshay KJ, Weitzman JN, Wilhelm JF, Hartranft J, Merritts DJ, Rahnis MA, Walter RC, Mayer PM (2022) Unearthing a stream-wetland floodplain system: increased denitrification and nitrate retention at a legacy sediment removal restoration site, Big Spring Run, PA, USA. Biogeochemistry 161:171–191. https://doi.org/10.1007/s10533-022-00975-z
Fraaije RGA, Poupin C, Verhoeven JTA, Soons MB, Yang Z (2019) Functional responses of aquatic and riparian vegetation to hydrogeomorphic restoration of channelized lowland streams and their valleys. J Appl Ecol 56:1007–1018. https://doi.org/10.1111/1365-2664.13326
Freyman WA, Masters LA, Packard S, Poisot T (2015) The Universal Floristic Quality Assessment (FQA) Calculator: an online tool for ecological assessment and monitoring. Methods Ecol Evol 7:380–383. https://doi.org/10.1111/2041-210x.12491
Fryirs K, Carthey A (2022) How long do seeds float? The potential role of hydrochory in passive revegetation management. River Res Appl 38:1139–1153. https://doi.org/10.1002/rra.3989
Gold AJ, Groffman PM, Addy K, Kellogg DQ, Stolt M, Rosenblatt AE (2001) Landscape attributes as controls on ground water nitrate removal capacity of riparian zones. J Am Water Resour Assoc 37:1457–1464. https://doi.org/10.1111/j.1752-1688.2001.tb03652.x
Goodson JM, Gurnell AM, Angold PG, Morrissey IP (2001) Riparian seed banks: structure, process and implications for riparian management. Prog Phys Geogr 25:301–325. https://doi.org/10.1177/0309133301025003
Göthe E, Timmermann A, Januschke K, Baattrup-Pedersen A (2015) Structural and functional responses of floodplain vegetation to stream ecosystem restoration. Hydrobiologia 769:79–92. https://doi.org/10.1007/s10750-015-2401-3
Groffman PM, Bain DJ, Band LE, Belt KT, Brush GS, Grove JM, Pouyat RV, Yesilonis IC, Zipperer WC (2003) Down by the riverside: urban riparian ecology. Front Ecol Environ 1:315–321. https://doi.org/10.1890/1540-9295(2003)001[0315:Dbtrur]2.0.Co;2
Groffman PM, Boulware NJ, Zipperer WC, Pouyat RV, Band LE, Colosimo MF (2002) Soil nitrogen cycling processes in urban riparian zones. Environ Sci Technol 36:4547–4552. https://doi.org/10.1021/es020649z
Gurnell AM, Boitsidis AJ, Thompson K, Clifford NJ (2006) Seed bank, seed dispersal and vegetation cover: Colonization along a newly-created river channel. J Veg Sci 17:665–674. https://doi.org/10.1111/j.1654-1103.2006.tb02490.x
Hammersmark CT, Dobrowski SZ, Rains MC, Mount JF (2010) Simulated Effects of Stream Restoration on the Distribution of Wet-Meadow Vegetation. Restor Ecol 18:882–893. https://doi.org/10.1111/j.1526-100X.2009.00519.x
Hartranft J, Merritts D, Walter R, Rahnis M (2011) The Big Spring Run Restoration Experiment: Policy, Geomorphology, and Aquatic Ecosystems in the Big Spring Run Watershed, Lancaster County, PA. Sustain Spring/Summer 24–30
Henriksen A, Kirkhusmo LA (2000) Effects of clear-cutting of forest on the chemistry of a shallow groundwater aquifer in southern Norway. Hydrol Earth Syst Sci 4:323–331. https://doi.org/10.5194/hess-4-323-2000
Holl KD, Luong JC, Brancalion PHS (2022) Overcoming biotic homogenization in ecological restoration. Trends Ecol Evol 37:777–788. https://doi.org/10.1016/j.tree.2022.05.002
Hunter DM, DeBerry DA (2023) Environmental drivers of plant invasion in wetland mitigation. Wetlands 43:81. https://doi.org/10.21203/rs.3.rs-2046029/v1
Hupp CR, Noe GB, Schenk ER, Benthem AJ (2013) Recent and historic sediment dynamics along Difficult Run, a suburban Virginia Piedmont stream. Geomorphology 180–181:156–169. https://doi.org/10.1016/j.geomorph.2012.10.007
Hutchinson RA, Fremier AK, Viers JH (2020) Interaction of restored hydrological connectivity and herbicide suppresses dominance of a floodplain invasive species. Restor Ecol 28:1551–1560. https://doi.org/10.1111/rec.13240
Inamdar S, Sienkiewicz N, Lutgen A, Jiang G, Kan J (2020) Streambank Legacy Sediments in Surface Waters: Phosphorus Sources or Sinks? Soil Systems 4:30. https://doi.org/10.3390/soilsystems4020030
Inamdar SP, Kaushal SS, Tetrick RB, Trout L, Rowland R, Genito D, Bais H (2023) More Than Dirt: Soil Health Needs to Be Emphasized in Stream and Floodplain Restorations. Soil Systems 7:36. https://doi.org/10.3390/soilsystems7020036
James LA, Beach TP, Richter DD (2020) Floodplain and terrace legacy sediment as a widespread record of anthropogenic geomorphic change. Ann Am Assoc Geogr 111:742–755. https://doi.org/10.1080/24694452.2020.1835460
Janssen P, Chevalier R, Chantereau M, Dupré R, Evette A, Hémeray D, Mårell A, Martin H, Rodrigues S, Villar M, Greulich S (2022) Can vegetation clearing operations and reprofiling of bars be considered as an ecological restoration measure? Lessons from a 10‐year vegetation monitoring program (Loire River, France). Restor Ecol 31:e13704. https://doi.org/10.1111/rec.13704
Johansson ME, Nilsson C, Nilsson E (1996) Do rivers function as corridors for plant dispersal? J Veg Sci 7:593–598. https://doi.org/10.2307/3236309
Johnson L, Trammell T, Bishop T, Barth J, Drzyzga S, Jantz C (2020) Squeezed from all sides: Urbanization, Invasive species, and Climate change threaten riparian forest buffers. Sustainability 12:1–23. https://doi.org/10.3390/su12041448
Kaase CT, Katz GL (2012) Effects of stream restoration on woody riparian vegetation of southern Appalachian Mountain streams, North Carolina, U.S.A. Restor Ecol 20:647–655. https://doi.org/10.1111/j.1526-100X.2011.00807.x
Kail J, Brabec K, Poppe M, Januschke K (2015) The effect of river restoration on fish, macroinvertebrates and aquatic macrophytes: A meta-analysis. Ecol Indicators 58:311–321. https://doi.org/10.1016/j.ecolind.2015.06.011
Kaushal SS, Groffman PM, Mayer PM, Striz E, Gold AJ (2008a) Effects of stream restoration on denitrification in an urbanizing watershed. Ecol Appl 18:789–804
Kaushal SS, Groffman PM, Mayer PM, Striz E, Gold AJ (2008b) Effects of stream restoration on denitrification in an urbanizing watershed. Ecol Appl 18:789–804. https://doi.org/10.1890/07-1159.1
Kaushal SS, Fork ML, Hawley RJ, Hopkins KG, Ríos-Touma B, Roy AH (2023) Stream restoration milestones: monitoring scales determine successes and failures. Urban Ecosyst 26:1131–1142. https://doi.org/10.1007/s11252-023-01370-8
Kuglerová L, Botková K, Jansson R (2016) Responses of riparian plants to habitat changes following restoration of channelized streams. Ecohydrology 10:e1798. https://doi.org/10.1002/eco.1798
Lammers RW, Bledsoe BP (2017) What role does stream restoration play in nutrient management? Crit Rev Environ Sci Technol 47:335–371. https://doi.org/10.1080/10643389.2017.1318618
Langland MJ, Duris JW, Zimmerman TM, Chaplin JJ (2020) Effects of legacy sediment removal and effects on nutrients and sediment in Big Spring Run, Lancaster County, Pennsylvania, 2009–15. U.S. Geological Survey Scientific Investigations Report 2020-5031, pp 28. https://doi.org/10.3133/sir20205031
Larson DM, Riens J, Myerchin S, Papon S, Knutson MG, Vacek SC, Winikoff SG, Phillips ML, Giudice JH (2019) Sediment excavation as a wetland restoration technique had early effects on the developing vegetation community. Wetlands Ecol Manage 28:1–18. https://doi.org/10.1007/s11273-019-09690-3
Leck MA (1989) Wetland Seed Banks. p. 283–305. In M. A. Leck, V. T. Parker and R. L. Simpson (eds.), Ecology of Soil Seed Banks. Academic Press. https://doi.org/10.1016/b978-0-12-440405-2.50018-x
Lesage JC, Howard EA, Holl KD (2018) Homogenizing biodiversity in restoration: the “perennialization” of California prairies. Restor Ecol 26:1061–1065. https://doi.org/10.1111/rec.12887
Lorenz AW, Haase P, Januschke K, Sundermann A, Hering D (2018) Revisiting restored river reaches - Assessing change of aquatic and riparian communities after five years. Sci Total Environ 613–614:1185–1195. https://doi.org/10.1016/j.scitotenv.2017.09.188
MacDougall AS, Turkington R (2005) Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 86:42–55. https://doi.org/10.1890/04-0669
Magurran AE (2004) Measuring Biological Diversity. Blackwell Publishing, Malden, MA
Matthews JW, Spyreas G (2010) Convergence and divergence in plant community trajectories as a framework for monitoring wetland restoration progress. J Appl Ecol 47:1128–1136. https://doi.org/10.1111/j.1365-2664.2010.01862.x
Matthews JW, Spyreas G, Endress AG (2009) Trajectories of vegetation-based indicators used to assess wetland restoration progress. Ecol Appl 19:2093–2107. https://doi.org/10.1890/08-1371.1
Mayer PM, Pennino MJ, Newcomer-Johnson TA, Kaushal SS (2022) Long-term assessment of floodplain reconnection as a stream restoration approach for managing nitrogen in ground and surface waters. Urban Ecosyst 25:879–907. https://doi.org/10.1007/s11252-021-01199-z
McCarty GW, Mookherji S, Angier JT (2006) Characterization of denitrification activity in zones of groundwater exfiltration within a riparian wetland ecosystem. Biol Fertil Soils 43:691–698. https://doi.org/10.1007/s00374-006-0151-0
McKown JG, Moore GE, Payne AR, White NA, Gibson JL (2021) Successional dynamics of a 35 year old freshwater mitigation wetland in southeastern New Hampshire. Plos One 16:e0251748. https://doi.org/10.1371/journal.pone.0251748
McLane CR, Battaglia LL, Gibson DJ, Groninger JW (2012) Succession of Exotic and Native Species Assemblages within Restored Floodplain Forests: A Test of the Parallel Dynamics Hypothesis. Restor Ecol 20:202–210. https://doi.org/10.1111/j.1526-100X.2010.00763.x
McMahon P, Beauchamp VB, Casey RE, Salice CJ, Bucher K, Marsh M, Moore J (2021) Effects of stream restoration by legacy sediment removal and floodplain reconnection on water quality. Environ Res Lett 16:035009. https://doi.org/10.1088/1748-9326/abe007
McMillan SK, Tuttle AK, Jennings GD, Gardner A (2014) Influence of Restoration Age and Riparian Vegetation on Reach-Scale Nutrient Retention in Restored Urban Streams. JAWRA J Am Water Resour Assoc 50:626–638. https://doi.org/10.1111/jawr.12205
Meisenbach WJ, Tychsen H, Siu C, Baker KH (2012) Failure of Reach-Scale Restoration to Improve Biotic Integrity in a Mid-Atlantic Stream. Environ Pollut 1:124–131. https://doi.org/10.5539/ep.v1n2p124
Merritts D, Walter R, Rahnis M, Hartranft J, Cox S, Gellis A, Potter N, Hilgartner W, Langland M, Manion L, Lippincott C, Siddiqui S, Rehman Z, Scheid C, Kratz L, Shilling A, Jenschke M, Datin K, Cranmer E, Reed A, Matuszewski D, Voli M, Ohlson E, Neugebauer A, Ahamed A, Neal C, Winter A, Becker S (2011) Anthropocene streams and base-level controls from historic dams in the unglaciated mid-Atlantic region, USA. Philos Trans A Math Phys Eng Sci 369:976–1009. https://doi.org/10.1098/rsta.2010.0335
Mitsch WJ, Zhang L, Stefanik KC, Nahlik AM, Anderson CJ, Bernal B, Hernandez M, Song K (2012) Creating Wetlands: Primary Succession, Water Quality Changes, and Self-Design over 15 Years. Bioscience 62:237–250. https://doi.org/10.1525/bio.2012.62.3.5
Mitsch WJ, Gosselink JG (2015) Wetlands Fifth edition. John Wiley and Sons, Inc.
Newcomer Johnson T, Kaushal S, Mayer P, Smith R, Sivirichi G (2016) Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis. Water 8:116. https://doi.org/10.3390/w8040116
Nilsson C, Brown RL, Jansson R, Merritt DM (2010) The role of hydrochory in structuring riparian and wetland vegetation. Biol Rev Camb Philos Soc 85:837–858. https://doi.org/10.1111/j.1469-185X.2010.00129.x
Noon KF (1996) A model of created wetland primary succession. Landscape Urban Plann 34:97–123. https://doi.org/10.1016/0169-2046(95)00209-x
O’Donnell J, Fryirs KA, Leishman MR (2016) Seed banks as a source of vegetation regeneration to support the recovery of degraded rivers: A comparison of river reaches of varying condition. Sci Total Environ 542:591–602. https://doi.org/10.1016/j.scitotenv.2015.10.118
Olivo A (2020) Stream restorations for Chesapeake Bay fuels debate in Fairfax. The Washington Post, January 25 2020.
Palmer MA, Filoso S, Fanelli RM (2014a) From ecosystems to ecosystem services: Stream restoration as ecological engineering. Ecol Eng 65:62–70. https://doi.org/10.1016/j.ecoleng.2013.07.059
Palmer MA, Hondula KL, Koch BJ (2014b) Ecological restoration of streams and rivers: Shifting strategies and shifting goals. Annu Rev Ecol, Evol, Syst 45:247–269. https://doi.org/10.1146/annurev-ecolsys-120213-091935
Pedersen ML, Andersen JM, Nielsen K, Linnemann M (2007a) Restoration of Skjern River and its valley: Project description and general ecological changes in the project area. Ecol Eng 30:131–144. https://doi.org/10.1016/j.ecoleng.2006.06.009
Pedersen ML, Friberg N, Skriver J, Baattrup-Pedersen A, Larsen SE (2007b) Restoration of Skjern River and its valley—Short-term effects on river habitats, macrophytes and macroinvertebrates. Ecol Eng 30:145–156. https://doi.org/10.1016/j.ecoleng.2006.08.009
Planty-Tabacchi AM, Tabacchi E, Naiman RJ, Deferrari C, Decamps H (1996) Invasibility of species rich communities in riparian zones. Conserv Biol 10:598–607. https://doi.org/10.1046/j.1523-1739.1996.10020598.x
Qian H, Guo Q (2010) Linking biotic homogenization to habitat type, invasiveness and growth form of naturalized alien plants in North America. Divers Distrib 16:119–125. https://doi.org/10.1111/j.1472-4642.2009.00627.x
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Reinartz JA, Warne EL (1993) Development of vegetation in small created wetlands in southeastern Wisconsin. Wetlands 13:153–164. https://doi.org/10.1007/bf03160876
Richardson DM, Holmes PM, Esler KJ, Galatowitsch SM, Stromberg JC, Kirkman SP, Pyšek P, Hobbs RJ (2007) Riparian vegetation: degradation, alien plant invasions, and restoration prospects. Divers Distrib 13:126–139. https://doi.org/10.1111/j.1366-9516.2006.00314.x
Robertson M, Galatowitsch SM, Matthews JW (2018) Longitudinal evaluation of vegetation richness and cover at wetland compensation sites: implications for regulatory monitoring under the Clean Water Act. Wetlands Ecol Manage 26:1089–1105. https://doi.org/10.1007/s11273-018-9633-8
Rusanen K, Finer L, Antikainen M, Korkka-Niemi K, Backman B, Britschgi R (2004) The effect of forest cutting on the quality of groundwater in large aquifers in Finland. Boreal Environ Res 9:253–261
Ruzicka KJ, Groninger JW, Zaczek JJ (2010) Deer browsing, forest edge effects, and vegetation dynamics following bottomland forest restoration. Restor Ecol 18:702–710. https://doi.org/10.1111/j.1526-100X.2008.00503.x
Sarneel JM, Hefting MM, Kowalchuk GA, Nilsson C, Van der Velden M, Visser EJW, Voesenek L, Jansson R (2019) Alternative transient states and slow plant community responses after changed flooding regimes. Glob Chang Biol 25:1358–1367. https://doi.org/10.1111/gcb.14569
Schenk ER, Hupp CR (2009) Legacy effects of colonial millponds on floodplain sedimentation, bank erosion, and channel morphology, mid-atlantic, USA. JAWRA J Am Water Resour Assoc 45:597–606. https://doi.org/10.1111/j.1752-1688.2009.00308.x
Swearingen JM, Fulton JP (2022) plant invaders of mid-atlantic natural areas, field guide, 4th edn. Passiflora Press
Tabacchi E, Planty-Tabacchi A-M, Roques L, Nadal E (2005) Seed inputs in riparian zones: implications for plant invasion. River Res Appl 21:299–313. https://doi.org/10.1002/rra.848
USDA, NRCS (United States Department of Agriculture, National Resources Conservation Service) (2020) The PLANTS Database http://plants.usda.gov. National Plant Data Team, Greensboro, North Carolina, USA
van der Valk AG (1981) Succession in wetlands: A gleasonian appraoch. Ecology 62:688–696. https://doi.org/10.2307/1937737
Vauclin S, Mourier B, Piégay H, Winiarski T (2020) Legacy sediments in a European context: The example of infrastructure-induced sediments on the Rhône River. Anthropocene 31:100248. https://doi.org/10.1016/j.ancene.2020.100248
Vidra RL, Shear TH, Wentworth TR (2006) Testing the paradigms of exotic species invasion in urban riparian forests. Nat Areas J 26:339–350. https://doi.org/10.3375/0885-8608(2006)26[339:TTPOES]2.0.CO;2
Violin CR, Cada P, Sudduth EB, Hassett BA, Penrose DL, Bernhardt ES (2011) Effects of urbanization and urban stream restoration on the physical and biological structure of stream ecosystems. Ecol Appl 21:1932–1949. https://doi.org/10.1890/10-1551.1
Voli M, Merritts D, Walter R, Ohlson E, Datin K, Rahnis M, Kratz L, Deng W, Hilgartner W, Hartranft J (2009) Preliminary reconstruction of a pre-European settlement valley bottom wetland, southeastern Pennsylvania. Water Resour Impact 11:11–13
Vosse S, Esler KJ, Richardson DM, Holmes PM (2008) Can riparian seed banks initiate restoration after alien plant invasion? Evidence from the Western Cape, South Africa. S Afr J Bot 74:432–444. https://doi.org/10.1016/j.sajb.2008.01.170
Walsh CJ, Roy AH, Feminella JW, Cottingham PD, Groffman PM, Morgan Ii RP (2005) The urban stream syndrome: Current knowledge and the search for a cure. J N Am Benthol Soc 24:706–723. https://doi.org/10.1899/04-028.1
Walter R, Merritts D, Rahnis M (2007) Estimating volume, nutrient content, and rates of stream bank erosion of legacy sediment in the Piedmont and Valley and Ridge physiographic provinces, southeastern and central PA. p. 40. A Report to the Pennsylvania Department of Environmental Protection. https://files.dep.state.pa.us/water/chesapeake%20bay%20program/lib/chesapeake/pdfs/padeplegacysedimentreport2007waltermerrittsrahnisfinal.pdf
Walter R, Merritts D, Rahnis M, Langland M, Galeone D, Gellis A, Hilgartner W, Bowne D, Wallace J, Mayer P, Forshay K (2013) Big Spring Run Natural Floodplain, Stream, and Riparian Wetland - Aquatic Resource Restoration Project Monitoring. Pennsylvania Department of Environmental Protection Final Report, 109. http://www.bsrproject.org/uploads/2/6/5/2/26524868/big_spring_run_aquatic_ecosystem_restoration_monitoring_report_2013.pdf
Walter RC, Merritts DJ (2008) Natural streams and the legacy of water-powered mills. Science 319:299–304. https://doi.org/10.1126/science.1151716
Weakley AS, Ludwig JC, Townsend JF, Crowder B (2012) Flora of Virginia, 1st, ed. Botanical Research Institute of Texas Press, Fort Worth, Tex
Wegmann KW, Lewis RQ, Hunt MC (2012) Historic mill ponds and piedmont stream water quality: Making the connection near Raleigh, North Carolina. p. 93–121. From the Blue Ridge to the Coastal Plain: Field Excursions in the Southeastern United States. Geological Society of America. https://doi.org/10.1130/2012.0029(03)
Weitzman JN, Kaye JP (2017) Nitrate retention capacity of milldam-impacted legacy sediments and relict A horizon soils. Soil 3:95–112. https://doi.org/10.5194/soil-3-95-2017
Weitzman JN, Forshay KJ, Kaye JP, Mayer PM, Koval JC, Walter RC (2014) Potential nitrogen and carbon processing in a landscape rich in milldam legacy sediments. Biogeochemistry 120:337–357. https://doi.org/10.1007/s10533-014-0003-1
Wentworth TR, Johnson GP, Kologiski RL (1988) Designation of wetlands by weighted averages of vegetation data: A preliminary evaluation. J Am Water Resour Assoc 24:389–396. https://doi.org/10.1111/j.1752-1688.1988.tb02997.x
Wheeler TB (2020) Stream restoration techniques draw pushback. Chesapeake Bay Journal, October 7, 2020
White L, Catterall C, Taffs K (2020) The habitat and management of hairy jointgrass (Arthraxon hispidus, Poaceae) on the north coast of New South Wales, Australia. Pac Conserv Biol 26:45–56. https://doi.org/10.1071/pc19017
Williams MR, Bhatt G, Filoso S, Yactayo G (2017) Stream restoration performance and its contribution to the chesapeake Bay TMDL: Challenges Posed by climate change in urban areas. Estuaries Coasts 40:1227–1246. https://doi.org/10.1007/s12237-017-0226-1
Wise M (2017) A field investigation into the effects of anthropogenic disturbances on biodiversity and alien invasions of plant communities. Bioscene: Journal of College Biology Teaching 43:3–14. https://eric.ed.gov/?id=EJ1170736
Wohl E, Castro J, Cluer B, Merritts D, Powers P, Staab B, Thorne C (2021) Rediscovering, Reevaluating, and Restoring Lost River-Wetland Corridors. Frontiers in Earth Science 9:653623. https://doi.org/10.3389/feart.2021.653623
Wood KL, Kaushal SS, Vidon PG, Mayer PM, Galella JG (2022) Tree Trade-Offs in Stream Restoration: Impacts on Riparian Groundwater Quality. Urban Ecosyst 25:773–795. https://doi.org/10.1007/s11252-021-01182-8
Zedler JB, Callaway JC (2002) Tracking wetland restoration: Do mitigation sites follow desired trajectories? Restor Ecol 7:69–73. https://doi.org/10.1046/j.1526-100X.1999.07108.x
Acknowledgements
We thank Ginny Jeppi, David Cote and Lisa Wheeler for field assistance and Charlie Davis for help with plant identification. Scott and Colin McGill at Ecotone Inc. were invaluable partners, and we thank Howard County, the City of Bel Air and the Edwards, Rigdon and Pitts families for access to their properties. We also appreciate the constructive feedback from two anonymous reviewers.
Funding
This project was supported with a Restoration Research grant (award #13974) from the Chesapeake Bay Trust and funding from Towson University.
Author information
Authors and Affiliations
Contributions
Vanessa Beauchamp and Joel Moore conceived and designed the research; Vanessa Beauchamp and Patrick Baltzer performed the field sampling and analyzed the data; Christopher Salice assisted with data analysis; Vanessa Beauchamp, Patrick Baltzer, and Joel Moore wrote and edited the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baltzer, P.J., Moore, J., Salice, C.J. et al. The Effects of Legacy Sediment Removal and Floodplain Reconnection on Riparian Plant Communities. Wetlands 44, 15 (2024). https://doi.org/10.1007/s13157-023-01768-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-023-01768-2